
[Mitosis y meiosis] [Microorganismos] [Plantas] [Invertebrados] [Peces] [Anfibios y reptiles] [Aves] [Humanos] [Enfermedades humanas]
Índice
Índice
||1|| Introducción ||2|| Generalidades ||3|| Las algas rodófitas y glaucófitas ||4|| Las algas verdes y los líquenes ||5|| Las briofitas ||6|| Helechos ||7|| Gimnospermas ||8|| Introducción a las Angiospermas | ||11|| Gametogénesis en las angiospermas ||12|| Polinización ||13|| Fertilización ||14|| Desarrollo embrionario en angiospermas ||15|| Las frutas ||16|| Las semillas ||17|| Dispersión de las semillas |
Portada

Portada
1. Introducción
1. Introducción
Para ponerme un poco pretencioso, esta unidad está dedicada a las plantas en sensu lato, es decir, vamos a trabajar a las plantas verdaderas “Viridiplantae” y sus parientes que poseen cloroplastos de endosimbiosis simple conocidos como arquiplastidos (Arana, Correa, & Oggero, 2014; Gololobova, 2015; Mackiewicz & Gagat, 2014). Sin embargo, el mayor peso recaerá en las plantas más exitosas que existen en la actualidad, las plantas con flor o angiospermas, las cuales demuestran una gran especialización en sus estructuras reproductivas.
Las plantas en sensu lato y en sensu estricto pueden reproducirse sexual y asexualmente, aunque depende mucho del grupo con el cual se esté trabajando. En esta serie de artículos se empleará la definición del reino plantas como Viridiplantae cuando hablemos de las plantas verdaderas o plantas en sensu estricto, es decir, todos los grupos de plantas terrestres más las algas verdes con cloroplastos de primera endosimbiosis, pero excluyendo a los glaucófitos y las algas rojas.

Figura 1.1. Dendrograma de los arquiplástidos. Los arquiplástidos se dividen en dos clados principales, los glaucófitos, que poseen cloroplastos con peptidoglicano; y el segundo clado que a su vez se divide en los rodófitos y en las plantas verdes verdaderas.
En términos de reproducción sexual la variabilidad es alta, de hecho, los mecanismos por medio de los cuales una planta se reproduce son el núcleo fundamental de las principales categorías taxonómicas de las plantas. Un aspecto latente en la reproducción y la evolución de las plantas, es que entre más reciente es el mecanismo reproductivo, más independiente es de la presencia de agua (De Duve & Pizano, 1995). Esto implica que los mecanismos reproductivos más antiguos son al mismo tiempo los que requieren mucho más de la presencia de agua. De hecho, la estructura de resistencia de las plantas, llamada semilla, evolucionó en la era geológica más seca de todos los tiempos, el pérmico (Linkies, Graeber, Knight, & Leubner‐Metzger, 2010; E. L. Taylor, Taylor, & Krings, 2009).
Carlos Lineo (1707-1778)
Fue un destacado botánico y zoólogo sueco, conocido como el padre de la taxonomía moderna. Sus contribuciones revolucionaron el campo de la biología al desarrollar un sistema de clasificación científica basado en la estructura y características de los organismos vivos. Su obra más famosa, "Systema Naturae", sentó las bases para la nomenclatura binomial que se utiliza hasta hoy en día. Linneo es considerado uno de los científicos más influyentes de la historia.

Contexto social
Carlos Linneo vivió en una época marcada por importantes cambios políticos, económicos, sociales, religiosos y culturales. A mediados del siglo XVIII, Europa estaba experimentando la Ilustración, un movimiento intelectual que promovía la razón, la ciencia y el pensamiento crítico.
En el ámbito político, Linneo vivió en una Suecia que se encontraba en proceso de transición hacia una monarquía absoluta. El país estaba tratando de consolidar su posición como potencia europea y expandir su influencia política y económica.
En términos económicos, el comercio y la navegación desempeñaban un papel central. El creciente comercio internacional y la explotación de colonias en América y otras partes del mundo brindaron nuevas oportunidades y riqueza a las naciones europeas, incluida Suecia.
En el ámbito social, la sociedad estaba estratificada en clases, con la nobleza y la burguesía ejerciendo gran poder. A pesar de esto, la Ilustración trajo consigo una creciente preocupación por la igualdad y los derechos individuales.
En cuanto a lo religioso, el protestantismo luterano era la religión dominante en Suecia y en gran parte de Europa. Sin embargo, la Ilustración también trajo consigo un espíritu de crítica hacia las instituciones religiosas y una mayor aceptación de ideas científicas que desafiaban las creencias tradicionales.
Culturalmente, la Ilustración estimuló un renacimiento intelectual y artístico. Se fomentaba la educación, el estudio de las ciencias y las artes, y se promovía la difusión de conocimientos a través de la imprenta y los salones literarios.
En este contexto, Carlos Linneo pudo desarrollar su trabajo científico y revolucionar la taxonomía. Sus ideas innovadoras y su enfoque científico riguroso se beneficiaron de la efervescencia intelectual de la época, así como del apoyo de mecenas y colegas interesados en los avances científicos.
Infancia
Carlos Linneo nació el 23 de mayo de 1707 en Råshult, Småland, Suecia. Provenía de una familia modesta y su infancia estuvo marcada por el amor hacia la naturaleza y la curiosidad por el mundo natural. Desde temprana edad, mostró un gran interés por las plantas y los animales, pasando largas horas explorando los campos y bosques cercanos a su hogar.
A pesar de las limitaciones económicas de su familia, Linneo tuvo la oportunidad de recibir una educación gracias al apoyo de su padre y de algunos profesores locales. Demostró una habilidad excepcional para el estudio y la memorización, lo que le permitió destacarse en sus estudios desde una edad temprana.
Durante su infancia, Linneo comenzó a recolectar y clasificar plantas, creando su propio herbario personal. Estas experiencias despertaron su pasión por la botánica y sentaron las bases para su futura carrera científica.
Aunque la educación formal en su época estaba dominada por el aprendizaje religioso y clásico, Linneo encontró en la naturaleza su principal fuente de inspiración. Explorar su entorno natural se convirtió en una constante fuente de descubrimiento y aprendizaje para él, lo que sentó las bases de su posterior desarrollo como botánico y zoólogo de renombre.
La infancia de Carlos Linneo estuvo marcada por su innato interés por la naturaleza, su capacidad de observación y su incansable curiosidad, cualidades que sentaron las bases para su futura contribución a la ciencia y la taxonomía.
Logros
Carlos Linneo recibió una educación académica en diversas instituciones. Estudió en la Universidad de Lund y posteriormente en la Universidad de Uppsala, ambas en Suecia. Durante su formación, se destacó por su habilidad para el estudio y la memorización, así como por su pasión por la botánica y la taxonomía.
Entre los logros científicos más destacados de Linneo se encuentran sus contribuciones a la taxonomía. Desarrolló el sistema de clasificación binomial, que consiste en asignar a cada especie un nombre científico compuesto por dos partes: el género y la especie. Esto permitió una organización sistemática y unificada de la diversidad biológica, sentando las bases para la taxonomía moderna.
Uno de sus principales instrumentos científicos fue el herbario, una colección de plantas secas y prensadas, que utilizó para describir y clasificar nuevas especies. Además, diseñó un sistema de símbolos para representar las características de las plantas, conocido como el "sistema sexual de Linneo", que permitía identificar la estructura reproductiva de las plantas.
Entre sus publicaciones más destacadas se encuentra "Systema Naturae", una obra en la que presentó su sistema de clasificación y nomenclatura. Esta obra tuvo múltiples ediciones y fue ampliada a lo largo de su vida. También escribió otras obras como "Species Plantarum" y "Genera Plantarum", que ampliaron y perfeccionaron su sistema de clasificación.
Si bien Linneo realizó grandes avances en la taxonomía, también cometió algunos errores, principalmente debido a las limitaciones de su época en términos de acceso a información y tecnología. Por ejemplo, algunos de sus conceptos de especies eran más amplios o estrechos de lo que se sabe actualmente. Sin embargo, sus contribuciones generales fueron fundamentales y sentaron las bases para futuros avances en la clasificación y el estudio de la diversidad biológica.
En cuanto a controversias, Linneo tuvo diferencias con otros científicos de la época, como Georges-Louis Leclerc Conde de Buffon, con quien discutió sobre la clasificación de algunas especies. Además, su sistema de clasificación binomial y su énfasis en la estructura reproductiva de las plantas también generaron controversias y críticas en su tiempo.
Carlos Linneo enfrentó controversias en relación con su concepto de sexualidad en las plantas, especialmente con otros científicos y con miembros del clero. En su época, el tema de la sexualidad en las plantas era un tema delicado debido a las creencias religiosas y a las concepciones tradicionales sobre la reproducción en el reino vegetal.
Linneo sostenía que las plantas tenían órganos sexuales y que la reproducción en ellas involucraba la unión de órganos masculinos y femeninos. Su sistema sexual de clasificación de las plantas, basado en la estructura de sus órganos reproductivos, generó controversias y oposición por parte de algunos miembros del clero y científicos conservadores.
En particular, Linneo tuvo desacuerdos con el naturalista francés Georges-Louis Leclerc Conde de Buffon, con quien debatió acerca de la clasificación y la interpretación de las flores y sus órganos sexuales. Leclerc sostenía una visión más conservadora, argumentando que las flores no tenían órganos sexuales claramente definidos.
Además, Linneo también enfrentó críticas de la Iglesia, ya que su concepción de la sexualidad en las plantas iba en contra de la interpretación religiosa tradicional. Algunos religiosos veían en sus teorías una amenaza a la concepción de la creación divina.
A pesar de estas controversias, Linneo defendió su posición y presentó sus argumentos en diversas publicaciones. Una de las obras más relevantes en las que abordó el tema de la sexualidad en las plantas fue "Philosophia Botanica" (1751), donde desarrolló su sistema de clasificación sexual y argumentó a favor de la presencia de órganos sexuales en las plantas.
Otra obra importante fue "Corollarium Generum Plantarum" (1737), en la que abordó el tema de la clasificación de las plantas y la importancia de la estructura de los órganos reproductivos.
Estas publicaciones y los debates que generaron son un testimonio de la controversia que rodeó las ideas de Linneo sobre la sexualidad en las plantas en su tiempo. Sin embargo, con el tiempo, la visión de Linneo fue ampliamente aceptada y sus contribuciones sentaron las bases para el estudio y la comprensión de la reproducción vegetal.
Comunidad científica
Carlos Linneo fue parte de una vibrante comunidad científica en su época, compuesta por destacados científicos, académicos y naturalistas. A través de su trabajo y sus contribuciones científicas, estableció relaciones con varios colaboradores, amigos, opositores y, en menor medida, enemigos.
Uno de los principales colaboradores y amigos de Linneo fue Daniel Solander, un destacado botánico sueco. Solander se convirtió en estrecho colaborador de Linneo y trabajó con él en la catalogación de plantas en el Jardín Botánico de Uppsala. Juntos, realizaron expediciones botánicas y clasificaron muchas especies de plantas. Solander fue uno de los primeros en adoptar y difundir el sistema de clasificación binomial de Linneo.
Otros colaboradores destacados de Linneo incluyen a Pehr Kalm, un botánico y explorador sueco, quien recopiló y envió valiosas muestras de plantas de América del Norte a Linneo. También colaboró con Anton Rolandsson Martin, un naturalista y médico sueco que participó en expediciones científicas en Suecia y África.
En términos de opositores, uno de los más notables fue Georges-Louis Leclerc, conocido como el Conde de Buffon, un influyente naturalista francés. Leclerc y Linneo tenían diferencias en relación con la clasificación de las especies y la interpretación de la sexualidad en las plantas. Tuvieron debates y discusiones acaloradas sobre estos temas, aunque también compartieron cierto respeto mutuo por sus contribuciones científicas.
Linneo también enfrentó críticas y oposición de algunos miembros del clero y religiosos conservadores debido a sus teorías sobre la sexualidad en las plantas, que desafiaban las creencias religiosas tradicionales sobre la creación divina.
Si bien no se conocen enemigos acérrimos de Linneo, sus ideas y su sistema de clasificación fueron objeto de controversias y críticas en su tiempo. Sin embargo, es importante destacar que Linneo también contó con un amplio apoyo y reconocimiento de otros científicos y académicos de renombre en Europa, quienes valoraron y adoptaron su sistema de clasificación y sus aportes a la botánica y la taxonomía.
En resumen, Carlos Linneo se involucró en una comunidad científica en constante interacción, donde tuvo colaboradores cercanos, como Solander, Kalm y Martin, así como opositores notables, como el Conde de Buffon. Aunque enfrentó críticas y controversias, su trabajo fue ampliamente reconocido y estableció la base para el desarrollo de la botánica y la taxonomía moderna.
Reconocimientos
Carlos Linneo recibió varios reconocimientos tanto en vida como póstumos debido a sus notables contribuciones científicas. Estos reconocimientos reflejaron la importancia y el impacto duradero de su trabajo en la comunidad científica y más allá.
En vida, Linneo fue honrado con diversos títulos y distinciones. En 1741, fue nombrado miembro de la Real Academia de Ciencias de Suecia y, más tarde, se convirtió en su presidente. Además, en 1753, fue nombrado caballero por el rey sueco Adolfo Federico, otorgándole el título de "Caballero Linneo".
Linneo también fue reconocido internacionalmente. En 1761, fue elegido miembro extranjero de la Real Academia de Ciencias de Francia y, en 1768, fue admitido en la Sociedad Real de Londres. Estos reconocimientos destacaron su influencia y prestigio en la comunidad científica más allá de las fronteras de Suecia.
Después de su fallecimiento en 1778, los reconocimientos póstumos a Linneo continuaron. En 1784, se erigió un monumento en su honor en Uppsala, Suecia, donde se encuentra la Universidad de Uppsala, en reconocimiento a su legado científico. Además, su hogar en Råshult se convirtió en un museo dedicado a su vida y trabajo.
El impacto duradero de Linneo en la ciencia también se evidencia en la cantidad de especies y géneros botánicos que llevan su nombre. Muchas plantas fueron bautizadas en su honor, y los términos "linnaeano" y "linnaeidae" se utilizan ampliamente en la taxonomía para hacer referencia a las clasificaciones y grupos establecidos por él.
En el campo de la botánica, la medalla Linneana, instituida en 1888, se otorga cada año en la Sociedad Linneana de Londres a científicos que han hecho contribuciones sobresalientes en esta disciplina.
El reconocimiento y la admiración hacia Linneo también se han manifestado a través de eventos espontáneos y homenajes por parte de la comunidad científica. En varias ocasiones, los botánicos y naturalistas se han reunido para celebrar el aniversario de su nacimiento y conmemorar su legado. Estos eventos demuestran la apreciación continua por su trabajo y su influencia en el campo de la taxonomía y la biología.
En resumen, Carlos Linneo recibió reconocimientos en vida, como títulos de nobleza y membresías en academias científicas, y fue homenajeado con monumentos y museos en su honor después de su fallecimiento. Su legado perdura a través de los nombres de plantas que llevan su nombre, así como de la medalla Linneana y los eventos conmemorativos realizados por la comunidad científica en reconocimiento a su destacada contribución a la botánica y la taxonomía.
¿Por qué él es importante?
El trabajo de Carlos Linneo ha tenido un impacto significativo en nuestra vida cotidiana, especialmente en los ámbitos de la biología, la botánica, la medicina y la agricultura. Sus contribuciones en la clasificación y nomenclatura de especies han proporcionado un marco de comprensión y comunicación universalmente aceptado sobre la diversidad de los seres vivos. Aquí hay algunas formas en las que el trabajo de Linneo nos afecta en nuestra vida diaria:
(a) Identificación y clasificación de especies: El sistema de clasificación binomial desarrollado por Linneo, que asigna un nombre científico único a cada especie, nos permite identificar y distinguir entre diferentes organismos. Esto es fundamental en campos como la ecología, la conservación, la agricultura y la medicina, ya que nos ayuda a comprender las interacciones y características de las diferentes especies.
(b) Agricultura y jardinería: La clasificación de las plantas por parte de Linneo ha sido invaluable para la agricultura y la jardinería. Nos permite identificar y seleccionar las plantas adecuadas para diversos propósitos, como la producción de alimentos, la ornamentación o la obtención de productos medicinales. Además, su sistema de nomenclatura nos permite comunicarnos de manera precisa sobre las especies de plantas que cultivamos.
(c) Medicina y farmacología: Linneo sentó las bases para la clasificación de plantas medicinales y su utilización en la medicina tradicional y la farmacología. Su sistema de clasificación nos ayuda a identificar las plantas con propiedades medicinales específicas y a comprender su uso histórico y potencial terapéutico. Muchos medicamentos modernos se derivan de plantas clasificadas por Linneo.
(d) Conservación y biodiversidad: La clasificación de las especies según Linneo ha sido esencial para comprender y conservar la diversidad biológica. Nos permite evaluar el estado de las poblaciones, identificar especies en peligro de extinción y diseñar estrategias de conservación efectivas.
(e) Investigación científica: El sistema de clasificación de Linneo proporciona un marco común para la investigación científica en biología y botánica. Permite a los científicos comunicarse y compartir información de manera precisa y eficiente, lo que impulsa el avance de la ciencia y facilita la colaboración internacional.
En resumen, el trabajo de Carlos Linneo nos afecta en nuestra vida cotidiana al brindarnos una comprensión estructurada y universalmente aceptada de la diversidad biológica. Su sistema de clasificación nos ayuda a identificar y comunicarnos sobre las especies, lo que tiene implicaciones en la agricultura, la medicina, la conservación y la investigación científica.
¿Qué debemos aprender de Lineo?
La vida de Carlos Linneo nos brinda importantes lecciones e inspiración para enfrentar los desafíos y buscar el éxito en nuestras propias vidas. Aquí hay algunas formas en las que podemos inspirarnos en su vida, limitaciones, obstáculos, logros e impacto:
Linneo demostró una pasión ardiente por la botánica y la ciencia en general. Nos inspira a perseguir nuestros propios intereses y cultivar una sed de conocimiento en el campo que nos apasione. La dedicación y el amor por lo que hacemos nos permiten superar obstáculos y alcanzar logros significativos.
A pesar de las limitaciones y desafíos que enfrentó en su infancia y juventud, Linneo no se rindió. Su determinación y voluntad de superar obstáculos nos enseñan la importancia de enfrentar las dificultades con valentía y encontrar soluciones creativas.
Linneo dedicó largas horas de trabajo y estudio para desarrollar su sistema de clasificación y realizar sus investigaciones botánicas. Su dedicación nos recuerda que el éxito no llega fácilmente y requiere esfuerzo constante y disciplina.
Linneo fue un innovador en su campo, desarrollando un sistema de clasificación revolucionario que aún se utiliza en la actualidad. Su capacidad para ver más allá de las ideas convencionales y desafiar las normas establecidas nos inspira a buscar nuevas perspectivas y enfoques en nuestras propias vidas y carreras.
El legado de Linneo continúa influyendo en la ciencia y en nuestras vidas hasta el día de hoy. Su trabajo ha sido fundamental en la clasificación y comprensión de la naturaleza, y ha tenido un impacto significativo en campos como la biología, la agricultura y la medicina. Esto nos anima a pensar en cómo podemos dejar un impacto duradero en nuestro propio entorno y contribuir al bienestar de la sociedad.
En resumen, la vida de Carlos Linneo nos enseña la importancia de la pasión, la superación de limitaciones, la persistencia, la innovación y el impacto duradero. Inspirándonos en su ejemplo, podemos buscar nuestras propias pasiones, enfrentar los obstáculos con determinación y trabajar arduamente para lograr nuestros objetivos, dejando un legado positivo en nuestro campo y en la sociedad en general.
2. Generalidades
2. Generalidades
La reproducción en plantas es un fenómeno fascinante que nos sumerge en un mundo lleno de diversidad y complejidad. Desde la sexualidad en las flores hasta la alternancia de generaciones entre el gametofito y el esporofito, este proceso vital es fundamental para la supervivencia y el éxito evolutivo de las plantas. Además, la formación de esporas y semillas desempeña un papel crucial en su dispersión y colonización de nuevos entornos.
Este artículo tiene como objetivo proporcionar una introducción completa a la reproducción en plantas, explorando los conceptos clave y desentrañando los mecanismos subyacentes. Comenzaremos adentrándonos en la sexualidad vegetal, donde descubriremos cómo las plantas se reproducen a través de la fertilización de gametos y cómo la diversidad de estrategias sexuales desempeña un papel crucial en su adaptación a diferentes entornos.
A continuación, nos sumergiremos en la alternancia de generaciones, un proceso único en las plantas que involucra la alternancia entre dos fases distintas: el gametofito y el esporofito. Exploraremos cómo estas dos generaciones se entrelazan en un ciclo vital complejo y cómo cada una cumple funciones específicas en la reproducción y la supervivencia de la planta.
Además, examinaremos el papel fundamental de las esporas y las semillas en la reproducción vegetal. Descubriremos cómo las esporas, estructuras reproductivas unicelulares, se forman y se dispersan para dar origen a nuevos individuos. También exploraremos el increíble poder de las semillas, estructuras adaptadas para la dispersión, la protección y la germinación, que han permitido a las plantas colonizar una amplia variedad de hábitats en todo el mundo.
A lo largo de este artículo, nos adentraremos en la complejidad y la belleza de la reproducción en plantas, revelando los mecanismos subyacentes que permiten a estos seres vivos prosperar y adaptarse en diversos entornos. Al comprender estos procesos fundamentales, podremos apreciar aún más la increíble diversidad y vitalidad del reino vegetal y su importancia para la vida en nuestro planeta.
Reproducción sexual y asexual
La reproducción en las algas verdes sigue un esquema sexual y asexual. La reproducción sexual puede ser de dos tipos, vegetativa cuando la colonia es fragmentada y enviada a otro lugar, o mediante la producción de zoosporas en las especies que son sedentarias. La reproducción sexual es caracterizada por una predominancia de la generación haploide sobre la diploide.

Figura 2.1. Macho y hembra. La distinción entre macho y hembra generalmente radica en el tamaño de uno de los gametos, el más grande generalmente es asociado a la hembra y el más pequeño al macho, casos que concuerdan con esto son la anisogamia y la ovogamia; pero en algunos protistas, existe isogamia, los gametos son iguales y por ende no existe criterio de macho o hembra.
De hecho, en la mayoría de las especies, la única célula de la generación diploide es el cigoto que también funciona como la estructura de resistencia, lo cual nuevamente confirma el hecho de que la reproducción sexual evolucionó como un mecanismo de emergencia ante un ambiente hostil. Las células vegetativas de las algas son haploides, lo cual hace que no tengan acceso a la compensación de dosis por la presencia de los cromosomas homólogos. En cualquier caso, las algas verdes son un grupo muy antiguo y diverso, por lo que a continuación solo se describirán unos pocos grupos (Bowman, Sakakibara, Furumizu, & Dierschke, 2016; Cock & Coelho, 2011; Qiu, Taylor, & McMANUS, 2012).
Alternancia de generaciones
Aunque ya discutimos el concepto en la introducción general en la unidad de reproducción en unicelulares y hongos, vale la pena repasarlo, ya que para las plantas la alternancia de generaciones es un concepto fundamental, ya que después de los hongos, estas son el mejor ejemplo del ciclo de vida más completo de los eucariotas (Coelho et al., 2007; L. K. E. Graham & Wilcox, 2000; Hughes & Otto, 1999; KENRICK, 1994; Mable & Otto, 1998; Niklas & Kutschera, 2010).
El ciclo de vida de alternancia de generaciones de las plantas se caracteriza por la ausencia de esporas asexuales altamente especializadas tanto en el cuerpo somático generador de esporas “esporofito” o en el cuerpo somático generador de gametos “gametofito”. Sin embargo, ambos cuerpos pueden regenerarse mitóticamente (10, 11) y en consecuencia aún pueden ejecutar la reproducción asexual por fragmentación dependiendo del grupo del cual estemos hablando.
El ciclo de vida de alternancia de generaciones tal como se muestra en la Figura 2.2 iniciará con la generación esporofítica, la cual puede o inducir todas sus células en meiosis, o especializar alguna parte de sus tejidos para la meiosis (2), esto dependerá de si estamos hablando de especias unicelulares o multicelulares. Asumiendo que trabajamos con multicelulares, el esporofito generalmente genera estructuras de generaciones de espora altamente desarrolladas.

Figura 2.2. Modelo de la alternancia de generaciones. Las plantas verdaderas, especialmente las más antiguas, se caracterizan por un ciclo de vida con dos cuerpos y dos tipos de propágulos, dependiendo del grupo, una de las generaciones se hace más grande, mientras que la más pequeña crece dentro de la más grande, hasta que se hace casi invisible.
Una vez que se ha efectuado la meiosis y las esporas han madurado, estas deben ser liberadas al ambiente (3). Estas esporas son sexuadas, ya sea de manera dioica en la cual cada espora dará lugar a un macho o a una hembra, o monoicas en la cual el individuo sexuado es hermafrodita. En algunas especies la sexualizacion puede ocurrir en el mismo esporofito, produciéndose solo esporas masculinas y solo esporas femeninas, aunque este nivel de especialización se observará principalmente en las plantas terrestres adaptadas a la vida en ambientas más secos como las gimnospermas y las angiospermas.
Una vez que las esporas sexuales llegan a un ambiente húmedo estas regeneran (4) un cuerpo gametofítico haploide (5). Una de las características de la evolución de las plantas, es que el gametofito pasa de ser una generación bastante conspicua y de casi el mismo tamaño que el esporofito en los musgos, a ser una estructura invisible, encerrada en el propio esporofito, por ejemplo el gametofito en las angiospermas y gimnospermas de la hembra siempre está encerrado en la recámara del ovario, mientras que el gametofito masculino se posa en los receptáculos y germina de manera microscópica al interior de los estróbilos o las flores. En otras palabras, la tendencia de las plantas terrestres es que la generación esporofítica diploide se haga más importante. Esto tiene algún sentido biológico ya que, al hacerse más grande, debe hacer frente a más parásitos y es necesaria mayor diversidad genética, y eso lo da células diploides.
El gametofito (5) por lo tanto se hace menos capaz de regenerarse en las plantas más recientes, pero aun así debe germinar y crecer de modo tal que algunas de sus células puedan especializarse para formar los gametos masculinos y femeninos (6). Los gametos experimentan la fecundación plasmogámica y cariogámica (7), dando lugar rápidamente a la primera célula de la generación diploide, el cigoto (8). Este cigoto puede desarrollarse directamente aprovechando las condiciones ambientales favorables, o permanecer encerrado al interior de todo el órgano reproductivo femenino en forma de una semilla. Por lo general las semillas almacenan embriones, no cigotos, de diversos estados de desarrollo, pero lo importante es que, a diferencia de las esporas, las semillas son mucho más resistentes a factores mecánicos, físicos, químicos y biológicos. Por lo general solo los linajes más recientes están protegidos con semillas, las demás simplemente regeneran (9) al esporofito (1).
Gametofito
Cuerpo somático que debe enfrentar a los factores ambientales y que en suma conocemos como selección natural, cuyas células poseen solo una copia de cada cromosoma, por lo que los denominamos haploides. El nombre de gametofito implica que es una planta que genera los gametos sexuales masculinos “espermas” y femeninos “óvulos”.
Como nota adicional en las plantas con semilla el gametofito masculino es el tubo de polen generado cuando el polen germina al interior de la flor o del estróbilo.
Dado lo anterior, si queremos ver gametofitos impresionantes, deberemos buscar en los clados de plantas sin semillas, especialmente, los helechos.
(a) Cyathea brownii es una especie de helecho arbóreo que se encuentra en Nueva Zelanda. Su gametofito, conocido como "peloteado", puede alcanzar tamaños extraordinarios, con un diámetro de hasta 1 metro. Esto lo convierte en uno de los gametofitos más grandes conocidos en el reino vegetal.

Figura 2.3. Cyathea brownii es una especie de helecho arbóreo endémica de Nueva Zelanda. También conocido como helecho gigante de tronco desnudo, puede alcanzar alturas impresionantes de hasta 20 metros. Sus frondas son grandes y plumosas, y su tronco es grueso y cubierto de escamas marrones. Cyathea brownii es apreciado por su presencia majestuosa en los bosques húmedos de Nueva Zelanda y su gametofito, conocido como "peloteado", puede alcanzar tamaños extraordinarios, con un diámetro de hasta 1 metro.
(b) Azolla filiculoides es una especie de helecho acuático flotante. Su gametofito, una pequeña estructura verde que flota en la superficie del agua, puede formar extensas colonias que cubren grandes cuerpos de agua. Estas colonias pueden ser tan grandes que se pueden ver desde una distancia considerable.
(c) Marchantia polymorpha es un tipo de hepática, una planta no vascular. Su gametofito, llamado "talo", es plano y de forma lobulada, con un tamaño que puede alcanzar varios centímetros de diámetro. Los tallos de M. polymorpha pueden formar extensas colonias en suelos húmedos y sombreados.
(d) Chara fragilis es una especie de alga verde de agua dulce. Su gametofito, conocido como "cuerda", tiene una apariencia filamentosa y puede crecer hasta alcanzar varios metros de longitud. Los gametofitos de Chara fragilis forman densas poblaciones en el fondo de lagos y ríos.
(e) Lycopodium clavatum es una especie de licopodio, un tipo de planta vascular sin semillas. Su gametofito, conocido como "protonema", es un filamento verde que crece horizontalmente sobre el suelo o en troncos de árboles. Aunque individualmente pueden ser pequeños, los protonemas de L. clavatum pueden extenderse en grandes áreas, formando una red densa y visible.
Sin embargo, los gametofitos actuales son solo una sombra de una biodiversidad más grande y esplendorosa, estas plantas proliferaron durante el carbonífero, por lo que para ver ejemplares grandes, debemos indagar en el registro fósil.

Figura 2.4. Leclercqia es un género extinto de helechos arborescentes que existió durante el período Carbonífero, hace aproximadamente 350 millones de años. Estos helechos pertenecían a la clase Polypodiopsida y mostraban características similares a los helechos modernos. Los restos fósiles de Leclercqia revelan una estructura con hojas ramificadas y frondas que se asemejan a los helechos arborescentes actuales. Este género fósil proporciona información importante sobre la evolución y diversidad de los helechos a lo largo del tiempo geológico.
El gametofito conocido más grande de todos los tiempos es el de la especie fósil de helecho arborescente, Leclercqia complexa. Este gametofito fósil, que existió durante el período Carbonífero, hace aproximadamente 350 millones de años, tenía un tamaño excepcionalmente grande en comparación con los gametofitos de las plantas actuales.
Los restos fósiles de Leclercqia complexa revelan gametofitos que alcanzaban diámetros de hasta 3 metros. Estos gametofitos fosilizados muestran una estructura similar a los helechos modernos, con hojas ramificadas que se asemejan a las frondas de los helechos arborescentes actuales.
La existencia de gametofitos tan grandes en el pasado sugiere que las condiciones ambientales y los procesos evolutivos pueden haber favorecido el desarrollo de gametofitos de mayores dimensiones en períodos antiguos. Leclercqia complexa es un ejemplo notable de cómo los gametofitos pueden variar en tamaño a lo largo de la historia evolutiva de las plantas.
Esporofito
Cuerpo somático que debe enfrentar a los factores ambientales y que en suma conocemos como selección natural, cuyas células poseen dos copias de cada cromosoma, por lo que los denominamos diploides. El nombre de esporofito significa planta generadora de esporas. Los esporofitos de los helechos no son muy impresionantes, siendo pequeños o microscópicos, pero en las plantas con semilla, la cosa es diferente. Algunos esporofitos impresionantes que podemos tener en cuenta son:
(a) Victoria amazonica (Victoria amazonica): Este lirio de agua gigante es nativo de las regiones tropicales de Sudamérica. Sus hojas flotantes son enormes y pueden alcanzar un diámetro de hasta 3 metros. Son de color verde intenso y tienen bordes ligeramente ondulados, con una estructura reforzada por nervaduras. Las flores son grandes y fragantes, de color blanco o rosado, y se abren durante la noche.
(b) Licopodio arbóreo (Lycopodium cernuum): Este licopodio arbóreo es una especie nativa de áreas tropicales de Asia y Australia. Su esporofito se caracteriza por ser alto y ramificado, alcanzando alturas de hasta 30 metros. Sus frondas son densas y cubiertas de pequeñas hojas en forma de escamas, que le dan una apariencia frondosa y exuberante.
(c) Sagú japonés (Cycas revoluta): esta planta es nativa de Japón. Su esporofito se presenta en forma de una palma arbórea con hojas grandes y arqueadas que pueden alcanzar longitudes de hasta 1,5 metros. Las hojas son de color verde oscuro y tienen un aspecto plumoso y frondoso, creando una apariencia majestuosa.
(d) Araucaria de hoja esmeralda (Araucaria heterophylla): también conocida como pino de Norfolk, es originaria de la isla de Norfolk, en el océano Pacífico. Su esporofito es un árbol alto y majestuoso que puede alcanzar alturas de hasta 60 metros. Tiene una forma distintiva y ramas horizontales con hojas en forma de aguja de color verde oscuro, lo que le da un aspecto frondoso y denso.
(e) Zamia furfuracea (Zamia furfuracea): Esta cícada es originaria de México. Su esporofito es una planta arbustiva con hojas en forma de abanico que pueden crecer hasta 1 metro de largo. Las hojas son de color verde brillante y están cubiertas por una capa de escamas cerosas que les da un aspecto distintivo y frondoso.
Sin embargo, ningún esporofito puede compararse con el de las magníficas secoyas. Su tamaño imponente y su longevidad han capturado la admiración de las personas y las han convertido en símbolos de fortaleza y longevidad en el reino vegetal.

Figura 2.5. El género Sequoia está compuesto por árboles coníferos perennes de gran tamaño que pertenecen a la familia Cupressaceae. Estas especies son conocidas por su imponente tamaño, longevidad y presencia majestuosa en los bosques. Son considerados algunos de los árboles más grandes y antiguos del mundo. Los árboles del género Sequoia son de crecimiento lento y pueden alcanzar alturas impresionantes, superando los 100 metros en algunos casos. Sus troncos son masivos y tienen una corteza gruesa y fibrosa que les proporciona protección contra incendios y enfermedades. Las ramas suelen ser horizontales y las hojas, en forma de escamas, se disponen de manera alterna a lo largo de las ramas.
Las secoyas son árboles con semillas de gran tamaño que se destacan por su imponente esporofito. A continuación, te presento dos ejemplos de secoyas:
(a) Secoya gigante (Sequoiadendron giganteum): Esta especie es nativa de las regiones montañosas del oeste de Estados Unidos. Es conocida por ser uno de los árboles más grandes del mundo en términos de volumen. Los esporofitos de la secoya gigante pueden alcanzar alturas de hasta 95 metros y tienen troncos masivos, con diámetros que pueden superar los 8 metros. Sus hojas son pequeñas y en forma de escamas, y forman una copa densa en la parte superior del árbol.
(b) Secoya roja (Sequoia sempervirens): Esta especie es nativa de la costa oeste de Estados Unidos, desde el sur de Oregón hasta el norte de California. Los esporofitos de la secoya roja pueden alcanzar alturas de hasta 115 metros, lo que las convierte en algunas de las especies de árboles más altas del mundo. Tienen troncos masivos con diámetros que pueden superar los 6 metros. Sus hojas son pequeñas, en forma de escamas y de color verde brillante.
Las secoyas son ejemplos impresionantes de árboles con esporofitos grandes y majestuosos. Su tamaño imponente y su longevidad han capturado la admiración de las personas y las han convertido en símbolos de fortaleza y longevidad en el reino vegetal.
Esporas
Las esporas son estructuras reproductivas esenciales para el ciclo de vida de las plantas y desempeñan un papel fundamental en su reproducción. Estas estructuras, también conocidas como propágulos reproductivos, pertenecen a la generación esporofítica y se distinguen de las semillas por sus características y funciones específicas. A través del proceso de meiosis, se generan las esporas, que constituyen una fase intermedia en el ciclo de vida de las plantas y darán lugar al desarrollo del gametofito, el cual sí produce las células reproductivas.
Un aspecto interesante sobre las esporas es su sensibilidad a la falta de agua. Debido a esta sensibilidad, no son muy resistentes y, por lo tanto, las plantas producen un gran número de esporas para aumentar las posibilidades de éxito reproductivo, pero únicamente en estaciones muy lluviosas o ambientes que sean húmedos permanentemente. Esta estrategia se ha desarrollado a lo largo de la evolución de las plantas, y se observa una tendencia en la que, inicialmente, tanto los géneros masculinos como los femeninos liberaban esporas. Sin embargo, a medida que evolucionaron para invadir terrenos más secos, la espora femenina quedó retenida en la planta esporofítica, mientras que la espora masculina se volvió más abundante y móvil, recibiendo el nombre de polen no-germinado.
Un ejemplo de una planta con esporas interesantes es el helecho arborescente (Cyathea brownii). Este helecho de gran tamaño produce esporas en soros, que son agrupaciones de estructuras esporangiales en la parte inferior de las frondas. Estas esporas tienen la capacidad de dispersarse a largas distancias, lo que permite la colonización de nuevos hábitats.

Figura 2.6. Lycopodium cernuum, conocido como licopodio arqueado o licopodio colgante, es una planta rastrera con tallos delgados y flexibles. Sus hojas pequeñas y puntiagudas crecen de forma alterna a lo largo del tallo. Se encuentra en regiones templadas y tropicales. Es apreciado por su forma arqueada y su aspecto delicado.
Otro ejemplo intrigante es el helecho Licopodio arbóreo (Lycopodium cernuum). Este helecho arbóreo produce esporas en conos llamados estrobilos, que se encuentran en la parte superior de la planta. Las esporas del Licopodio arbóreo son extremadamente ligeras y pueden ser dispersadas por el viento a distancias sorprendentes.
A medida que las plantas evolucionaron y se adaptaron a nuevos entornos, se produjeron cambios significativos en las características y funciones de las esporas. Estos cambios estuvieron influenciados por factores como la disponibilidad de agua, la colonización de tierra firme y la necesidad de sobrevivir en el inhóspito desierto del Pérmico en el supercontinente Pangea. En general aunque las esporas se adaptaron a ambientes con menor dispobilidad de agua, no pudieron desprenderse de esta, por lo que las plantas productoras de esporas como los helechos se vieron confinados a ambientes costeros altamente húmedos. Sin embargo cierto grupo de plantas logró adaptar otro tipo de propágalo llamado, semilla.
Semilla
Las semillas son estructuras altamente complejas que desempeñan un papel fundamental en el ciclo de vida de las plantas. Están compuestas por un embrión en desarrollo y diversos órganos protectores que trabajan en conjunto para salvaguardar al embrión de los efectos adversos del ambiente. Una de las características más destacadas de las semillas es su resistencia a la pérdida de agua. Además de contar con una cubierta dura que evita la deshidratación del embrión, las semillas poseen reservas de alimento que brindan nutrición al embrión durante su germinación y desarrollo inicial.
Las primeras semillas evolucionaron hace aproximadamente 360 millones de años, durante el período Devónico. Fue en esta época que las plantas desarrollaron la capacidad de producir semillas, lo que representó un hito importante en la historia evolutiva de las plantas terrestres. Las semillas proporcionaron a las plantas una ventaja selectiva significativa al permitirles resistir períodos de sequía, dispersarse a nuevos territorios y proteger a los embriones en desarrollo.

Figura 2.7. El superdesierto de Pangea, durante el Pérmico, influyó en la evolución de las semillas. Las condiciones extremadamente secas llevaron a la proliferación de especies con semillas resistentes a la desecación. Estas estructuras protegieron los embriones y permitieron a las plantas sobrevivir en entornos áridos. Además, las semillas desarrollaron mecanismos de dispersión para colonizar áreas más favorables. Este evento marcó un hito en la historia de las plantas terrestres y sentó las bases para la diversidad y éxito de las plantas con semillas en la actualidad.
Esta resistencia a la pérdida de agua es de vital importancia para las plantas, especialmente en entornos secos y desafiantes como el desierto de Pangea durante el período Pérmico. En ese tiempo, el continente Pangea experimentó condiciones áridas y escasez de agua. Las plantas que habitaban ese entorno hostil tuvieron que desarrollar mecanismos de supervivencia para adaptarse a la falta de agua. Las semillas jugaron un papel crucial en esta adaptación, ya que permitieron a las plantas dispersar sus progenies y colonizar nuevas áreas, superando las limitaciones de la dependencia del agua para la reproducción.
Existen numerosos ejemplos de semillas destacables en el reino vegetal. Uno de ellos es el roble (Quercus spp), un género de árboles que produce semillas conocidas como bellotas. Estas semillas poseen una cubierta dura y una estructura interna que alberga el embrión en desarrollo y las reservas de alimento necesarias para su germinación y crecimiento inicial. Otro ejemplo interesante es el coco (Cocos nucifera), una planta tropical que produce grandes semillas en forma de nueces. Estas semillas tienen una cubierta fibrosa y resistente que protege el embrión y almacena una cantidad significativa de agua y nutrientes, lo que les permite sobrevivir largos períodos flotando en el agua y colonizar nuevas áreas costeras.
En resumen, las semillas son estructuras altamente complejas y resistentes, que permiten a las plantas sobrevivir y adaptarse en entornos desafiantes. Su resistencia a la pérdida de agua, su capacidad para almacenar nutrientes y su importancia en la evolución de las plantas son aspectos destacados. Ejemplos notables de semillas incluyen las bellotas de los robles y las nueces de coco, que demuestran la diversidad y la adaptación de las semillas en el reino vegetal.
3. Las algas rodófitas y glaucófitas
3. Las algas rodófitas y glaucófitas
En este capítulo examinaremos los ciclos de vida y modos reproductivos de algunas algas que no son clasificadas como plantas verdaderas, pero que están relacionadas taxonómicamente al ser todos poseedores de cloroplastos de primera endosimbiosis, los cuales son dos linajes, los rodófitos o algas rojas y los glaucófitos (De Clerck, Bogaert, & Leliaert, 2012; Hackett, Su Yoon, Butterfield, Sanderson, & Bhattacharya, 2007; Kim, Park, Bhattacharya, & Yoon, 2014; Ragan & Gutell, 1995).
Algas rojas
Son un grupo primitivo de pequeñas algas unicelulares esféricas que presentan una coloración similar a las algas glaucófitas que va de verde a verde azulada (o verde cian). Se caracterizan por ser extremófilas debido a su condición termoacidófila (son termófilas e hiperacidófilas), habitando en aguas termales, calderas volcánicas y algunos entornos ácidos producto de la actividad humana, mostrando además resistencia a la presencia de metales tóxicos. Su reproducción es asexual por división mitótica, formando en ocasiones endosporas en número y a veces hasta 16 células hijas. Son organismos muy simples, y en la mayoría de ocasiones poseen un solo cloroplasto y una sola mitocondria, los cuales se reproducen por división sincronizada entre la célula y estos mismos organelos. La simplicidad de estas algas estaría en relación con la pérdida evolutiva de genes. Se estima que Cyanidiophytina habría perdido veces más genes que Rhodophytina. Las algas rojas poseen dos tipos de ciclo de vida dependiendo de la especie, pero aunque ambos involucran la alternancia de generaciones, uno es más complejo que el otro, además de que las estructuras cambian (CHAPMAN, 1961; L. E. Graham, 1985; Searles, 1980).
Algas rojas digénicas
La expresión "alga roja digénica" se refiere a un tipo de alga roja que tiene un ciclo de vida que involucra dos fases distintas, conocidas como generación diploide y generación haploide. La fase diploide se caracteriza por tener células con un par de juegos completos de cromosomas, mientras que la fase haploide contiene células con un solo juego de cromosomas.
Este ciclo de vida digénico es común en las algas rojas y es una característica distintiva de este grupo de organismos. Durante la fase diploide, se produce la reproducción sexual y se forman esporas haploides. Estas esporas haploides germinan y dan lugar a la fase haploide, donde ocurre la reproducción asexual y se generan nuevas células haploides. Estas células haploides pueden fusionarse nuevamente para formar la fase diploide y completar el ciclo.
Algunos ejemplos importantes de algas rojas digénicas son:
(a) Chondrus crispus: También conocida como musgo de Irlanda, es una alga roja ampliamente distribuida en las costas del Atlántico Norte. Tiene un ciclo de vida digénico en el que la fase diploide forma estructuras laminadas y la fase haploide se presenta como pequeños talos filamentosos.
(b) Porphyra spp.: Este género de algas rojas incluye varias especies de importancia económica, como Nori (Porphyra yezoensis) y Laver (Porphyra umbilicalis). Estas algas tienen un ciclo de vida digénico en el que la fase diploide forma estructuras laminadas y la fase haploide se presenta como pequeñas láminas filamentosas.
(c) Gracilaria spp.: Este género de algas rojas incluye varias especies utilizadas en la industria de la alimentación y la biotecnología. Tienen un ciclo de vida digénico similar al de otras algas rojas, con una fase diploide que forma talos y una fase haploide que produce estructuras filamentosas.
Esporofito
El cuerpo somático dominante es el esporofito diploide, que se trata de una estructura macroscópica y pluricelular, pero con carecía de una estructura tisular altamente especializada, lo cual es análogo a muchas algas verdes verdaderas. La función del esporofito es la producción de esporas sexuales al interior de esporangios, además de poder sobrevivir hasta la edad reproductiva, siendo la estructura dominante que debe soportar la mayor parte de la selección natural. La principal ventaja de las algas oceánicas es que no deben preocuparse por la disponibilidad de agua, de forma tal que el principal propágulo reproductivo son las esporas, las cuales al estar maduras se liberan en forma de zoosporas.
Zoosporas
Estas zoosporas son células de la generación gametofítica, pero no son gametos, sino estructuras que al encontrar un sustrato adecuado proliferaran para dar lugar a los gametofitos. Estructuralmente se asemejan a zoosporas de otros protistas, poseen dos flagelos, uno con estructuras semejantes que se describen como vellos, y otro flagelo liso. Al ser zoosporas están pueden nadar activamente hasta encontrar un sustrato adecuado.
Gametofito
Los gametofitos son microscópicos, siendo de unos cuantos cientos de células. Por lo general estamos tratando con ciclos dioicos, por lo que cada espero producirá un gametofito de un sexo específico, siendo uno generador de machos y otro generador de hembras. Una vez que los gametofitos han proliferado lo suficiente deben iniciar la especialización de algunas de sus células en la espermatogénesis y la ovogénesis. La estructura reproductiva masculina se denominará anteridio y liberará espermatozoides que nadarán atraídos por feromonas a la estructura femenina denominada ovogonio.
Gametos
La fecundación ocurre en los ovogonios, donde los óvulos permanecen inmovilizados. Estas algas son por lo tanto de tipo anisogámicos, donde los gametos masculinos se caracterizan típicamente por tener menor calidad de citoplasma, ser móviles y ser producidos en mayor cantidad, además son de tipo zoospórico con un flagelo liso y otro con vellos; mientras que el gameto femenino se encuentra inmovilizado, posee mayor calidad de citoplasma y se produce en menores cantidades.
Parasitismo generacional
La fecundación da lugar rápidamente a la generación esporofítica diploide, pero el gameto no es protegido ni convertido en un propágulo, en su lugar inicia su crecimiento aprovechando nutrientes que fluyen desde el gametofito, y por tanto algunos autores denominan a este tipo de relaciones como parasitismo generacional. En este sentido la generación diploide obtiene nutrientes de la generación gametofítica en mayores cantidades hasta que la mata. Esto se da porque el esporofito crece muchos miles de veces más que el gametofito aplastándolo en cualquier caso.
Ciclo de vida
Al madurar lo suficiente, el esporofito (1) dominante genera esporangios (2) en su interior o bordes a nivel microscópico. Las células al interior de los esporangios realizan la meiosis (3) produciendo esporas sexuales dioicas (4).

Figura 3.1. Ciclo de vida de las algas dígenas.
Unas esporas regeneran (5a) al gametofito masculino que al madurar genera anteridios (6), este anteridio posee capacidades regenerativas limitadas (5b) en caso de daños mecánicos, pero formalmente sus propágulos son los espermatozoides de tipo zoospora (7). Las zoosporas también pueden regenerar gametofitos femeninos que al madurar generan el ovogonio (8), el cual produce óvulos inmóviles en sus vértices (9). La fecundación (10) ocurre sobre el ovogonio y el cigoto no se propaga (11a). En lugar de ello, el nuevo esporofito se regenera (12a) encima del viejo gametofito, alimentándose a sus expensas, gasta que crece y lo aplasta, formando el esporofito maduro (1), el cual también puede regenerar partes dañadas (11b).
Algas rojas trigénicas
Las algas rojas trigénicas poseen un ciclo de vida relativamente complejo, que involucra la formación de tres tipos de talos somáticos diferentes.
Tetrasporofito
Es el primer talo y el más dominante, perteneciendo a la generación diploide. En consecuencia, es la estructura que debe afrontar la mayor parte de la selección natural. Al microscopio el tetrasporofito es semejante al talo del gametofito. Cuando el tetrasporofito madura produce los tetrasporangios. En el interior de los tetrasporangios, las células son inducidas a realizar la meiosis, produciendo las esporas que inician la etapa haploide del ciclo de vida. Las tetrasporas del ciclo trigénico son muy diferentes, pues carecen de flagelos, por lo que dependen de las corrientes para dispersarse, pero al igual que las zoosporas sexuales del ciclo de vida anterior, se desarrollan en estructuras dioicas de sexos separados que denominamos gametofitos.
Gametofito
Los gametofitos masculino y femenino en este caso no son aplastables, ya que crecen casi a la par con el esporofito, y poseen una estructura macroscópica similar. El gametofito masculino prosigue con la espermatogénesis en estructuras especializadas denominadas espermatogonios. Los espermatogonios se asemejan a grupos de uvas muy densos, cada espermatogonio contiene un solo espermatio que funcionará como un gameto inmóvil. La estructura reproductora femenina es denominada carpoginia, y es producida en el talo del gametofito femenino. Cada carpogonio contiene una única célula que se ve como si fuera una botella microscópica con un cuello largo denominado tricogino, la función del tricógino es la de ser como una caña de pescar a la cual se pegan las espermatias cuando pasan cerca, cuando se fecunda el óvulo en el interior del carpogonio esta estructura desaparece. Un solo núcleo en la base del carpogonio funciona como un gameto único, el óvulo.
Fecundación
Debido a que los espermatio no poseen flagelos, depende exclusivamente el flujo estocástico de las corrientes para poder alcanzar el talo femenino, donde se encuentran los óvulos. Si un espermatio llega lo bastante cerca de un tricogino, las paredes de ambos se rompen, permitiendo que el núcleo haploide del espermatio migre al núcleo del óvulo, permitiendo la cariogamia y la formación de la primera célula de la generación diploide denominada cigoto.
Propágulos sexuales
Tal vez la diferencia más marcada de los ciclos digénicos y trogénicos, es el destino del gametofito y del cigoto. En el ciclo anterior vimos que el cigoto crece como un parasito de su gametofito y no se transforma en un propágulo reproductivo. En el ciclo trigeno la cosa es diferente, pues el cigoto se desarrolla en una estructura denominada quistocarpo. Cada quistocarpo resulta del desarrollo combinado del embrión, un pericarpio que es un grupo de células producido por el gametofito femenino alrededor del embrión. Cuando el embrión se desarrolla se denomina carposporangio el cual al combinarse con el pericarpio femenino da lugar al quistosporangio. En esta etapa se da una reproducción asexual obligatoria en la cual el embrión único da lugar a muchas esporas denominadas carposporas, las cuales son liberadas al medioambiente. Cuando la carpospora llega a un sustrato adecuado, germina para dar lugar al tetrasporofito.
Ciclo de vida
A pesar de todo, el ciclo de vida dígeno y trígeno es bastante semejante, las diferencias radican en gametofitos grandes, que no mueren aplastados y en que el embrión da lugar a esporas que funcionan como propágulos asexuales.

Figura 3.2. Ciclo de vida de las algas trígenas.
Iniciamos el ciclo de vida con el cuerpo somático o talo denominando tetrasporofito, que, aunque lleva un nombre más complicado, funciona como cualquier esporofito (1), generando estructuras especializadas llamadas tetrasporangios (2), donde ocurre la meiosis (3). El resultado son tetrasporas (4) que flotan en el agua. Las tetrasporas son sexuadas, por lo que algunas regeneran mitoticamente (5a) un gametofito masculino (6) o un gametofito femenino (9). Ambos gametofitos son somáticos y semejantes al esporofito. El gametofito masculino genera estructuras donde se genera la espermatogénesis llamadas espermatangios (7) que producen las espermatias (8) las cuales flotan en el mar hasta encontrar un gametofito femenino de la misma especie. El gametofito femenino genera carpogonios (10) que están equipados con una caña para pescar las espermatias llamada tricógino. Cuando un tricógino pesca una espermatia se da la unión (11). La unión es un proceso complejo que permite la fusión plasmogámica de la espora y el carpogonio permitiendo el ingreso del núcleo masculino.
Con la cariogamia (12) que ocurre inmediatamente se forma un cigoto (13) que va a desarrollar una estructura llamada quistocarpo (14). El quistocarpo es el propio carpogomnio que empieza a cambiar, pierde el tricógino y se combina con el quistoicarpo que en lugar de crear un individuo único lo que hace es una mitosis para generar esporas asexuales. Cuando las esporas han madurado la estructura se denomina carposporangio (14), y sus esporas se denominan carposporas (15), las cuales pueden regenerar mitoticamente (16) el tetrasporofito.
Reproducción y ciclo de vida de los glaucófitos
Lamentablemente no hay información disponible a cerca de los glaucófitos con respecto a sus ciclos de vida, sin embargo, al ser algas unicelulares es posible que tengan ciclos de vida semejante a otros protistas de vida libre. En estos casos una célula de vida libre realiza la meiosis y libera cuatro gametos, los cuales buscan a otros para combinarse y generar un cigoto que a su vez desarrolla nuevamente a la célula somática.
4. Las algas verdes y los líquenes
4. Las algas verdes y los líquenes
Estas son algas que están dentro del linaje de las plantas verdaderas o viridiplantae. Las hay unicelulares o multicelulares (Bonner, 2015; Bowman et al., 2016; Herron, 2016; Umen, 2014).
Reproducción y ciclo de vida de clamidomonas
Clamidomonas es un género de algas verdes unicelulares perteneciente a la familia Chlamydomonadaceae. Se caracterizan por su forma ovalada o periforme y su tamaño oscila entre 5 y 10 micrómetros. Estas algas poseen un flagelo, que les permite moverse en el agua, y un ocelo, un pequeño orgánulo que les ayuda a detectar la luz.
Clamidomonas es un organismo fotosintético y contiene cloroplastos que contienen clorofila a y b, lo que les da su color verde característico. Realizan la fotosíntesis para obtener energía, utilizando la luz solar y el dióxido de carbono.
Estas algas se encuentran en diversos hábitats acuáticos, como estanques, lagos y charcas. Se reproducen principalmente por división celular, donde una célula madre se divide en dos células hijas idénticas. Sin embargo, también pueden reproducirse sexualmente en ciertas condiciones.
Clamidomonas es objeto de estudio en la investigación científica debido a su importancia en los ecosistemas acuáticos y su utilidad en la comprensión de la biología celular y la evolución. Además, su capacidad de movimiento y su relativa facilidad de cultivo las convierten en organismos modelo en estudios experimentales y genéticos.
Generación haploide o gametofítica
Las algas verdes como las clamidomonas (YouTube) son seres vivos unicelulares y a diferencia de la mayoría de los miembros del reino Plantas, son motiles gracias a la presencia de flagelos. Antes de iniciar la reproducción asexual, la célula de la clamidomona pierde sus flagelos, posteriormente se realiza la mitosis al interior de la pared celular que permanece intacta, y solo se rompe cuando las dos células hijas han formado sus nuevos flagelos. Como cualquier mitosis, el proceso se repite una vez que cada célula hija ha alcanzado un tamaño crítico.

Figura 4.1. Colonia de clamidomonas.
Ocasionalmente, los flagelos no se generan, y las algas generan una colonia estable. Esto ocurre si las condiciones del ambiente son estables y propicias, pero si el ambiente se hace hostil, los flagelos se generan y las células escapan de la colonia para iniciar la etapa sexual. Algunas especies de clamidomonas forman estructuras multicelulares con una membrana externa que mantiene la colonia bajo coheción, en este caso no se las considera zoosporas ya que no han generado los flagelos. Las células vegetativas son por lo general haploides.
Generación diploide
Durante condiciones hostiles de luz, temperatura y otras fuerzas ambientales no conocidas, muchas células independientes de una colonia dispersa se pueden congregar nuevamente. Cada una de las células individuales sirve como gameto, y estos son atraídos por sus flagelos gracias a la producción de hormonas, semejante a como ocurre en los hongos.
Las células somáticas son al mismo tiempo gametos, y se los clasifica como isogametos ya que no hay diferencias entre los conjugantes, por tal razón se emplea la designación (+) y (-) para representar la conjugación fecundante, sin embargo, esta designación implica que los conjugantes son compatibles genéticamente, y estas células lo reconocen por medio de hormonas y receptores de membrana, las cuales les permite nadar hasta tocarse y luego reconocerse específicamente.
Aunque decimos que la célula vegetativa funciona como gameto, en realidad antes de hacer la fecundación deben completar su gametogénesis, para ello lo que hacen es que después de reconocer su compatibilidad genética, cada conjugante se despoja de la membrana exterior, lo cual puede ser interpretado como su etapa final de gametogénesis, la célula que emerge del interior si puede realizar la plasmogamia formando así el cigoto. Inmediatamente se forma una nueva pared celular. El cigoto mismo se convierte en una estructura de resistencia llamada zigospora y puede permanecer en dormancia desde varios días hasta meses, esperando que las condiciones ambientales se tornen favorables. Una vez el cigoto se reactiva da lugar la cariogamia y la meiosis formando cuatro zoosporas, las cuales al nacer se separan lo más posible unas de otras, y luego se desarrollan en clamidomonas vegetativas haploides.
Ciclo de vida
El ciclo de vida es dominado por células vegetativas haploides (1) que bajo condiciones hostiles inician su etapa sexual reconociendo a un conjugante genéticamente adecuado (2), cuando los conjugantes se reconocen se despojan de la membrana externa y realizan la fecundación (3) que da lugar a una célula diploide llamada zigospora (4) en honor a la zigospora de los zigomicetos. Esta zigospora realiza inmediatamente la meiosis (5) liberando de su interior cuatro células vegetativas funcionales.

Figura 4.2. Ciclo de vida de las clamidomonas. Tenga en cuenta que el tamaño y el nivel de especialización de las colonias de clamidomonas varía mucho entre los diferentes clados, desde simples masas, hasta cuerpos con tejidos casi verdaderos.
Adicionalmente hay dos tipos de reproducción asexual, una mitosis en la cual la célula vegetativa pierde el flagelo (6) produciendo dos células nuevas de su interior, o muchas más células en su interior generando una colonia (7) la cual puede disgregarse. En cualquiera de los dos casos, una vez liberadas, las células hijas regeneran los flagelos reiniciando el ciclo de vida.
Reproducción y ciclo de vida en ultroxidos
Ulothrix es un género de algas verdes filamentosas perteneciente a la clase de las Chlorophyceae. Estas algas son comunes en diversos hábitats acuáticos, como ríos, arroyos y estanques.
Ulothrix se caracteriza por su morfología filamentosa, formando largos filamentos compuestos por células en cadena. Cada célula tiene una forma alargada y cilíndrica. Al igual que otras algas verdes, Ulothrix contiene cloroplastos con clorofila a y b, lo que les confiere un color verde intenso.
Estas algas son fotosintéticas y utilizan la luz solar y el dióxido de carbono para realizar la fotosíntesis, produciendo oxígeno y azúcares como fuente de energía. Además, Ulothrix se reproduce tanto de forma asexual como sexual. En la reproducción asexual, los filamentos se fragmentan y cada fragmento puede dar lugar a un nuevo individuo. En la reproducción sexual, se forman gametos masculinos y femeninos que se fusionan para producir una nueva generación.
Ulothrix es un componente importante en los ecosistemas acuáticos, contribuyendo a la producción de oxígeno y sirviendo como alimento para organismos acuáticos. También se utiliza en estudios científicos y educativos como organismo modelo para entender diversos aspectos de la biología de las algas verdes.
Generación haploide o gametofítica
Los ulotrixos son algas verdes multicelulares en las cuales la generación gametofítica es dominante sobre la generación diploide, el ciclo de vida es semejante a clamidomonas, pero la forma colonial cambia de una estructura esférica a otra filamentosa y sésil. En la estructura filamentosa cada célula se organiza en una hilera semejante a la hifa de un hongo, la única célula especializada es la célula gancho “holdfast” (Figura 4.3) que evita que la corriente de agua se lleve al filamento.

Figura 4.3. Filamento de Ultrox.
Todas las células excepto el gancho son capaces de realizar la reproducción asexual y sexual. El proceso de mitosis se realiza al interior de cada segmento, formándose dos o más células hijas que forman flagelos motiles. Cuando se alcanza el punto crítico estas células llamadas zoosporas nadan alejándose lo más posible una de las otras, llegando a nadar durante días. Liberación de zoosporas, dependiendo del tipo de reproducción la zoospora cambia, si es asexual la zoospora tiene cuatro flagelos, pero si es sexual la zoospora tiene dos flagelos. Finalmente, cuando encuentran un buen sustrato realizan una primera mitosis para formar una nueva célula gancho y comienzan a realizar otras mitosis para reconstituir el filamento.
Reproducción sexual en ulotrixidos
La reproducción sexual y asexual en las algas verdes del linaje de los ulotrixos inicia del mismo modo. Los contenidos de cualquiera de las células del filamento excepto en el gancho se condensan y dividen. Lo cual produce esporas móviles. La diferencia entre la espora sexual y la asexual es la cantidad de flagelos, 4 en la asexual y 2 en la sexual. Un filamento puede llegar a producir 64 esporas sexuales, las cuales rompen las paredes del viejo filamento, y funcionarán como gametos sexuales. Cada gameto busca a un gameto de un tipo diferente por medio de hormonas segregadas en el agua, realizando la plasmogamia e inmediatamente la cariogamia.

Figura 4.4. Ciclo de vida de los ultróxidos.
El cigoto flota en el agua hasta encontrar un sustrato adecuado, donde inicia la meiosis. Las cuatro células hijas son zoosporas de cuatro flagelos que retomarán el ciclo de vida asexual. Los gametos de los ulotrixidos son isogámicos, eso implica que los gametos (+) y (-) son semejantes entre sí, y su única diferencia radica a nivel molecular. Al igual que los clamidomonidos, las células vegetativas son haploides, y no tienen acceso a la compensación de dosis, por lo que la presencia de alelos recesivos letales no es enmascarada.
Ciclo de vida
El ciclo de vida inicia cuando un filamento somático (1) genera la gametogénesis en sus células, liberando gametos zoospóricos de dos flagelos (2a) que se comportará ingual que las células somáticas de clamidomonas, los gametos isogámicos denominados “+” y “-“ (2b) experimentan la plasmogamiia y la cariogamia (3) generando un cigoto que mantiene los flagelos parentales, por lo que hablamos de un cigoto propagulante de tipo zoospórico (4). El cigoto nada activamente hasta encontrar un lugar donde asentarse (5). Una vez asentado el cigoto actúa como si fuera un esporofito, haciendo la meiosis (6) y liberando de su interior cuatro zoosporas de cuatro flagelos pero haploides (7). Estas zoosporas se asientan (8), y regeneran mitóticamente (9) el filamento (1). El filamento también puede regenerar esporas asexuales (7) generando un epiciclo asexual. Adicionalmente la colonia puede fragmentarse y regenerar dos o más colonias dependiendo del fragmento, lo cual implica un segundo epiciclo asexual, pero que no es obligatorio para completar el ciclo de vida.
Reproducción en espirogiridos
Spirogyra es un género de algas verdes filamentosas perteneciente a la clase de las Chlorophyceae. Son comunes en hábitats acuáticos dulces, como estanques, lagos y arroyos.
Las especies de Spirogyra se caracterizan por sus filamentos largos y delgados formados por células cilíndricas que se agrupan en hileras. Estas células están unidas por puentes citoplasmáticos y están rodeadas por una cubierta gelatinosa llamada mucílago.
Cada célula de Spirogyra contiene uno o más cloroplastos en forma de espiral, de ahí el nombre del género. Estos cloroplastos contienen clorofila a y b, lo que les da un color verde intenso y les permite llevar a cabo la fotosíntesis. A través de la fotosíntesis, las algas Spirogyra convierten la luz solar y el dióxido de carbono en oxígeno y azúcares, desempeñando un papel crucial en el equilibrio de los ecosistemas acuáticos.
Spirogyra se reproduce tanto de forma asexual como sexual. En la reproducción asexual, los filamentos se fragmentan y cada fragmento puede desarrollar una nueva colonia. En la reproducción sexual, se forman estructuras especializadas llamadas gametos, que se fusionan para formar zigotos que luego germinan y forman nuevas colonias.
Estas algas verdes tienen importancia ecológica y también son objeto de estudio en investigación científica debido a su estructura filamentosa, su capacidad de reproducción y su papel en los ecosistemas acuáticos.
Reproducción asexual en espirogiridos
A diferencia de los clamidomonidos y ulotrixidos, los espirogiridos (Figura 4.5) no producen zoosporas u otras células con flagelos. Al igual que los ulotrixidos, los esporogiridos poseen un modo de vida vegetativo filamentoso. Cualquier célula es capaz de dividirse por mitosis, pero la única reproducción asexual posible es la que resulta del rompimiento mecánico de un filamento. El fragmento flotante puede reconstruir el gancho y vivir en un nuevo sustrato.

Figura 4.5. Filamento de Spirogyra.
Reproducción sexual en espirogiridos
Las colonias de filamentos de los espirogiridos crecen muy cerca los unos con los otros, lo cual es vital para su peculiar forma de reproducción sexual. Cuando la reproducción sexual da inicio, las células individuales de los filamentos adyacentes empiezan a producir una serie de abultamientos denominados papillae “singular, papilla”. Estas papilas crecen unas contra otras, primero forzando los filamentos a separarse, y finalmente permitiendo que la pared celular se rompa creando un puente para el citoplasma de los compartimentos adyacentes.
El contenido de cada compartimento es una célula haploide individual que funciona tanto como órgano vegetativo como gameto. Debido a diferencias genéticas, uno de los gametos se mantiene estacionario “y se lo considera femenino” mientras que el otro “considerado masculino” atraviesa el puente para realizar la plasmogamia. El cigoto entra inmediatamente en meiosis, pero de forma similar a los óvulos de las mujeres humanas, solo uno de los cuatro productos de la meiosis llega a buen término. Una vez se ha determinado el sobreviviente, inicia una etapa de mitosis, lo cual implica que un nuevo filamento crecerá encima del viejo. A este tipo de reproducción sexual se lo denomina conjugación, y no debe ser confundida con la otra conjugación que llevan a cabo las bacterias.
Reproducción y ciclo de vida de los oedogonidos
Los oedogonidos también son algas filamentosas. Su reproducción asexual se realiza por medio de zoosporas o por medio de fragmentación vegetativa. Las células reproductivas asexuales o zoosporas se producen únicamente en las células de la punta de cada filamento, pero su característica más definitoria es que la zoospora se moviliza por la presencia de hasta 120 pequeños flagelos que forman una superficie aterciopelada en uno de los polos de la célula. Una vez que las zoosporas se han separado de la colonia original (Figura 4.7) y encontrado un sustrato adecuado proceden a asentarse de forma similar a como realizan los ulotrixidos.

Figura 4.6. Colonia de Oedogonium.

Figura 4.7. Zoospora de Oedogonium.
Reproducción y ciclo de vida de los líquenes
Los líquenes son una relación simbiótica entre un hongo, generalmente un ascomiceto y un alga, lo cual implica un ciclo de vida relativamente complejo, que involucra la conjunción de dos ciclos de vida dependientes el uno del otro. Adicionalmente, ha sido complicado determinar las etapas del ciclo del alga, por lo que el mejor esquema que he encontrado sobre el ciclo de vida de los líquenes marca como dominante al hongo, y el alga es un acompañante (Bonner, 2015; Klees & Piepenbring, 2017; Sanders, 2014).
De acuerdo a Sanders (2011) el ciclo de vida del ascomicético liquenizado posee bastantes modificaciones con respecto a un ascomiceto normal. Tal vez lo más raro es que la ruta de la reproducción sexual del ascomiceto sigue por la producción de los conodios.

Figura 4.8. El ciclo de vida de los líquenes. Un líquen es una simbiosis entre un hongo y un alga, de allí que su ciclo de vida sea complejo y paralelo.
En cualquier caso, el ciclo de vida inicia con el talo somático (1) que es con conjunción entre un alga y el micelio (1a-b) el cual posee varias formas, que crecen sobre las rocas (1c). Desde aquí, puede haber dos rutas, una que ha sido reportada y que es asexual, y una presumida debido a análisis genéticos, pero cuyas estructuras aún no han sido aisladas que es la sexual. Iniciamos pues con el ciclo sexual, aunque se presuma, el talo produce estructuras reproductoras esporulantes (2) que se denominan picnidios (3a-b) los cuales crecen casi al interior del talo como si fueran botellas pigmentadas, a dentro de las cueles están los conodioforos y los conidios apretujados. En el esquema original de Sanders (2011) muestran dos picnidios de diferentes colores lo cual implicaría la existencia de picnidios diferentes que generan conodiosporas sexualizadas (4) diferentes, las cuales al germinar producen las hifas gaméticas (5?) las cuales se presume que experimentan la plasmogamia (6?). Desde este punto se supondría que el hongo sigue parte de su ciclo ascomicete normal, con la formación del micelio dicarionte (7) que regenera (8) un ascocarpo especializado denominado apotecio (9a-c).
partir de aquí se seguiría el proceso de formación de la ascospora (10-16), aunque la ascospora es diferente de la ascospora normal (17a-b), ya que es grande y germina en varios puntos (18) aunque solo prospera si logra asociarse a un alga, además el talo puede regenerarse por fragmentación (19). El alga es denominada fotobionte y el hongo micobionte. A pesar de que el ciclo sexual mayor tiene varias etapas por aislar, el ciclo asexual menor parece ser el mejor reportado, en este caso el talo produce estructuras asexuales denominadas campilidios (20a-c) en los cuales se desarrollan esporas que crecen hasta ser hifas lo bastante grandes como para estar asociadas a una célula del alga, cuando son expulsados, estos propágulos complejos son denominados hifa aposimbiótica (21) la cual también puede regenerar el talo somático.
5. Las briofitas
5. Las briofitas
Los musgos y en general los briofitos son uno de los grupos de plantas terrestres más antiguos, sino el más antiguo, y su modo de vida no ha cambiado mucho, por lo que muchos de sus rasgos se consideran ancestrales. Las briofitas, que incluyen musgos, hepáticas y antocerotas, son un grupo de plantas no vasculares que se consideran uno de los linajes más antiguos de plantas terrestres. Se cree que las briofitas evolucionaron hace aproximadamente 470 millones de años, durante el período Ordovícico de la era Paleozoica. Aunque las briofitas pueden haber experimentado una disminución en su diversidad y abundancia durante el Gran Desierto Pérmico, lograron sobrevivir en hábitats refugio y posteriormente repoblaron las áreas disponibles cuando las condiciones mejoraron. Su adaptación a ambientes húmedos y su capacidad de reproducción asexual les permitieron persistir y continuar evolucionando hasta el día de hoy.

Figura 5.1. Musgo creciendo en tierra húmeda.
Dos rasgos importantes empiezan a ser evidentes en los musgos: En primera instancia, son plantas con una especialización celular notable, en la que la dependencia del agua empieza a limitarse, es decir, empiezan a ser más independientes de los ambientes húmedos. En segunda instancia, la generación diploide empieza a ser mucho más importante que un único cigoto, en los musgos la generación diploide ya compone una estructura multicelular con especialización de tejidos y órganos, aunque aún se encuentra fuertemente asociada a la generación haploide sobre la cual crece. En última instancia, la reproducción sexual ya no es simplemente un mecanismo de emergencia para producir descendientes variables cuando el ambiente se torna hostil, en los musgos la reproducción se torna predominantemente sexual (Bowman et al., 2016; Haig, 2016; Harholt, Moestrup, & Ulvskov, 2016; Kofuji & Hasebe, 2014; Petersen & Burd, 2016; Willis & McElwain, 2014).
El musgo
Los musgos son las plantas más tipológicas de los briofitos y por lo tanto las analizaremos primero (YouTube). Sus ciclos de vida poseen dos características, el primero es que el gametofito es la etapa fotosíntética somática que afronta la selección natural en su mayor parte, por lo que se dice que es un ciclo de vida en el cual domina el gametofito. Y la segunda es que el cigoto no es un propágulo, así que debe crecer como un parásito a partir de los nutrientes que sustenta el gametofito, así que decimos que el esporofito es parasito del gametofito. Los musgos se caracterizan por un ciclo de vida complejo con alternancia de generaciones.

Figura 5.2. Esporulación (YouTube).
Generación gametofítica
Esta es la generación de un cuerpo capaz de crear gametos. El gametofito del musgo es la etapa dominante y más conspicua de su ciclo de vida. Es la fase haploide y sexualmente reproductiva de la planta.
(a) Nacimiento a partir de una espora. Cuando la cabeza del esporangio explota libera esporas haploides. Las esporas son propágulos, pero no estructuras de resistencia, de hecho, todo este proceso ocurre en una temporada lluviosa, ya que se requiere agua para la fecundación y también se requiere agua para que las esporas puedan germinar, y además se requiere agua para que los gametofitos puedan sobrevivir, es por esto que el musgo está limitado a zonas muy húmedas.
(b) Crecimiento del gametofito. Las esporas que germinan por sí mismas se desarrollan por mitosis hasta volver a formar el gametofito completando el ciclo de los musgos. Cuando la espora germina (A) se reproduce por mitosis generando una fila de células semejante a las hifas de los hongos (B, C, D, E, F). Pero a diferencia de los hongos las células en mitosis forman rápidamente tejidos especializados como los rozoides para sostenerse del sustrato y las estructuras fotosintéticas del gametofito inmaduro (G). Finalmente, el gametofito vuelve a crecer (H).

Figura 5.3. Desarrollo del gametofito.

Figura 5.4. El gametofito. Los gametofitos son la generación visible, pero son enanos y se ven como un terciopelo verde sobre las rocas.
En resumen, los musgos son plantas terrestres que dependen mucho de los ecosistemas acuáticos, siendo un linaje de transición, viven en tierra seca. pero sin agua constante nunca se reproducen.
(c) El gametofito. El cuerpo que generalmente vemos de los musgos se denomina gametofito que significa cuerpo generador de gametos, es una estructura multicelular que se alimenta por fotosíntesis, carece de sistema vascular y por lo tanto es extremadamente pequeño. El gametofito del musgo tiene un tallo corto y ramificado, a menudo cubierto por células alargadas llamadas filamentos. Estos filamentos proporcionan soporte y ayudan a la absorción de agua y nutrientes. En el gametofito del musgo, las hojas son simples y se disponen en espiral alrededor del tallo. Son generalmente delgadas, con una forma lanceolada o ovada, y pueden tener una estructura laminar o filamentosa.


Figura 5.5. El arquegonio. Los arquegonios se agrupan en una estructura de forma de botella grande y agrupa a varios arquegonios individuales. A su vez, cada arquegonio individual es una estructura en forma de botella donde se realiza la gametogénesis femenina.

Figura 5.6. El anteridio y los espermatozoides. Los anteridios en plural también se forman como racimos estrechamente agrupados que contienen anteridios individuales, los cuales poseen forma de sacos al interior de los cuales se prosigue con la gametogénesis.

Figura 5.7. Los gametos masculinos son flagelados y móviles, pero deben esperar a que el gametofito esté sumergido para poder desplazase.
El musgo es una planta de estatura muy baja que forma tapices aterciopelados en zonas húmedas y poco contaminadas. Gametofito es el cuerpo que se alimenta y su genotipo es haploide (n).
Los gametofitos de los musgos son dioicos, por lo que habrá individuos masculinos y femeninos, pero las diferentes son microscópicas y solo se manifiestan en el ápice o punta de la planta. En las puntas del gametofito se desarrollan de forma microscópica estructuras especializadas llamadas anteridios para los machos y arquegonios para las hembras.
El gametofito más impresionante de los musgos actuales y de todos los tiempos es el de la especie Dawsonia superba, también conocida como "Gran musgo de Tasmania" o "Gran musgo de Nueva Zelanda". Este musgo se encuentra en las regiones templadas y tropicales de Australia, Nueva Zelanda y otras áreas del Pacífico.

Figura 5.8. Dawsonia superba, conocida como "Gran musgo de Tasmania" o "Gran musgo de Nueva Zelanda", es una especie de musgo que se encuentra en las regiones templadas y tropicales de Australia, Nueva Zelanda y otras áreas del Pacífico. Su gametofito es excepcional en tamaño, pudiendo alcanzar alturas de hasta 50 centímetros. Tiene un tallo ramificado con filamentos foliares que forman una estructura frondosa densa. Es una adaptación al hábitat de los bosques húmedos y cumple un papel importante en la retención de agua y la creación de microhábitats.
El gametofito de Dawsonia superba es excepcional en términos de tamaño y estructura. Puede alcanzar alturas de hasta 50 centímetros, lo que lo convierte en uno de los gametofitos más grandes entre todas las especies de musgos. Además, tiene un tallo ramificado con filamentos foliares que forman una estructura frondosa densa.
La impresionante apariencia del gametofito de Dawsonia superba se debe a su adaptación al hábitat de los bosques húmedos. Sus grandes dimensiones y su frondosidad le permiten captar la máxima cantidad de luz solar en áreas sombreadas y competir por recursos con otras plantas. Este musgo tiene un papel ecológico importante al retener agua y ayudar a mantener la humedad en su entorno.
Es importante destacar que Dawsonia superba es solo una especie notable entre la diversidad de gametofitos de musgos existentes. Aunque es uno de los más impresionantes en términos de tamaño, otros musgos también tienen características fascinantes y únicas en sus gametofitos.
Generación diploide, el esporofito
Cuando llueve lo suficiente como para que el gametofito se sumerja los espermatozoides nadan hacia los arquegonios guiados por hormonas. Una vez que se fertiliza al óvulo el embrión en crecimiento comienza a desarrollarse de manera parásita al gametofito, con una estructura en forma de bombón llamada esporofito, una estructura diploide.

Figura 5.9. El esporofito. El esporofito es pequeño y crece como un parásito del gametofito.
El esporofito crece por meiosis alimentándose por nutrientes del gametofito de manera parasítica y/o por fotosíntesis propia si es que posee los pigmentos necesarios. La punta del esporofito maduro se denomina cápsula y es un esporangio, allí las células realizan la meiosis, pero sin desarrollarse en gametos, es decir en la meiosis intermedia los conceptos de meiosis y gametogénesis son diferentes. Estructuralmente hablando el único miembro de la generación diploide es el esporofito, ya que los propágulos que este genera son productos meióticos denominados esporas, los cuales regenerarán a los gametofitos de la generación diploide. Las células haploides no gaméticas se almacenan hasta que el esporofito esté listo, luego la cabeza del esporangio explota liberan a estas células conocidas como esporas.
Reproducción asexual en los musgos
Cierta reproducción en los musgos puede realizarse independientemente del ciclo sexual, sin embargo, su potencialidad solo ha sido demostrada en condiciones de laboratorio. Debido a que la diferenciación celular de los tejidos de los arquegonios, anteridios, rizomas y parafisos “hojas” pueden reasumir el crecimiento de individuos completos cuando crecen en medios de cultivo adecuados. Sin embargo, esta propiedad es muy limitada, se ha reportado que solo el 12% de un grupo de muestras de musgos del ártico pudieron reasumir el crecimiento a individuos completos. Se ha demostrado que esta reproducción vegetativa por vegetación puede llegar a ser más importante en regiones con climas más favorables, donde el viento es un factor más importante. Debido a que la reproducción vegetativa no es un factor relevante, este es uno de los primeros ciclos de vida en los que podemos afirmar que la reproducción depende casi de manera exclusiva en el ciclo sexual.
Ciclo de vida
El esporofito (1) en su ápice o cápsula (2) induce a las células internas a realizar la meiosis con la formación y liberación (3) de esporas (4). Las esporas germinan y regeneran (5) un gametofito dioico (6a-b) que pose estructuras masculinas y femeninas simultáneamente. La estructura masculina es denominada anteridio (7) los anteridios crecen justo en el ápice (7a) como si se tratara de un plato muy agrupado, mientras que los anteridios propiamente dichos (7b) son bolsas que contienen las células espermáticas.

Figura 5.10. El ciclo de vida del musgo
El anteridio produce la gametogénesis masculina para generar a los espermatozoides (8), que deben esperar a que el gametofito esté sumergido en agua para poder nadar hacia los arquegonios (9). Los arquegonios son las estructuras sexuales femeninas (9), estos se encuentran también en el ápice protegidos y apretujados (9a), mientras que el anteridio propiamente dicho guarda en su interior a los óvulos (9c). Ambos gametofitos son cortos, por lo que pueden ser cubiertos de agua, permitiendo que los espermatozoides naden y fecunden a los óvulos (10). El cigoto no propagúla, sino que germina internamente en el gametofito femenino, desarrollando (11) un esporofito parasítico (1).
Los antoceros
Los antóceros, también conocidos como antocerotofitas, son un grupo de plantas no vasculares que pertenecen a la división Anthocerotophyta. Aunque comparten algunas características con los musgos y las hepáticas, también presentan diferencias distintivas.
Una de las principales diferencias entre los antóceros y los musgos o las hepáticas radica en su morfología. Mientras que los musgos y las hepáticas tienen una estructura foliar más compleja, con hojas y tallos bien diferenciados, los antóceros carecen de tallos y hojas verdaderas. En cambio, tienen un cuerpo gametofítico aplanado, similar a una lámina, conocido como talo. Este talo tiene una forma plana y generalmente forma una especie de cinturón o banda en el suelo.
Otra diferencia es la disposición de los órganos reproductores. Los antóceros tienen estructuras reproductivas llamadas esporófitos, que son más complejas que las de los musgos y las hepáticas. Estos esporófitos están compuestos por un cápsula que contiene esporas y un pie que las sostiene. A diferencia de los musgos y las hepáticas, los antóceros no tienen una fase esporofítica prolongada y la mayor parte de su ciclo de vida está dominada por el gametofito.
Además, los antóceros tienen células que producen un pigmento oscuro llamado antocianina, de ahí su nombre. Este pigmento les da un color característico, a menudo oscuro o negro, que contrasta con el verde más común en los musgos y las hepáticas.
Los antoceros poseen ciclos de vida semejantes, pero su principal característica distintiva es que su gametofito es monóico.

Figura 5.11. Ciclo de vida de los antoceros.
El esporofito (1) en su ápice o cápsula (2) induce a las células internas a realizar la meiosis con la formacvión y liberación (3) de esporas (4). Las esporas germinan y regeneran (5) un gametofito hermafrodita (6) que pose estructuras masculínas y femeninas simultáneamente. La estructura masculina es denominada anteridio (7) el cual se ubica hacia el centro del gametofito, ya que este crece horizontalmente y no verticalmente como los musgos. El anteridio produce la gametogénesis masculina para generar a los espermatozoides (8), que deben esperar a que el gametofito esté sumergido en agua para poder nadar hacia los arquegonios (9). Los arquegonios son las estructuras sexuales femeninas, y en su interior se encuentran los óvulos (10). Los arquegonios también se encuentran hacia el centro del gametofito y cuando experimentan la fecundación (11) generan un esporofito diploide (1) y parasítico que se ancla al gametofito y crece alimentándose de él hasta alcanzar la madurez y completar el ciclo de vida.
Hepáticas
Las hepáticas son plantas no vasculares que pertenecen al grupo de las Bryophyta. Son conocidas por su apariencia similar a un hígado, de ahí su nombre. Las hepáticas se encuentran en una amplia variedad de hábitats terrestres, desde bosques húmedos hasta desiertos áridos.

Figura 5.12. Gametofito y esporofito de las hepáticas. Gametofito (parece una hoja grande abajo) y esporofito (parece una flor). La forma del portador de gametos “gametangioforo” revela si es femenino o masculino.
A diferencia de los musgos y los antóceros, las hepáticas tienen un cuerpo gametofítico más plano y a menudo forman estructuras parecidas a almohadillas o tapetes. Estas estructuras están compuestas por células que forman láminas, a veces con forma lobulada, en lugar de tener una forma alargada como los musgos.
Otra característica distintiva de las hepáticas es la presencia de estructuras reproductivas especializadas llamadas gametangios. Los gametangios en las hepáticas se conocen como arquegonios y anteridios. Los arquegonios son estructuras que contienen los óvulos, donde se produce la fertilización, y los anteridios son estructuras que producen los gametos masculinos (espermatozoides). Estas estructuras reproductivas pueden estar ubicadas en tallos, en los bordes de las hojas o en estructuras en forma de disco llamadas yemas.
En cuanto a la reproducción, las hepáticas pueden reproducirse tanto asexualmente como sexualmente. La reproducción asexual ocurre a través de fragmentación o por la formación de yemas. En la reproducción sexual, los espermatozoides nadan a través del agua para llegar a los óvulos y fertilizarlos, lo que da lugar a la formación de esporofitos.
Generación gametofítica
Como en las demás briofitas, la generación del gametofito haploide es la más visible, aunque en estas se conoce como el talo.
(a) Gametofito tálico. La hepática tipo que se representa generalmente es Marchantia spp. En este caso tenemos un cuerpo somático haploide que denominamos gametofito, el gametofito de las hepáticas es dioico. La principal diferencia de las hepáticas con respecto a otros briófitos es que además del talo gametofítico fotosintético, también se generan estructuras altamente especializadas que crecen del talo llamados portadores (foros). Dependiendo de si el talo es masculino o femenino los clasificamos como anteridioforos (portadores masculinos) o arquegonioforos (portadores femeninos).

Figura 5.13. Marchantia polymorpha: Conocida comúnmente como hepática común, es una especie ampliamente distribuida en todo el mundo. Tiene un aspecto distintivo con un cuerpo plano y lobulado, que parece un tapiz verde en el suelo. Sus células reproductivas se encuentran en estructuras llamadas gametofitos, que forman pequeñas copas en la superficie de la planta.
A parte de los gametangioforos masculinos y femeninos, los gametofitos tálicos poseen otras partes, la parte principal del talo es una estructura en forma de hoja aplanada que se encarga de la fotosíntesis, pero aparte hay otras dos partes menos evidentes. Los rizoides que se encargan de afianzar la estructura al sustrato y las copas de gemación. Las copas de gemación producen estructuras especializadas para la reproducción asexual vegetativa llamadas gemas, que son fragmentos de tejido que pueden regenerar al talo. Dado que tratamos con fragmentos de tejido que no poseen mayor capacidad de resistencia no las denominamos ni esporas ni quistes.

Figura 5.14. Ciclo de vida de las hepáticas. En este modelo tenemos una representación detallada de los gametofitos masculino y femenino.
(b) Anteridioforos y anteridios. Los anteridioforos poseen dos partes, un tallo y el anteridioforo propiamente dicho, siendo en general estructuras con forma de plato, los anteridios se ubican en la parte superior del anteridioforo y exponen sus aperculos liberadores de gametos hacia “arriba”. Al interior de los anteridios se da la gametogénesis masculina para producir los espermatozoides que deben esperar a que el anteridioforo esté sumergido en agua.

Figura 5.15. Gametangios de las hepáticas. (Izquierda) Anteridioforos y anteridios; (Derecha) Arquegonioforos y arquegonios.
(c) Arquegonioforos y arquegonios. Los arquegonioforos también poseen un tallo de crecimiento y en su ápice el arquegonio propiamente dicho, poseen una forma de sombrilla con unos vértices que parecen dedos carnosos. Debajo de la sombrilla del arquegonioforo se encuentran los arquegonios ubicados hacia abajo. Por lo general la fertilización se da antes de que el tallo del arquegonioforo hubiera terminado de crecer.
Generación esporofítica
El esporofito crece debajo del arquegonioforo, siendo absolutamente parasítico del gametofito, y además lo único que desarrolla realmente es su cápsula. Cuando el esporofito se desarrolla completamente la única evidencia de su presencia es que los palpos del arquegonioforo se abren exponiendo las cápsulas al aire. Por esto podemos afirmar que en las hepáticas es donde podemos ver la mayor dominancia del gametofito sobre el esporofito.

Figura 5.16. Esporofito parásito.
Ciclo de vida
El esporofito dependiente del arquegonioforo (1) realiza la meiosis y liberar (2) las esporas sexuales (3) que flotan en el agua hasta encontrar un lugar donde germinar (4) y regenerar así un gametofito tálico (5a-b). El talo es dioico y pueden generar estructuras reproductivas asexuales llamadas gemas (6) que regeneran más talos o generar los gametangioforos que serían algo así como los verdaderos gametofitos.

Figura 5.17. Ciclo de vida de las hepáticas.
El gametofito masculino (7) se llama anteridioforo y sobre su estructura en forma de plato crecen los anteridios (8) que generan los espermatozoides (9). El gametofito femenino (10) es denominado arquegonioforo, el cual posee forma de sombrilla, debajo de la sombrilla se desarrollan los arquegonios (11) que deben ser fecundados (12) antes de que el tallo del arquegonioforo haya terminado de crecer. El cigoto diploide genera (13) un esporofito imperfecto (1) que solo desarrolla la cápsula completando del ciclo de vida.
6. Helechos
6. Helechos
Helechos, licopodios y antócereos representan un momento crucial en la evolución de las plantas. El esporofito es la clave en esta revolución, ya que adquiere tejidos vasculares llamados floema y xilema, el cual le permite crecer a tal nivel que aplasta a su gametofito. Evidentemente, en estas plantas el esporofito domina sobre el gametofito como cuerpo somático, aunque la mayoría de los procesos se dan en la generación gametofítica (Bateman & DiMICHELE, 1994; Cousens, 1988; Essig, 2015; L. E. Graham, 1985; Ligrone, Duckett, & Renzaglia, 2012; Mehltreter, Walker, & Sharpe, 2010; Niklas & Cobb, 2016; Schneider, Pryer, Cranfill, Smith, & Wolf, 2002; Sessa & Der, 2016).
Los helechos son plantas aún más independientes del agua que los musgos, antoceros y hepáticas, pero aun la requieren para que sus gametos puedan encontrarse entre sí. A diferencia de las plantas con semillas, los helechos pasan por una fase en que sus gametos masculinos deben nadar en el agua para encontrar a los femeninos, ergo, sin agua no hay reproducción. Adicionalmente los tamaños de las generaciones siguen cambiando. En las hepáticas el gametofito es más grande que el esporofito, en los musgos los tamaños se hacen menos conspicuos y en los antoceros son casi del mismo. En los helechos y parientes cercanos, el esporofito es mucho más grande que el gametofito que permanece enterrado e invisible. Los helechos también son capaces de la reproducción asexual.
Los helechos son plantas vasculares que se caracterizan por una alternancia de generaciones en las que el esporofito “cuerpo diploide donde se realiza la meiosis y se generan las esporas” es dominante sobre el gametofito que empieza a hacerse más pequeño. Aunque al inicio el proceso es semejante en el sentido de que el esporofito inicia su vida al interior del gametofito, siendo inicialmente parasítico con respecto a su gametofito, la diferencia es que en este caso el esporofito crece desproporcionadamente, posiblemente destruyendo al gametofito del cual se originó. Otra característica del ciclo de vida de los helechos es que el gametofito es hermafrodita produciendo simultáneamente espermatozoides y óvulos en diferentes partes de su estructura.
Evolución y registro fósil
Los helechos son plantas vasculares antiguas que han existido en la Tierra durante millones de años. Su evolución se remonta al período Devónico, hace aproximadamente 360 millones de años. Los primeros fósiles de helechos muestran características distintivas, como hojas grandes y divididas en pequeños segmentos llamados pinnas. Estas características se han mantenido en muchas especies de helechos a lo largo del tiempo.
Uno de los especímenes fósiles más impresionantes es el helecho arborescente del género Psaronius. Estas plantas gigantes del período Carbonífero tenían troncos parecidos a palmeras y hojas que podían alcanzar hasta 10 metros de longitud. También se han descubierto fósiles de helechos con estructuras reproductivas bien conservadas, como esporangios, que revelan cómo se reproducían y dispersaban en el pasado.

Figura 6.1. La especie de Psaronius más grande conocida es Psaronius brasiliensis. Esta planta del período Carbonífero era un helecho arborescente que crecía en los bosques pantanosos de lo que ahora es Brasil. Se estima que alcanzaba alturas impresionantes de hasta 15 metros, lo que la convierte en una de las plantas más altas de su época. Psaronius brasiliensis tenía un tronco robusto y erecto, similar a una palmera, con una corona de hojas frondosas en la parte superior. Sus hojas eran grandes, llegando a medir hasta 2 metros de longitud, y estaban divididas en pequeños segmentos llamados pinnas. Esta especie de Psaronius prosperaba en un ecosistema de bosque pantanoso y húmedo. Los suelos fértiles y la abundancia de agua proporcionaban las condiciones ideales para su crecimiento. Estos helechos arborescentes formaban parte de una comunidad vegetal diversa que incluía otras plantas del período Carbonífero, como licofitas y equisetos. Psaronius brasiliensis desempeñaba un papel importante en el ecosistema al proporcionar refugio y alimento para diversas especies animales. Sus grandes frondas ofrecían sombra y protección a criaturas terrestres y acuáticas, como anfibios, insectos y pequeños reptiles. La existencia de Psaronius brasiliensis y otras especies de Psaronius en los bosques pantanosos del pasado nos brinda una visión fascinante de la diversidad y el tamaño impresionante que las plantas pueden alcanzar en ecosistemas antiguos.
Durante la era de los dinosaurios, los helechos experimentaron una diversificación y proliferación significativas. Muchas especies se adaptaron a diferentes hábitats y ocuparon nichos ecológicos diversos. Los helechos fueron una parte importante de los ecosistemas terrestres y proporcionaron alimento y refugio a numerosas criaturas, incluidos los dinosaurios.

Figura 6.2. Cyathea brownii: Conocido como helecho de tronco negro, es nativo de Nueva Zelanda y puede alcanzar alturas impresionantes de hasta 20 metros. Tiene un tronco delgado y negro que se eleva sobre el dosel del bosque, y sus frondas largas y elegantes se extienden en todas las direcciones, creando una apariencia majestuosa.
A lo largo del tiempo, los helechos han cambiado en su tamaño y diversidad, y algunas especies se han adaptado a condiciones ambientales cambiantes. En la actualidad, se estima que existen alrededor de 12,000 especies de helechos en todo el mundo, que van desde pequeñas especies herbáceas hasta helechos arborescentes de gran tamaño.
Generación gametofítica en helechos
La generación gametofítica de los helechos es un fascinante aspecto de su ciclo de vida. En este artículo, exploraremos en detalle la fase gametofítica de los helechos, centrándonos en su estructura, desarrollo y función en el proceso de reproducción. Analizaremos cómo los gametofitos producen y albergan las células reproductivas masculinas y femeninas, y cómo ocurre la fertilización. También examinaremos la diversidad de formas y tamaños de los gametofitos en diferentes especies de helechos, destacando aspectos interesantes y peculiares. Comprender la generación gametofítica de los helechos nos brinda una visión más completa de su complejo ciclo de vida y su importancia en la conservación y evolución de estas fascinantes plantas.

Figura 6.3. Angiopteris evecta: También conocido como helecho rey, es nativo de regiones tropicales de Asia y el Pacífico. Esta especie puede alcanzar alturas de hasta 6 metros. Sus frondas son grandes y brillantes, con una disposición en forma de abanico, y su apariencia exuberante y llamativa lo convierte en una especie impresionante.
Esporas
Al interior de los soros se encuentran varios espoirangios, y al interios de estos se encuentran los esporocitos que llevan a cabo la meiosis, usualmente produciendo entre 48 a 64 esporas por cada esporangio, sin embargo, especies de estrategia de vita r pueden producir hasta 15000 esporas por esporangio. En especies semiacuáticas de helechos “Marsilea y Salvinia” pueden producirse dos tipos de esporas, las megasporas y mircosporas generando gametofitos dioicos, pero en la vasta mayoría se generan esporas monoicas que producirán a su vez gametofitos hermafroditas. Las esporas sin dispersadas por el aire, pero deben caer en ambientes húmedos que permitan el desarrollo del gametofito.
Gametofito
El gametofito o protalo es la fase más pequeña del talo, mide tan solo unos centímetros y su vida es bastante breve, ya que rara vez sobrevive unas semanas. Tiene forma acorazonada y está fijado al sustrato por unos rizoides. En él se forman los anteriodos, que forman los gametofitos masculinos, y los arquegonios, que forman los femeninos. En algunos casos estos órganos sexuales aparecen en protalos diferentes, pero lo normal es que aparezcan en el mismo. La llegada de agua a los órganos sexuales hace que se liberen los espermatozoides, de forma helicoidal, que nadan hasta los arquegonios, lugar donde se produce la fecundación. Tras ella, se formará de nuevo un esporófito.
Los gametofitos generalmente se enroscan levemente en sus bordes, y pueden llegar a medir entre 5 y 6 milímetros, por lo que son visibles sin microscopio, aunque muchos de sus detalles finos requieren ser expuestos a un estereoscopio. Muchos gametofitos son muy delgados, siendo de tan solo una célula de grosor, por lo que carecen de tejidos vasculares de transporte, de allí que aun cuando son fotosíntéticos, sus tamaños no superan al de sus parientes más viejos briófitos.

Figura 6.4. Gametofito. Una tendencia de los grupos más recientes de plantas, y también más terrestres, es el hecho de que su gametofito se hace más pequeño, y la generación dominante es el esporofito diploide, posiblemente debido a que al tener dos copias de cada gen son más adaptables de los gametofitos haploides.

Figura 6.5. Regiones sexuales del Gametofito. El gametofito de los helechos no solo es más pequeño, también es hermafrodita, portando ambos gametangios.
Gametangios
Los anteridios se agrupan de manera aletaoria de manera cercana a los rizoides, aunque en algunas especies puede distribuirse en todo el gametofito, mientras que los arquegonios se ubican centra al vértice central de la estructura en forma de corazón. Los arquegonios tienen forma de botella de cuello corto con una abertura que permite el paso de los espermas, siempre y cuando el gametofito se encuentre sumergido en agua. Un solo anteridio piede producir de 32 a varios cientos de espermas, cada uno con pocos o muchos flagelos. El óvulo por su parte se encuentra bloqueado e inmobilizado al interior del arquegonio. Como es tradición, las células masculinas son de menor calidad y mas numerosas, a demás de que se pueden mobilizar, mientras que la célula femenina es menor en número, de mayor calidad e inmóbil.


Figura 6.6. Detalles de los anteridios (arriba) y los arquegonios (abajo).

Figura 6.7. Espermas de los helechos.
Fertilización
La fertilización solo se da cuando el gametofito queda sumergido en agua, igual que sucede con las briofitas mas primitivas. Con agua, los espermas de los anteridios pueden nadar atraidos por hormonas segregadas por los arquegonios. Esta es una carrera contra el reloj, pues aun cuando cada gametofito puede producir varios arquegonios, solo uno de ellos podrá generar un esporofito.
Generación esporofítica en helechos
La generación esporofítica de los helechos es un aspecto fundamental de su ciclo de vida. Estos organismos presentan una alternancia de generaciones entre el esporofito y el gametofito. En este artículo, exploraremos en detalle la fase esporofítica de los helechos, centrándonos en su estructura, desarrollo y función en la reproducción. Analizaremos cómo los esporofitos producen esporas a través de los esporangios y cómo estas se dispersan en el entorno. También examinaremos la diversidad de formas y adaptaciones de los esporofitos en diferentes especies de helechos, destacando aspectos interesantes y peculiares. Comprender la generación esporofítica de los helechos nos permitirá apreciar su importancia en el ciclo de vida y en la ecología de estos fascinantes organismos.
Embrión
Una vez que se fertiliza al óvulo el embrión en crecimiento comienza a desarrollarse. Lo primero en suceder es que toda la región por debajo del arquegonio se inflama, y las células diploides inician un proceso de especialización, el pie radicular se ubica hacia el cuerpo del gametofito, mientras que el primordio forliar emergerá por el vértice de la estructura en forma de corazón. Este crecimiento inicialmente procede igual que en varias briofitas ancestrales y algunas algas verdes, pues el esporofito crece de manera parasítica al gametofito.

Figura 6.8. Desarrollo inicial del esporofito de los helechos. El embrión al interior del gametofito es inicialmente parasítico, pero pronto inicia su crecimiento, hasta literalmente aplastar a su gametofito.
El esporofito no dura como un parásito por mucho tiempo, rápidamente desarrolla su propio sistema radicular y sus propias estructuras fotosíntéticas, además, al poseer sistema vascular, puede crecer de manera más marcada que el gametofito que morirá rápidamente.
Esporofito maduro
Esporofito maduro, es el cuerpo que siempre vemos de forma macroscópica de un helecho, la especialización celular para sobrevivir en el ambiente en ambientes secos y llenos de parásitos y depredadores es más marcada, pero aun necesitan agua para poder reproducirse. En el esporofito las hojas se denominan frondes y poseen una organización fractalizada, mientras que los tallos se denominan raquis. Las frondes, que son todos macrófilos, se van desarrollando a medida que crecen (crecimiento circinado), y que pequeñas tienen una forma característica de báculo. Tienen abundante nerviación y suelen estar divididas en pinnas una vez (unipinnados), dos (bipinnados), tres (tripinnados) o incluso más en otros segmentos más pequeños llamados pínnulas. El raquis es el nervio medio de estas frondes divididas.

Figura 6.9. Cibotium glaucum: Conocido como helecho de árbol hawaiano, es endémico de Hawái y puede crecer hasta una altura de aproximadamente 5 metros. Tiene un tronco grueso y frondas grandes que se despliegan en forma de abanico. Esta especie es apreciada por su aspecto tropical y su capacidad para crear un ambiente exuberante en los bosques hawaianos.
Estas hojas o frondes tienen una clara función fotosintética o alimenticia por su color verde, pero también tienen la función de formar y proteger las esporas, por lo que son además de trofofilos (hojas alimentadoras), esporofilos (hojas productoras de esporas), siendo pues troposporófilos. Existen casos en los que estas funciones están separadas en hojas diferentes.

Figura 6.10. El esporofito maduro de los helechos. El esporofito crece tanto que aplasta al gametofito, siendo la generación dominante de los helechos.
Esporangios
Los esporangios suelen aparecer agrupados formando soros en el envés de las frondes aunque a veces se encuentran solitarios o soldados formando los llamados sinangios. Tienen una protección, llamada indusio, cuando aparecen en este lugar, y que no es más que una excrecencia de la fronde; cuando los soros son terminales existe un pseudoindusio, que es el margen de la fronde doblado protegiendo el soro.


Figura 6.11. Esporangios y soros de los helechos. (Arriba) Los esporangios se ubican al interior de racimos llamados soros, que son visibles en el envés de las hojas de los helechos maduros; (abajo) Vistas de las esporas y los esporangios en un sus respectivos racimos o soros.
Los esporangios pueden ser de dos tipos en función, principalmente, de su tamaño: (1) Eusporangios: son grandes, sésiles o con un pedúnculo macizo. Albergan en su interior gran número de esporas y su pared es gruesa y está formada por varios estratos al menos durante las primeras etapas de la formación de las mismas. (2) Leptosporangios: más pequeños que los anteriores, tienen un pedúnculo delgado y el número de esporas que tienen es menor y múltiplo de 2. Su pared es más pequeña, formada por un sólo estrato de células, y por lo general tienen un mecanismo de dehiscencia o apertura característico que está formado por unas células más engrosadas que se llaman anillo mecánico. Cuando las esporas están maduras, las células del anillo se secan, provocando una brusca contracción de las células que forman dicho anillo, dispuestas por lo general a lo largo del mismo, haciendo que se rompa la pared del esporangio, liberando de esta forma las esporas al exterior con relativa potencia.
En resumen, los helechos son plantas con adaptaciones más marcadas al ambiente terrestre, como un sistema vascular que les permite crecer y un sistema de sostenimiento que mantiene a la planta en pie. Sin embargo, la adaptación de los helechos al ambiente terrestre no es completa, aun necesitan agua para que el gametofito pueda crecer adecuadamente y que este sumergido pueda liberar a los espermatozoides que nadan por el ambiente acuoso desde el anteridio al arquegonio (4). Estas plantas se desarrollaron mucho en el periodo geológico llamado carbonífero contribuyendo a las altas concentraciones de presión parcial de oxígeno durante este período de tiempo.
Reproducción asexual de los helechos
Los helechos se reproducen asexualmente por medio de la fragmentación de alguna estructura de sus cuerpos, pero la que con mayor éxito logra el truco es el rizoma. Una vez que un rizoma es separado de una planta madre, este prosigue su crecimiento para formar un esporofito nuevo. El tallo de los esporofitos de los helechos produce bulbos o crecimientos que pueden desprenderse con gran facilidad y permiten la propagación del individuo de forma asexual. Los bulbos también le permiten a la planta volver a crecer en caso de que las secciones superiores del bulbo mueran.
Una última forma de reproducción asexual es llevada a cabo in vitro, y es el cultivo de tejido del esporofito en un medio de cultivo con fines de investigación o industriales y es denominada propagación.
Ciclo de vida de los helechos
El esporofito maduro (1) es la planta que denominamos propiamente helecho, la cual posee hojas o frondes que poseen función fotosintética en el haz de la hoja y funciones reproductivas en su envés, donde genera estructuras reproductivas denominados esporangios (2), los cuales se encuentran agrupados densamente en paquetes coloreados y macroscópicos denominados soros (3). Al interior de los esporangios se realiza la meiosis (4) para producir esporas (5), las cuales deben caer en ambientes húmedos para producir el gametofito hermafrodita (6). El mismo gametofito produce anteridos (7) donde prosigue la espermatogénesis para generar los espermas (8). Tambie´n se producen en el mismo gametofito los aquegonios (9) que retienen un óvulo. En presencia de agua, los espermas son liberados, y nadan hacia los arquegonios para realizar la fecundación (10).

Figura 6.12. Ciclo de vida de los helechos (YouTube).
Solo hay un cigoto (11) ganador que bloquea a los demás químicamente y por demanda de nutrientes, el cual crecerá parasíticamente al gametofito (12) hasta que desarrolla su propia raíz y sus propias hojas fotosintéticas, igualmente el gametofito muere después de dos semanas de vida. El esporofito a su vez puede reproducirse asexualmente fragmentando (13) su sistema radicular, pues varios frondes pueden desarrollarse a partir de un mismo sistema de radículas.
7. Gimnospermas
7. Gimnospermas
Las gimnospermas se cuentan como el linaje de plantas con semillas y sus parientes. Evolucionaron en la era geológica más seca de todos los tiempos, el pérmico, por lo que desarrollaron una serie de adaptaciones que las hacen independientes del agua lo más posible. Dos adaptaciones hacen de las gimnospermas muy independientes del agua en términos de sus necesidades reproductivas. La primera es el polen que se dispersa por medio del aire y no del agua, y la segunda es la semilla que protege a la joven planta de la desecación (Biswas & Johri, 2013; Brenner, Stevenson, & Twigg, 2003; W G Chaloner, Hill, & Lacey, 1977; William G Chaloner & Pettitt, 1987; De Duve & Pizano, 1995; DiMichele & Bateman, 1996; Lee et al., 2011; Leslie, Beaulieu, & Mathews, 2017; Linkies et al., 2010; Rueda, Godoy, & Hawkins, 2017; Wang & Ran, 2014).
Los bosques del carbonífero de helechos fueron barridos en su mayoría por la desertificación del continente Pangea durante el pérmico, lo cual contribuyó a la diversificación de plantas que no necesitan agua para reproducirse como las gimnospermas. Las primeras gimnospermas eran semejantes a la araucaria, pero en términos taxonómicos se encuentran cercanamente emparentadas con el Ginko biloba. Por lo demás en las gimnospermas se marca la tendencia, el esporofito es enorme y lo llamamos árbol, mientras que el gametofito es ya microscópico y se encuentra escondido, rodeado y protegido por el esporofito. De hecho, el único gametofito que pasa algo de su vida fuera del esporofito es el gametofito masculino, vulgarmente conocido como el polen.

Figura 7.1. El Ginkgo biloba es un árbol único y antiguo que se cree que existe desde hace más de 270 millones de años. Es ampliamente conocido por su distintivo follaje en forma de abanico y sus propiedades medicinales. Este árbol caducifolio puede alcanzar alturas de hasta 30 metros y tiene una corteza grisácea y rugosa. Sus hojas, en forma de abanico, son de color verde brillante en verano y se vuelven amarillas en otoño antes de caer. El Ginkgo biloba es apreciado por sus beneficios para la salud, ya que se utiliza en la medicina tradicional para mejorar la memoria, la circulación y la salud cerebral. Además, es considerado un fósil viviente, ya que es el único miembro sobreviviente de la división Ginkgophyta.
Los descendientes vivos de este segundo ejército incluyen los helechos de semilla, las cicas que son palmas similares a árboles, los ginkos y especialmente, los pinos, cipreses, abetos y otras coníferas. Juntos, estos grupos forman la superfamilia de las gimnospermas (del griego gymnos, desnudo, y sperma, semilla). En la actualidad, sus semillas distan mucho de ser desnudas; se describen como tales a diferencia de las angiospermas (del griego aggeion, envoltura), cuyas semillas se encuentran contenidas dentro de frutos.
Generación esporofítica
La generación esporofítica en las gimnospermas es un aspecto esencial de su ciclo de vida. Estas plantas, que incluyen coníferas, cicas, ginkgos y gnetofitas, exhiben una alternancia de generaciones entre el esporofito y el gametofito. En este artículo, exploraremos en profundidad la fase esporofítica de las gimnospermas, centrándonos en su estructura, desarrollo y función en la reproducción. Analizaremos cómo los esporofitos producen esporas a través de estructuras como conos o estróbilos y cómo estas esporas dan lugar a gametofitos. También discutiremos la diversidad de formas y adaptaciones de los esporofitos en las diferentes divisiones de las gimnospermas. Comprender la generación esporofítica en las gimnospermas es fundamental para apreciar su éxito evolutivo y su importancia en los ecosistemas terrestres.
La semilla
La semilla corresponde al carácter más importante de las gimnospermas, ya que continúa con las adaptaciones contra la ausencia de agua. A diferencia de las esporas, las semillas poseen una protección mecánica contra depredadores y contra la desecación, además poseen nutrientes almacenados con lo que pueden resistir meses o años hasta que las condiciones mejoren. La formación de la semilla continua el proceso de formación del embrión, cuando uno de los embriones logra la victoria sobre sus hermanos “selección natural”, la membrana del megaesporangio se fortalece para formar la concha de la semilla. Las semillas poseen una variedad de formas, algunas poseen membranas que facilitan su dispersión por el aire, otras pueden ser liberadas por herbívoros como las ardillas que abren los conos.

Figura 7.2. La semilla. Las semillas son estructuras de resistencia ante la falta de agua prolongada, evolucionaron en el periodo geológico conocido como pérmico, donde los continentes formaron uno solo Pangea, y la mayoría de ese continente estuvo dominada por desiertos de diversos grados.
Las semillas son estructuras de resistencia con una buena cantidad de nutrientes que facilitan su desarrollo y extienden su periodo de dormancia. También son muy resistentes a los herbívoros y sus sistemas digestivos. Dado que las semillas evolucionaron para soportar los periodos secos del pérmico, lo que activa el desarrollo de la semilla es la presencia de agua. Una vez que la semilla germina, el nuevo esporofito crece hasta poder reiniciar el ciclo. El nombre de gimnospermas significa semilla desnuda, esto se debe a que la escama del estrobilo femenino que lo protegía se rompe al poco tiempo, liberando las semillas.
Esporofito
Los esporofitos dominan de manera tan absoluta a los gametofitos, que en realidad solo vemos una planta, el esporofito. A diferencia de otras plantas que vimos anteriormente, el gametofito jamás podrá ser encontrado en el suelo, la relación parasítica entre generaciones se invierte, ahora es el gametofito que crece como parásito del esprofito, pero de manera absoluta. El esporofito provee al gametofito interiorizado de todos los nutrientes que necesita, a tal punto que podríamos decir que el gametofito, especialmente el femenino es un mero órgano especializado del esporofito.

Figura 7.3. El esporofito de la gimnosperma. El esporofito de las gimnospermas es una planta leñosa de gran porte que llamamos genéricamente “árbol”. Habitan regiones frías latitudinales y montañosas.
Los esporofitos de las gimnospermas son muy reconocibles, pues se trata de árboles enormes, de expectativas de vida colosales. Las gimnospermas vivientes consisten en las cícadas, el ginkgo y un clado de coníferas y gnétidas. Estas plantas crecen en todo el mundo, de los 72 grados norte a los 55 grados sur, y son la vegetación dominante en muchas regiones frías y árticas. Muchas son familiares como ornamentales para madera de alta calidad. Entre las gimnospermas se encuentran los individuos más altos y de más larga vida de las plantas. El éxito evolutivo de las angiospermas en relación a los otros grupos de espermatofitas puede ser atribuido a los vasos en el xilema y a sus atributos para la reproducción. Todas las gimnospermas excepto los gnétidas tienen traqueidas en su xilema. Los vasos son más eficientes para el transporte de agua bajo algunas circunstancias, por lo que son más eficientes que otros linajes en ambientes secos y fríos con baja disponibilidad de agua.
Estas adaptaciones son tan poderosas, que han permitido la evolución de plantas enormes, como las colosales secoyas que pueden alcanzar más de 100 metros de altura, todo a un sistema vascular mejorado con respecto a los helechos y otras plantas con caracteres ancestrales, que dependen fuertemente de un ambiente húmedo. Sin embargo, para crecer no solo se necesita un bien sistema vascular, también el desarrollo especializado de una estructura de soporte, la lignina que da lugar a la madera, los esporofitos de las gimnospermas son conocidos por sus maderas de alta calidad.
La piña
Vulgarmente conocidas como piñas, y si, en nuestro idioma se confunde mucho con el fruto del género Anana de las angiospermas, pero es una estructura completamente diferente, conforman la principal estructura reproductiva de las gimnospermas, también se las llama comúnmente conos. En realidad, se las denominas estróbilos, y se dividen en dos dependiendo del sexo, para el masculino tenemos el microestróbilo y para el femenino tenemos el megaestróbilo. Los estróbilos comparten como característica común el estar hechos por escamas fuertemente ligninizadas, lo cual los hace muy duros, lo cual ayuda a proteger las estructuras reproductivas. El estróbilo hace parte del esporofito diploide, en otras palabras, no es el verdadero gametofito, el cual se encuentra escondido en su interior.

Figura 7.4. Los conos o estróbilos. Hay dos clases de piñas, la grande o femenina llamada megaestróbilo y la pequeña o masculina llamada microsestróbilo.
(a) Microestróbilo. El estróbilo masculino consiste en escamas membranosas o de textura acartonada organizados en una estructura espiral alrededor de un eje central. Estos conos comienzan a ser producidos durante la primavera. Cada una de las escamas del estróbilo en su base posee un par de microsporangios, que tampoco son el gametofito masculino. La función del microestróbilo es proteger los microesporángios encargados a su vez de producir el polen.

Figura 7.5. El cono masculino. La piña pequeña es la que produce el molesto polen de primavera que dispara las alergias de las regiones templadas del mundo.
Si hablamos de unidades reproductivas, esas deberían ser las escamas. Cada una de las escamas del microstróbilo protege en su interior al microsperorangio encargado de la producción del polen, y a su vez, las escamas se organizan geométroicamente en una espiral de Fibonacci alrededor del eje del cono. Vistas al microscopio, las escamas masculinas pueden ser descritas más como bolsas que como escamas, las cuales contienen muchísimas esporas.
(b) Megaestróbilo. Las megasporas que producirán el megasponrangio se encuentran localizadas en la base de las escamas de los conos formadores de semilla, también conocidos como estróbilos femeninos.

Figura 7.6. El cono femenino. El polen de desplaza por el viento al azar para ver si algún grano se filtra en las hendiduras de la piña grande, donde están las estructuras femeninas.
Los estróbilos femeninos son mucho más grandes que los masculinos debido a que deben acumular una mayor biomasa para la producción de las semillas, por esta razón también reciben el nombre de megastróbilos. Su desarrollo es complejo, pues requieren ser polinizados para seguir creciendo. Los estróbilos femeninos son también más duros, en contraste con la textura acartonada del masculino, las escamas de los conos femeninos son muy duras, con una textura de madera.

Figura 7.7. La escama. A diferencia de las microsporas, las megasporas se producen en cantidades limitadas al interior de los arquegonios de cada una de las escamas, esta es una microfotografia de varias escamas de un megastróbilo.
La unidad reproductiva vuelve a ser la escama, en cuyo interior se almacena el megasporangio, donde se encuentran las células reproductivas de la planta femenina. Cada escama es por lo tanto un protector de megasporangio y una semilla en potencia.
Generación gametofítica
La generación gametofítica en las gimnospermas desempeña un papel fundamental en su ciclo de vida. Estas plantas, que incluyen coníferas, cicas, ginkgos y gnetofitas, muestran una alternancia de generaciones entre el esporofito y el gametofito. En este artículo, exploraremos en detalle la fase gametofítica de las gimnospermas, centrándonos en su estructura, desarrollo y función en la reproducción. Investigaremos cómo los gametofitos producen células sexuales masculinas y femeninas y cómo se lleva a cabo la fertilización para dar lugar a la formación de la semilla. También discutiremos la diversidad de formas y adaptaciones de los gametofitos en las diferentes divisiones de las gimnospermas. Comprender la generación gametofítica en las gimnospermas nos permitirá apreciar la complejidad de su reproducción y su importancia en la persistencia de estas fascinantes plantas a lo largo del tiempo.
Esporas
Ocultos al interior de cada escama, son los encargados de producir las células reproductivas, aunque tampoco son el gametofito. Dado que cada sexo produce esporas de distinta calidad los esporangios se clasifican como microesporángios y megaesporángios, según producen mircsoesporas masculinas o megaesporas femeninas, unas de menor calidad, pero más numerosas, y las segundas de mayor calidad. pero en menor número.


Figura 7.8. Esporas y polen. (Arriba) El polen es una espora masculina, destinada a literalmente “germinar” si encuentra un cono femenino, en su interior transporta los espermas; (Abajo) Dado que el polen de las gimnospermas es transportado principalmente al azar, los machos producen enormes cantidades de polen para asegurarse de que algunos lleguen al blanco femenino, mientras que lo demás se desperdicia.
(a) La microespora o grano de polen. Al interior del microsporangio, las células madre llamadas microsporocitos llevan a cabo la meiosis masculina produciendo cuatro células haploides llamadas microsporas. Las microsporas no son homólogas a los espermatozides en los animales, ya que estas no son liberadas en la estructura femenina.
En su lugar, la microspora por mitosis se desarrolla en un grano de polen “los cuatro productos meióticos en un solo grano”, el cual en sí mismo es un gametofito masculino microscópico inmaduro. Cada grano de polen consiste en un par de sacos aéreos, los cuales incrementan el rozamiento del polen contra el aire y les permite flotar con el viento. Debido a la naturaleza fundamentalmente azarosa con la que el polen debe alcanzar la estructura femenina, los microsporangios producen cantidades astronómicamente inmensas de polen. Por ejemplo, un solo nido de conos es capaz de producir más de un millón de granos de polen para desgracia de aquellos individuos alérgicos a él.
Un grano de polen es microscópico, no mayor que 50 micras, pero las gimnospermas producen enormes cantidades en la primavera generando nubes de polen que inducen a fuertes alergias. Después de unas semanas de producción incesante, los estróbilos masculinos caen secos al suelo.
(b) El megasporangio y la megaspora. En la base de cada escama femenina se encuentra el megasponrangio, que no es el gametofito femenino. Dentro de cada megasporangio se ubica el macrosporocito o célula madre. El megasporangio es mucho más complejo que su homólogo masculino o microsporangio y está compuesto por el macrosporocito y por un tejido de reserva llamada nucela o saco embrionario. El megasporangio se encuentra encerrado por una cubierta muy dura, que posteriormente será la concha de la semilla. Sin embargo, esa concha posee un orificio tubular llamado micrópilo, el cual crece hacia el eje apical del cono, y es básicamente la puerta de entrada del polen.

Figura 7.9. Estructura interna de una escama de una piña femenina.
El megasporangio es una estructura dinámica, por ejemplo, la nucela es reemplazada por el gametofito que surge de la proliferación de la megaspora. Resulta interesante que es la megaspora el lugar donde tanto el gametofito masculino y femenino proliferan a sus estados multicelulares para hacer entrar en contacto sus células germinativas.
Los gametofitos y la polinización
Hasta el momento todo lo qeu hemos descrito se produce con células del esporofito, excepto claro, las esporas respectivas. Sin embargo, ¿Cuál es la generación gametofítica en las gimnospermas?
(a) Formación del gametofito femenino. El macrosporocito lleva a cabo la meiosis al interior de la nucela y el macrosporangio, produciendo cuatro células llamadas megasporas. De forma homóloga a lo que sucede en los mamíferos, pronto tres de las megasporas se degeneran mientras que la restante absorbe sus nutrientes. La megaspora restante producirá entonces el gametofito femenino, que consiste en cientos de células. La nucela sirve como el tejido de reserva para la masa del gametofito femenino en crecimiento hasta que se desarrollan unos seis arquegonios, los cuales quedan abiertos hacia el micrópilo.
Cada arquegonio desarrolla un óvulo. Por lo general en las microfotografías de los arquegonios no se puede ver el micrópilo debido a que es muy difícil hacer un corte justo a la mitad de la estructura. El espacio ocupado por la nucela en el megasporangio es reemplazado por el gametofito a medida que crece, y a su vez ese gametofito se convierte en el tejido de reserva que mantendrá a la semilla viva durante su dormancia y primeros días de desarrollo.
(b) Polinización en las gimnospermas. Los conos o estróbilos femeninos tardan aproximadamente dos años en madurar desde una coloración rojiza → verde → castaña. Durante la primera primavera las escamas inmaduras se abren permitiendo que los granos de polen puedan hacer contacto con el eje apical de cada escama.
Para asegurar la retención del polen, la escama produce un material ceroso y muy pegajoso que crea burbujas de polen aglomerado. A medida que el fluido se evapora el polen es tragado al interior del micrópilo, mientras que la abertura del micrópilo se degenera hasta cerrarse. Este proceso es denominado polinización.
Después de la polinización, las escamas crecen juntas y se cierran, protegiendo el óvulo en desarrollo. De hecho, la polinización es el evento que gatilla la meiosis en el megasporangio. Más aun, la maduración del megaesporangio femenino hasta poseer arquegonios femeninos maduros no ocurre sino hasta un año después de la polinización.
(c) Formación del gametofito masculino en las gimnospermas. El gametofito inmaduro o grano de polen también madura durante el año posterior a la polinización. En primera instancia debe producir un tubo polínico que le dé acceso por el micrópilo. Al llegar al megasporangio debe digerir la nucela restante en dirección a algún arquegonio maduro que esté produciendo suficientes hormonas. De las cuatro células meióticas, dos se desarrollan para ingresar al arquegonio. Una de ellas es la célula generativa, la cual a su vez por mitosis genera dos células, una estéril y la otra la célula espermática vegetal. Este primer espermatozoide se vuelve a dividir produciendo dos espermas secundarios, que sirven como los verdaderos gametos masculinos.
El esperma no se le pone el “zoide” debido a que carece de flagelos, y se encuentran confinados en el interior del tubo polínico hasta que este irrumpe muy cerca del ovulo al interior del arquegonio. El gametofito masculino se considera maduro solo hasta haber alcanzado el arquegonio y conteniendo dos espermas. En resumen, es como si fuera una plantita que crece al interior del cono femenino.

Figura 7.10. Germinación del grano de polen. (Irriba) Grano de polen germinando; (Abajo) En resumen tenemos el siguiente semiciclo de vida del polen, (a, b y c) ocurren en el estróbilo masculino y corresponde a la meiosis de la microspora (a hasta b) y la formación del grano de polen (c). Este viaja por el viento hasta que por azar alcanza un estróbilo femenino de la misma especie, allí empieza su desarrollo (i hasta j) como gametofito masculino en maduración.
Este tipo de reproducción tiene la ventaja de estar aislado del agua. En otras plantas los gametofitos debían estar húmedos para liberar los espermatozoides que debían nadar hasta la estructura femenina. Aquí no, es la planta completa la que viaja en forma de espora hasta llegar a la estructura femenina, y aun allí los espermas son transportados por el gametofito masculino en crecimiento. Gracias a este mecanismo inmune a la disponibilidad de agua, las plantas con semillas pudieron colonizar regiones que eran imposibles para otros linajes más arcaicos.
Fecundación y desarrollo del embrión vegetal
La fecundación se da retardada, el tubo polínico no ingresa en el anteridio sino hasta 15 meses después de la polinización, por esta razón la fecundación y la polinización son procesos completamente independientes.
Uno de los espermas fecunda al óvulo, mientras que la esperma restante, junto con las células del arquegonio se degenera. Los espermas de otros granos de polen pueden fecundar perfectamente los demás arquegonios, y cada cigoto comienza a desarrollarse en un propio embrión. El proceso es similar a la formación de los gemelos fraternales e los animales, aunque en este caso generalmente solo uno de los embriones sobrevive. A pesar que posteriormente el embrión se puede dividir de forma similar al proceso que conlleva a la formación de los gemelos idénticos, solo un embrión en un nido de un megasporangio logra desarrollarse en la mayoría de los casos. Una vez que el óvulo ha sido fecundado rápidamente comienza la proliferación del embrión por clivaje. Al mismo tiempo el embrión se mueve a la punta apical del arquegonio y crece hasta que lo rompe.
Una vez allí empieza a alimentarse del gametofito femenino. La punta en crecimiento formará los cotiledones de la semilla, mientras que la punta que crece en el sentido del grano de polen será la que forme la raíz "radícula". Una vez ha crecido lo suficiente el embrión formando el eje de los cotiledones-radícula, la semilla ha madurado a su estado normal de dispersión.
Ciclo de vida de las gimnospermas
A diferencia de las plantas con sistemas reproductivos más antiguos, las gimnospermas poseen un gametofito muy pequeño. El masculino es llamado polen y es transportado desde su lugar de producción en el cono masculino hasta el femenino.
El esporofito (1) puede ser diocio o monoico, pero en cualquier caso, genera estructuras generadores de esporas independientes, el megaestróbilo femenino (2) y el microestrobilo masculino (3). Al interior de las escamas de los estróbilos ocurre la meiosis (4) para generar esporas.
Las esporas de las gimnospermas son anisospóricas, de forma tal que las microsporas (5) o granos de polen son liberadas al ambiente como siempre había sido en las plantas, pero la macrospora (6) permanece fija al interior de la escama del estróbilo femenino. Cuando las microsporas llegan a un estróbilo femenino y se filtran por las hendiduras de las escamas germina, regenerando (7) un gametofito masculino (8) muy pequeño llamado tubo polínico, el cual porta núcleos generativos haploides, no espermas. Por otra parte, el gametofito femenino (10) prolifera (9) al interior de la escama, generando un tejido accesorio muy simple y un par de arquegonios con sus correspondientes óvulos haploides.
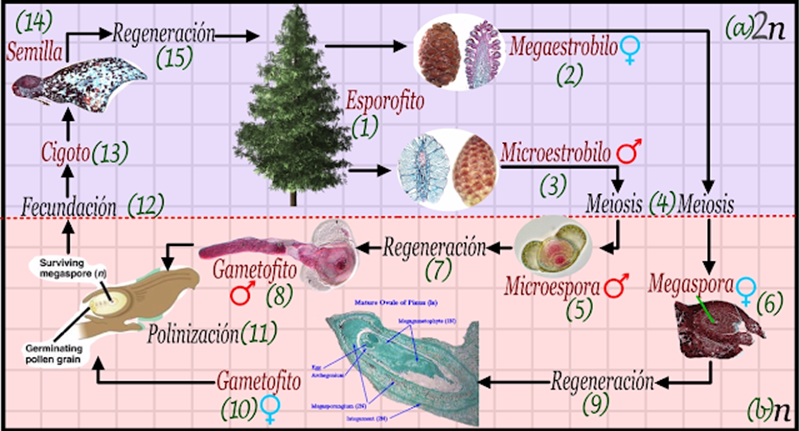
Figura 7.11. Ciclo de vida de las gimnospermas.
La llegada del polen a la escama se denomina polinización (11), pero solo hasta que ambos gametofitos se han desarrollado lo suficiente como para que los núcleos generativos del polen hacen contacto y realizan la cariogamia con los dos óvulos, es que se genera la fecundación (12). El cigoto (13) reemplaza al gametofito femenino, desplazándolo, y conformando el embrión, que incia una etapa de dormancia, mientras que el megasporangio conformará la membrana dura de la semilla (14). Cuando la semilla es liberada puede resistir condiciones adversas por meses o años hasta que germina y regenera (15) al esporofito (1).
8. Introducción a las Angiospermas
8. Introducción a las Angiospermas
Al igual que las gimnospermas, las angiospermas son heterospóricas, produciendo dos tipos de esporas, sin embargo, estas no son liberadas directamente, en su lugar se desarrollan en el interior del esporofito, ya sea para producir un arquegonio muy pequeño en la parte femenina de la flor, o produciendo los granos de polen. Sin embargo, a diferencia de los conos de las gimnospermas, las angiospermas poseen un órgano altamente especializado para la producción de semilla, y esta a su vez es producida encerrada en una superestructura llamada fruto.

Figura 8.1. La flor. La flor es una adaptación reproductiva, generada con el fin de manipular a otros seres vivos por medio de la simbiosis, de esta manera se generan vectores de transmisión de polen, evitando tener que producir tanto polen como las gimnospermas.

Figura 8.2. El pasto es una planta con flor, pero las flores están protegidas en estructuras llamadas espigas o inflorescencias. Estas se encuentran en la parte superior de los tallos y son pequeñas y poco vistosas. Aunque no llaman mucho la atención, son esenciales para la reproducción de la planta y la producción de semillas.
La fertilización de las angiospermas es uno de los procesos más complejos, ya que interviene un proceso llamado fertilización doble. Sin embargo, antes de hablar de ello, ahondaremos un poco más en la anatomía de la flor. De todas las plantas superiores, las plantas con flor o angiospermas son las que reciben una mayor atención por parte de los textos botánicos, esto se debe a dos razones principales, son extremadamente diversas y muchas de sus especies son de importancia civilizadora para nuestra sociedad moderna (Ames, 1963; Chang, 1987; Harshberger, 1896; Karling & Beuschlein, 1956; Mattu, 2017). Para no ir más lejos el arroz es una de las plantas de cultivo más importantes del mundo, también es un tipo de pasto y una angiosperma.
En esta serie de artículos realizaremos un breve análisis de la estructura, las partes y algunas modificaciones de las flores. También se realizará una breve descripción de los grupos principales de angiospermas basado en la estructura de la semilla. Posteriormente se ahondará en el desarrollo de la flor en los frutos, así como en su anatomía y fisiología generales. Finalmente se realiza una descripción de la estructura y la función de la semilla. Esta secuencialidad tiene que ver con el ciclo de vida de las plantas con flor, ya que la flor se convierte en el fruto que a su vez contiene las semillas, que en últimas son las estructuras de propagación y resistencia.
Diversidad de las platas con flor
El número de especies de plantas con flor varía entre 250000 a 400000 debido a que muchas aún no han sido catalogadas y también a la facilidad con que las nuevas especies pueden evolucionar en este linaje de seres vivos. En comparación con los musgos con 12000 especies conocidas, los helechos “y parientes” con 11000 especies, mientras que el número de especies de gimnospermas apenas sobrepasan las 100 especies vivas. Por mucho son las plantas más diversas a pesar de haber surgido recientemente en el registro fósil (Brock, 2014; Henry, 2005; Soltis & Soltis, 2014).

Figura 8.3. Wolffia columbiana es una especie de planta acuática flotante perteneciente a la familia de las lentejas de agua (Lemnaceae). Es considerada la planta con flores más pequeña del mundo. Sus frondas son extremadamente diminutas, midiendo tan solo alrededor de 0.5 a 1.5 milímetros de longitud. Presenta una forma ovalada y un color verde claro. Esta planta flota en la superficie de cuerpos de agua dulce, como lagos, estanques y arroyos. Al igual que otras especies de Wolffia, se reproduce rápidamente por división vegetativa, formando colonias densas. A pesar de su tamaño diminuto, Wolffia columbiana desempeña un papel importante en los ecosistemas acuáticos, proporcionando alimento y refugio para diversas especies de animales acuáticos.

Figura 8.4. Rafflesia arnoldii es una especie de planta parásita y sin hojas que se encuentra en las selvas tropicales de Sumatra y Borneo. Es conocida como la flor más grande del mundo debido a su gigantesco tamaño. Esta especie puede alcanzar un diámetro de hasta un metro y pesar hasta 11 kilogramos. Su aspecto distintivo incluye grandes pétalos rojos y una apertura central que revela una estructura interna con apariencia de carne y aspecto desagradable. Además, Rafflesia arnoldii emite un olor fuerte y fétido, similar al de la carne en descomposición, para atraer a los insectos polinizadores. La floración de Rafflesia arnoldii es un evento raro y espectacular, lo que la convierte en una atracción para los investigadores y los amantes de la naturaleza.
La mayoría de las especies aun no descritas se encuentran en los trópicos o en lugares de difícil acceso. Las flores mismas pueden variar entre miniaturas de 0.5-0.7 milímetros como en Wolffia columbiana (Figura 8.3), hasta flores enormes de hasta un metro y pesos de 9 kilos como en Rafflesia arnoldoni (Figura 8.4).

Figura 8.5. La especie de planta con la flor más grande de Colombia es la Victoria regia (Victoria amazonica). Esta planta acuática es conocida por sus enormes hojas flotantes y sus impresionantes flores. Las flores de la Victoria regia pueden alcanzar un diámetro de hasta 40 centímetros y tienen pétalos de color blanco o rosado. Sus flores desprenden un agradable aroma atraer polinizadores, como los escarabajos. Además de su belleza, la Victoria regia es admirada por su capacidad para soportar el peso de un humano en sus hojas flotantes. Esta especie es un tesoro natural de Colombia y representa la exuberante biodiversidad de los ecosistemas acuáticos del país.
El abominable misterio
A pesar de ser las más jóvenes en el registro fósil su amplia diversidad creó un desconcierto para los primeros darwinistas, pues el propio Charles Darwin describió su origen y evolución posterior como un “abominable misterio”, hoy tal misterio permanece en su origen último, mas no en su diversificación posterior (Cascales-Miñana, Cleal, & Gerrienne, 2016; Davies et al., 2004; Edwards, 2014; Friedman, 2009; Oliver, McCOMB, & Greene, 2014; D. E. Soltis, 2016).
Más allá de su origen último que como en la mayoría de los orígenes, se depende de la evidencia fósil, el abominable misterio hace referencia a todo el proceso evolutivo de las plantas con flores que aparenta un sistema discontinuo y a saltos. La razón para esto es que las flores de las plantas con flor poseen una mala tendencia, o buena según sea el punto de vista, que evitaban todas las plantas predecesoras, y es la autofertilización. Las flores hermafroditas de las plantas con flor tienden a la autofertilización, pero no autofertilizaciones cualquiera, sino a autofertilizaciones entre gametos no disyuntados, que provoca la formación de poliploides. Las semillas poliploides generan automáticamente una pequeña población nueva, y debido a las copias extras cromosomales, las primeras generaciones se ven protegidas ante la acumulación de mutaciones deletéreas por una compensación de dosis cuádruple. Adicionalmente los mecanismos de reparación genéticos provocan alteraciones secundarias, que generan la especiación instantánea (Buerki, Forest, & Alvarez, 2014; Cascales-Miñana, Cleal, & Gerrienne, 2016; Crepet, n.d.; Debate, 2012; Magadum, Banerjee, Murugan, Gangapur, & Ravikesavan, 2013; Moghe & Shiu, 2014, 2014; Oliver, McComb, & Greene, 2013; Roulin et al., 2013).

Figura 8.6. Si bien Charles Darwin fue uno de los científicos que estudió y discutió la "abominable misterio" de la evolución de las plantas con flor, no fue él quien acuñó el término. La expresión "abominable misterio" se atribuye al botánico inglés John Lindley, quien la utilizó en su obra "The Vegetable Kingdom" publicada en 1846. Lindley se refirió a la aparente rápida aparición y diversificación de las angiospermas en comparación con las plantas sin flores. En cuanto a la comprensión de la rápida evolución de las plantas con flor, varios científicos y botánicos contribuyeron a esta idea antes de Darwin. Algunos de ellos fueron Joseph Dalton Hooker, Ernst Haeckel y Thomas Huxley, quienes plantearon la noción de que las angiospermas experimentaron una radiación evolutiva acelerada. Sin embargo, Darwin amplió y profundizó esta idea en su obra "El origen de las especies", enfatizando la importancia de la selección natural en la evolución de las plantas con flor.
Esta especiación instantánea y cambios rápidos en las primeras generaciones aceleran el tempo evolutivo de las plantas con flor, provocando que evolucionen rápido y que se diversifiquen en periodos de tiempo geológico muy cortos en comparación con otras plantas, de allí el abominable misterio, para Darwin la evolución debía ser un proceso lento, dado que no conocía los mecanismos genéticos que permiten que el proceso evolutivo se dé, sin embargo las condiciones genéticas y anatómicas exclusivas de las plantas con flor han servido como un acelerador a marchas forzadas de su evolución y adaptabilidad.
Importancia de las plantas con flor para el hombre
Las plantas con flor son vitales para el ser humano, sin importar si su civilización es tribal o sedentaria. Muchas sociedades tribales dependen de frutas silvestres para completar sus dietas, y esos frutos son producidos por plantas con flor.

Figura 8.7. Seguridad alimentaria. Las plantas con flor, especialmente los pastos Poaceae son la base de la civilización moderna.
Las sociedades nómadas de pastores también dependen de las plantas con flor de forma indirecta, esto se debe a que los pastos son de hecho plantas con flores y frutos. El ganado se debe alimentar de estos pastos constantemente, y por eso deben movilizarse por enormes extensiones de terreno para encontrarlo, ejemplos de este tipo de sociedades son los mongoles, y algunas tribus aún sobreviven con este estilo de vida.
La importancia es mucho más capital en las sociedades sedentarias, de hecho, sin la domesticación de una serie de pastos la civilización sedentaria hubiera sido imposible. De hecho, la mayor parte de la humanidad depende de tan solo 11 especies de pastos de la familia Poaceae, los cuales suplen cerca del 80% de la biomasa alimentaria mundial. Algunos ejemplos son el arroz, el trigo, la cebada, el centeno, el maíz, la avena y el sorgo (Clayton & Renvoize, 1986; DeWet, 1981; Khush, 2016; Lapierre & Signoret, 2004; LU & Ellstrand, 2014; Stallknecht, Gilbertson, & Eckhoff, 1993; Watson, 1990).
Diversidad de los pastos comestibles
Los pastos comestibles desempeñan un papel fundamental en la alimentación humana desde tiempos ancestrales. Dentro del clado Poaceae, se han domesticado diversos miembros a lo largo de la historia, proporcionando una amplia gama de cultivos agrícolas de importancia económica y nutricional. En este artículo, exploraremos la diversidad de los pastos comestibles y su orden cronológico de domesticación.
A medida que examinamos la evolución y domesticación de los miembros del clado Poaceae, podremos apreciar la riqueza de especies que han sido cultivadas por las sociedades humanas en diferentes épocas y regiones geográficas. Desde los cereales fundamentales en la antigüedad, la diversidad de los pastos comestibles ha sido moldeada por la selección humana y ha evolucionado para adaptarse a una amplia gama de condiciones ambientales.
Acompáñanos en este recorrido por la historia de los pastos comestibles y descubriremos los hitos clave en su domesticación, explorando su importancia histórica, su valor nutricional y su impacto en la seguridad alimentaria global. Adentrémonos en el mundo de los pastos comestibles y su papel vital en la alimentación humana a lo largo del tiempo. Los miembros del clado Poaceae domesticados se pueden organizar en orden cronológico de la siguiente manera.
(a) El trigo (Triticum spp.), cuya domesticación tuvo lugar alrededor del 8000 a.C. en el Creciente Fértil de Oriente Medio ha sido un cultivo de gran importancia histórica, ya que desempeñó un papel crucial en el desarrollo de las primeras civilizaciones agrícolas, como las de Mesopotamia y Egipto. En la actualidad, el trigo sigue siendo uno de los cereales más importantes a nivel mundial, utilizado para la producción de harina y otros productos alimenticios.
(b) La cebada (Hordeum vulgare), domesticada alrededor del 7000 a.C. en el Creciente Fértil de Oriente Medio, ha sido un cultivo fundamental en la historia de la agricultura. Además de su uso para la alimentación humana y animal, la cebada tiene una amplia gama de aplicaciones, incluyendo la fabricación de cerveza y whisky. Es uno de los cereales más cultivados en todo el mundo y sigue siendo una fuente importante de alimento y materia prima en diversas industrias.
(c) La caña de azúcar (Saccharum officinarum), domesticada alrededor del 6000 a.C. en el sudeste asiático. La caña de azúcar ha tenido una gran importancia histórica como fuente de azúcar y ha influido en el comercio y la economía global durante siglos. En la actualidad, la caña de azúcar sigue siendo un cultivo de importancia económica significativa en muchas partes del mundo, siendo utilizada principalmente para la producción de azúcar y etanol.
(d) El arroz (Oryza sativa) fue domesticado alrededor del 5000 a.C. en Asia Oriental, específicamente en China y/o India. Históricamente, el arroz ha sido un cultivo fundamental para la alimentación humana en muchas culturas asiáticas, desempeñando un papel crucial en el desarrollo de la agricultura y el establecimiento de civilizaciones en estas regiones. En la actualidad, el arroz sigue siendo uno de los cultivos más importantes a nivel económico, siendo una fuente básica de alimento para una gran parte de la población mundial.
(e) Otro miembro destacado es el maíz (Zea mays), domesticado alrededor del 5000 a.C. en Mesoamérica, específicamente en México. El maíz ha sido un cultivo central en la historia de las civilizaciones precolombinas en América, como los mayas y los aztecas. Hoy en día, el maíz es uno de los cultivos más ampliamente cultivados y consumidos en todo el mundo, utilizado en una amplia variedad de alimentos, piensos y productos industriales.
(f) El centeno (Secale cereale), domesticado alrededor del 4000 a.C. en Europa, es un cereal utilizado principalmente para la alimentación animal y la producción de harina para panificación. Aunque su importancia económica moderna es menor en comparación con otros cereales, el centeno es apreciado en ciertas regiones por sus características de cultivo resistente y su adaptabilidad a suelos pobres.
(g) El sorgo (Sorghum bicolor), domesticado alrededor del 3000 a.C. en África, es un cultivo versátil que se utiliza tanto para la alimentación humana como para la alimentación animal. Es resistente a la sequía y se cultiva ampliamente en regiones áridas y semiáridas. Además, el sorgo tiene una importancia económica moderna debido a su uso en la producción de etanol y como materia prima para la industria del papel.
(h) Avena (Avena sativa): La avena fue domesticada alrededor del año 2000 a.C. en Europa. La avena es un cereal ampliamente cultivado en todo el mundo. Se utiliza principalmente para la alimentación animal y la producción de alimentos como avena para el desayuno, harinas y productos horneados.

Figura 8.8. Es válido decir que la civilización humana descansa sobre los pastos comestibles debido a su importancia fundamental en la alimentación y la agricultura. Los pastos comestibles, pertenecientes al clado Poaceae, han sido domesticados y cultivados durante miles de años, proporcionando una fuente primaria de alimentos para la humanidad. Desde los cereales básicos como el trigo, el arroz y el maíz, hasta pastos utilizados como forraje para el ganado, estos cultivos han sustentado comunidades, alimentado a civilizaciones enteras y han sido la base de sistemas agrícolas sostenibles o insostenibles. Los pastos comestibles son esenciales para la seguridad alimentaria, la economía y el desarrollo humano, lo que los convierte en la piedra angular de nuestra civilización.
(i) Mijo (Pennisetum glaucum): fue domesticado alrededor del año 2000 a.C. en África y Asia. El mijo es un grupo de cereales pequeños que se cultivan en muchas regiones del mundo, especialmente en África y Asia. Son una fuente importante de alimento básico para muchas comunidades y se utilizan en la alimentación humana y animal.
Importancia histórica
Estos miembros del clado Poaceae domesticados han dejado un impacto duradero en la historia de la humanidad, tanto desde una perspectiva histórica como económica, demostrando la importancia y la diversidad de este grupo de plantas en nuestra sociedad. Dos ejemplos históricos de imperios que se vieron afectados por una mala cosecha de plantas del clado Poaceae son:
(a) El Imperio Romano: Durante el siglo III d.C., el Imperio Romano sufrió una serie de malas cosechas que contribuyeron a su debilitamiento y eventual colapso. Las sequías y la falta de producción agrícola, incluyendo los cultivos de trigo (Triticum spp.), afectaron la estabilidad económica y social del imperio. Esto llevó a la escasez de alimentos, el aumento de los precios y la descontento generalizado entre la población, lo que a su vez generó conflictos internos y dificultades en la gobernabilidad del imperio.
(b) El Imperio Maya: En el período Clásico Tardío (aproximadamente del 600 al 900 d.C.), el Imperio Maya en Mesoamérica experimentó un declive significativo. Uno de los factores que contribuyeron a este declive fue la falta de éxito en la producción agrícola, incluyendo los cultivos de maíz (Zea mays). Las sequías prolongadas y las prácticas agrícolas insostenibles agotaron los recursos naturales y disminuyeron la capacidad de los mayas para sostener su población. Esto llevó a conflictos internos, migraciones y el eventual abandono de muchas ciudades mayas.
Estos ejemplos ilustran cómo la dependencia de los cultivos del clado Poaceae y los desafíos relacionados con la producción agrícola pueden tener consecuencias significativas en la estabilidad y sostenibilidad de los imperios en diferentes períodos históricos y regiones geográficas.
Color, forma, textura y olor, la variabilidad de la flor
Lo primero que se hace evidente es la estructura externa de la flor, así que vamos a ver un poco de su diversidad anatómica y funcional. La flor es una estructura con una variabilidad extrema. Sus colores pueden ser cualquiera de los del arcoíris y más, esto se debe a que los polinizadores pueden detectar radiaciones electromagnéticas de mayor amplitud y con mayor precisión que el ojo humano. Las flores pueden tener cualquier textura vegetal, desde una textura suave y transparente de láminas muy finas, hasta texturas de cuero acartonadas, o incluso más duras. Otras pueden ser aterciopeladas o incluso húmedas al tacto o pegajosas. Las flores de muchos árboles y pastos no son conspicuas y carecen de olores característicos, pero otras flores son maravillosamente bellas, particularmente cuan son examinadas con un microscopio. Los patrones de coloración cambian dependiendo del filtro de luz con que las miras, una flor puede presentar patrones completamente diferentes vistas con un filtro que permite ver el ultravioleta.

Figura 8.9. Belleza escondida. Dependiendo de cómo podamos ver la radiación electromagnética, una misma flor poseerá un diferente patrón de color.
Las fragancias de las flores también son inmensamente variables, algunas son extremadamente placenteras y otras huelen a carne en putrefacción. De hecho, muchas de estas fragancias son la base de las industrias de aromas y perfumes.
Flores raras
Las flores, con su diversidad de colores, formas y aromas, son una de las maravillas más hermosas que nos ofrece la naturaleza. Sin embargo, dentro de este mundo floral, existen especies que destacan por su rareza y singularidad. En este artículo, exploraremos el fascinante mundo de las flores raras, aquellas que sorprenden por sus características únicas y extraordinarias.
Desde flores con formas inusuales hasta colores vibrantes que desafían la norma, estas especies nos cautivan con su rareza y nos invitan a contemplar la asombrosa creatividad de la naturaleza. A medida que nos adentremos en este viaje, descubriremos ejemplos de flores raras de diferentes rincones del mundo, explorando sus adaptaciones, sus misteriosos procesos de polinización y el papel crucial que desempeñan en sus ecosistemas.
Prepárate para sumergirte en un mundo lleno de exotismo floral y descubrir las joyas ocultas que nos regala la naturaleza. Exploraremos cómo estas flores raras han fascinado y cautivado a científicos, amantes de la naturaleza y coleccionistas a lo largo de la historia. Acompáñanos en este apasionante recorrido por las flores más singulares y extraordinarias que adornan nuestro planeta.
Flores bajo tierra
Varios años atrás, un granjero australiano llamado Jack Trott mientras araba su plantación empezó a notar un patrón de lo más extraño en su campo, era como si existieran flores bajo la tierra y él no se hubiera dado cuenta. Cuando se bajó de su tractor y miró más de cerca se dio cuenta de que lo que veía no era meramente un patrón al azar, en efecto se trataba de una flor bajo tierra. Más aun, la planta de la cual germinaba era completamente pálida y no poseía ningún tipo de clorofila. El reportó su descubrimiento a una Universidad donde los botánicos determinaron que el granjero se topó con la primera especie de planta con flor que crece bajo tierra, y una de las pocas plantas con un modo de vida mico-heterótrofo, fue denominada Rhizanthella gardneri (Balogh, 1982; Dixon, 2003; Mursidawati, 2004).

Figura 8.10. Flores bajo tierra. Flor de Rhizanthella gardneri, planta parasítica que ataca a algunos hongos.
La planta misma era una orquídea que vive de parasitar a un hongo extrayéndole sus nutrientes por medio de una micorriza y es polinizada por pequeñas moscas que tenían acceso a sus flores por medio de las grietas que se desarrollan durante la temporada seca. Aunque las orquídeas parásitas no son raras, si lo son aquellas que son parásitos obligados sin clorofila, y peor aún aquellas que parasitan a los hongos.
Flores con olor a muerto
Una especie única denominada la flor cadáver Amorphophallus titanum (Gandawijaja, Idris, Nasution, Nyman, & Arditti, 1983)produce una inflorescencia que merece mención a parte debido a su tamaño y olor particularmente desagradable. La inflorescencia misma puede abarcar dos metros, la cual protege a un grupo de flores en su interior. El retoño emerge del suelo y puede permanecer oculto por décadas.
Los escarabajos carroñeros piensan que es un cadáver debido a su aroma semejante a la carne en putrefacción, por lo que presumiblemente son los principales polinizadores, al viajar de una flor a otra. La rareza y espectacularidad de la flor cadáver inspira a espectadores curiosos a visitar una de estas rarezas botánicas en los jardines botánicos alrededor del mundo. Esto se debe a que solo se han reportado poco más de 100 retoños de esta flor.

Figura 8.11. Flores con olor a muerto. El ecoturismo podría salvar a la flor más grande del mundo (YouTube).
Flores parásitas
Las plantas con flor habitan ambientes tan diversos como sus formas. Algunas plantas epifitas entablan relaciones simbióticas con otras plantas, dentro de todos los rangos, algunas como comensales y otras como parásitos (Joel, Gressel, & Musselman, 2015; Musselman, Press, Press, & Graves, 1995). Las orquídeas tienden a formar este tipo de relaciones, aunque sus flores pueden llegar a ser muy bellas.

Figura 8.12. Flores pálidas. Estas flores pertenecen a plantas sin clorofila parásitas de hongos-árboles micorrízicos. También cabe resaltar que las plantas con flor poseen metabolismos de una productividad superior como es el caso de las plantas CAM y C4. Adaptabilidad de las plantas con flor o angiospermas
También pueden sobrevivir tanto en agua dulce como en agua salada, pueden aferrarse a muchas superficies difíciles como las grietas de rocas y edificaciones, pueden sobrevivir en ambientes con baja disponibilidad de agua como en los desiertos cálidos o en los desiertos fríos de la tundra. Algunas pueden tolerar temperaturas de hasta 43°C y otras pueden soportar hasta -18°C. De hecho, pueden encontrarse especies de plantas con flor prácticamente en cualquier parte, incluso en ambientes sin luz. Sin embargo, debido a la estructura general de sus hojas, la mayoría de las especies se encuentran particularmente adaptadas a los trópicos.
Tiempo de desarrollo de una planta con flor
Las plantas con flor pueden ir desde la germinación de la semilla hasta la primera reproducción de la planta adulta en menos de un mes o en más de 150 años dependiendo de la especie. Las flores anuales generalmente viven en un ciclo anual, la planta nace, crece, se reproduce y muere en un año. Las plantas bianuales tardan dos años en completar sus ciclos de vida y morir.

Figura 8.13. Un árbol viejo. Las plantas con flor no viven mucho en comparación con otros linajes, pero el higo de Sri Lanka muestra el potencial de longevidad de un ser vivo.
Las plantas perennes toman varios años para desarrollarse y pueden reproducirse por numerosos ciclos anuales. De hecho, la planta con flor más vieja registrada fue plantada en el año de 288 después de Cristo y ha vivido por más de 2.300 años, se trata de un higo “Ficus religiosa” de Sri Lanka llamado Jaya Sri Maha Bodhi (Chandrakanth & Romm, 1991; Kaur, Rana, Tiwari, Sharma, & Kumar, 2011; Sharma, Rikhari, & Palni, 1999).
Clasificación de las plantas con flor
Tres grupos de plantas con flor han sido designados clásicamente, las acotiledóneas o magnolidas, las monocotiledóneas o magnoliopsidas y las dicotiledóneas o liliopsidas. La distribución de especies no es homogénea, cerca del 75% de las especies se clasifican como liliopsidas, el 23% como magnoliopsidas y menos del 2% se clasifican como magnolidas. Al interior de las 42 familias de plantas con flor más importantes, la mayoría pertenecen a las Asteracea o compuestas, también designables como la familia de los girasoles y parientes cercanos con 25 000 especies conocidas. La segunda familia más numerosa es la de las orquídeas con 28 000 especies conocidas. Las leguminosas poseen 19 4500 especies, y los pastos 12 000 especies.
9. La flor, anatomía y función
9. La flor, anatomía y función
La flor es la estructura reproductiva de las plantas que pertenecen al linaje de las angiospermas también denominadas Magnoliofitas. La función de la flor es la de permitir el proceso reproductivo mediante la provisión del espacio para la fertilización de la esperma y el óvulo. Sin embargo, la flor es mucho más, esta se desarrolla en flor y fruto. La reproducción de las angiospermas es un proceso altamente especializado y la mayoría de las veces requiere de entablar relaciones simbióticas con animales. Esto se ha llevado a tal punto que se ha propuesto que las abejas evolucionaron de un grupo de avispas que se especializó en obtener el néctar de las primeras flores.
Figura 9.1. La simbiosis mutualista. La flor evolucionó para solucionar el problema del polen, las gimnospermas producen mucho polen con pocas probabilidades de alcanzar a las partes femeninas, la flor se encarga de aumentar la probabilidad atrayendo polinizadores y haciéndolos específicos para cada especie de flor, disminuyendo así la necesidad de generar mucho polen.

Figura 9.2. Anatomía de la flor hermafrodita. Según la especie, las flores pueden ser hemafroditas o dioicas, exponiendo partes masculinas (estamen) y femeninas (Carpelo), o ser monóicas, y tener flores masculinas y femeninas especializadas.
Los polinizadores simbióticos son un mecanismo que reduce la aleatoriedad en la transmisión del polen de una flor a otra, lo cual disminuye la cantidad de polen que una flor planta debe producir para reproducirse ahorrando energía metabólica. Algunas plantas han sido tan extremas que han especializado polinizadores únicos para su especie, lo cual es un arma de doble filo, aunque el polinizador en este caso transmite el polen con 0 azar, si la especie polinizadora se extingue también lo hace la especie de la flor. Muchas flores son hermafroditas, por lo que facilita la autofertilización y la generación de autoploliploides y alopoliploides, lo cual facilita su especiación a gran velocidad. Esta capacidad de generar nuevas especies con facilidad es una de las partes del rompecabezas para solucionar ese problema misterioso del origen de las plantas con flor que Darwin denominó, un abominable misterio.
Las flores están compuestas en general por dos grupos de estructuras, las partes vegetativas y las partes reproductivas. Las partes vegetativas o perianto son aquellas que no están involucradas con la fertilización o la polinización, pero que apoyan el proceso haciéndolo más probable o simplemente protegiendo a las partes reproductivas. Las dos secciones más importantes del perianto son el cáliz y la corola. Las partes reproductivas de la flor son aquellas involucradas con la producción de los gametos reproductivos y con la fertilización, se dividen en dos de acuerdo a los sexos. La parte masculina que produce el polen se llama androceo “estambres” y la femenina “gineceo” “pistilo”.

Figura 9.3. Izquierda, flore sin pedicelo y derecha, flor pedicelada.
Partes vegetativas de una flor o perianto
y fascinante compuesta por diferentes partes. Entre estas partes se encuentran las denominadas partes vegetativas de la flor o perianto. En este artículo, nos sumergiremos en el estudio detallado de estas estructuras y descubriremos su importancia en la biología floral.
El perianto comprende las partes de la flor encargadas de proteger y atraer a los polinizadores, formando una especie de envoltura que rodea a las partes reproductivas. Estas partes pueden incluir los sépalos, que forman el cáliz, y los pétalos, que conforman la corola. A través de una exploración minuciosa, analizaremos la morfología, función y variabilidad de estas partes vegetativas en diferentes especies de plantas.
Además, abordaremos la diversidad de colores, texturas y formas que pueden presentar los sépalos y pétalos, así como su adaptación a diferentes estrategias de polinización. También examinaremos casos de modificaciones y adaptaciones especializadas en estas estructuras, que han permitido a las plantas conquistar diversos ambientes y atraer a sus polinizadores específicos.
Únete a nosotros en este emocionante viaje hacia el interior de la flor y descubre las maravillas y secretos que las partes vegetativas del perianto nos revelan sobre la biología y la evolución de las plantas con flores. Prepárate para adentrarte en un mundo de formas y colores sorprendentes, donde cada detalle desempeña un papel esencial en la perpetuación de la vida vegetal.
El eje de la flor
En el maravilloso mundo de las flores, el eje que da origen al brote de la flor juega un papel fundamental. Aunque no se considera una parte formal del perianto, merece su reconocimiento debido a su importancia en la arquitectura floral. En esta sección, exploraremos en detalle el eje de la flor y su contribución esencial en el desarrollo y organización de las estructuras florales.
El eje de la flor puede manifestarse de diferentes maneras según su función y ubicación. Si el tallo que da origen a la flor es el principal y no está específicamente dedicado a la producción floral, se conoce como pedúnculo. Por otro lado, si el pedúnculo no genera una flor directamente, sino un tallo de menor envergadura del cual emerge la flor, se le denomina pedicelo. Ambas variantes del eje de la flor desempeñan un papel crucial en la presentación y sostén de la flor.
El eje central de todas las estructuras florales se conoce como receptáculo. Este receptáculo es un punto focal en el cual convergen la corola, el cáliz y las estructuras reproductivas. Actuando como un eje de simetría radial, el receptáculo proporciona la base sobre la cual se organizan las partes de la flor, asegurando una disposición armónica y equilibrada.
A través de la comprensión y estudio detallado del eje de la flor, podemos apreciar su contribución esencial en la morfología y función floral. Su presencia permite una distribución adecuada de las estructuras reproductivas y un soporte estructural para las partes vegetativas de la flor. Sin duda, el eje de la flor es un elemento vital en la arquitectura floral y merece nuestra atención al explorar la diversidad y complejidad de las flores en el reino vegetal..
El primer círculo, protector
Justo a partir del receptáculo, emerge el primer círculo de hojas especializadas que desempeñan un papel protector en la flor. En la mayoría de las plantas, estas hojas, que pueden llegar a ser fotosintéticas y bastante resistentes, se conocen como sépalos. Los sépalos, que suelen ser de tres a cinco en número, rodean completamente el primordio de la flor cuando aún se encuentra en su etapa de botón en formación.

Figura 9.4. Sépalos. Los sépalos son las hojas verdes que sostienen la flor.
En ciertas plantas, como los pastos, donde las flores no son fácilmente perceptibles, los sépalos se presentan en forma de dos hojas grandes y completamente funcionales conocidas como lemas (la más grande) y palea (la más pequeña). Juntas, forman un estuche que envuelve a la flor y la protege. El conjunto de sépalos se conoce como cáliz y constituye la primera sección formal del perianto, la envoltura floral.

Figura 9.5. El cáliz. En este racimo podemos ver varias flores con diferentes estados de desarrollo, al inicio cuando la flor es muy joven los sépalos cubren todo el primordio de la flor en desarrollo, la cual al crecer emerge por la región de la punta apical del cáliz abriéndolo paulatinamente.
El cáliz desempeña un papel crucial al brindar protección a las delicadas estructuras reproductivas en desarrollo dentro del capullo floral. Además de su función protectora, los sépalos también pueden presentar características ornamentales, contribuyendo a la belleza y diversidad de las flores. A medida que exploramos la maravillosa diversidad de los patrones y formas de los sépalos y el cáliz, nos adentramos en un fascinante mundo de adaptaciones evolutivas y funciones vitales en el reino vegetal.
El segundo círculo, guía
La siguiente sección del perianto está compuesta por los pétalos, que son hojas mucho más especializadas que los pétalos. A la colectividad de pétalos se los denominan corola.
La corola es la parte más bella de la flor y lo que nos hace llamarla flor, pero no hay que olvidar que es una parte de la flor, es más, existen flores sin corolas y pétalos. Las corolas poseen pigmentaciones de todos los colores posibles, más allá incluso de los que puede percibir el ojo humano, y su función es la de servir como faro para los polinizadores, especialmente las abejas y otros artrópodos capaces de percibir rangos más amplios y detallados del espectro electromagnético. En otras palabras, la mayoría de las flores guardan más colores que no podemos ver, más patrones que los que podemos percibir, y más belleza de la que podemos notar.
Los patrones de coloración de las flores cambian dependiendo del tipo de receptor que posea un ojo, por ejemplo, muchos insectos polinizadores ven en ultravioleta "lo cual genera un patrón diferente en la foto de la derecha", ahora no es que vean en morado, los filtros UV para fotos transforman el ultravioleta invisible en un color visible para nuestros ojos.

Figura 9.6. La corola. La corola es la parte llamativa de las flores, y es altamente variable, aun en plantas cercanamente emparentadas, los colores de los pétalos no solo involucran el espectro visible, sino también infrarojos, ultravioletas, e intraespectros que nuestros ojos no son capaces de percibir.
La flor no visible de los pastos
La colora en ocasiones puede no ser conspicua o ni siquiera estar presenten como ocurre en muchos árboles o en los pastos “Poaceae”. Estas especies dependen del viento para su polinización. Lo que nos lleva a un detalle interesante.

Figura 9.7. La flor del pasto.
Muchas personas, incluso estudiantes de primeros semestres de biología no consideran a los pastos como angiospermas debido a que no pueden ver sus flores. La razón de esto es que el perianto de los pastos está muy modificado (Figura 9.7 A-B). En los pastos la estructura reproductora o espiga es una infloresencia (Figura 9.7 A), cada floreta está compuesta por un cáliz de dos sépalos llamados lema “la más grande” y palea “la más pequeña” (Figura 9.7 B) los cuales encierran los órganos sexuales como si fueran una caja (Figura 9.7 D). Estos sépalos también fungen como las hojas fotosintéticas principales de los pastos. Adicionalmente debido a que el cáliz encierra toda la flor no se requiere de corola, y en consecuencia estas flores no tienen pétalos coloridos. El asunto es que la mayoría asocia corola = flor, pero esta asociación es falsa, los pastos poseen flores, pero no corolas.
Partes reproductivas de una flor
Las flores son estructuras fascinantes y bellas que desempeñan un papel fundamental en la reproducción de las plantas con flores. Son el escenario donde se lleva a cabo la compleja danza de la fertilización y la formación de nuevas generaciones. Para comprender a fondo este proceso vital, es esencial familiarizarse con las partes reproductivas de una flor y su función en el ciclo de vida de las plantas.
En este artículo, exploraremos detalladamente las diferentes partes reproductivas de una flor y su significado en el contexto de la reproducción sexual de las plantas. Desde los órganos masculinos hasta los órganos femeninos, descubriremos cómo cada componente desempeña un papel único y esencial en el proceso de polinización y fecundación.
Sumérgete en el mundo intrincado de los estambres, pistilos, estigmas, anteras y otros componentes reproductivos de las flores. A través de su estudio, podremos apreciar la belleza y la complejidad de la reproducción en las plantas, así como la increíble diversidad de estrategias y adaptaciones que han evolucionado a lo largo del tiempo. ¡Acompáñanos en este viaje a través de las partes reproductivas de una flor y desvela los secretos de la perpetuación de la vida vegetal!
Androceo, estambre y antera
Las partes vegetativas de la flor sirven como estructura de apoyo y protección a las zonas reproductivas. Las flores pueden ser monoicas “generar un solo tipo de estructura sexual” o dioicas “generan ambos tipos de estructura sexual”. Por lo general los diagramas de flores ideales se realizan con una flor de tipo dioico. A las partes masculinas se las denomina androceo y a las femeninas gineceo. El androceo tiene como función principal la producción del gametofito masculino o polen. El androceo está compuesto por el estambre y la antera. El estambre son filamentos delgados que se encuentran unidos a la base o receptáculo y parece que emergieron del pistilo que conforma el eje de la flor. Los estambres pueden ser semirrígidos o muy flexibles y en su punta portan un saco especializado llamado antera.

Figura 9.8. Flor masculina. El androceo compuesto por estambre y antera. Es en la antera donde se producen los granos de polen. En la mayoría de las flores, el polen es liberado a lo largo de aberturas o poros microscópicos de la antera.
Gineceo o pistilo
El pistilo o eje central de la flor generalmente tiene forma de vaso cerrado en la punta y consiste en tres regiones que convergen mutuamente llamadas estigma, estilo y ovario. De igual forma un pistilo está formado por uno o más carpelos.

Figura 9.9. Flor femenina. El gineceo también denominado pistilo es la parte central de la flor femenina o hermafrodita y se divide en estigma, estilo y ovario.
El estigma se encuentra conectado a una estructura delgada y con forma de aguja llamada estilo, el cual se conecta a su vez con una base vacía llamada ovario. El estigma puede ser tan pequeño como un punto, un pequeño abultamiento o puede consistir en muchas prolongaciones o secciones ramificadas. El ovario es la estructura que se convierte en el fruto. Existe evidencia de que los primeros ovarios se desarrollaron cuando las márgenes de las hojas que portaban los óvulos se enrollaron hacia el interior de la estructura reproductiva. Tales hojas portadoras de ovarios fueron llamadas carpelos.
En algunos casos dos o más carpelos se fusionaron y muchos ovarios son ahora compuestos, que consistente en muchos carpelos unidos (B). El número de carpelos presentes es posteriormente evidenciado por el número de lóbulos o divisiones internas del estigma. Cada ovario maduro como el tomate o la naranja representa un carpelo.
El eje de la fruta es donde estuvo ubicado el pistilo y cada una de las secciones es un carpelo con sus propios ovarios y semillas. El ovario es una parte del pistilo que contiene el óvulo vegetal y puede localizarse arriba o detrás del punto de conexión con la base de los pétalos y sépalos llamado receptáculo. El ovario se denomina superior o súpero si se ubica por encima del receptáculo e inferior o ínfero cuando se encuentra por detrás del receptáculo, en caso de estar en medio se lo llama semiínfero.
Ejemplos de ovarios súperos son presentados por las flores de la alverja y la primavera. Ejemplo de ovarios ínferos son presentados por las flores del cactus y la zanahoria. Una cavidad que contiene uno o más carpelos que poseen a su vez uno o más óvulos; los óvulos se encuentran unidos a una pared de la cavidad por medio de unos tallos cortos. Cuando un óvulo se desarrolla después de la fertilización se transforma en semilla, sin embargo, el proceso es mucho más complejo que en las gimnospermas.

Figura 9.10. Partes del gineceo. (1) Carpelos; (2) Corte transversal de la base del carpelo u ovario; (3) Modelo de la estructura interna de un carpelo.
Estructura de la inflorescencia
Una inflorescencia es un grupo de flores anidadas en torno a un solo tallo organizador, el cual libera varias ramificaciones en torno a un patrón complejo. Básicamente se trata de varias flores relativamente pequeñas que maduran en progresión. El tallo que mantiene a toda la inflorescencia es denominado pedúnculo y los tallos principales que mantienen a las flores o a más ramificaciones se llama raquis. El tallo de cada flor individual se denomina pedicelo. Cuando los ovarios de una inflorescencia maduran a su estado de fruto, la estructura en su conjunto es denominada infrutescencia. La flor que pertenece a una inflorescencia es llamada floreta, especialmente cuando las flores particulares son muy pequeñas y nacen en un grupo anidado muy estrecho.

Figura 9.11. La flor que no es flor, pero si muchas flores. Las inflorescencias son flores apiñadas en torno a un tallo común llamado pedúnculo.
10. Controversia sobre la sexualidad de la flor
10. Controversia sobre la sexualidad de la flor
Durante muchos años, los botánicos y científicos estuvieron inmersos en una intrigante controversia sobre la sexualidad de las flores. Aunque hoy en día nos resulte evidente que las flores poseen órganos sexuales, hubo un tiempo en el que esta afirmación generó un intenso debate y especulaciones en diversos ámbitos científicos, filosóficos, religiosos y morales.
Argumentos religiosos
En la controversia sobre la sexualidad de las flores, los argumentos religiosos también desempeñaron un papel significativo. Diferentes interpretaciones de los textos sagrados generaron opiniones encontradas sobre si las flores podían considerarse sexualmente activas.
Algunas religiones, como ciertas ramas del hinduismo y el taoísmo, veían en la sexualidad de las flores un reflejo de la divinidad y la perfección de la creación. Estas religiones consideraban que la reproducción en todas sus formas era un acto sagrado y un recordatorio de la continuidad de la vida en el universo. Desde esta perspectiva, las flores y su capacidad para reproducirse eran vistas como manifestaciones de la divinidad misma.
Sin embargo, otras corrientes religiosas tenían interpretaciones más restrictivas. Algunas creencias consideraban que la reproducción solo podía tener lugar entre seres dotados de alma o conciencia, excluyendo así a las plantas, incluyendo las flores, de la categoría de seres sexualmente activos.
Estas divergencias religiosas reflejaban las diferentes cosmovisiones y enfoques en relación con la sexualidad y la reproducción. Mientras algunas religiones celebraban la diversidad y la sacralidad de todas las formas de vida, otras enfatizaban la centralidad de los seres humanos en el orden cósmico y limitaban la sexualidad a los seres dotados de alma.
A lo largo de la historia, numerosos teólogos y eruditos religiosos han participado en el debate sobre la sexualidad de las flores, aportando diferentes interpretaciones basadas en los textos sagrados de sus respectivas tradiciones religiosas. Estos eruditos, con sus estudios y publicaciones, han dejado una huella significativa en la forma en que las diferentes religiones abordan esta cuestión.

Figura 10.1. Agustín de Hipona, también conocido como San Agustín, fue un influyente teólogo, filósofo y obispo nacido el 13 de noviembre de 354 en Tagaste, una ciudad de la provincia romana de Numidia (en lo que hoy es Argelia). Es considerado uno de los pensadores más destacados de la Iglesia Católica y uno de los Padres de la Iglesia más influyentes. Agustín se destacó por su habilidad intelectual y se dedicó al estudio de la filosofía, la retórica y la literatura en su juventud. Tras una vida marcada por la búsqueda de la verdad y una serie de experiencias espirituales, se convirtió al cristianismo en el año 386 y fue bautizado por San Ambrosio en Milán. A lo largo de su vida, Agustín escribió numerosas obras teológicas y filosóficas que abordaban temas como la naturaleza de Dios, la gracia divina, la moralidad y la doctrina cristiana. Sus escritos más famosos incluyen "Confesiones", una autobiografía espiritual, y "La Ciudad de Dios", donde reflexiona sobre la relación entre la Iglesia y el mundo secular. San Agustín fue reconocido por su profunda influencia en el pensamiento cristiano y en la teología occidental. Sus ideas han sido estudiadas y debatidas a lo largo de los siglos, y su legado sigue siendo relevante en la actualidad. Falleció el 28 de agosto del año 430 en Hipona (hoy Annaba, en Argelia), dejando un impacto duradero en la historia del pensamiento religioso y filosófico.
Entre ellos, se destaca el teólogo cristiano Agustín de Hipona (354-430 d.C.), quien en su obra "La Ciudad de Dios" abordó la relación entre la sexualidad y la naturaleza, argumentando que las flores y otras formas de vida vegetal no poseían una sexualidad en el sentido humano, sino que se reproducían de manera puramente funcional.
En el ámbito islámico, el teólogo y filósofo persa Al-Farabi (872-950 d.C.) planteó su visión en su obra "La ciudad ideal". Sostenía que la sexualidad de las flores era un reflejo de la sabiduría y la perfección divina en la creación, y que cada forma de vida cumplía con su propósito y función designados por Alá.
Otro destacado erudito religioso es el filósofo indio Ramanuja (1017-1137 d.C.), quien en sus escritos védicos abordó la relación entre la sexualidad de las flores y la idea de la interconexión de todas las formas de vida en el universo. Según Ramanuja, la sexualidad de las flores era una manifestación de la armonía y la unidad de la creación divina. A medida que la comprensión científica y filosófica de la sexualidad se ha desarrollado, muchas religiones han adaptado sus perspectivas y han incorporado una visión más inclusiva de la sexualidad en la naturaleza, reconociendo que las flores también forman parte del vasto y diverso espectro de la reproducción en el mundo natural.

Figura 10.2. Ramanuja fue un filósofo y teólogo indio nacido en el siglo XI en la región de Tamil Nadu, en el sur de la India. Es considerado uno de los más influyentes líderes religiosos en la tradición del vaisnavismo, una rama del hinduismo dedicada a la adoración de Vishnu. Desde temprana edad, Ramanuja mostró un gran interés por la espiritualidad y el estudio de los textos sagrados. Se convirtió en discípulo del renombrado maestro Yamunacharya, quien le enseñó las enseñanzas del vaisnavismo y lo inició en la filosofía vedanta. Ramanuja desarrolló sus propias interpretaciones y enseñanzas, enfatizando la devoción amorosa a Vishnu y la creencia en la unidad de todos los seres con la divinidad. Promovió la idea de la rendición total a Dios y la importancia de la comunidad y la igualdad en la práctica religiosa. Sus enseñanzas se plasmaron en numerosos escritos y comentarios sobre los textos sagrados, como el Bhagavad-gita y los Vedas. Ramanuja viajó extensamente por la India, difundiendo sus enseñanzas y atrayendo seguidores devotos. Ramanuja dejó un legado duradero en la historia religiosa y filosófica de la India. Sus enseñanzas han influido en el desarrollo del vaisnavismo y continúan siendo estudiadas y seguidas por millones de personas en la actualidad. Su enfoque en la devoción, la igualdad y la unidad espiritual ha dejado una profunda huella en la tradición hindú y en el pensamiento religioso en general.
Argumentos morales
Desde una perspectiva moral, existían preocupaciones sobre la concepción de la sexualidad en las plantas. Algunos argumentaban que otorgarles una sexualidad a las flores podría abrir la puerta a interpretaciones inmorales o pervertidas de la naturaleza. Se temía que asignarles una sexualidad implicara reconocerles una agencia o voluntad consciente que no les correspondía.
Uno de los filósofos influyentes en este debate fue Jean-Jacques Rousseau, quien en su obra "Discurso sobre el origen y los fundamentos de la desigualdad entre los hombres" (1755) planteó que las plantas no tenían una vida moral o sexual, y que cualquier atribución de sexualidad a las flores era una proyección humana sobre la naturaleza. Rousseau argumentaba que al atribuirles una sexualidad a las flores, se desvirtuaba la belleza y la pureza de la naturaleza.

Figura 10.3. Jean-Jacques Rousseau fue un filósofo, escritor y músico suizo-francés nacido en Ginebra en 1712. Es conocido por sus ideas influyentes en los campos de la filosofía política, la educación y la literatura. Rousseau creció en un entorno modesto y recibió una educación limitada, pero su pasión por el aprendizaje y su talento innato lo llevaron a convertirse en uno de los pensadores más destacados de su tiempo. Sus escritos abordaron temas como la libertad, la igualdad y la naturaleza humana. Una de sus obras más importantes es "El contrato social", donde expuso su visión sobre el origen y la organización legítima de la sociedad. Rousseau argumentaba que el poder soberano residía en el pueblo y que las instituciones políticas debían estar basadas en el consentimiento popular. Además de su trabajo filosófico, Rousseau también fue un novelista y compositor notable. Su novela "La nueva Eloísa" y su autobiografía "Las confesiones" son ejemplos destacados de su habilidad literaria y su introspección emocional. Las ideas de Rousseau tuvieron un profundo impacto en la Ilustración y en el movimiento romántico posterior. Sus ideas sobre la educación también influyeron en la pedagogía moderna, promoviendo un enfoque más centrado en el desarrollo individual y la conexión con la naturaleza. Jean-Jacques Rousseau falleció en 1778, pero su legado perdura en la historia de la filosofía y la literatura. Sus ideas sobre la libertad, la igualdad y la autenticidad continúan siendo objeto de debate y reflexión en la actualidad.
Además, varios pensadores religiosos prominentes se opusieron firmemente a la idea de atribuir una sexualidad a las flores. Uno de ellos fue San Agustín, un influyente teólogo y filósofo cristiano del siglo IV. En su obra "Confesiones", San Agustín sostenía que la sexualidad era una característica exclusiva de los seres humanos y otros seres dotados de alma racional, y que las plantas carecían de esta capacidad. Para él, asignar una sexualidad a las flores sería una distorsión de la jerarquía moral establecida por Dios.
Asimismo, el teólogo y filósofo Santo Tomás de Aquino, en su obra "Summa Theologica", argumentaba que la sexualidad era un don divino otorgado solo a los seres humanos para la procreación y el disfrute dentro del matrimonio. Según su visión, las flores no tenían una sexualidad genuina, ya que carecían de alma y no estaban destinadas a la reproducción sexual.

Figura 10.4. Santo Tomás de Aquino fue un teólogo y filósofo medieval nacido en Italia en el año 1225. Es considerado uno de los pensadores más influyentes de la historia, especialmente en el ámbito de la filosofía y la teología católica. Nacido en una familia noble, Tomás de Aquino ingresó a la Orden de los Dominicos a una edad temprana. Durante su formación académica, estudió en diversas universidades destacadas de la época, como la Universidad de Nápoles y la Universidad de París. Sus obras más importantes incluyen la "Summa Theologiae" y la "Summa contra Gentiles", donde desarrolló una síntesis entre la filosofía aristotélica y la teología cristiana. Su enfoque fue un intento de reconciliar la razón y la fe, demostrando la coherencia entre la filosofía natural y la revelación divina. Santo Tomás de Aquino también hizo importantes contribuciones en otros campos, como la ética, la metafísica y la teoría política. Su método de pensamiento riguroso y su capacidad para abordar una amplia gama de temas lo convirtieron en una figura destacada en el pensamiento medieval. Fue canonizado como santo por la Iglesia Católica en el año 1323. Su legado se extiende más allá de su época y sus enseñanzas siguen siendo estudiadas y debatidas en la actualidad. Santo Tomás de Aquino dejó una profunda huella en la historia del pensamiento occidental y continúa siendo una referencia importante en la teología y la filosofía.
Estos pensadores religiosos, entre otros, tuvieron una influencia significativa en la comprensión moral y religiosa de la sexualidad de las flores. Sus argumentos se basaban en la creencia de que la sexualidad era una facultad exclusiva de los seres humanos y otros seres dotados de alma, y asignarles una sexualidad a las flores podría llevar a una confusión moral y una distorsión de los roles establecidos por Dios en la jerarquía moral.
No obstante, a medida que la ciencia botánica avanzó y se descubrieron más detalles sobre la estructura y función de las flores, se hizo evidente que las plantas sí poseen órganos sexuales y participan en la reproducción sexual. Esto llevó a una revisión de las perspectivas morales y religiosas, y a una comprensión más completa de la diversidad y complejidad de la sexualidad en el reino vegetal. Estas perspectivas morales influyeron en la forma en que se abordó la sexualidad de las flores durante mucho tiempo, restringiendo la investigación científica y limitando la comprensión de su papel en la reproducción. Sin embargo, a medida que la ciencia botánica avanzó y se realizaron observaciones más detalladas, se evidenció la presencia de órganos sexuales en las flores y se pudo comprender mejor su función en el proceso reproductivo.
A través de la investigación científica rigurosa, se logró superar las barreras morales y se demostró que las flores tienen una sexualidad intrínseca que desempeña un papel crucial en la reproducción de las plantas. Esta comprensión más profunda ha permitido un avance significativo en el conocimiento botánico y ha desafiado las concepciones morales y filosóficas previas sobre la sexualidad de las flores.
En última instancia, la superación de los argumentos morales en esta controversia ha permitido una apreciación más completa y precisa de la sexualidad en el mundo vegetal, enriqueciendo nuestro entendimiento de la naturaleza y su diversidad. Al romper con las restricciones morales, la ciencia botánica ha logrado avanzar en el conocimiento y la comprensión de las flores y su función vital en la perpetuación de la vida vegetal.
Argumentos biologicos
Durante la controversia sobre la sexualidad de las flores, varios científicos plantearon argumentos desde una perspectiva biológica para respaldar su postura. Uno de ellos fue Carl Linnaeus en el siglo XVIII, quien en su obra "Systema Naturae" (1758) sostenía que las flores no podían considerarse sexualmente activas, ya que carecían de los órganos reproductivos típicos de los animales.
En el siglo XIX, Augustin Pyramus de Candolle desafió esta idea en su libro "Théorie élémentaire de la botanique" (1813), argumentando que las flores eran estructuras reproductivas complejas y que los órganos reproductivos se encontraban dentro de ellas.

Figura 10.5. Augustin Pyramus de Candolle fue un botánico y naturalista suizo nacido el 4 de febrero de 1778 en Ginebra. Es conocido por sus contribuciones significativas en el campo de la botánica, especialmente en la taxonomía y la sistemática de las plantas. De Candolle provenía de una familia con tradición en el estudio de las plantas, lo que le permitió desarrollar un interés temprano en la botánica. Estudió medicina en la Universidad de Montpellier, pero su pasión por las plantas lo llevó a dedicarse por completo a la botánica. En 1813, publicó su obra más influyente, "Prodromus Systematis Naturalis Regni Vegetabilis", donde propuso un sistema de clasificación de las plantas basado en características morfológicas y fisiológicas. Su enfoque revolucionario sentó las bases para la taxonomía moderna y tuvo un impacto duradero en el campo de la botánica. De Candolle también realizó importantes estudios sobre la distribución geográfica de las plantas y la influencia de factores ambientales en su crecimiento y supervivencia. Contribuyó a la comprensión de la adaptación de las plantas al medio ambiente y su interacción con otros organismos. Además de su trabajo científico, De Candolle fue un destacado profesor y mentor de numerosos botánicos, lo que le permitió ejercer una gran influencia en el desarrollo de la botánica como disciplina. También fundó el Jardín Botánico de Ginebra, que se convirtió en un centro importante de investigación botánica. Augustin Pyramus de Candolle falleció el 9 de septiembre de 1841 en Ginebra, dejando un legado duradero en la botánica. Sus contribuciones científicas y su enfoque sistemático sentaron las bases para el estudio y la comprensión de las plantas, y su trabajo sigue siendo reconocido y valorado en la actualidad.
Sin embargo, fue Charles Darwin en su obra revolucionaria "El origen de las especies" (1859) quien proporcionó una sólida evidencia científica para respaldar la sexualidad de las flores. Darwin describió detalladamente los órganos reproductivos de las flores y explicó cómo la polinización y la fecundación sexual eran esenciales para la reproducción de las plantas con flores.
Los trabajos posteriores de Gregor Mendel sobre la herencia genética en los guisantes (1865) también respaldaron la idea de la sexualidad de las flores, al demostrar cómo los caracteres se transmitían a través de la reproducción sexual.
Estos avances científicos y las publicaciones de Linnaeus, De Candolle, Darwin y Mendel finalmente superaron los argumentos biológicos que negaban la sexualidad de las flores, estableciendo de manera concluyente que las flores son organismos altamente especializados en la reproducción sexual y desempeñan un papel crucial en la perpetuación de las especies vegetales..
Argumentos filosóficos
La controversia sobre la sexualidad de las flores también involucró argumentos desde una perspectiva filosófica. Algunos filósofos plantearon cuestionamientos sobre si las flores eran capaces de experimentar placer o deseo sexual, y si podían ser consideradas sexualmente activas.
Uno de los principales exponentes de esta postura fue Arthur Schopenhauer, quien en su obra "El mundo como voluntad y representación" (1818) argumentó que solo los seres vivos superiores y complejos eran capaces de experimentar la sexualidad debido a su conciencia y capacidad de deseo. Según Schopenhauer, las plantas, incluidas las flores, no poseían esta capacidad y, por lo tanto, no podían considerarse sexualmente activas.
Sin embargo, la visión filosófica comenzó a cambiar con el surgimiento de nuevas ideas y enfoques. Filósofos como Friedrich Nietzsche y Henri Bergson desafiaron la noción de que la sexualidad estaba exclusivamente ligada a la conciencia y el deseo humano. Estos filósofos propusieron una visión más amplia de la sexualidad, considerándola como una fuerza vital presente en todas las formas de vida, incluidas las plantas.
A medida que avanzaba el siglo XX, filósofos como Ludwig Wittgenstein y Martin Heidegger abordaron la cuestión de la sexualidad en términos de lenguaje y existencialismo. Estos filósofos argumentaron que la sexualidad no se limitaba a la experiencia subjetiva, sino que también tenía un significado y una función objetiva en la naturaleza.
Si bien la controversia filosófica sobre la sexualidad de las flores persistió durante un tiempo, los argumentos planteados por Nietzsche, Bergson y otros filósofos contribuyeron a una mayor comprensión de la sexualidad como un fenómeno presente en toda la naturaleza, incluidas las flores. Esta visión más amplia y holística de la sexualidad finalmente se integró en el entendimiento científico y filosófico, superando así los argumentos filosóficos que negaban la sexualidad de las flores.

Figura 10.6. Henri Bergson fue un filósofo francés nacido el 18 de octubre de 1859 en París. Es conocido por sus contribuciones al campo de la filosofía y su enfoque en la comprensión de la experiencia y la percepción humana. Bergson estudió en la École Normale Supérieure y se especializó en filosofía. Su obra más destacada, "La evolución creadora", publicada en 1907, tuvo un impacto significativo en la filosofía y le valió el reconocimiento internacional. Bergson desarrolló la idea de que la realidad es una corriente continua de cambio y movimiento, y argumentó que la experiencia humana no puede ser plenamente capturada por conceptos estáticos o racionales. Propuso que la intuición y la experiencia directa son fundamentales para comprender la realidad y la naturaleza del ser. Sus ideas sobre la duración, el tiempo y la memoria también fueron influyentes. Sostenía que el tiempo no puede ser medido ni reducido a unidades fijas, sino que debe ser entendido como una experiencia subjetiva y fluida. Bergson recibió numerosos reconocimientos a lo largo de su carrera, incluido el Premio Nobel de Literatura en 1927. Sus ideas filosóficas tuvieron un impacto duradero en la filosofía, la psicología y otros campos relacionados, y su influencia se extendió más allá de su tiempo. Henri Bergson falleció el 4 de enero de 1941 en París, dejando un legado como uno de los filósofos más importantes del siglo XX. Su enfoque en la experiencia vivida y su crítica a la visión mecanicista de la realidad continúan siendo objeto de estudio y debate en la filosofía contemporánea.
En resumen, la controversia sobre la sexualidad de las flores fue un tema de debate multidisciplinario que involucró aspectos biológicos, filosóficos, religiosos y morales. Sin embargo, gracias a los avances científicos y las contribuciones de destacados biólogos, hoy en día no existen dudas
11. Gametogénesis en las angiospermas
11. Gametogénesis en las angiospermas
Al igual que en los animales, las plantas deben pasar por un proceso complejo para el desarrollo de sus gametos masculinos y femeninos. La gametogénesis, el proceso mediante el cual se producen los gametos en las angiospermas, ha sido objeto de gran interés y estudio en el campo de la biología vegetal. Esta fascinante etapa del ciclo de vida de las plantas con flores es fundamental para la reproducción sexual y la diversidad genética de estas especies.
En este artículo, exploraremos en detalle el proceso de gametogénesis en las angiospermas, desde la formación de las células madre hasta la diferenciación de los gametos masculinos y femeninos. Analizaremos los eventos clave, las estructuras y las interacciones moleculares que ocurren durante la gametogénesis, así como su importancia evolutiva y las implicaciones en la mejora genética de las plantas. A través de esta investigación, esperamos ampliar nuestra comprensión de la gametogénesis en las angiospermas y su papel crucial en la perpetuación de la vida vegetal.
Desarrollo de las estructuras femeninas
En el fascinante mundo de las angiospermas, el desarrollo de las estructuras femeninas desempeña un papel fundamental en el proceso de reproducción. Estas estructuras, que incluyen el pistilo y los óvulos, son responsables de la formación de las semillas y la continuidad de la especie. En esta sección, exploraremos en detalle el desarrollo de estas estructuras, desde la formación del óvulo hasta la madurez del pistilo. Analizaremos los eventos clave, las transformaciones morfológicas y las interacciones moleculares que ocurren durante este proceso. Además, examinaremos la importancia evolutiva de estas estructuras y cómo han contribuido a la diversificación y adaptación de las angiospermas en su entorno. A través de este estudio, esperamos descubrir los secretos y mecanismos detrás del desarrollo de las estructuras femeninas en las angiospermas y apreciar su crucial papel en la perpetuación de la vida vegetal.
Integumento y micrópilo
A medida que el gametofito femenino se desarrolla, el megasporangio “semilla inmadura” se desarrolla. Dos capas de células crecen alrededor del megasporangio denominadas integumentos. Los integumentos darán lugar a la cascara de la semilla posteriormente. Los integumentos dejarán abierta una cavidad o conducto que permite al tubo polinífero encontrar su camino denominado micrópilo. En este punto al interior del megasporangio hay una célula octacarionte, es decir una sola célula con ocho núcleos, los cuales proceden a repartirse en parejas.

Figura 11.1. Modelo del integumento y el micrópilo
La meiosis en las angiospiermas desde el estado diploide al haploide
Mientras que la flor aún continúa siendo un botón, en su interior unas células madre denominadas megasporocito realizan una diferenciación de las demás células del ovario y subsecuentemente entra en meiosis.

Figura 11.2. La megaspora es originalmente una célula con ocho núcleos.
El producto de la meiosis se denomina megasporas y son cuatro. Poco después de haberse producido una de las megasporas absorbe los nutrientes mientras que las otras tres se degeneran y desaparecen finalmente. Posteriormente está célula superviviente “haploide” inicia la mitosis para generar en el gametofito femenino. Al igual que en las gimnospermas y en contraste con las demás plantas, el gametofito femenino se queda atrapado al interior del esporofito “al interior de la flor” por lo que no crece demasiado.
La megaspora como una célula octacarionte
La megaspora remanente crece, pero la división por mitosis no es tan simple como podría llegar a esperarse. Inicialmente solo los núcleos se dividen mitóticamente sin que se dé la citocinesis, lo que implica la formación de una célula octacarionte “ocho núcleos haploides en una misma célula”. Para el momento en que los núcleos se han dividido la célula ha crecido hasta que llena la cavidad del megasporangio.
El gametofito femenino
Los ocho núcleos de la megaspora forman dos grupos de cuatro células, posteriormente dos de esos núcleos migran al centro y serán llamados núcleos centrales, mientras que los dos grupos de tres células restantes recibirán el nombre de células polares. En algunas especies los dos núcleos centrales de fusionan para formar un solo núcleo diploide lo cual es raro para un gametofito. En cualquier caso, la megaspora procede a dividir su citoplasma en torno a 6 núcleos independientes y en torno a los dos núcleos centrales formando siete células. Las que se encuentran abajo se llaman sinérgidas, la sinérgida que está en contacto con el micrópilo se denomina óvulo. Las células opuestas a las sinérgidas se denominan antipódales. Todas juntas forman el gametofito femenino maduro o saco embrionario, el cual se encuentra a la espera de la llegada del gametofito masculino maduro.
Desarrollo de las estructuras masculinas
En el fascinante mundo de las angiospermas, el desarrollo de las estructuras masculinas es un proceso crucial para la reproducción y la diversidad de esta vasta clase de plantas con flores. En esta sección, exploraremos en detalle el desarrollo de estas estructuras, que incluyen los estambres y los granos de polen. Investigaremos cómo se forman, maduran y liberan los granos de polen, así como las etapas clave en la diferenciación y desarrollo de los estambres. Además, examinaremos las interacciones moleculares y los mecanismos genéticos que regulan este proceso, y cómo contribuyen a la viabilidad y eficiencia reproductiva de las angiospermas. También exploraremos las adaptaciones evolutivas de las estructuras masculinas en diferentes especies y su importancia para la polinización y la perpetuación de la vida vegetal. A través de este estudio, esperamos desentrañar los secretos detrás del desarrollo de las estructuras masculinas en las angiospermas y apreciar su impacto en la extraordinaria diversidad de estas fascinantes plantas.
La meiosis en las angiospiermas desde el estado diploide al haploide
Los procesos para formar el polen y el óvulo ocurren en paralelo en la mayoría de las especies dioicas. El proceso de la formación del polen o gametofito masculino toma lugar en las anteras. A medida que la antera madura, cuatro parches de tejido maduran formando cuatro sacos a lo largo de la antera llamados microsporangios.
Desarrollo de los sacos poliníferos
Cuando se realiza un corte cruzado de una antera es posible visualizar cuatro cámaras o cavidades denominadas sacos poliníferos o microsporangios alineados con células nutritivas denominadas tapetales. A medida que la antera va madurando, las paredes entre las cámaras adyacentes desaparecen dejando solo dos grandes sacos llamados microsporangios. Al interior del microsporangio se encuentran los microsporocitos que llevan a cabo el proceso de meiosis para producir después de una serie de alteraciones las microsporas. Estas microsporas haploides van madurando por un periodo de unas pocas semanas hasta producir el polen.


Figura 11.3. Diagrama y fotografía del corte transversal de una antera. Arriba a la izquierda la antera inmadura, y abajo la antera madura. Los microsporangios contienen los microsporocitos que son células diploides. Cada uno realiza la meiosis para producir una cuarteta o cuarteta de microsporas haploides.
Alteraciones de la microspora para convertirse en polen
Algunos de los cambios más importantes son: El núcleo de cada microsporocito se divide por mitosis para formar posteriormente la célula tubular y la célula vegetativa. Los miembros de cada cuarteta meiótica se separan al degradarse el cuerpo calloso, aunque de forma inusual algunas especies los mantienen unidos. Al madurar la microspora una segunda capa con una estructura esculpida o tallada es formada alrededor.
El grano de polen en las angiospermas
Cuando los eventos descritos en los artículos anteriores se han completado se tiene un grano de polen maduro. La capa externa del grano de polen se denomina exina que posee una estructura intrincada, contiene químicos que posteriormente se van a acoplar con químicos del estigma de la flor. Como resultado de estas reacciones, el grano de polen puede germinar rápido, lento o no hacerlo dependiendo de las limitaciones “si las hay” del estigma a reproducirse con sigo misma, con miembros cercanos de su familia entre otros factores evolutivos.
Utilidades de los granos de polen
El citoplasma del grano de polen es rico en vitaminas, por lo que el polen es recolectado para emplearlo como suplemento nutricional en algunos alimentos especializados. La exina de muchos granos de polen es virtualmente indestructible, por lo que granos de polen han sido encontrados en yacimientos con miles de años. El patrón de la exina es dependiente de la especie, por lo que puede emplearse para identificar la especie a la cual pertenece. Este rasgo ha sido importante para las investigaciones forenses en casos de conexión de un criminal a un lugar o fecha específicos.

Figura 11.4. El polen. (A) Microsporocitos en meiosis, la mayoría se encuentra aún en meiosis 1, pero a la derecha puede verse ya una cuarteta que ha finalizado la meiosis 2. (B) Un grano de polen al microscopio electrónico, los patrones de acanalado son criterios de clasificación taxonómica. (C) La polinización y la fertilización son procesos independientes, el primero es la llegada del polen y la segunda es la llegada de los espermas al óvulo por medio del tubo polínico.
El núcleo vegetativo y el núcleo generativo en el grano de polen
Los dos núcleos al interior del grano de polen se denominan: generativo y vegetativo/tubular. El núcleo generativo es aquel que al dividirse producirá dos núcleos que serán rodeados por la membrana plasmática y funcionarán como esperma “sin flagelos por eso no se los llama espermatozoides”. Dado que no pueden nadar por sí mismos, los espermas deben ser llevados hasta su blanco en el gametofito femenino. Esta función será realizada con el apoyo de la célula que rodea de facto a la célula germinativa y se denomina vegetativa. Es decir, la célula vegetativa tiene en su interior a la célula generativa, una vez activada, la célula vegetativa se estira formando el tubo polínico de forma similar a una hifa sin septos de un hongo.
12. Polinización
12. Polinización
Uno puede leer en la prensa o en revistas de que cuando una flor ha sido polinizada se sigue necesariamente de que también ha sido fertilizada, como si ambos procesos fueran el mismo. Al igual que en las gimnospermas la polinización y la fertilización son procesos independientes que dependen de una secuencia cronológica de hechos. La polinización se refiere a la llegada del polen al estigma de la flor, donde este puede o no ser activado mediante complejos mecanismos de reconocimiento hormonal.
La polinización se realiza con la ayuda de factores bióticos y abióticos que son los mismos que describimos para la dispersión de las semillas, aunque en algunos casos con grados de especialización aún mayores. La fertilización involucra la unión del óvulo con el espermatozoide, fenómeno que en las angiospermas es más complejo ya que involucra el proceso de fertilización doble que será discutido en artículos futuros.

Figura 12.1. Polinizadores. Las adaptaciones entre los polinizadores y as flores pueden llegar a ser muy intrincadas. La longitud de la lengua de este murciélago le asegura a la flor, que el llevará el polen solo a plantas con formas semejantes a la suya, lo cual disminuye el azar en el proceso de fertilización, y por ende debe gastar menos energía en la producción de polen (YouTube)
Adaptaciones a la polinización en las angiospermas
Aunque el viento sirve como uno de los polinizadores más importantes al igual que en las gimnospermas, una de las tendencias evolutivas más marcadas en las plantas con flor es a disminuir el factor azaroso en la reproducción.
Figura 12.2. Un abominable misterio. Dendrograma cronocalibrado que describe la cantidad de especies de cada linaje de plantas verdes, se puede apreciar que las plantas con flor experimentaron una diversificación rápida en comparación con los demás grupos de plantas.
Para esto las flores emplean polinizadores más especializados, los animales, mediante el establecimiento de una simbiosis mutualista. Los animales polinizadores son muy diversos, principalmente artrópodos y aves, pero también han sido registrados mamíferos.
Coevolución entre flores y polinizadores
La coevolución entre flores y polinizadores es un fenómeno fascinante que ha moldeado la evolución de numerosas especies de plantas y animales a lo largo de millones de años. Esta estrecha relación simbiótica entre las flores y los polinizadores es fundamental para la reproducción exitosa de muchas especies de plantas y la perpetuación de la vida vegetal en general.
El uso de polinizadores animales por parte de las plantas ha permitido una notable reducción en la cantidad de polen producido. En lugar de producir grandes cantidades de polen para asegurar la fertilización, las plantas invierten su energía en la generación de néctar, una sustancia dulce y rica en nutrientes que actúa como una especie de "moneda de pago" para los polinizadores. Al visitar las flores en busca de néctar, los polinizadores transportan inadvertidamente el polen de una flor a otra, facilitando así la polinización cruzada y la transferencia de material genético entre individuos.
Sin embargo, la interacción entre las flores y los polinizadores va más allá de la simple recompensa de néctar. Muchas especies de plantas han desarrollado adaptaciones especializadas para atraer y manipular a polinizadores específicos, actuando como verdaderos agentes selectores. Estas adaptaciones pueden incluir la formación de corolas con formas intrincadas que solo son compatibles con probóscides, picos o lenguas especializadas de determinados polinizadores. De esta manera, las plantas favorecen la polinización por parte de un grupo selecto de polinizadores, promoviendo la especialización tanto en las poblaciones de polinizadores como en las plantas que los atraen.
La coevolución entre flores y polinizadores no se limita únicamente a la forma y la morfología de las flores. Las interacciones químicas también desempeñan un papel crucial en esta relación simbiótica. Algunas flores son capaces de segregar químicos que imitan las feromonas reproductivas de los polinizadores, atrayéndolos de manera más efectiva. Estos compuestos químicos engañan a los polinizadores al hacerles creer que han encontrado una pareja reproductiva potencial, lo que los incita a visitar más flores y aumentar la eficacia de la polinización.
Esta coevolución entre flores y polinizadores tiene importantes consecuencias para la diversidad y la evolución de las especies. Las plantas dependen en gran medida de los polinizadores para su reproducción y supervivencia, mientras que los polinizadores dependen de las flores como fuente de alimento y recursos. Esta relación simbiótica ha llevado a la evolución de una gran variedad de formas y colores de flores, así como a la especialización de polinizadores en determinadas especies de plantas. Además, la desaparición de ciertos polinizadores puede tener efectos devastadores en las poblaciones de plantas que dependen exclusivamente de ellos, lo que destaca la fragilidad y la interdependencia de esta relación.
La coevolución entre flores y polinizadores abarca una amplia variedad de organismos, incluyendo diferentes tipos de artrópodos y aves.
Polinizadores artrópodos:
Los insectos son los polinizadores más comunes y diversos en la coevolución con las flores. Entre los insectos polinizadores se encuentran las abejas, los escarabajos, las mariposas, las polillas y los mosquitos. Cada grupo de insectos polinizadores tiene sus propias características morfológicas y comportamentales que se han adaptado a las diferentes estructuras de las flores.
(a) Abeja europea (Apis mellifera): Las abejas melíferas son polinizadores clave para una amplia variedad de plantas, incluyendo cultivos agrícolas y plantas silvestres.
(b) Abeja orquídea (Euglossa spp.): Estas abejas son conocidas por su papel en la polinización de orquídeas, especialmente en las especies de la subfamilia Epidendroideae.

Figura 12.3. Euglossa dilemma es una especie de abeja conocida como abeja de las orquídeas, debido a su estrecha relación con las flores de orquídeas. Esta abeja se encuentra principalmente en América Central y América del Sur, incluyendo regiones de Colombia. La Euglossa dilemma tiene un tamaño mediano, con aproximadamente 10-12 mm de longitud. Su cuerpo es delgado y está cubierto de vellosidad, y presenta colores llamativos y variados. El abdomen puede mostrar tonalidades metálicas brillantes, como verde o azul, mientras que la cabeza y el tórax pueden presentar segmentos de color azul o verde. Esta especie de abeja juega un papel importante en la polinización de las orquídeas, ya que visita las flores en busca de néctar y polen. A medida que se alimenta, recoge polen en su cuerpo y lo transporta de una flor a otra, permitiendo así la fertilización cruzada y la reproducción de las orquídeas. La Euglossa dilemma es considerada una especie clave en los ecosistemas donde se encuentra, contribuyendo a la diversidad y preservación de las orquídeas y otros tipos de plantas. Su presencia es un indicador de la salud y la biodiversidad de los ecosistemas en los que habita.
(c) Escarabajo de flores (Melolontha melolontha): Este escarabajo se alimenta de pétalos y polen de diversas flores, contribuyendo a la polinización de especies vegetales.
(d) Escarabajo estercolero (Scarabaeus spp.): Estos escarabajos polinizan plantas que atraen a los escarabajos mediante el olor a materia orgánica en descomposición, como algunas especies de flores cadáver.

Figura 12.4. Melolontha melolontha, comúnmente conocido como escarabajo común de mayo, es una especie de escarabajo perteneciente a la familia Scarabaeidae. Este escarabajo es ampliamente distribuido en Europa, incluyendo regiones de España, Francia, Alemania y el Reino Unido. El escarabajo común de mayo tiene un cuerpo robusto y redondeado, con una longitud de aproximadamente 2 a 3 centímetros. Su exoesqueleto es de color marrón oscuro o negro, con un ligero brillo. Posee antenas largas y delgadas, y sus patas son fuertes y adaptadas para excavar en el suelo.
Estos escarabajos son conocidos por su actividad durante el mes de mayo, cuando emergen de sus larvas subterráneas y se vuelven abundantes en áreas rurales y boscosas. Son principalmente herbívoros y se alimentan de una variedad de plantas, incluyendo hojas, flores y raíces. El ciclo de vida de Melolontha melolontha es de aproximadamente tres años. Las hembras depositan sus huevos en el suelo, y las larvas, conocidas como gusanos blancos, se desarrollan bajo tierra alimentándose de materia orgánica. Una vez completada su etapa larval, se transforman en pupas y finalmente emergen como adultos.
Aunque el escarabajo común de mayo puede ser considerado una plaga en algunos cultivos agrícolas, también juega un papel importante en el ecosistema como descomponedor, contribuyendo a la descomposición de materia orgánica y al ciclo de nutrientes en el suelo. Además, sus larvas son una fuente de alimento para diversas especies de aves y mamíferos.
(e) Mariposa monarca (Danaus plexippus): Durante su migración, las mariposas monarca polinizan diversas especies de flores, especialmente aquellas pertenecientes a la familia Asclepiadaceae.
(f) Mariposa cola de golondrina (Papilio machaon): Esta mariposa poliniza plantas de la familia Apiaceae, como el hinojo y el eneldo.
(g) Polilla esfíngida del tabaco (Manduca sexta): Esta polilla es un polinizador nocturno que juega un papel importante en la polinización de plantas como el tabaco y el datura.
(i) Polilla del rosal (Cacoecimorpha pronubana): Esta polilla poliniza flores de rosas y otras especies de plantas relacionadas.

Figura 12.5. Cacoecimorpha pronubana, conocida comúnmente como polilla del racimo de la vid o polilla del racimo europea, es una especie de lepidóptero de la familia Tortricidae. Esta polilla es nativa de Europa, pero se ha extendido a otras partes del mundo, incluyendo América del Norte y del Sur, África y Australia. La polilla del racimo de la vid tiene una envergadura de alrededor de 1.5 a 2 centímetros. Sus alas delanteras tienen un patrón distintivo de coloración marrón con manchas y rayas más oscuras. Las alas traseras son más pálidas en comparación. Además, presenta un patrón de escamas de color plateado en la cabeza y el tórax. Esta especie es considerada una plaga agrícola, ya que se alimenta de los racimos de uvas y otros frutos, causando daños significativos en los cultivos de vid. Las larvas de la polilla se desarrollan dentro de los racimos de uvas, alimentándose de la pulpa y dejando daños y pudrición en el fruto. Esto puede resultar en pérdidas económicas para los viticultores. El ciclo de vida de Cacoecimorpha pronubana comienza con la puesta de huevos en los racimos de uvas. Las larvas emergen y se alimentan de la pulpa, pasando por varias etapas de desarrollo antes de pupar en el suelo o en refugios cerca de los cultivos. Finalmente, emergen como adultos y comienza un nuevo ciclo. El control de esta plaga se realiza a través de diversas estrategias, como el uso de insecticidas, trampas de feromonas y la implementación de prácticas culturales para reducir la población de polillas. Es importante destacar que Cacoecimorpha pronubana es una especie introducida en muchas regiones fuera de su área de distribución original, lo que ha causado problemas en la agricultura y ha requerido la implementación de medidas de control para mitigar sus impactos.
(j) Alfred Russel Wallace predijo que la polilla de la yuca (Goniopteryx yuccasella) y la flor asociada es la yuca (Yucca sp.) tenían una relación de polinización mutua en la cual la polilla era el polinizador principal de la flor de la yuca. Esta predicción se basó en la observación de la estructura especializada de la flor de la yuca y las características de la polilla, como su aparato bucal adaptado para recolectar polen y su comportamiento de búsqueda de néctar en las flores de la yuca. Esta relación mutualista entre la polilla de la yuca y la yuca ha sido ampliamente estudiada y es considerada un ejemplo clásico de coevolución entre polinizadores y plantas.
(j) Mosquito de los pantanos (Wyeomyia smithii): Este mosquito poliniza la planta llamada Vaccinium stamineum, también conocida como "mosquito-berry" por su asociación con este insecto.
(j) Mosquito Aedes albopictus: Aunque es principalmente conocido por ser un vector de enfermedades, algunos estudios sugieren que los mosquitos Aedes albopictus también pueden desempeñar un papel en la polinización de algunas especies vegetales.
Estos son solo algunos ejemplos de parejas polinizador-insecto altamente especializados. Cabe mencionar que existen muchas más especies de insectos que desempeñan un papel crucial en la polinización de una amplia variedad de plantas en diferentes ecosistemas.
Polinizadores aves:
Los polinizadores aves desempeñan un papel vital en la polinización de muchas especies de plantas. A diferencia de los insectos, las aves tienen la capacidad de volar largas distancias y acceder a flores con características específicas adaptadas a su morfología y forma de alimentación. Este grupo de polinizadores se caracteriza por su atractivo colorido, su capacidad de vuelo y su relación simbiótica con las plantas. En esta sección, exploraremos los diferentes tipos de polinizadores aves, sus características únicas y su importancia en la reproducción de numerosas especies vegetales.
(a) Colibríes: Los colibríes son aves especializadas en la polinización de flores. Poseen picos largos y delgados que les permiten acceder al néctar en corolas tubulares. Su capacidad de volar en el aire estacionario les facilita la visita a múltiples flores.
La especie de colibrí más importante en la polinización del café en Colombia es el colibrí esmeralda (Amazilia tzacatl). Este colibrí es nativo de América Central y del Sur y se encuentra ampliamente distribuido en Colombia. Su tamaño y comportamiento lo hacen ideal para la polinización del café, ya que su pico largo y curvado le permite acceder al néctar en las flores tubulares del cafeto.

Figura 12.6. El colibrí esmeralda (Amazilia tzacatl) es una especie de colibrí presente en América Central y del Sur, incluyendo Colombia. Con su plumaje verde brillante y vientre blanco, este colibrí de tamaño mediano deslumbra con su vuelo rápido y ágil. Su pico curvado le permite alimentarse del néctar de flores tubulares, desempeñando así un papel crucial en la polinización de plantas como flores tubulares y árboles frutales. Su visita a las flores facilita la transferencia de polen, promoviendo la fertilización cruzada y la reproducción de las plantas. Este colibrí añade color y movimiento a los paisajes naturales, siendo un componente esencial para la diversidad y reproducción de las plantas que dependen de su polinización.
El colibrí esmeralda es un polinizador eficiente y activo, visitando numerosas flores de café en busca de néctar. Durante sus visitas, recoge polen en su cabeza y plumaje, transfiriéndolo de una flor a otra a medida que se mueve de planta en planta. Esta actividad de polinización es esencial para la fertilización cruzada del café y, por lo tanto, para el desarrollo adecuado de los frutos.
La presencia del colibrí esmeralda en las plantaciones de café es un indicador de la salud del ecosistema y de la calidad de la biodiversidad. Su papel en la polinización del café contribuye a mantener la producción sostenible y la diversidad genética de los cultivos de café en Colombia.
(a) Aves nectarívoras: Además de los colibríes, existen otras aves nectarívoras que participan en la polinización de ciertas flores. Algunas especies de loros y pájaros carpinteros también pueden actuar como polinizadores en determinadas regiones.

Figura 12.7. El Colaptes melanochloros, conocido comúnmente como pájaro carpintero verde, es una especie de ave de tamaño mediano que se encuentra en varias regiones de América del Sur, incluyendo Argentina, Bolivia, Brasil y Paraguay. Su apariencia distintiva incluye un plumaje predominantemente verde, con tonos más claros en la parte inferior y un patrón de barras negras y blancas en su espalda. Tiene un pico largo y puntiagudo, ideal para buscar alimento y construir nidos en árboles. Estas aves se alimentan principalmente de insectos y larvas, a los que atrapan con su lengua pegajosa. Además de su importancia ecológica al controlar las poblaciones de insectos, el pájaro carpintero verde también desempeña un papel significativo en la dispersión de semillas y la polinización de plantas mientras busca néctar en las flores. Su presencia vibrante y su distintivo llamado resonante añaden color y sonido a los bosques donde habita.
El pájaro carpintero verde (Colaptes melanochloros) es un ave nectarívora que desempeña un papel agroindustrialmente importante en Colombia. Aunque no es un colibrí, este pájaro se alimenta de néctar de diversas flores y juega un papel crucial en la polinización de plantaciones agrícolas. Su presencia es especialmente beneficiosa en plantaciones de árboles frutales, como los cítricos y los aguacates.
Estas aves se alimentan de insectos y larvas que pueden dañar los cultivos, ayudando así a controlar las plagas de manera natural. Además, su actividad de búsqueda de alimento y construcción de nidos en los árboles contribuye a la salud y estructura de los bosques, promoviendo un entorno propicio para el desarrollo de los cultivos. También se ha observado que el pájaro carpintero verde juega un papel importante en la dispersión de semillas y la polinización de plantas, lo que favorece la diversidad y el mantenimiento de la vegetación en las plantaciones. En resumen, la presencia del Colaptes melanochloros en las plantaciones de frutas en Colombia es esencial para el equilibrio ecológico y el buen desarrollo de los cultivos.
Polinizadores anfibios
Existen polinizadores anfibios, aunque su papel en la polinización es menos conocido y estudiado en comparación con los polinizadores insectos y aves. Algunas especies de anfibios, como las ranas y los sapos, pueden visitar flores en busca de néctar, y en el proceso pueden transportar polen de una flor a otra. Sin embargo, su contribución a la polinización es considerada incidental en la mayoría de los casos, ya que su principal interacción con las flores es para obtener alimento, como néctar o insectos. Además, su movilidad limitada y su dependencia del agua para reproducirse limitan su capacidad de ser polinizadores eficientes en comparación con otros grupos de animales. Aunque los polinizadores anfibios no son tan comunes o destacados como los insectos o las aves, su papel en la polinización sigue siendo objeto de estudio y se reconoce su contribución a la reproducción de algunas plantas.
Polinizadores reptiles:
Algunos lagartos, como los geckos, pueden visitar flores en busca de néctar, y aunque su papel en la polinización es menos frecuente, se ha observado en algunas situaciones.
(a) Lagarto de la especie Uta stansburiana: Este lagarto, también conocido como lagarto de collarejo, ha sido registrado como polinizador en áreas desérticas de Estados Unidos y México. Se ha observado que mientras se alimenta de néctar de ciertas flores, su cabeza se cubre de polen, transportándolo inadvertidamente a otras flores.
(b) Lagarto de la especie Pogona vitticeps: Este lagarto, comúnmente conocido como dragón barbudo, ha sido reportado como polinizador en su hábitat natural en Australia. Se ha observado que durante su búsqueda de néctar, su lengua se cubre de polen, permitiendo la transferencia del mismo entre las flores.

Figura 12.8. El Pogona vitticeps, o dragón barbudo, es un reptil de tamaño medio originario de Australia. Posee un cuerpo robusto, cabeza triangular y una barba que puede inflar en momentos de excitación. Su coloración varía de tonos marrones a naranjas y rojos brillantes. Son omnívoros y se alimentan de insectos, vegetación y frutas. Son excelentes escaladores y pasan tiempo en el suelo para tomar el sol. Son conocidos por su naturaleza dócil y capacidad para interactuar con sus cuidadores. Los Pogona vitticeps han ganado popularidad como mascotas en todo el mundo debido a su apariencia única y su comportamiento interesante. Son reptiles fascinantes que aportan diversidad y belleza a los entornos domésticos.
Cabe destacar que la contribución de los lagartos a la polinización es menos frecuente y relevante en comparación con otros polinizadores, y su participación varía según las especies de lagartos y las plantas involucradas.
Mamíferos
Algunas especies de murciélagos se alimentan de néctar y polen, y durante sus vuelos nocturnos, pueden transportar el polen de una flor a otra. En Colombia, tres especies de murciélagos nectarívoros desempeñan un papel crucial en la polinización de plantas de importancia económica, especialmente en la agroindustria. Estas son:
(a) Glossophaga soricina: Conocido como el murciélago pescador, es un polinizador clave para cultivos como el aguacate y la guayaba. Su lengua larga y delgada le permite alcanzar el néctar en las flores, facilitando así la transferencia del polen entre plantas.
(b) Lonchophylla robusta: Este murciélago es un polinizador esencial para el banano y otras plantas de la familia de las Musáceas. Su actividad de polinización contribuye al desarrollo de frutas de alta calidad y buen rendimiento en las plantaciones de banano.
(c) Anoura caudifer: Conocido como el murciélago cola de ratón, es un polinizador importante para cultivos como el cacao. Su rol en la polinización asegura la producción de flores femeninas y la formación adecuada de los frutos de cacao.
Estos murciélagos nectarívoros desempeñan un papel fundamental en la polinización de estas plantaciones, y su ausencia podría tener consecuencias significativas, incluyendo la disminución de la producción y pérdidas económicas para la agroindustria.

Figura 12.9. Glossophaga soricina, conocido como murciélago de lengua larga, es un nectarívoro importante en Colombia. Con su hocico puntiagudo y lengua tubular, se alimenta del néctar de las flores. Esta especie desempeña un papel esencial en la polinización de plantas como el aguacate y la guayaba, cruciales para la agroindustria del país. Su capacidad de recolectar néctar y transportar polen de una flor a otra asegura una reproducción exitosa y una buena producción de frutos. Su vuelo ágil y metabolismo rápido le permiten visitar múltiples flores en busca de alimento. La presencia de Glossophaga soricina beneficia a los productores agrícolas al aumentar la producción y calidad de los cultivos, generando importantes beneficios económicos.
En algunas regiones, ciertos roedores como los ratones del género Melomys han sido identificados como polinizadores de plantas específicas.
Es importante destacar que la coevolución entre flores y polinizadores es altamente específica y depende de la adaptación conjunta de las características morfológicas, fisiológicas y de comportamiento de ambos grupos. Cada polinizador ha desarrollado una relación simbiótica particular con ciertas especies de plantas, lo que ha llevado a la diversidad y especialización de las flores para atraer y recompensar a sus polinizadores específicos.
Autopolinización y el abominable misterio de Darwin
Aunque algunas especies plantean bloqueos hormonales que impiden la endogamia entre polen y estigmas, muchas plantas con flor tienen la capacidad de autopolinizarse, el ejemplo típico es la alverja o guisante, el grosor representa la cantidad de especies de cada linaje, mientras que el eje (y) representa la escala geológica-cronológica. En contraste con los linajes de gimnospermas que o conservaron su diversidad o perdieron especies después del límite cretáceo terciario, l angiospermas han experimentado una diversificación incesante. Las implicaciones genéticas de este proceso generalmente involucran procesos de especiación instantáneos en caso de que se generen poliploides. Esta especiación instantánea es una de las respuestas al abominable misterio de Darwin, a diferencia de las gimnospermas, las angiospermas tienden a evolucionar muy rápido por poliploidias. Por esta razón, aunque las gimnospermas son más viejas, tienen menos especies y son menos diversas.
El ser humano como polinizador
El ser humano desempeña un papel importante como polinizador de plantas, especialmente en cultivos agrícolas y huertos. A través de actividades como la agricultura, la jardinería y la apicultura, los humanos intervienen en la transferencia de polen de las partes masculinas a las partes femeninas de las flores, facilitando la polinización y la producción de frutos y semillas.
Esta intervención humana en la polinización es especialmente crucial en especies como el manzano (Malus domestica), el peral (Pyrus communis), el melocotonero (Prunus persica) y otros árboles frutales, así como en cultivos como el tomate (Solanum lycopersicum), el pimiento (Capsicum annuum) y las plantas de calabaza (Cucurbita spp.). Estas plantas dependen en gran medida de la polinización realizada por los humanos o por abejas domesticadas, ya que sus flores pueden requerir una transferencia de polen precisa y controlada.
La importancia del ser humano como polinizador radica en garantizar la productividad y calidad de los cultivos, asegurando la producción de alimentos y otros productos agrícolas. Además, el manejo de las colmenas y la cría de abejas por parte de los humanos también contribuyen a la polinización de diversas plantas, ya que las abejas son polinizadores clave en muchos ecosistemas. En resumen, la intervención humana en la polinización es esencial para el mantenimiento de la seguridad alimentaria y el sustento económico en numerosas regiones del mundo.
En Colombia, algunos cultivos requieren la polinización por parte de trabajadores humanos debido a diversas razones. Uno de los casos más destacados es el cultivo del café (Coffea spp.), donde la polinización manual es fundamental. A diferencia de otros países productores de café donde se utilizan abejas como polinizadores naturales, en Colombia, debido a la altitud y otras condiciones ambientales, las abejas no son eficientes para la polinización del café. Por lo tanto, se requiere la intervención humana para realizar la polinización manualmente, mediante la transferencia del polen de una flor a otra. Aunque en Colombia, los colibríes desempeñan un papel importante en la polinización de algunas plantaciones de café, pero no logran polinizar todas debido a varias razones.

Figura 12.10. Hypothenemus hampei, conocido comúnmente como la "broca del café", es un pequeño escarabajo que pertenece a la familia Curculionidae. Es considerado uno de los principales insectos plagas en las plantaciones de café en todo el mundo, incluyendo en Colombia. Sin embargo, es importante destacar que la broca del café no es un polinizador, sino un insecto perjudicial que se alimenta de los granos de café. Este escarabajo tiene un ciclo de vida completo que involucra la infestación de los frutos del café, donde la hembra deposita sus huevos. Las larvas se desarrollan dentro de los granos de café, causando daños significativos a la calidad y cantidad de la cosecha. La presencia de la broca del café puede resultar en pérdidas económicas importantes para los productores de café. Para controlar la infestación de la broca del café, se emplean diversos métodos de manejo integrado de plagas, como la recolección y destrucción de frutos afectados, el uso de trampas y la aplicación de insecticidas selectivos. Estas medidas son fundamentales para proteger las plantaciones de café y asegurar una producción de alta calidad.
Una de ellas es que la distribución geográfica de las especies de colibríes puede variar, y es posible que algunas áreas de plantaciones de café no cuenten con la presencia de colibríes polinizadores. Además, los colibríes tienen preferencias específicas en cuanto a los tipos de flores que visitan, y pueden mostrar una mayor afinidad por ciertas especies de plantas que por otras. Esto significa que pueden concentrarse en polinizar ciertos cultivos o especies de plantas que se ajustan a sus necesidades alimenticias, dejando de lado otras plantaciones de café. Otro factor a considerar es que la densidad de colibríes puede ser limitada en algunas regiones, lo que puede dificultar una polinización completa en todas las plantaciones de café. Es importante destacar que los colibríes no son los únicos polinizadores del café. También intervienen otros agentes polinizadores, como abejas, moscas y algunas especies de escarabajos. La combinación de diferentes polinizadores es fundamental para asegurar una adecuada polinización y producción de café en Colombia.
Otro ejemplo es el cultivo del cacao (Theobroma cacao). En Colombia, la principal especie de mosquito (Forcipomyia spp.) que suele ser responsable de la polinización del cacao no es abundante en todas las regiones cacaoteras, lo que limita la polinización natural. Por tanto, los agricultores y trabajadores deben realizar la polinización manual para asegurar una buena producción de cacao.

Figura 12.11. Forcipomyia saetheri es una especie endémica de la región amazónica y se encuentra ampliamente distribuida en las áreas donde se cultiva el cacao. Estos mosquitos son atraídos por el aroma del néctar producido por las flores del cacao y se alimentan del néctar y del polen de estas flores. Durante este proceso, se produce la polinización cruzada, que es esencial para la fertilización y el desarrollo de los frutos de cacao. La presencia y actividad de Forcipomyia saetheri en las plantaciones de cacao contribuye significativamente a la producción de frutos de alta calidad. Por lo tanto, su presencia y su papel como polinizadores del cacao son de gran importancia para la industria del cacao en Colombia y otros países productores.
En ambos casos, la falta de polinizadores naturales adecuados en ciertas regiones de Colombia, debido a factores como la altitud, la temperatura y la distribución geográfica, hace necesaria la intervención humana para garantizar una polinización efectiva y un rendimiento adecuado de los cultivos. Esto resalta la importancia del papel de los trabajadores humanos como polinizadores en estos casos específicos.
13. Fertilización
13. Fertilización
Una cosa es que el polen llegue a la flor, y otra cosa diferente que el óvulo de la flor sea fecundado, en esta sección discutiremos la serie de eventos que vinculan a estas dos ideas.
Condiciones para el desarrollo del gametofito masculino maduro
Una vez que el polen hace contacto con el estigma de la flor, este debe segregar y recibir una serie de compuestos químicos que sirven como señalización, esto permite identificar la compatibilidad de especie entre polen y flor. Sin embargo, las plantas con flor poseen mecanismos como la alopoliploidía que permiten la fertilización entre especies distintas altamente emparentadas.
Las condiciones también pueden establecer que el polen no se desarrolla si es el que produce la misma planta, sin embargo. las plantas con flor tienen una fuerte tendencia a la endogamia, y más a la endogamia absoluta “autofertilización”. En resumen, las dos condiciones son:
(1) Polen de una especie diferente;
(2) Polen de un individuo diferente.

Figura 13.1. Germinación del gametofito masculino. (A-D) Germinación del tubo polínico al microscopio óptico. (E) Polen con tubo polínico germinado. (F) Camino que debe cubrir el tubo polínico en crecimiento. (G) Grano de polen y tubo polínico con el núcleo generativo a la vista del microscopio óptico.
Pero dichas condiciones no son en ningún caso absolutas, por ejemplo, dos especies diferentes llamadas Raphanous y Brassica pueden permitir el crecimiento de su polen, respectivos sin bloquearlos. Una vez que el polen es activado absorbe agua y otros fluidos suministrados por el estigma activando una de las células en su interior, la cual prolifera. La estructura generada infla el polen y lo abre por una ranura.
Tamaño y tiempo de maduración del gametofito masculino, el tubo polínico
La estructura que rompe el grano de polen se denomina tubo polínico. Se trata de una célula única que crece en la región apical, el núcleo de esta célula se encontrará siempre en la región apical permitiendo su crecimiento, detrás del núcleo se encuentra la célula germinativa.
En el maíz el tubo polínico crece más de 50 centímetros antes de arribar al carpelo. Al igual que sucedía con las esporas de otros linajes vegetales, el camino del tubo polínico es guiado por hormonas segregadas por el ovario femenino ubicado en los carpelos. En la mayoría de las plantas la distancia entre el estigma y los carpelos es considerablemente inferior.
El tubo polínico debe viajas desde el estigma en la punta del pistilo, a través de todo el pistilo hasta el carpelo donde se ubican las semillas inmaduras o mesgasporangios donde fertilizará uno de los óvulos del gametofiito femenino al interior del megasporangio. El viaje del tubo polínico también es variable, en algunas especies puede llegar a tomar menos de 24 horas, mientras que en otras puede llegar a tomar más de un año tal como ocurre en las gimnospermas. Aun así, el promedio no es mayor a dos días.
Gametofito masculino maduro y el tubo polínico
A medida que el tubo polínico crece “célula vegetativa”, la mayor parte del contenido del grano de polen es descargado en la punta del tubo polínico. El núcleo vegetativo se mantiene en la punta, mientras que la célula generativa “dentro del citoplasma vegetativo” realiza una mitosis para formar dos células que serán llamadas espermas. En algunas especies el núcleo generativo se divide antes de que el tubo polínico haya comenzado a formarse. El grano de polen con su tubo polínico, el núcleo vegetativo y dos células espermáticas constituye el gametofito masculino maduro. Existen estructuras accesorias que mantienen a las células generativas y al núcleo vegetativo en la punta como una especie de tampón.
Doble fertilización y formación del endospermo triploide
Cuando el tubo polínico alcanza el carpelo encuentra su camino hacia el megagametofito gracias a las hormonas liberadas a través del micrópilo. El tubo atraviesa el micrópilo destruyendo las dos células sinérgidas que se encuentran en estado de degeneración.
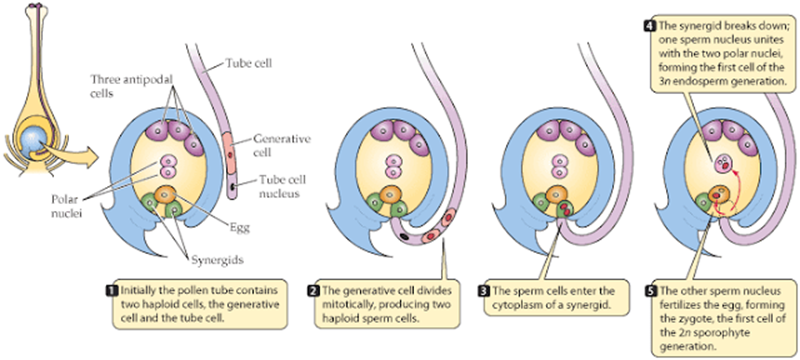
Figura 13.2. La doble fertilización. Llegada del tupo polínico (1) llevando el núcleo generativo, que se divide en dos células espermáticas (2), las cuales entran en el citoplasma de una sinergida; la sinergida se rompe y el núcleo del esperma se une con dos cuerpos polares generando un núcleo 3n (4), mientras que el otro núcleo se une al óvulo verdadero (5).
El proceso que sigue a continuación es único de las angiospermas y algunas gnetofitas y se denomina fertilización doble. A diferencia de lo que sucede en las gimnospermas donde uno de los espermas se pierde, en las angiospermas ambas espermas fecundan. El primer esperma fecunda al óvulo para formar el embrión del nuevo individuo, mientras que el segundo esperma fertiliza la célula central que o posee dos núcleos haploides o ya posee un núcleo diploide. Los dos resultados de la doble fertilización son, una célula diploide o cigoto y una célula triploide o endospermo.
14. Desarrollo embrionario en angiospermas
14. Desarrollo embrionario en angiospermas
Destinos del endospermo triploide
El endospermo triploide se hace particularmente activo y se divide repetidamente por mitosis”, con ciclos celulares que se completan en unas pocas horas, usualmente sin la formación de paredes celulares. En endospermo forma una capa protectora que rodea al embrión en desarrollo, convirtiéndose en algunas especies en una estructura nutritiva y/o carnosa.
En algunas monocotiledóneas como los pastos el endospermo se convierte en un tejido integral de la semilla “frutos llamados granos no distinguibles de la semilla”, pero en la mayoría de las dicotiledóneas como las leguminosas el endospermo se convierte en la estructura nutritiva la cual posteriormente es absorbida por los cotiledones. Esta estructura nutritiva mantendrá alimentado el embrión durante su desarrollo temprano.
Formación del embrión en las angiospermas
Paralelo a la formación del endospermo triploide el embrión empieza a desarrollarse, tres segmentos son importantes. Inicialmente el embrión se especializa en dos partes, el embrión en sí que crece hacia la punta apical de la semilla.

Figura 14.1. Embrión 2n y endospermo 3n. Los integumentos de la semilla son diploides y maternos, mientras que el integumento es triploide con doble cromosoma femenino y una serie masculino, el embrión si es diploide masculino y femenino.
La punta opuesta al crecimiento se denomina tejido suspensor y su función inicial es la de transferir nutrientes desde el endospermo hasta el embrión. A medida que el embrión madura empieza a absorber nutrientes directamente del endospermo sin la intervención del suspensor. En este punto se diferencia lo que va a ser el embrión en si del cotiledón. La diferenciación se realiza con la formación de la plúmula “shoot tip”, más arriba de la plúmula está el cotiledón y por debajo de la plúmula el embrión hasta el suspensor.
Maduración de los integumentos y maduración de la semilla
Los dos integumentos que rodean al megagametofito proceden a endurecerse, mientras que en el interior el gametofito femenino restante se degenera. De las siete células femeninas solo son empleadas la célula central originada por los dos núcleos polares y el óvulo de las sinérgidas, las otras dos sinérgidas y las tres antipodales se degeneran a medida que el endospermo inicia su desarrollo. Como conclusión de estos eventos, el óvulo se ha convertido en una semilla, y al mismo tiempo todos los carpelos que rodean a las semillas se transforman en una fruta.
Apomixia y partenocarpia
Tienen que ver con la semilla y el fruto. En la apomixia se generan semillas viables clonales a la célula madre, por lo que puede clasificarse como otro tipo de reproducción asexual de las angiospermas. En esta el embrión se forma a partir de una célula diploide de la madre, ejemplos de este tipo de reproducción lo dan los dientes de león. Partenocarpia significa fruto virgen, en este sentido hace referencia a procesos usualmente artificiales que buscan la formación de frutos carnosos sin semillas. Sin embargo, la partenocarpia también ocurre en la naturaleza, algunos ecólogos han propuesto que es un mecanismo de defensa contra los depredadores de semilla que omiten los frutos. Si un pájaro que no se alimenta del fruto encuentra un fruto sin semilla lo asociará con un esfuerzo sin recompensa y no lo atacará más. Otra ventaja de un fruto partenocárpico es que hace que la planta genere frutos aun en periodos con baja producción de semilla lo que permite reforzar las relaciones de simbiosis con los dispersadores de semilla, que comen el fruto, pero no la semilla. En un año con baja producción de semilla y sin partenocarpia, los dispersadores de semilla se morirían de hambre o emigrarían, haciendo que, en otros años con más producción de semillas, la planta encuentre más dificultades para reproducirse.
15. Las frutas
15. Las frutas
Por lo general tenemos una serie de ideas preconcebidas sobre lo que debe ser una fruta, un vegetal y en algunos casos un grano. Un ejemplo de estos conflictos aparece de forma bastante cómica en un caso sucedido en 1893 en el país de los abogados, Estados Unidos. El tomate por lo general es considerado un vegetal y se emplea como saborizante picante o salado para ciertas comidas, por otro lado, la idea intuitiva de las frutas es que son dulces y carnosas, sin embargo, desde el punto de vista biológico el tomate es una fruta. En 1893 la Corte Suprema de los Estados Unidos falló en el caso Nix contra Hedden, sobre si el tomate debía ser considerado un vegetal o una fruta. A pesar de lo estúpido que puede llegar a sonar el asunto era muy serio, pues trataba de dinero. Las frutas no pagaban impuestos, pero los vegetales si, reclasificar el tomate como fruta ahorraría dinero a muchas personas. La corte adoptó el significado común del tomate como un vegetal, pero como veremos en esta serie de artículos, el tomate es una fruta, así como muchas otras estructuras que normalmente no poseen este tipo de tratamiento.

Figura 15.1. Frutas secas. Los frutos no necesariamente deben ser dulces y carnosos, las vainas son un ejemplo de frutas secas.
Las frutas se definen botánicamente como el desarrollo de cualquier ovario y sus partes accesorias a partir de una flor hasta un estado maduro que contiene semillas. Bajo esta definición muchos así llamados vegetales incluyendo los tomates, las vainas de los verdes “ajotes”, los pepinos, las calabazas, y las vieiras entre muchas otras son en realidad frutas. Por otra parte, el nombre vegetal es un nombre común para varias estructuras modificadas de una planta como hojas “lechuga y repollo”, pecíolos de las hojas “apio”, hojas especializadas “cebolla”, tallos “patatas”, raíces “batatas”, raíces y tallos “remolacha”, flores y pedúnculos “brócoli”, botones de flor “alcachofa” entre otros.
Las vainas de semillas como las de los frijoles o las alverjas también son frutas, aunque carecen de la dulce carnosidad con que asociamos la palabra fruta, estas son frutas secas. Continuando con la definición botánica, las frutas son el producto del desarrollo de los ovarios de las plantas angiospermas mediante mecanismos relativamente complejos. La fertilización de los óvulos de una fruta debe realizarse en grupo, si algunos de los óvulos no son fertilizados, el ovario como un todo no se desarrolla, siendo cortado de la planta al secarse.

Figura 15.2. Las frutas son en general el desarrollo de la flor una vez que ha sido fertilizada, aunque se puede engañar a la flor para generar fruta sin semilla (YouTube).
Una vez que la mayoría de las semillas son fertilizadas y se han segregado las suficientes hormonas, el ovario crece acumulando nutrientes para formar una fruta carnosa. Los granos de polen cuando llegan en suficiente cantidad a la flor segregan hormonas que estimulan el desarrollo de la fruta. Los botánicos han logrado encontrar técnicas para engañar a la flor para convertirse en fruto sin semilla, como la administración de polen muerto. En algunos casos como en los cultivos de bananos las frutas pueden desarrollarse sin la fertilización mediante un proceso llamado partenocarpia, sin embargo, aunque una fruta sin semilla es más valiosa, para reproducir el cultivo se aplican técnicas asexuales, que hacen de estos cultivares más vulnerables a las plagas.
Anatomía de las frutas
La mayoría de las frutas maduras tienen cuatro regiones, el problema es que en ocasiones esas regiones se pueden fusionar y pueden llegar a ser muy difíciles de distinguir unas de otras. La piel de una fruta se denomina exocarpio, es una superficie dura cuya función es proteger a la fruta de invasores microscópicos como los hongos durante un lapso lo bastante largo como para que los agentes dispensadores de semilla hagan su trabajo.
En las frutas carnosas existe una región intermedia llamada mesocarpio, su tejido es carnoso y jugoso, poseen gran cantidad de nutrientes que sirven como aliciente para que animales ataquen la fruta y dispersen la semilla. Debajo del mesocarpio se encuentra en endocarpio que puede llegar a ser tan duro como una roca “como en las naranjas” o puede tener la misma textura del mesocarpio. En otras ocasiones es el endocarpio la estructura carnosa con nutrientes, mientras que el mesocarpio es seco y sirve como estructura de protección, esto sucede en las naranjas.
Colectivamente a estas tres regiones se las denomina pericarpio. La cuarta parte de la fruta se encuentra al interior del pericarpio, y no es otra cosa que su región más importante, la semilla. Las frutas secas tienen un pericarpio muy delgado por lo que en su totalidad parece más un estuche al cual se le da el nombre de vaina. Algunas frutas poseen pericarpios aún más simplificados consistiendo únicamente en el ovario y las semillas. La razón para esta diferenciación en las frutas es la relación de la planta con otras especies. Las frutas carnosas se han desarrollado como adaptación a la coevolución con herbívoros, es decir, es el cambio de una relación de forrajeo a una relación de simbiosis comensalista. En ese caso el herbívoro consume la fruta y transporta las semillas a otros lugares.


Figura 15.3. Partes principales de las frutas carnosas: exocarpo o epicarpo “cáscara externa”, mesocarpo, endocarpo y semillas. Las naranjas a la derecha no poseen semillas debido a un tratamiento genético para producirlas de ese modo.
Los primates son de los mayores dispersadores de semilla a través de frutas, y de todos, el ser humano es el más eficiente de todos, llegando incluso a afectar el curso evolutivo de muchos linajes de angiospermas. Dependiendo de la estructura del pericarpio se define si una fruta es carnosa o seca en su madurez, adicionalmente las frutas pueden realizar otros trucos. En frutos que no son consumidos por herbívoros la dispersión puede llegar a depender de que el excocarpio de la fruta se abra por si solo al secarse exponiendo las semillas en el proceso. Una fruta puede derivarse de un solo ovario o puede requerir de la mayoría de los ovarios de la flor sean fertilizados. Tradicionalmente muchas de estas características fueron empleadas por los botánicos para clasificar a las frutas. Dado el marcado interés en la taxonomía filogenética, muchos de estos sistemas de clasificación se han abandonado en favor de la clasificación molecular.

Figura 15.4. Del mismo modo que la flor es un truco para que los animales dispersen el polen, la fruta carnosa es un truco para que los animales transporten la semilla.
Frutas carnosas
En las frutas carnosas los mesocarpios “y en algunas especies también los endocarpios” son total o parcialmente carnosas, es decir poseen una estructura fibrosa llena de agua y nutrientes dispuestos no para el desarrollo de las semillas, sino para la atracción de animales que dispersarán la semilla. En las frutas carnosas simples la fruta se crea a partir de un solo pistilo, el ovario puede ser súpero, semiínfero o ínfero, el ovario mismo puede ser simple “formado por un solo carpelo” o compuesto “formado por varios carpelos”. En algunas ocasiones otras partes de la flor permanecen en la fruta como el cáliz o el mismo pistilo.
Drupas
El ejemplo más típico de una fruta carnosa simple es la drupa, la cual posee una sola semilla aislada por un endocarpio muy duro. Por lo general se forman de flores con ovarios súperos que contienen un solo carpelo con un solo ovario.

Figura 15.5. Ejemplos de drupas son albaricoques, cerezas, melocotones, ciruelas, aceitunas y almendras.
El mesocarpio no es en ocasiones tan obviamente carnoso, pero en los cocos el mesocarpio es bastante fibroso. La semilla de coco es hueca y contiene un endospermo acuoso comúnmente, aunque incorrectamente llamado leche de coco.
Pomos
Los pomos son frutas simples carnosas en donde la parte carnosa se desarrolla a partir de partes florales alargadas o del receptáculo del ovario. El endocarpio alrededor de la semilla puede tener una textura de papel o de cuero. Ejemplos de este tipo de frutas son las manzanas, las peras y los membrillos.

Figura 15.6. Las peras y las manzanas son ejemplos de los pomos.
En una manzana el ovario consiste en un núcleo con un tejido adyacente. El resto de la fruta se desarrolla a partir del tubo floral con poco tenido contribuido por el receptáculo. Algunos textos botánicos pueden clasificar a los pepos, algunas bayas y otras frutas derivadas de más de un ovario como frutas accesorias o frutas con tejido accesorio.
Bayas
Las bayas “berries” se desarrollan a partir de un ovario compuesto por varios carpelos y comúnmente contienen más de una semilla. En estas frutas la totalidad del pericarpio es carnoso y es difícil distinguir sus partes constituyentes. Pueden reconocerse varios tipos diferentes de bayas. Las bayas verdaderas, los pepos y los hesperidios. Las bayas verdaderas son frutas con una piel delgada y un pericarpio suave al madurar. Aunque la mayoría contiene más de una semilla existen excepciones notables que poseen una sola semilla. Ejemplos típicos de las bayas verdaderas son los tomates, las uvas, caquis, pimientos y berenjenas. Otras frutas que en ingles reciben el nombre de baya al finalizar “berries” no son bayas verdaderas. Algunas bayas se derivan de flores con ovarios ínferos por lo que otras partes de la flor contribuyen a la formación de la piel de la fruta. Esto puede evidenciarse por la permanencia de partes florales en la fruta como el cáliz, ejemplos: grosellas, arándanos, granadas y plátanos. Las bananas cultivadas por el hombre generalmente no tienen semilla cuando son criadas para la venta, pero las bananas silvestres poseen muchas semillas, así como un mesocarpio más seco, duro y hasta venenoso.

Figura 15.7. Algunos ejemplos de bayas son las uvas, las grosellas y los plátanos.
Los pepos
Los pepos son bayas con cortezas relativamente duras. Frutas de linaje de las calabazas incluyendo calabazas, pepinos, sandías, calabazas y melones son pepos.

Figura 15.8. El melón es un ejemplo de pepo.
Hesperidios
Los hesperidios son bayas con una piel apergaminada que contiene aceites. Numerosos crecimientos de las líneas del ovario crecen a costa del mesocarpio, estos forman sacos llenos de nutrientes y jugos frutales. Todos los miembros de la familia de los cítricos producen este tipo de fruta: ejemplos naranjas, limones, limones, toronjas, mandarinas y naranjas chinas.

Figura 15.9. Los cítricos dan fruto de tipo hesperidio
Frutas secas
En las frutas secas el mesocarpio y el endocarpio son muy secos al madurar la fruta. Por lo general se subdivide en dos grupos las frutas que se abren al madurar y las frutas que no se abren al madurar.
Frutas dehiscentes
Las frutas que se abren al madurar se denominan frutas dehiscentes, y sus subgrupos se clasifican de acuerdo al mecanismo de apertura de la fruta. En este grupo se encuentran frutos que generalmente no se clasifican como tales como las vainas de las semillas de los frijoles o las alverjas.
(a) Folículos. Las semillas foliculares se abren a lo largo de una sola sutura exponiendo las semillas en el interior. Ejemplos de este tipo de fruto son: delfinios aquilegias, algondoncillos “Figura 15.10” y peonias.

Figura 15.10. Algodoncillo.
(b) Legumbres. En las legumbres la fruta se abre a lo largo de los dos lados. Este tipo de fruta tiene representantes en cientos de especies de la familia de las legumbres “Fabaceae”. Ejemplos incluyen guisantes “alverjas Figura 15.11”, habas, garbanzos, lentejas, algarroba.
El fruto de la alverja al igual que las demás leguminosas es una vaina donde se encuentran las semillas en serie, esta vaina se abre a medida que se va secando exponiendo las semillas. Los cacahuetes o maní también son legumbres, pero presentan frutas atípicas que maduran bajo tierra. Las frutas típicamente son liberadas en la naturaleza gracias a la destrucción de la fruta realizada por las bacterias.

Figura 15.11. Arvejas.
(c) Silicuas. Las silicuas se abren a lo largo de los dos lados o costuras de la fruta, pero las semillas están encerradas en la región central, la cual es expuesta cuando las dos mitades de la fruta se separan de forma semejante a las leguminosas.


Figura 15.12. Vista de una silicua. Las silicuas y los siliclos son producidos por miembros de la familia de la mostaza “Brassiaceae” que incluye el brócoli, la col, el rábano, la bolsa de pastor, y el berro. Abajo, comparación entre una vaina de leguminosa “izquierda” y de silicua “derecha”.
A simple vista las silicuas son semejantes a las vainas de las legumbres "Figura 15.12 arriba", pero su diferencia interna es que la semilla no cuelga como tal, sino que está inmovilizada en compartimentos. Tales frutas cuando son al menos tres veces más largas que anchas son denominadas siliclos. En el siguiente modelo podemos apreciar la diferencia entre los frutos de una legumbre y un silicuo, en las legumbres las semillas cuelgas ya que no hay una matriz que las rodee y las inmovilice, cosa que si sucede en los silicuos.
(c) Cápsulas. Las capsulas son las frutas secas más comunes que se abren por si solas. Consisten en mínimo dos carpelos y se abren con una variedad de mecanismos. Algunas se abren a lo largo de las particiones entre los carpelos, mientras que otras se abren a través de cavidades en los carpelos llamados lóculos.

Figura 15.13. Cápsulas. La cápsula más famosa es la adormidera de opio Papaver spp.
Otras forman una capa delante de una punta expulsando las semillas a presión o forman una línea de poros a través de los cuales las semillas son expulsadas a medida que el fruto es sacudido por el viento. Ejemplo de estos son los lirios, orquídeas, amapolas, violetas y anturios.
Frutas indehiscentes
Las frutas secas que no se abren por si solas al madurar se denominan indehiscentes. En estas frutas las semillas se encuentran unidas fuertemente al pericarpio seco.
(a) Aquenios. En los aquenios la semilla única está almacenada en un hueco en el pericarpio. En la mayoría de las especies la semilla está levemente unida al pericarpio, por lo que este es fácil de desprender de la semilla. Los aquenios tienden a tener estructuras accesorias que incrementan su flotabilidad ya sea para desplazarse en el agua o en el aire. Ejemplos de este tipo de semilla incluyen las semillas de los girasoles, botón de oro, y el trigo sarraceno.

Figura 15.14. Las semillas de los aquenios flotan en el aire, fueron empleados como modelo de nave espacial por Carl Sagan en la serie original de “Cosmos, un viaje personal”.
(b) Nueces. Las nueces son frutas con una semilla única similar a los aquenios, pero generalmente son más grandes y el pericarpio es incomparablemente más duro y sólido. Las semillas se desarrollan unidas a una copa que limita sus movimientos. Ejemplos incluyen las bellotas, avellanas y nueces de nogal. Botánicamente hablando muchas nueces en el sentido popular "especialmente en el idioma inglés donde casi todos estos frutos terminan con el sufijo -nut que significa nuez" no son nueces verdaderas. Hemos visto anteriormente los cacahuetes que son típicas legumbres mientras que los cocos y almendras que son drupas. Las pecanas también son drupas cuya parte carnosa se seca después de que la semilla madura. Las nueces del Brasil son las semillas de una capsula muy grande, y los marañones son la única semilla de una sola drupa. Por lo cual resulta una tendencia general confundir semillas únicas muy grandes con frutos denominados nueces, otro ejemplo son las nueces de pistacho que son las semillas de una drupa.

Figura 15.15. Las nueces son semillas muy diversas.
(c) Granos. La reducción del pericarpio en algunos frutos puede llegar al extremo de hacerlo indistinguible del tejido de las semillas. En las semillas cuyo fruto se convierte en una parte más de la semilla se los denomina granos. Todos los miembros de la familia de los pastos “Poaceae” incluyendo el arroz, el maíz, el trigo, el centeno producen granos.

Figura 15.16. Los granos no son exactamente “la semilla” sino una fruta que rodea a una semilla.
(d) Granos. En las sámaras el pericarpio que rodea a las semillas se extiende hacia afuera formando un ala membranosa que ayuda en la dispersión de la semilla. En los maples las sámaras son producidas en parejas, pero en los cenizos, olmos, y en árboles del cielo son producidos individualmente.

Figura 15.17. Las samaras fueron elegidas por Neil D: Tyson como modelo de nave espacial en la serie “Cosmos una aventura de tiempo y espacio”.
(e) Esquizocarpios. Son frutas que se organizan como gemelos siameses, cada mitad se denomina mericarpio, es único de una familia de árboles polinesios llamados perejil “Apiaceae”. Miembros de esta familia incluyen el perejil, la zanahoria, el anís, la alcaravea, y el eneldo. Cuando se secan las frutas gemelas se rompen en dos fragmentos de una sola semilla llamados mericarpios.

Figura 15.18. Los esquizocarpios parecen dos samaras pegadas.
Frutas agregadas
Una fruta agregada es aquella que se deriva de una sola flor con muchos pistilos. Los pistilos individuales se desarrollan en pequeñas drupas u otras formas frutales, pero al madurar lo hacen en un grupo anidado en un único receptáculo. Ejemplos de frutas agregadas frambuesas, moras y fresas. En una fresa el receptáculo posee tiene forma de cono, la fruta se hace carnosa y roja mientras que cada pistilo se convierte en un pequeño aquenio en la superficie. En otras palabras, en las fresas las frutas aparte de ser agregadas también son compuestas.

Figura 15.19. Las frutas agregadas son incorrectamente denominadas bayas.
Frutas múltiples
Las frutas múltiples se derivan de muchas flores individuales producidas en un grupo anidado de floretas, es decir en una inflorescencia. Cada flor tiene su propio receptáculo, pero a medida que las flores maduran de forma separada en fruteletas se fusionan en una sola fruta agregada. Ejemplos de este tipo de frutas son moras, naranjas de Osage, piñas, y los higos. Las piñas al igual que las bananas tienen el potencial de desarrollarse por partenocarpia y en consecuencia no desarrolla semillas. Las flores individuales de los higos están unidas en torno a un eje carnoso y las fruteletas se fusionan en un único receptáculo con una apertura en la punta. Tal arreglo se denomina siconio.

Figura 15.19. Algunos ejemplos de frutas múltiples.
16. Las semillas
16. Las semillas
La semilla, simiente o pepita es cada uno de los cuerpos que forman parte del fruto que da origen a una nueva planta; es la estructura mediante la cual realizan la propagación de las plantas que por ello se llaman espermatofitas (plantas con semilla). La semilla se produce por la maduración de un óvulo de una gimnosperma o de una angiosperma. Una semilla contiene un embrión del que puede desarrollarse una nueva planta bajo condiciones apropiadas. También contiene una fuente de alimento almacenado y está envuelta en una cubierta protectora.
Anatomía de las semillas
Las semillas evolucionaron originalmente en el linaje de las angiospermas como estructura de latencia y resistencia, específicamente para resistir periodos de escases de agua. Christian de Duve en su libro Polvo Vital, la Historia de la Vida en la Tierra nos dice que la semilla de las plantas terrestres es el equivalente vegetal al huevo. Las razones de esto fueron que ambas estructuras hicieron a sus portadores exitosos y capaces de sobrevivir con mejores condiciones a través de la gran desertificación que tuvo el planeta durante la era geológica del pérmico, así como la más grande extinción en masa ocurrida durante esa misma era geológica. Las semillas en las angiospermas también son estructuras de latencia y resistencia, en esta serie de artículos discutiremos la anatomía de la semilla tipo “dicotiledónea” así como las propiedades de latencia y resistencia generales de las semillas.

Figura 16.1. Estructura eterna de una semilla dicotiledónea. La función de la semilla es proteger el embrión paralizado, y sostener su crecimiento una vez se detectan condiciones favorables, proceso denominado germinación (YouTube) (YouTube) (YouTube).
Partes externas de la semilla de las dicotiledóneas
Por lo general la semilla tipo que se emplea para ejemplificar las estructuras de una semilla es el frijol y sus semejantes, que son semillas con forma de riñón. La parte cóncava de estas semillas poseen una cicatriz blanca llamada hilium. El hilium Mareca el punto donde el óvulo que dio origen a la semilla estaba conectado con la pared del ovario.
Muy cerca del hilum se encuentra un poro diminuto llamado micrópilo. Si la semilla es colocada en el interior del agua por una hora o dos absorberá suficiente agua como para romper la pared de la semilla. Una vez que la pared de la semilla ha sido removida, las dos mitades llamadas cotiledóneas pueden distinguirse con claridad.

Figura 16.2. Tipos de semilla. Existen dos categorías, las monocotiledóneas de un solo lóbulo endospermio, y las dicotiledóneas con dos lóbulos endospermios.
El embrión de las dicotiledóneas
Los cotiledones se encuentran unidos justo debajo de la plúmula. Una parte muy corta del tallo arriba de los cotiledones se llama epicótilo, mientras que la parte del tallo debajo de la unión con los cotiledones se llama hipocótilo.
Ambas estructuras hacen parte del embrión, pero es difícil saber desde que punto se le puede llamar tallo y a que parte raíz. Por lo general solo la punta del hipocótilo recibe el nombre de radícula. Cuando la semilla germina lo que emerge en primera instancia es el hipocótilo, el cual se curva y adquiere una forma de garfio.
El garfio se entierra empujando los cotiledones fuera del suelo. Una vez que los cotiledones han emergido, el garfio se fortalece y sigue enterrándose.
Estructura de soporte vital en las semillas dicotiledóneas
Al inicio de la germinación la planta no hace fotosíntesis, de hecho, en muchas ni siquiera hay clorofila aun, entonces ¿de dónde sacan la energía y la biomasa para crecer? Si se observa la semilla misma, el espacio dedicado al embrión es minúsculo, el resto es ocupado por un tejido carnoso extremadamente nutritivo “por eso nos alimentamos de esas semillas”, en las leguminosas el tejido carnoso y nutritivo son los cotiledones.
Sin embargo, al igual que sucede con las frutas, el tejido nutritivo varia de posición entre los diferentes linajes, por ejemplo, en el maíz al tener un fruto tan íntimamente ligado a la semilla, es una parte del fruta llamada endiospermio la que proporciona la energía y la masa para el embrión en crecimiento.
Estructuras protectoras en las semillas dicotiledóneas
La plúmula es la estructura que dará lugar a las primeras hojas de la planta, mientras que la punta del hipocótilo formará la primera raíz. Ambas se encuentran rodeadas por estructuras tubulares llamadas caleoptilo y coleorrizo. Estas estructuras protegen los tejidos delicados en su interior en el momento de la germinación.

Figura 16.3. Germinación y crecimiento.
El coleoptilo forma el primer tallo o soporte rígido que levanta la punta de la planta, permitiendo de la plúmula se expanda. En el sentido inverso el coleorrizo se sigue enterrando hasta que termina de desarrollarse, esta no es una raíz verdadera ya que es abortiva, más bien es un ancla a la espera que las raíces secundarias emerjan de la semilla o de sí misma.
Germinación de la semilla
La germinación es el proceso de inicio o retoma del crecimiento de una semilla, lo cual depende de la intervención de factores bióticos y de factores abióticos internos y externos a la semilla. Para poder germinar una semilla debe poder ser viable. La viabilidad de una semilla implica que el embrión en su interior está vivo, de lo contrario, aunque los tejidos nutritivos estén intactos, la semilla no va a germinar. La pérdida de la viabilidad depende también del tiempo. Muchas semillas requieren factores como un periodo de latencia prolongado antes de poder germinar, otras semillas deben ser corroídas por bacterias o los ácidos estomacales de los herbívoros para poder germinar. En resumen, la germinación está mediada por factores mecánicos, químicos, fisiológicos y cronológicos.
Rompimiento de la latencia de una semilla por factores mecánicos
Algunas semillas poseen cascaras extremadamente duras que mantienen al embrión latente. El mejor método para romper el estado de latencia es mediante el proceso de escarificación “scarification → creación de cicatrices”, que requiere el rompimiento sutil de la concha de la semilla o sumergirla en ácido concentrado por unos segundos o minutos.

Figura 16.4. Escarificación. Algunas semillas deben rasparse para poder germinar, lo que imita en la naturaleza el efecto de las rocas, el agua y el viento.
Rompimiento de la latencia por factores bioquímicos
La activación de la semilla para su germinación puede requerir que su coraza externa sea degradada parcialmente por los ácidos digestivos u por las enzimas pépticas de los estómagos e intestinos de herbívoros que se alimentan de las frutas. Muchas de estas adaptaciones son tan externas, que las semillas de algunas especies jamás germinan si su herbívoro asociado no está presente (De Castro & Marraccini, 2006; Nogales et al., 2017; Otani, 2004; Traveset & Verdú, 2002; Zungu, 2014).
Rompimiento de la latencia por factores físicos no mecánicos
La latencia es un fenómeno común en muchas semillas y es el período en el que permanecen en estado de dormancia, incapaces de germinar incluso en condiciones favorables. Sin embargo, existen diversos factores físicos no mecánicos que pueden romper esta latencia y desencadenar el proceso de germinación. En este artículo, exploraremos en detalle cómo ciertos factores físicos, distintos al mecanismo tradicional de ruptura de la cubierta de la semilla, pueden estimular y acelerar la germinación.
A menudo, se piensa que el rompimiento de la latencia depende exclusivamente de la acción mecánica, como la abrasión o el corte de la cubierta de la semilla. Sin embargo, numerosos estudios han revelado que factores físicos no mecánicos, como la temperatura, la luz, el agua y la composición química del medio ambiente, también desempeñan un papel crucial en el proceso de germinación.
Exploraremos cómo la variación de la temperatura, la exposición a diferentes longitudes de onda de luz, la disponibilidad de agua y la presencia de ciertos compuestos químicos pueden influir en el rompimiento de la latencia y desencadenar la germinación. Además, examinaremos ejemplos concretos de plantas y semillas que han demostrado ser especialmente sensibles a estos factores físicos no mecánicos.
A través de este estudio, obtendremos una comprensión más profunda de los mecanismos subyacentes que permiten el rompimiento de la latencia por factores físicos no mecánicos, lo cual tiene importantes implicaciones en la propagación de plantas, la agricultura y la conservación de especies vegetales.
(a) Gases metabólicos. La germinación de una semilla está estrechamente ligada a la disponibilidad de oxígeno, ya que el proceso de respiración celular aeróbica es fundamental para activar el metabolismo y permitir el crecimiento del embrión. Sin embargo, existe una relación compleja entre el contenido de agua y la disponibilidad de oxígeno en la semilla durante el proceso de germinación.
Cuando una semilla se encuentra con una alta humedad relativa y está cargada de agua, se crea un ambiente propicio para la competencia entre el vapor de agua y el oxígeno molecular. El exceso de vapor de agua puede ocupar el espacio que normalmente estaría ocupado por el oxígeno, lo que dificulta su entrada a la semilla. Como resultado, el embrión no puede iniciar la respiración celular aeróbica y el metabolismo celular se paraliza nuevamente.
En términos simples, las semillas necesitan oxígeno para germinar. La disponibilidad adecuada de oxígeno permite que las enzimas responsables de la respiración celular funcionen correctamente, proporcionando la energía necesaria para activar los procesos metabólicos y permitir el crecimiento y desarrollo del embrión.
Es importante destacar que el equilibrio entre la humedad relativa y la disponibilidad de oxígeno es crucial para el rompimiento de la latencia y la germinación exitosa de las semillas. El control adecuado de estos factores es fundamental en prácticas agrícolas y técnicas de propagación de plantas, donde se busca promover una óptima relación entre la humedad y la oxigenación del sustrato para garantizar un desarrollo saludable de las semillas y plántulas.
(b) Temperatura. La temperatura es otro factor crítico que influye en el proceso de germinación de las semillas. Muchas semillas tienen requisitos específicos de temperatura para poder romper la latencia y comenzar el proceso de germinación. Las temperaturas extremas, tanto muy bajas como muy altas, pueden tener efectos adversos en el metabolismo celular y alterar la forma y función de las proteínas, lo que impide el desarrollo adecuado del embrión.
En general, se ha observado que la mayoría de las semillas requieren temperaturas dentro de un rango óptimo que oscila entre 0°C y 45°C para poder germinar. Fuera de este rango, el metabolismo celular se ve afectado y la germinación no ocurre. Los extremos de temperatura pueden desnaturalizar las proteínas y afectar su capacidad para catalizar las reacciones bioquímicas necesarias para el crecimiento y desarrollo de la planta.
La relación entre la temperatura y el porcentaje de germinación de las semillas a menudo se representa mediante una gráfica de campana de Gauss. Esto significa que hay una temperatura óptima en la cual la germinación es máxima, y a medida que la temperatura se aleja de este punto óptimo, el porcentaje de germinación disminuye. Para muchas semillas, la temperatura óptima se encuentra alrededor de los 22.5°C, que es una temperatura típica de las áreas tropicales donde muchas plantas con flores prosperan.
En el caso de las plantas de cultivo, la mayoría de ellas crecen mejor en un rango de temperatura que se sitúa entre los 20°C y los 30°C. Estas temperaturas permiten un metabolismo celular eficiente, una correcta absorción de nutrientes y una óptima fotosíntesis, lo que contribuye al crecimiento saludable y productivo de las plantas cultivadas.
Es fundamental comprender y controlar la temperatura durante la germinación de las semillas, especialmente en el contexto de la agricultura y la producción de cultivos. Los agricultores deben garantizar que las condiciones de temperatura sean adecuadas para maximizar el potencial de germinación y establecimiento de las semillas, lo que contribuye a una mayor productividad y calidad de los cultivos.
(c) Luz. La luz desempeña un papel crucial en el proceso de germinación de las plantas, aunque su influencia puede variar significativamente dependiendo de la especie. Cada planta está adaptada a condiciones específicas de luz, lo que significa que algunas especies requieren ciertas intensidades de luz para germinar adecuadamente, mientras que otras necesitan condiciones de sombra.
En el caso de las semillas de ciertas variedades de lechuga, por ejemplo, la germinación se ve favorecida por la presencia de luz. La luz actúa desactivando inhibidores del desarrollo presentes en las semillas, lo que permite que el embrión comience a crecer y se inicie el proceso de germinación. Estas variedades de lechuga necesitan una exposición directa a la luz para su correcto desarrollo.
Por otro lado, algunas semillas, como las de la amapola de California, requieren condiciones de sombra para germinar. En este caso, la luz estimula la secreción de inhibidores del crecimiento, lo que retarda la germinación y permite que la planta se establezca en áreas sombreadas. Estas especies están adaptadas a entornos con menor intensidad de luz y necesitan estas condiciones para desencadenar su proceso de germinación.
La respuesta de las semillas a la luz durante la germinación es un mecanismo de adaptación que les permite aprovechar al máximo su entorno y asegurar su supervivencia. La sensibilidad a la luz puede variar incluso dentro de una misma especie, lo que refleja las diferentes estrategias de crecimiento y reproducción de las plantas en su hábitat natural.
En el ámbito de la agricultura, comprender los requisitos de luz de las semillas es esencial para optimizar la germinación y el crecimiento de los cultivos. Los agricultores deben proporcionar las condiciones adecuadas de luz, ya sea exposición directa a la luz solar o sombreado controlado, según las necesidades de las diferentes especies cultivadas. Esto garantiza un desarrollo saludable de las plantas y un rendimiento óptimo en la producción agrícola.

Figura 16.5. Valle de Cocora Quindío, Colombia. Los bosques tropicales son el ambiente ideal para la germinación de las plantas con flor debido a su disponibilidad de agua y temperatura estables durante todo el año.
Las frutas y cáscaras pueden inhibir la germinación de las semillas
La latencia puede ser inducida y mantenida químicamente mediante sustancias producidas por la cáscara de la semilla o por la fruta donde están contenidas. Muchas plantas del desierto poseen inhibidores en las cascaras de sus semillas. Estas semillas tienen que ser lavadas antes de poder ser plantadas. Esto implica que los inhibidores actúan como una señal de la existencia de agua lluvia, cuando la lluvia lava los inhibidores la semilla puede crecer. Esto evita que el embrión germine en un ambiente con poca agua, característico de los desiertos.
Las frutas carnosas como las manzanas, las peras, los cítricos, los tomates y muchas otras contienen inhibidores que previenen la germinación de la semilla al interior del fruto. Esto evita que un embrión germinado muera asfixiado o consumido por un herbívoro. Una vez que las semillas han sido removidas de la fruta y lavadas adecuadamente “algunas incluso sometidas a tratamientos con ácidos” pueden germinar rápidamente. Los embriones de algunas semillas pueden encontrarse en un estado muy simple de desarrollo cuando la cascara se rompe. Las plantas del desierto deben germinar rápidamente después de llover, pero mantenerse en espera lo más posible cuando no hay agua disponible.

Figura 16.6. Disponibilidad de agua. Las semillas de las plantas del desierto tienen bloqueada su germinación hasta asegurarse el contacto con el agua.
Postmaduración de una semilla
Las semillas no madurarán, aunque la cáscara se hubiese roto hasta que el embrión expuesto no se halla desarrollado lo suficiente con la ayuda de los nutrientes de los cotiledones o el endospermo. A tal proceso se le da el nombre de postmaduración. Muchas plantas leñosas “generan maderas” en las regiones templadas requieren de estimuladores para iniciar su desarrollo embrionario. Los parámetros que determinan el inicio de la postmaduración son la humedad relativa y el contenido de aceites interno, la naturaleza de la cáscara de la semilla y la temperatura. La postmaduración es arrestada por una semilla con poca disponibilidad de agua, por lo que se necesita de cierto valor crítico de humedad para que la semilla reinicie su desarrollo embrionario. Sin embargo, lo contrario también es cierto, una humedad relativa muy alta evita que la postamuración inicie “lo cual evita que una semilla germine cuando se encuentra en un ambiente acuoso que la asfixiaría.
El agua
Normalmente las semillas no se desarrollan hasta que la humedad relativa es alta y la temperatura es baja “una estación lluviosa”, usualmente este período debe durar entre unas 4 a 6 semanas. La latencia en tales semillas puede romperse artificialmente colocándolas en un refrigerador, preferiblemente en arena mojada por unas cuantas semanas. Debido a que el embrión se alimenta del endospermio o de los cotiledones, los retoños no requieren luz para crecer. Una semilla puede absorber tanta agua como para aumentar su propio peso unas 10 veces antes de iniciar su desarrollo embrionario nuevamente.

Figura 16.7. Postmaduración. El control de la postmaduración es un parámetro crítico a la hora de almacenar correctamente las semillas para la plantación de la cosecha de años venideros.
Después de que la semilla ha absorbido suficiente agua, el citoplasma de las células embrionarias se rehidrata, lo cual altera todas las condiciones fisicoquímicas celulares. Las proteínas que son las sustancias encargadas de realizar las funciones son muy sensibles a las condiciones fisicoquímicas. La forma de las proteínas está condicionada por el pH o acidez y este parámetro, así como otros que también afecta su forma dependen de la cantidad de agua. Sin agua las proteínas adquieren formas no funcionales que paralizan el metabolismo de la célula hasta un mínimo vital. La presencia de agua reactiva las proteínas y reinicia el metabolismo activo. En algunas plantas, el metabolismo mínimo vital es la respiración anaeróbica, con este metabolismo la semilla reinicia sus actividades hasta poder realizar la respiración aeróbica nuevamente. Una vez el metabolismo está en marcha, el ciclo celular de las células da inicio, y en correspondencia la mitosis que permite el crecimiento del embrión vegetal.
Longevidad y latencia de las semillas
Existen leyendas de internet que dice que algunas semillas han podido ser recuperadas de antiguas tumbas egipcias o americanas y mantienen su viabilidad después de miles de años. Existe evidencia de que ratos y otros roedores han transportado las semillas a dichos lugares, dejándolas abandonadas. Sin embargo, un caso muy convincente ocurrió en una tumba japonesa, esto debido a que la planta producida por dicha semilla es diferente de todas sus congéneres en el número de pétalos, ya sea producto de la evolución de la población o de alteraciones en el desarrollo genético durante la latencia prolongada.
Viabilidad de la semilla, condiciones para su conservación
La viabilidad se define como el potencial de una semilla para reiniciar su desarrollo una vez que las condiciones bióticas y abióticas para su germinación se han cumplido. Los periodos de latencia en los cuales la semilla retiene su viabilidad cambian mucho dependiendo de la especie y del individuo.

Figura 16.8. Longevidad de las semillas. Como cualquier carácter evolutivo darwiniano, la viabilidad es muy variable al interior de una misma población, por lo que el tiempo de viabilidad disminuye de forma gradual entre un grupo de semillas. Las curvas de viabilidad contra tiempo en meses, muestra como la viabilidad de cuatro especies disminuye de forma gradual. Las germinaciones más antiguas han sido reportadas en la tundra ártica con semillas de más de 10.000 años de antigüedad que mantienen su viabilidad (YouTube).
Las semillas de especies de sauces, álamos, orquídeas, y plantas de te permanecen viables por unos pocos días o algunas semanas como máximo, sin importar que posteriormente las condiciones ambientales sean favorables no germinarán. El periodo de latencia en el que la semilla retiene su viabilidad es mucho más prolongado en la mayoría de las especies, como mínimo de algunos meses mientras que otras pueden extenderse por años siempre y cuando se mantengan condiciones de baja temperatura y humedad.
Almacenamiento y venta de las semillas
La normatividad de varios países determina que las semillas empacadas para la venta deben tener un registro en sus paquetes que informe la fecha de producción y el posible periodo de tiempo promedio en el cual las semillas conservan si viabilidad. Las semillas del linaje de las calabazas “calabaza, melón, pepino” retienen un porcentaje relativamente alto de su viabilidad por muchos años, mientras que los miembros del linaje de los lirios “cebolla, puerro, cebollino” retienen su viabilidad en un rango de 2 a 3 años.
El 30% de las semillas de trigo almacenadas apropiadamente pueden retener su viabilidad después de unos 30 años, algunos pastos pueden retener su viabilidad por periodos mucho más prolongados. El almacenamiento de la semilla requiere de negarle los requerimientos para la postmaduración, una humedad relativa baja, una temperatura muy baja que impida que las bacterias la ataquen, un bajo nivel de oxígeno que impida la activación del metabolismo y bajos índices de luz solar.
Semillas vivíparas
Algunas especies de plantas con flor ya sean dicotiledóneas o monocotiledóneas no poseen un estado de latencia como tal. Esto implica que el embrión jamás detiene su crecimiento llegando a germinar al interior del fruto, este fenómeno es denominado viviparía vegetal. El mangle rojo, un árbol tropical asociado a los ecotonos costeros de agua dulce-salada poseen frutas con semillas únicas. El embrión sigue creciendo incluso hasta que la nueva planta germina, aunque el fruto aún se encuentre colgado del árbol.


Importancia evolutiva de los periodos de latencia en las semillas
La latencia prolongada de las semillas es uno de los mejores métodos de los seres vivos para tolerar cambios ambientales catastróficos que conllevan a las extinciones en masa. Es probable que tanto para las extinciones en masa del pérmico como del cretáceo la semilla hubiese jugado un papel fundamental en la supervivencia de las gimnospermas y las angiospermas. Como la mayoría de las innovaciones evolutivas, la semilla ya había aparecido tiempo antes de la extinción pérmica, y muchos autores sostienen que las semillas con largos periodos de latencia le permitieron a las angiospermas sobrevivir y recuperarse muy rápido después de la extinción en masa del cretáceo.
17. Dispersión de las semillas
17. Dispersión de las semillas
Los vegetales se caracterizan por su falta de movilidad, y aun así son capaces de dispersarse de un lugar a otro al reproducirse. Algunas especies se encuentran confinadas en unos pocos lugares del mundo, siendo tan endémicos que sus especies no abarcan más de unas pocas hectáreas; mientras que otras especies son tan eficientes en su dispersión que sus descendientes han llegado virtualmente a todos los continentes. ¿Por qué algunas orquídeas son tan raras, mientras que los pastos son tan comunes?
Las respuestas a estas preguntas recaen en las adaptaciones de las plantas para poder aprovechar a los factores bióticos y abióticos en el transporte de sus semillas de un lugar a otro. De hecho, cuando una semilla es transportada por un factor biótico como los animales, a esta relación se la denomina foresis “movimiento o transporte” y es un tipo de simbiosis, sin embargo, los factores abióticos como el viento o el agua también pueden mover las semillas de un lugar a otro. Cinco métodos principales para la dispersión de las frutas y las semillas han sido propuestos, cabe indicar que el 5 es una subclase del 2 dado que los seres humanos somos animales, pero igual los clasificaremos aparte por la tradición: dispersión por: (1) viento, (2) animales, (3) agua, (4) propulsión y (5) por seres humanos.

Figura 17.1. Medios de dispersión de semillas. Algunos mecanismos para la dispersión de las semillas, de izquierda a derecha: viento, animales, agua, expulsión mecánica y por humanos.
Viento
El viento es una corriente de aire que con la suficiente intensidad puede llegar a mover con facilidad estructuras pequeñas como las semillas. En términos de un ecosistema, el viento es un factor abiótico, es decir un fenómeno que no está vivo pero que afecta a los seres vivos de múltiples formas, lo que en última instancia es uno de los factores que componen a la selección natural.
La Figura 17.2 muestra una simulación computarizada del efecto del viento sobre la dispersión de las semillas. Si el árbol es el centro, las semillas caerán con más probabilidad cerca del árbol. Sin embargo, este patrón aleatorio puede aumentar su aleatoriedad, si las semillas son producidas con una variedad de modificaciones que les permiten aprovechar la fuerza del viento y movilizarse a grandes distancias.

Figura 17.2. Probabilidad de germinación contra espacio. La probabilidad de germinación es inversamente proporcional a la distancia del foco de producción de semillas.
Adaptaciones al viento
Las sámaras son frutos con una adaptación bastante evidente para la dispersión por el viento, con este método las sámaras pueden movilizarse cerca de 10 kilómetros de su planta madre y hasta poder encontrar un lugar para germinar.

Figura 17.3. Algunas adaptaciones de las semillas para la dispersión por viento (YouTube).
Números de distancias semejantes han sido estimados para las semillas de los botones de oro y los girasoles “Ranunculaceae “imagen siguiente” y Asteraceae”. Las frutas poseen estructuras semejantes a plumas, mientras que en los sauces las frutas están rodeadas por una estructura algodonosa que ayudan a mejorar la flotabilidad en una corriente de aire. Al igual que cualquier especie biológica, la mayor parte de las frutas caen en lugares malos o son devoradas por los depredadores, y solo unas pocas logran sobrevivir. Otro aspecto de la dispersión por viento es que es muy aleatoria y la mayoría de las frutas son dispersadas en un rango muy cercano de la planta madre.
A parte de la estructura de la fruta, las semillas individuales al ser lo bastante pequeñas pueden ser arrastradas por el viento sin necesidad de poseer una costosa estructura secundaria que incremente su flotabilidad. Las flores de algunas orquídeas y brezales producen semillas tan pequeñas que son comparables en tamaño, pero y densidad al polvo. Los árboles de catalpa y jacaranda la semilla posee modificaciones que incrementar su flotabilidad en una corriente de aire, en este caso se trata de membranas alares como en la sámara “que es una fruta”.
La nave de la imaginación
Las fruteletas de la inflorescencia del diente de león son tal vez el ejemplo más típico de una semilla con una estructura accesoria que le permite incrementar su flotabilidad en una corriente de aire tan tenue como una pequeña briza. Resulta interesante que en ambas series de Cosmos emplean semillas y frutas transportadas por el viento como el templado para sus naves de la imaginación. Lo anterior tal vez es para representar que la imaginación es la semilla que sirve para dispersar el conocimiento humano, y que las propias naves espaciales serán las semillas que transporten a la humanidad a nuevos mundos de manera simultánea.


Figura 17.4. La nave de la imaginación. La “nave de la imaginación” de Carl Sagan era un diente de león; mientras que la nave de Tyson aparenta tener la estructura de una sámara única. La idea es que las naves espaciales serán la semilla que disperse la vida en el universo.
Agua
El agua (YouTube) constituye el otro gran factor abiótico al cual las plantas se han adaptado para la dispersión de sus frutos y/o semillas. El modo más simple es que la planta no tenga una adaptación como tal, y sea simplemente el efecto de una corriente de agua lo bastante potente como para romper partes de la planta y conducirlas a otras partes del planeta. Algunas frutas contienen aire atrapado en su interior, lo cual disminuye su densidad relativa y en consecuencia pueden flotar con mucha mayor facilidad en el agua. Los mecanismos que permiten atrapar aire incluyen sacos que le permiten a las semillas flotar como en la fruta de las ciperáceas. Otras semillas poseen materiales cerosos en la superficie de las semillas, las cuales previenen temporalmente que la semilla absorba agua mientras flotan. Otras plantas poseen un pericarpio duro y esponjoso que absorbe agua lentamente desviándola del delicado pericarpio. En caso de que el agua sea salada solo es cuestión de tiempo para que cualquier adaptación falle y la sal mate al embrión.
Los animales
Al contrario de las gimnospermas donde tanto la fertilización como la dispersión de las semillas obedecen a fenómenos relativamente aleatorios como el viento y el agua, las plantas con flor han desarrollado métodos para aprovechar sus propios depredadores en su beneficio (YouTube) (YouTube). Lo anterior implica que las relaciones ecológicas son mucho más complejas de lo que podría esperarse, ya que a pesar de que muchos herbívoros afectan negativamente la aptitud reproductiva de las plantas al comerse sus semillas, también actúan como potenciales agentes de dispersión de semillas y en consecuencias de colonización de nuevos territorios, lo cual en ultimas beneficia la aptitud reproductiva de una especie.
Las adaptaciones de las frutas y semillas para la dispersión por animales son numerosas. Aves, mamíferos y artrópodos actúan como diseminadores. Se puede llegar a extremos en que la semilla no puede germinar a menos que entre en contacto con un animal, si se extingue al animal que dispersa la semilla también lo hace la planta.
Ingestión simple
Los frutos carnosos se convierten en el mecanismo más evidente en una relación mutualista para la dispersión de una planta y uno de sus depredadores en la relación de forrajeo. Muchas aves y mamíferos al alimentarse de las frutas que son estructuras altamente nutritivas no son capaces de digerir la semilla, la cual pasa intacta a través del tracto gastrointestinal, siendo expulsada con las heces en un lugar alejado de la planta madre.
Esto es un ejemplo de una relación en red en los ecosistemas, donde las relaciones ecológicas son muy complejas. La fruta no es meramente un agente pasivo en esta relación, algunas frutas poseen agentes químicos laxantes que aceleran el paso de la semilla a través del tracto gastrointestinal en caso de que exista el potencial de daños por el paso a través del tracto gastrointestinal. En las aves especialmente estos laxantes pueden inducir a la expulsión de la semilla en tan solo 15 minutos. Esto se debe a que las aves poseen mollejas, órganos especializados en triturar semillas.
Ingestión especializada
Por otra parte, los mamíferos y varios reptiles no poseen órganos especializados para triturar semillas y por lo tanto el periodo entre la ingesta de la fruta y la explosión de la semilla puede rondar las 24 horas. Otro aspecto importante de esta relación es que las semillas en algunos casos se vuelven dependientes de su paso a través de un tracto gastrointestinal que debilite lo suficiente la cascara de la misma para poder germinar.

Figura 17.5. Ingestión especializada. Sin los lémures, muchas semillas endémicas de Madagascar se extinguirían.

Figura 17.6. Ingestión especializada 2. Igual que con los lémures y muchos frugívoros, muchas semillas deben pasar por las enzimas y jugos digestivos de un animal para activarse y germinar.
O lo que es lo mismo, si se le retira a estas especies de planta su depredador “herbívoro” la planta también se extingue pues pierde su capacidad reproductiva. Los lémures, por ejemplo, son primates que dispersan de forma obligatoria varias semillas endémicas de Madagascar, su extinción significaría la muerte de dichas especies de plantas En las tortugas gigantes de los Galápagos, por ejemplo, las semillas de varias especies de plantas deben pasar cerca de dos semanas o más hasta ser expulsadas. La semilla no germinará a menos que haya sido digerida parcialmente en el tracto digestivo de una de estas tortugas. Ejemplos de esto han sido reportados en otras especies como primates.
Abandono de herbívoros
Algunas semillas son recolectadas y almacenadas sistemáticamente por forrajeadores como los roedores y las hormigas. Entre los principales roedores con este comportamiento se cuentan a los ratones de campo y las ardillas. Dado que el almacenamiento de la semilla es bajo tierra, y no es raro que el pequeño cerebro de estos animales olvide el lugar donde escondieron todas sus semillas, lo cual les permite germinar. Las aves también son capaces de realizar esto como en el caso de los arrendajos azules, pájaros carpinteros y otras aves que abandonan frutas duras como las nueces a mitad del vuelo. Las hormigas también transportan algunas semillas, más para alimentarse de sus apéndices, dado que la concha de la semilla impide el acceso de estos animales al embrión en desarrollo. La fruta como tal queda almacenada y abandonada hasta que germina.
Adhesión mecánica
La adhesión mecánica es completamente involuntaria para el animal y no le confiere ningún beneficio, por lo cual se la puede clasificar como una foresis comensalista. Existen muchas formas en que una fruta o semilla puede adherirse a un animal.


Figura 17.7. Transporte mecánico. Otras semillas son transportadas como polizones de los animales por medio de garfios.
En algunas frutas y semillas otro factor abiótico como el lodo sirve como pegamento para adherirse al plumaje de las aves o al pelaje de los mamíferos que avanzan en su territorio. Muchas frutas y semillas poseen adaptaciones fisiológicas para la adhesión independientes del lodo como ganchos. Las semillas de la planta unicornio “imagen siguiente” poseen dos extensiones curvas de unos 15 centímetros de largo, las cuales se enganchan al largo pelaje de los ciervos y otros animales grandes.
Las lineanas gemelas y la linaza poseen frutos que han continuado con la idea del lodo, pero en lugar de depender de este como pegamento, ellas producen sus propios apéndices pegajosos que les permiten la adhesión. Otras frutas son un poco más molestas como en el caso del vino punzante “Tribulus terrestris” cuyas púas penetran en el cuero, tal como podemos apreciar en la siguiente imagen. Otras plantas emplean aceites con feromonas que atraen a los artrópodos solo para quedarse pegadas a estos por ciertos periodos de tiempo.
Autodispersión
La dispersión por aire puede verse apoyada por mecanismos fisiológicos de las plantas que expulsan las semillas con gran potencia lejos de la planta madre (YouTube) (YouTube). Las frutas de algunas leguminosas como los miembros del grupo de las mimosas o no me toques “Mimosa pudica” eyectan las semillas de forma mecánica, algunas veces con fuerza considerable. Por ejemplo, la separación de fruto de la avellana puede expulsar las semillas 12 metros alrededor de la planta madre. En la familia de las calabazas las semillas son liberadas como si fueran el polvo de un geiser.
Intervención humana
Los seres humanos también son importantes dispersadores de fruto y semilla. Nuestra afición por las frutas se debe a que al igual que los primates del viejo mundo dependemos de ellas para la obtención de nutrientes esenciales como algunas vitaminas. Esta afición por las frutas ha hecho que a lo largo de la historia los viajeros y comerciantes buscaran en los nuevos territorios frutas exóticas que pudieran acoplarse a los métodos de producción más eficientes de sus respetivos periodos históricos. Uno de los primeros productos que llamaron la atención de los españoles conquistadores fue la piña, lamentablemente en el siglo XV no existían medios para preservar las frutas en un viaje de dos meses en alta mar. Algunas plantas no pueden reproducirse sin nuestra ayuda, esto se debe a que el proceso de domesticación ha sido tan fuerte que sin nuestro cuidado la semilla no puede salir del fruto. El transporte de semilla y fruto planificado también acarrea el no planificado, muchas veces el transporte de semillas de especies invasoras afecta la diversidad de regiones apartadas, destruyendo las especies nativas.
18. Ciclo de vida de las angiospermas
18. Ciclo de vida de las angiospermas
Ya hemos visto en detalle muchos de los pasos del ciclo de vida de las angiospermas, ahora vamos a realizar un resumen para colocar las cosas en orden a manera de resumen.

Figura 18.1. Ciclo de vida de las angiospermas.
(YouTube) El ciclo de vida comienza con el desarrollo del esporofito (1) “en las plantas dioicas” o de los esporofitos “en las monoicas”. Recuerde que una planta monoica solo produce flores con un solo sexo.
(YouTube) Cuando se da la maduración sexual las plantas generan sus estructuras sexuales que denominamos flores (2). En las flores dioicas se generan tanto estambres “masculinos” (4) como pistilos-carpelos “femeninos” (3). Hay que recordar que las flores no son siempre conspicuas y algunas carecen de pétalos coloridos.
(YouTube) El estambre induce la meiosis (5) produciendo la microespora (6) mejor conocida como grano de polen que es transportado al pistilo por múltiples factores bióticos y abióticos. Una vez en la punta o estigma del pistilo el polen crece (7) desarrollando el tubo polínico (8).
(YouTube) Simultáneamente al interior de la semilla inmadura se encuentra la megaespora (9) la cual se desarrolla (10) en un gametofito (11), a diferencia del gametofito femenino de las gimnospermas, el gametofito de las angiospermas es minúsculo y está compuesto solo por 7 células, y solo 2 de estas se emplean en el proceso reproductivo.
(YouTube) La polinización se define como el momento en que el polen alcanza el pistilo (12), iniciando su crecimiento, cuando el tubo polínico alcanza la semilla libera dos células espermáticas que fecundan al óvulo y a dos núcleos haploides en el centro del gametofito, proceso denominado fecundación doble (13). El embrión surge del cigoto (14) que es diploide 2n, y de la segunda fertilización surge un endospermo triploide 3n.
(YouTube) Posteriormente el endospermo nutre al embrión en crecimiento que se diferencia en el embrión como tal y en el cotiledón o cotiledones a través de una plúmula en medio. En las dicotiledóneas, los cotiledones absorben lo que queda del endospermo. Una vez la semilla crece y ha perdido agua su cascara se endurece (15).
(YouTube) Simultáneamente la fruta crece (16), en caso de una fruta carnosa toda el área alrededor de la semilla es rodeada por un tejido altamente nutritivo que facilita procesos de simbiosis y dispersión, otras frutas llamadas secas simplemente dejan que los factores abióticos hagan su trabajo. Cuando la semilla es liberada pasa por un periodo de latencia que se rompe cuando las condiciones se tornan favorables y comienza el desarrollo (17) de un nuevo esporofito (1).
Generación esporofítica
Al igual que pasa con las gimnospermas, en las angiospermas es el esporofito quien domina, a tal punto que los gametofitos maduros se deben desarrollar en su interior.
Desarrollo de fruta y semilla
Del cigoto surge el embrión y de la célula triploide se desarrolla un endospermo, que es un tejido nutritivo para la semilla en desarrollo. Cuando el o los embriones inician su desarrollo la flor cambia, pierde todas sus hojas especializadas y se hincha para formar el fruto, que puede ser seco o carnoso. El fruto carnoso posee tejidos extra para alimentar a los animales que ayuden a dispersar la semilla, mientras que los frutos secos se abren solos y dejan salir semillas que se dispersan por otros medios. La semilla es una estructura de resistencia que porta un embrión que ha paralizado momentáneamente su desarrollo a la espera de mejores condiciones ambientales.
Semilla
La semilla, simiente o pepita es cada uno de los cuerpos que forman parte del fruto que da origen a una nueva planta; es la estructura mediante la cual realizan la propagación de las plantas que por ello se llaman espermatofitas (plantas con semilla). La semilla se produce por la maduración de un óvulo de una gimnosperma o de una angiosperma. Una semilla contiene un embrión del que puede desarrollarse una nueva planta bajo condiciones apropiadas. También contiene una fuente de alimento almacenado y está envuelta en una cubierta protectora.
Las semillas de las angiospermas vienen protegidas por varias capas, las cuales pueden ser muy diferentes de las propias semillas en forma de vainas y frutos, o estar reducidas a cascarones poco diferenciables de las semillas propiamente dichas como sucede con muchos granos de pastos. Todas las angiospermas tienen flores (aunque no siempre corresponden a la idea común que todos tenemos de una flor), que producen semillas encerradas y protegidas por la pared del ovario (carpelos) que, posteriormente, se convierte en fruto. Los mecanismos de dispersión son diversos, y puede consultarlos en el capítulo 11 de la presente unidad. Cabe resaltar que lo que se encuentra en el interior de una semilla es un embrión altamente desarrollado y no un óvulo, además de una gran cantidad de tejido nutritivo que ayuda a que esta sirva como propágulo de resistencia.
Esporofito
A diferencia de lo que sucede con las gimnospermas tipo, el esporofito de las angiospermas puede ser muy diverso en forma, estilo de vida, adaptaciones nutricionales y expectativa de vida. Pueden ser plantas herbáceas, arbustivas o arbóreas.
La diversificación que sufrió este grupo es asombrosa. En el registro fósil aparecen a principios del Cretácico (hace unos 130 millones de años), momento a partir del cual aparecen enormes cantidades de fósiles de especies muy diversas, como si hubieran aparecido con toda su diversidad de golpe, lo que Darwin llamó en su momento un "abominable misterio". Aun hoy cerca del 90% de las plantas terrestres pertenecen a este grupo. Con unas 257.000 especies vivientes (Judd et al. 2002), las angiospermas son las responsables de la mayor parte de la diversidad en espermatofitas, en embriofitas y en viridofitas.
Así como posee muchas especies, este grupo se caracteriza por una enorme diversidad de hábitos, y haber ocupado prácticamente todos los nichos ecológicos posibles. Hay plantas arbustivas y herbáceas, las hay terrestres y acuáticas, se encuentran tanto en los desiertos como en los pantanos, en el nivel del mar como en lo alto de las montañas. Su diversidad de especies es mucho más alta en zonas tropicales y húmedas (alrededor del 60% de las especies son de zonas tropicales y un 75% tiene un crecimiento óptimo en climas tropicales), donde dominan completamente el paisaje, y va disminuyendo su número hacia las latitudes altas, llegando a poseer una representación empobrecida en las floras más frías como la de la tundra (que todavía hoy está dominada por las coníferas). Los miembros de este grupo son la fuente de la mayor parte de los alimentos consumidos por el hombre, así como de muchas materias primas y productos naturales. El grueso de la alimentación mundial procede de sólo quince especies.
El esporofito es quien se encarga de la mayoría de las funciones no reproductivas e incluso de muchas funciones reproductivas, afrontando la mayor parte de la selección natural, como enfrentar a competidores, depredadores y parásitos. Al igual que los esporofitos de las gimnospermas, las angiospermas poseen esporofitos complejos con partes altamente especializadas:
(a) la raíz, es la parte que se encuentra debajo de la tierra. Su función es sujetar la planta y absorber las sales minerales y el agua del suelo.
(b) Transporta agua y nutrientes de las raíces a las hojas y el alimento producido por estas al resto de la planta, para esto emplea un sistema vascular dividido entre un floema y un xilema. Muchas angiospermas poseen vasos xilemáticos además de traqueidas, carácter derivado dentro del grupo. En los vasos, el agua puede fluir sin necesidad de atravesar una membrana, lo que los vuelve muy eficientes en el transporte de fluidos dentro del esporofito pero probablemente también más propensos a recibir daño (en especial por embolias de aire) cuando están sujetos a estrés hídrico. El floema de las angiospermas difiere del de todas las demás plantas en que los elementos del tubo criboso (que son células vivas pero sin núcleo, encargadas del transporte de azúcares) están acompañadas por una o más células acompañantes, que nacen de la misma célula madre que el elemento.
(c) las hojas, contiene el pigmento verde llamado clorofila, que absorbe la energía de la luz solar y la usa para convertir el dióxido de carbono en oxígeno. También absorben y difunden agua y gases. Son muy variadas en forma, color y tamaño. Su riqueza es absoluta en la Tierra. Las hojas pueden experimentar modificaciones secundarias para resistir al clima, los depredadores o incluso para cazar alimento.
(d) las flores que se encargan de la fabricación de esporas, frutos y semillas, adicionalmente muchas de las partes de los frutos y las semillas se fabrican con partes del esporofito.
(e) Flor Es la versión del macroestrobilo y el microestróbilo, pero muchísimo más especializado. Al igual que con los estróbilos, las flores se especializan tanto en la producción de esporas como en la producción de semilla, y además también se da el hecho de que las macroesporas femeninas se encuentran atrapadas. Las flores pueden ser monoicas con un solo tipo de estructuras sexuales, o dioicas, presentando ambos tipos, con una alta posibilidad y tendencia a la autofertilización, lo cual ha sido una clave para su alta diversificación y evolución rápida por medio de los procesos de poliploidías, que conlleva a especiaciones instantáneas.
Generación gametofítica
La generación gametofítica es microscópica, al ambiente solo se liberan las microsporas masculinas, conocidas como granos de polen.
Microspora o grano de polen
El grano de polen es el propágulo de tipo microespora, es producida por las anteras, que es el esporangio masculino producido por las flores. Las microsporas son liberadas al viento o emplear otros mecanismos como simb iosis mutuañlistas con artrópodo, aves y mamíferos para aumentar la probabilidad de polinización, y disminuir la energía invertida en producir muchos granos de polen como ocurre con las gimnospermas.
Estas microesporas deben llegar al estigma de una flor de la misma especie para germinar. Varios granos pueden germinar en un mismo estigma, lo que conlleva a una carrera por la fertilización. El proceso de transporte del polen de una flor a otra se conoce como polinización, y es un evento separado temporan y espacialmente de la fecundación.
Megaspora
Se encuentra al interior de cada micópilo, siendo una estructurua anclada, e inmóvil, pero con un desarrollo bastante complejo.
Gametofito masculino
Es el grano de polen ya germinado, en otras palabras, el tubo polinígfero que transporta no gametos, sino núcleos generatuivos gaméticos haploides, siendo estos dos por cada grano de polen. Un núcleo fertilizará el óvulo, y otro fertilizará una celula diploide que acompaña al gametofito produciebndo un endospermo triploide.
Gametofito femenino
El gametofito femenino nunca de ser una célula es una estructura compleja compuesta por ocho células únicamente, una de ellas es un óvulo, pero también se debe tener en cuenta a dos de las tres células polares, ya que son vitales en el desarrollo de la semilla no desnuda.
Fertilización
La fertilización es doble, siendo este un carácter apomórfico de las plantas con flores angiospermas. La primera fertilización estándar es la que produce in cigoto diploide a partir de los núcleos gaméticos hamploides como en cualquier otro ciclo de vida. La segunda fertilización se da entre el segundo núcleo generativo del grano de polen con dos núcleos polares, dando lugar a una célula triploide.
Ciclo de vida
El ciclo de vida de las angiospermas es similar al de las gimnospermas, el esporofito es la fase dominante, mientras que el gametofito es microscópico. Solo el gametofito masculino inmaduro es visible en forma de los granos de polen fuera de la planta, el gametofito femenino siempre se encuentra escondido al interior de la semilla y los carpelos de la flor.
R. Referencias bibliográficas
R. Referencias bibliográficas
Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molecular biology of the cell (5th ed.). Garland Science.
Ames, O. (1963). The significance of the angiosperm seed. Economic Botany, 17(1), 3–10.
Arana, M. D., Correa, A. L., & Oggero, A. J. (2014). EL REINO PLANTAE:¿ QUÉ ES UNA PLANTA Y CÓMO SE CLASIFICAN? Revista de Educación En Biología, 17(1), pp-9.
Balogh, P. (1982). Rhizanthella RS Rogers, a misunderstood genus (Orchidaceae). Selbyana, 7(1), 27–33.
Bateman, R. M., & DiMICHELE, W. A. (1994). Heterospory: the most iterative key innovation in the evolutionary history of the plant kingdom. Biological Reviews, 69(3), 345–417.
Belk, C., & Maier, V. B. (2013). Biology Science for Life with physiology. (Pearson, Ed.) (4th ed.).
Berta, A., Sumich, J. L., & Kovacs, K. M. (2015). Marine mammals: evolutionary biology. Academic Press.
Biswas, C., & Johri, B. M. (2013). The gymnosperms. Springer Science & Business Media.
Bonner, J. T. (2015). Life cycles: reflections of an evolutionary biologist. Princeton University Press.
Bowman, J. L., Sakakibara, K., Furumizu, C., & Dierschke, T. (2016). Evolution in the Cycles of Life. Annual Review of Genetics, 50, 133–154.
Brenner, E. D., Stevenson, D. W., & Twigg, R. W. (2003). Cycads: evolutionary innovations and the role of plant-derived neurotoxins. Trends in Plant Science, 8(9), 446–452.
Brock, K. (2014). Tracking the Evolutionary History of Development Genes: Implications for the Diversification of Fruits and Flowers in the Brassicaceae and Cleomaceae. University of Alberta.
Brusca, R., Brusca, G. J., & Haver, N. J. (2003). Invertebrates (2nd ed.). Sinauer Associates.
Buerki, S., Forest, F., & Alvarez, N. (2014). Proto‐South‐East Asia as a trigger of early angiosperm diversification. Botanical Journal of the Linnean Society, 174(3), 326–333.
Campbell, M. K., & Farrell, S. O. (2009). Biochemistry (6th ed.). USA: Thomsom Brooks/Cole.
Cascales-Miñana, B., Cleal, C. J., & Gerrienne, P. (2016). Is Darwin’s ‘Abominable Mystery’still a mystery today? Cretaceous Research, 61, 256–262.
Chaloner, W. G., Hill, A. J., & Lacey, W. S. (1977). First Devonian platyspermic seed and its implications in gymnosperm evolution. Nature, 265(5591), 233–235.
Chaloner, W. G., & Pettitt, J. M. (1987). The inevitable seed. Bulletin de La Société Botanique de France. Actualités Botaniques, 134(2), 39–49.
Chandrakanth, M. G., & Romm, J. (1991). Sacred forests, secular forest policies and people’s actions. Nat. Resources J., 31, 741.
Chang, T.-T. (1987). The impact of rice on human civilization and population expansion. Interdisciplinary Science Reviews, 12(1), 63–69.
CHAPMAN, V. J. (1961). Life histories in the algae. Annals of Botany, 25(4), 548–561.
Clayton, W. D., & Renvoize, S. A. (1986). Genera graminum. Grasses of the World. Genera Graminum. Grasses of the World., 13.
Cock, J. M., & Coelho, S. M. (2011). Algal models in plant biology. Journal of Experimental Botany, 62(8), 2425–2430.
Coelho, S. M., Peters, A. F., Charrier, B., Roze, D., Destombe, C., Valero, M., & Cock, J. M. (2007). Complex life cycles of multicellular eukaryotes: new approaches based on the use of model organisms. Gene, 406(1), 152–170.
Cousens, M. I. (1988). Reproductive strategies of pteridophytes. Plant Reproductive Ecology. Edited by J. Lovett-Doust and L. Lovett-Doust. Oxford University Press, New York, 307–328.
Cox, M. M., Doudna, J. A., & O’Donnell, M. (2012). Molecular biology (1st ed.). Freeman.
Crepet, W. L. (n.d.). Advances in Flowering Plant Evolution. ELS.
De Castro, R. D., & Marraccini, P. (2006). Cytology, biochemistry and molecular changes during coffee fruit development. Brazilian Journal of Plant Physiology, 18(1), 175–199.
De Clerck, O., Bogaert, K. A., & Leliaert, F. (2012). Diversity and evolution of algae: primary endosymbiosis. Adv Bot Res, 64, 55–86.
De Duve, C., & Pizano, M. (1995). Polvo vital: origen y evolución de la vida en la tierra. Norma, Bogotá.
Debate, A. (2012). Morphological Transformation, the Fossil Record, and the. Evolutionary Biology, 7, 295.
DeWet, J. M. J. (1981). Grasses and the culture history of man. Annals of the Missouri Botanical Garden, 87–104.
DiMichele, W. A., & Bateman, R. M. (1996). Plant paleoecology and evolutionary inference: two examples from the Paleozoic. Review of Palaeobotany and Palynology, 90(3–4), 223–247.
Dixon, K. (2003). Plate 468. Rhizanthella Gardneri. Curtis’s Botanical Magazine, 20(2), 94–100.
Essig, F. B. (2015). Plant life: a brief history. Oxford University Press.
Gandawijaja, D., Idris, S., Nasution, R., Nyman, L. P., & Arditti, J. (1983). Amorphophallus titanum Becc.: a historical review and some recent observations. Annals of Botany, 51(3), 269–278.
Gololobova, M. A. (2015). The position of “lower plants” on the tree of life. Biology Bulletin, 42(6), 500–507.
Goodenough, J., & McGuire, B. (2012). Biology of Humans, Concepts, Applications and Issues (4th ed.). San Francisco: Pearson, Benjamin Cummings.
Graham, L. E. (1985). The Origin of the Life Cycle of Land Plants: A simple modification in the life cycle of an extinct green alga is the likely origin of the first land plants. American Scientist, 73(2), 178–186.
Graham, L. K. E., & Wilcox, L. W. (2000). The origin of alternation of generations in land plants: a focus on matrotrophy and hexose transport. Philosophical Transactions of the Royal Society B: Biological Sciences, 355(1398), 757–767.
Hackett, J., Su Yoon, H., Butterfield, N. J., Sanderson, M., & Bhattacharya, D. (2007). Evolution of primary producers in the sea. In Elsevier Inc.
Haig, D. (2016). Living together and living apart: the sexual lives of bryophytes. Phil. Trans. R. Soc. B, 371(1706), 20150535.
Harholt, J., Moestrup, Ø., & Ulvskov, P. (2016). Why plants were terrestrial from the beginning. Trends in Plant Science, 21(2), 96–101.
Harshberger, J. W. (1896). The purposes of ethno-botany. Botanical Gazette, 21(3), 146–154.
Henry, R. J. (2005). Plant diversity and evolution: genotypic and phenotypic variation in higher plants. Cabi Publishing.
Herron, M. D. (2016). Fitness and individuality in complex life cycles. Philosophy of Science, 83(5), 828–834.
Hoefnagels, M. (2015). Biology: concepts and investigations (3rd ed.). McGraw-Hill New York.
Hughes, J. S., & Otto, S. P. (1999). Ecology and the evolution of biphasic life cycles. The American Naturalist, 154(3), 306–320.
Joel, D. M., Gressel, J., & Musselman, L. J. (2015). Parasitic orobanchaceae. Springer.
Kardong, K. V. (2011). Vertebrates, comparative anatomy, function, evolution (6th ed.). McGraw-Hill New York.
Kardong, K. V. (2014). Vertebrates: Comparative Anatomy, Function, Evolution (7th ed.). McGraw-Hill Education.
Karling, J. S., & Beuschlein, M. (1956). Plants and man. The American Biology Teacher, 18(1), 9–13.
Karp, G. C. (2013). Cell and Molecular Biology, Concepts and Experiments (7th ed.). USA: Wiley Online Library.
Kaur, A., Rana, A. C., Tiwari, V., Sharma, R., & Kumar, S. (2011). Review on Ethanomedicinal and Pharmacological Properties of Ficus religiosa.
KENRICK, B. (1994). Alternation of generations in land plants: new phylogenetic and palaeobotanical evidence. Biological Reviews, 69(3), 293–330.
Khush, G. S. (2016). Biodiversity Use for Food Security. Indian Journal of Plant Genetic Resources, 29(3), 251–252.
Kim, K. M., Park, J.-H., Bhattacharya, D., & Yoon, H. S. (2014). Applications of next-generation sequencing to unravelling the evolutionary history of algae. International Journal of Systematic and Evolutionary Microbiology, 64(2), 333–345.
Klees, G., & Piepenbring, M. (2017). Animated life cycles of fungi and plants with spores for teaching. Journal of Biological Education, 1–13.
Kofuji, R., & Hasebe, M. (2014). Eight types of stem cells in the life cycle of the moss Physcomitrella patens. Current Opinion in Plant Biology, 17, 13–21.
Lapierre, H., & Signoret, P.-A. (2004). Viruses and virus diseases of Poaceae (Gramineae). Editions Quae.
Lee, E. K., Cibrian-Jaramillo, A., Kolokotronis, S.-O., Katari, M. S., Stamatakis, A., Ott, M., … McCombie, W. R. (2011). A functional phylogenomic view of the seed plants. PLoS Genet, 7(12), e1002411.
Leslie, A. B., Beaulieu, J. M., & Mathews, S. (2017). Variation in seed size is structured by dispersal syndrome and cone morphology in conifers and other nonflowering seed plants. New Phytologist.
Ligrone, R., Duckett, J. G., & Renzaglia, K. S. (2012). Major transitions in the evolution of early land plants: a bryological perspective. Annals of Botany, mcs017.
Linkies, A., Graeber, K., Knight, C., & Leubner‐Metzger, G. (2010). The evolution of seeds. New Phytologist, 186(4), 817–831.
Lovette, I. J., & Fitzpatrick, J. W. (2016). Handbook of bird biology. John Wiley & Sons.
LU, B., & Ellstrand, N. (2014). World food security and the tribe Triticeae (Poaceae): Genetic resources of cultivated, wild, and weedy taxa for crop improvement. Journal of Systematics and Evolution, 52(6), 661–666.
Mable, B. K., & Otto, S. P. (1998). The evolution of life cycles with haploid and diploid phases. BioEssays, 20(6), 453–462.
Mackean, D. G., & Hayward, D. (2014). Biology (3rd ed.). IGCSE Cambridge.
Mackiewicz, P., & Gagat, P. (2014). Monophyly of Archaeplastida supergroup and relationships among its lineages in the light of phylogenetic and phylogenomic studies. Are we close to a consensus? Acta Societatis Botanicorum Poloniae, 83(4).
Mader, S. S. (2010). Biology (10th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2015). Biology (12th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2018). Essentials of biology (5th ed.). McGraw-Hill Education.
Magadum, S., Banerjee, U., Murugan, P., Gangapur, D., & Ravikesavan, R. (2013). Gene duplication as a major force in evolution. Journal of Genetics, 92(1), 155–161.
Marieb, E. N., Wilhelm, P. B., & Mallatt, J. (2017). Human Anatomy (8th ed.). Pearson.
Mason, K. A., Losos, J. B., Singer, S. R., & Raven, P. H. (2014). Biology (7th ed.). McGraw-Hill New York.
Mattu, N. (2017). Impact of Global Climate Change on Pollinator Diversity and Conservation: Current Issues and Future Challenges. Journal of Research: THE BEDE ATHENAEUM, 8(1), 76–88.
Mehltreter, K., Walker, L. R., & Sharpe, J. M. (2010). Fern ecology. Cambridge University Press.
Moghe, G. D., & Shiu, S. (2014). The causes and molecular consequences of polyploidy in flowering plants. Annals of the New York Academy of Sciences, 1320(1), 16–34.
Murray, R. K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V., & Weil, A. (2012). Harpers Illustrated Biochemistry (29th ed.). McGraw-Hill Medical.
Mursidawati, S. (2004). Mycorrhizal association, propagation and conservation of the myco-heterotrophic orchid Rhizanthella gardneri. University of Western Australia.
Musselman, L. J., Press, M. C., Press, M. C., & Graves, J. D. (1995). Parasitic plants. Parasitic Plants.
Niklas, K. J., & Cobb, E. D. (2016). The evolutionary ecology (evo-eco) of plant asexual reproduction. Evolutionary Ecology, 1–16.
Niklas, K. J., & Kutschera, U. (2010). The evolution of the land plant life cycle. New Phytologist, 185(1), 27–41.
Nogales, M., González‐Castro, A., Rumeu, B., Traveset, A., Vargas, P., Jaramillo, P., … Heleno, R. (2017). Contribution by vertebrates to seed dispersal effectiveness in the Galápagos Islands: A community‐wide approach. Ecology.
Oliver, K. R., McComb, J. A., & Greene, W. K. (2013). Transposable elements: powerful contributors to angiosperm evolution and diversity. Genome Biology and Evolution, 5(10), 1886–1901.
Otani, T. (2004). Effects of macaque ingestion on seed destruction and germination of a fleshy‐fruited tree, Eurya emarginata. Ecological Research, 19(5), 495–501.
Petersen, K. B., & Burd, M. (2016). Why did heterospory evolve? Biological Reviews.
Pollard, T. D., Earnshaw, W. C., Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell Biology (3rd ed.). Elsevier.
Qiu, Y., Taylor, A. B., & McMANUS, H. A. (2012). Evolution of the life cycle in land plants. Journal of Systematics and Evolution, 50(3), 171–194.
Ragan, M. A., & Gutell, R. R. (1995). Are red algae plants? Botanical Journal of the Linnean Society, 118(2), 81–105.
Reece, J. B., Urry, L. A., Wasserman, S. A., Cain, M. L., Minorsky, P. V, & Jackson, R. B. (2014). Campbell Biology (10th ed.). Pearson Higher Ed.
Ritchison, G. (2015). Bird biogeography. RITCHISON, G. Avian Biology: Everything You Wanted to Know about Birds… and More. Eastern Kentucky University,[Sd]. Disponível Em:< Http://People. Eku. Edu/Ritchisong/Birdbiogeography1. Htm>. Acesso Em, 9.
Roulin, A., Auer, P. L., Libault, M., Schlueter, J., Farmer, A., May, G., … Jackson, S. A. (2013). The fate of duplicated genes in a polyploid plant genome. The Plant Journal, 73(1), 143–153.
Rueda, M., Godoy, O., & Hawkins, B. A. (2017). Spatial and evolutionary parallelism between shade and drought tolerance explains the distributions of conifers in the conterminous United States. Global Ecology and Biogeography, 26(1), 31–42.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of Biology (10th ed.). Sinauer & MacMillian.
Sadler, T. W. (2012). Medical Embryology. (C. Taylor, Ed.) (12th ed.). Baltimore: Lippincott Williams & Wilkins.
Sanders, W. B. (2014). Complete life cycle of the lichen fungus Calopadia puiggarii (Pilocarpaceae, Ascomycetes) documented in situ: Propagule dispersal, establishment of symbiosis, thallus development, and formation of sexual and asexual reproductive structures. American Journal of Botany, 101(11), 1836–1848.
Schneider, H., Pryer, K. M., Cranfill, R., Smith, A. R., & Wolf, P. G. (2002). Evolution of vascular plant body plans: a phylogenetic perspective. Developmental Genetics and Plant Evolution, 330–364.
Searles, R. B. (1980). The strategy of the red algal life history. The American Naturalist, 115(1), 113–120.
Sessa, E. B., & Der, J. P. (2016). Chapter Seven-Evolutionary Genomics of Ferns and Lycophytes. Advances in Botanical Research, 78, 215–254.
Sharma, S., Rikhari, H. C., & Palni, L. M. S. (1999). Conservation of natural resources through religion: a case study from Central Himalaya. Society & Natural Resources, 12(6), 599–612.
Shier, D., Butler, J., & Lewis, R. (2015). Hole’s Human Anatomy & Physiology (14th ed.). McGraw-Hill Education.
Simon, E. J., Dickey, J. L., & Reece, J. B. (2013). Essential biology (Pearson).
Solomon, E., Martin, C., Martin, D. W., & Berg, L. R. (2014). Biology (10th ed.). Cengage Learning.
Soltis, P. S., & Soltis, D. E. (2014). Flower diversity and angiosperm diversification. Flower Development: Methods and Protocols, 85–102.
Stallknecht, G. F., Gilbertson, K. M., & Eckhoff, J. L. (1993). Teff: Food crop for humans and animals. New Crops. Wiley, New York, 5, 231–234.
Starr, C., Evers, C., & Starr, L. (2013). Biology: Today and Tomorrow With Physiology (4th ed.). Brooks/Cole.
Taylor, E. L., Taylor, T. N., & Krings, M. (2009). Paleobotany: the biology and evolution of fossil plants. Academic Press.
Taylor, M. R., Simon, E. J., Dickey, J. L., Hogan, K., & Reece, J. B. (2018). Biology, Concepts and conections (9th ed.). Boston: Pearson education.
Traveset, A., & Verdú, M. (2002). 22 A Meta-analysis of the Effect of Gut Treatment on Seed Germination. Seed Dispersal and Frugivory: Ecology, Evolution, and Conservation, 339.
Umen, J. G. (2014). Green algae and the origins of multicellularity in the plant kingdom. Cold Spring Harbor Perspectives in Biology, 6(11), a016170.
Vitt, L. J., & Caldwell, J. P. (2014). Herpetology An Introductory Biology of Amphibians and Reptiles (4th ed.). Oklahoma: Elsevier Inc.
Wang, X.-Q., & Ran, J.-H. (2014). Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution, 75, 24–40.
Watson, L. (1990). The grass family, Poaceae. Reproductive Versatility in the Grasses, 1–31.
Weaver, R. F. (2012). Molecular biology (5th ed.). McGraw-Hill Education.
Willis, K., & McElwain, J. (2014). The evolution of plants. Oxford University Press.
Zungu, M. M. (2014). Aspects of fruit digestion and selection in selected southern African avian frugivores. Citeseer.
No hay comentarios:
Publicar un comentario