
Indice
|
||1|| Introducción ||2|| Generalidades ||3|| El
citosol ||4|| Organelos
procariotas, el flagelo ||5|| La
membrana celular procariota ||6|| Pared
celular procariota ||7|| El
citoplasma ||8|| El
ribosoma procariota |
||9|| Los
retículos endoplasmáticos eucariotas ||10|| El
aparato de Golgi eucariota ||11|| El
núcleo celular eucariota ||12|| Vesículas
celulares eucariotas ||13|| Organelos
energéticos y endosimbióticos ||14|| Otros
organelos de las células eucariotas ||R|| Referencias |
Portada

1. Introducción
|| Regresar al índice ||
El sistema de endomembranas constituye el
principal sistema de organelos celulares, partes especializadas en funciones
concretas y está compuesto por las diferentes membranas que están suspendidas
en el citoplasma dentro de una célula eucariota. Estas membranas dividen la
célula en compartimentos funcionales y estructurales, u orgánulos.
1.1 Christian René Marie Joseph, Vizconde de Duve (2 October 1917 – 4 May
2013)
Fue un citólogo y
bioquímico belga ganador del Premio Nobel. Hizo los descubrimientos de dos
orgánulos celulares, peroxisomas y lisosomas, por los que compartió el Premio
Nobel de Fisiología o Medicina en 1974 con Albert Claude y George E. Palade
("por sus descubrimientos sobre la organización estructural y funcional de
la célula"). Además de peroxisoma y lisosoma, inventó nombres científicos
como autofagia, endocitosis y exocitosis en una sola ocasión.
Hijo de refugiados
belgas durante la Primera Guerra Mundial, de Duve nació en Thames Ditton,
Surrey, Inglaterra. Su familia regresó a Bélgica en 1920. Fue educado por los
jesuitas en Our Lady College, Amberes, y estudió medicina en la Universidad
Católica de Lovaina. Al obtener su título de médico en 1941, se unió a la
investigación en química, trabajando en la insulina y su papel en la diabetes mellitus.
Su tesis le valió el título universitario más alto agrégation de
l'enseignement supérieur (equivalente a PhD) en 1945.
Con su trabajo sobre
la purificación de la penicilina, obtuvo una maestría en ciencias en 1946. Se
formó con (posteriormente ganadores del Premio Nobel) Hugo Theorell en el
Karolinska Institutet en Estocolmo, y Carl y Gerti Cori en la
Universidad de Washington en St. Luis. Se unió a la facultad de medicina de
Lovaina en 1947. En 1960 fue invitado al Instituto Rockfeller (ahora
Universidad Rockefeller). Por mutuo acuerdo con Lovaina, se convirtió en
profesor en ambas universidades a partir de 1962, dividiendo su tiempo entre
Lovaina y Nueva York. En 1974, el mismo año en que recibió su Premio Nobel,
fundó el ICP, que más tarde pasaría a llamarse Instituto de Duve. Se convirtió
en profesor emérito de la Universidad de Lovaina en 1985 y de Rockefeller en 1988.
De Duve recibió el
rango de Vizconde en 1989 por el rey Balduino de Bélgica. También
recibió el Premio Francqui, el Premio Internacional de la Fundación
Gairdner, el Premio Heineken y el Premio E.B. Medalla Wilson. En 1974
fundó el Instituto Internacional de Patología Celular y Molecular en Bruselas,
eventualmente rebautizado como Instituto de Duve en 2005. Fue el presidente
fundador de los Premios L'Oréal-UNESCO para Mujeres en la Ciencia. Murió por
eutanasia legal después de sufrir durante mucho tiempo cáncer y fibrilación
auricular.
2. Generalidades
|| Regresar al índice ||
En los eucariotas, los orgánulos del
sistema endomembrana incluyen: la membrana nuclear, el retículo endoplásmico,
el aparato de Golgi, los lisosomas, las vesículas, los endosomas y la membrana
plasmática (celular), entre otros. El sistema se define con mayor precisión
como el conjunto de membranas que forman una sola unidad funcional y de
desarrollo, ya sea que se conectan directamente o intercambian material a
través del transporte de vesículas.
👉 Es importante destacar que el sistema de
endomembrana no incluye las membranas de los cloroplastos o las mitocondrias,
pero podría haber evolucionado a partir de este último.
👉 La membrana nuclear contiene una bicapa
lipídica que abarca el contenido del núcleo.
👉 El retículo endoplásmico (ER) es un orgánulo
de síntesis y transporte que se ramifica hacia el citoplasma en las células
vegetales y animales.
👉 El aparato de Golgi es una serie de
compartimentos múltiples donde las moléculas se empaquetan para su entrega a
otros componentes celulares o para la secreción de la célula.
👉 Las vacuolas, que se encuentran en las células
vegetales y animales (aunque mucho más grandes en las células vegetales), son
responsables de mantener la forma y estructura de la célula, así como de
almacenar los productos de desecho.
👉 Una vesícula es un saco relativamente pequeño,
encerrado en una membrana, que almacena o transporta sustancias.
👉 La membrana celular es una barrera protectora
que regula lo que entra y sale de la célula.
👉 También hay un organelo conocido como
Spitzenkörper que solo se encuentra en hongos y está relacionado con el crecimiento
de la punta de la hifa.
En las procariotas, las endomembranas son
raras, aunque en muchas bacterias fotosintéticas la membrana plasmática está
muy plegada y la mayor parte del citoplasma celular está lleno de capas de membrana
que recoge la luz. Estas membranas de recolección de luz pueden incluso formar
estructuras cerradas llamadas clorosomas o tilacoides.
Los orgánulos del sistema de endomembranas
están relacionados por contacto directo o por transferencia de segmentos de
membrana como vesículas. A pesar de estas relaciones, las diversas membranas no
son idénticas en estructura y función. El grosor, la composición molecular y el
comportamiento metabólico de una membrana no son fijos, pueden modificarse
varias veces durante la vida útil de la membrana. Una característica
unificadora que comparten las membranas es una bicapa lipídica, con proteínas
unidas a ambos lados o atravesando ellas.
El sistema de endomembranas no es el único
sistema de organelos, pues podemos tener organelos dependientes del citoesqueleto,
así como el propio citoesqueleto, así como organelos derivados de la
endosimbiosis con bacterias.
3. El citosol
|| Regresar al índice ||
Hasta ahora, nos hemos centrado en las características generales de las células procariotas y eucariotas. En el resto de este capítulo, examinaremos los diversos compartimentos de las células eucariotas con un mayor énfasis en la estructura y la función. La Figura 3.1 destaca una célula animal y vegetal de acuerdo con cuatro regiones diferentes.

Figura 3.1. Compartimentos dentro de (a) células
animales y (b) vegetales. El citosol, que está fuera de los orgánulos, pero
dentro de la membrana plasmática, se muestra en amarillo. Las membranas del
sistema de endomembranas se muestran en púrpura y los interiores llenos de
líquido son de color rosa. El peroxisoma es de color púrpura oscuro. El
interior del núcleo es azul. Los organelos semiautónomos se muestran en naranja
(mitocondrias) y verde (cloroplastos).
Comenzaremos con el citosol (que se
muestra en amarillo), la región de una célula eucariota que está fuera de los
orgánulos unidos a la membrana, pero dentro de la membrana plasmática. Las
otras regiones de la célula, que examinaremos más adelante en este capítulo,
incluyen el interior del núcleo (azul), el sistema de endomembranas (púrpura y
rosa) y los orgánulos semiautónomos (naranja y verde). Como en las células
procariotas, el término citoplasma se refiere a la región encerrada por la
membrana plasmática. Esto incluye el citosol y los orgánulos.
3.1 Un espacio para la química
El metabolismo se define como la suma de
las reacciones químicas por las cuales las células producen los materiales y
utilizan la energía necesaria para mantener la vida. Aunque también se producen
muchos pasos del metabolismo en los orgánulos celulares, el citosol es una
región central de coordinación para muchas actividades metabólicas de las
células eucariotas. El metabolismo a menudo implica una serie de pasos llamados
vías metabólicas. Cada paso en una ruta metabólica es catalizado por una enzima
específica, una proteína que acelera la velocidad de una reacción química.
Algunas vías implican la descomposición de
una molécula en componentes más pequeños, un proceso denominado catabolismo. Dichas vías son necesarias
para que la célula utilice energía y también para generar moléculas que
proporcionen los bloques de construcción para construir macromoléculas. Por el
contrario, otras vías están involucradas en el anabolismo, la síntesis de moléculas y macromoléculas. Por ejemplo,
los polisacáridos se hacen uniendo moléculas de azúcar. Para producir
proteínas, los aminoácidos se conectan covalentemente para formar un
polipéptido, utilizando la información dentro de un ARNm. La traducción ocurre
en los ribosomas, que se encuentran en varios lugares de la célula. Algunos ribosomas
pueden flotar libremente en el citosol, otros están unidos a la membrana
externa de la envoltura nuclear y la membrana del retículo endoplásmico, y
otros se encuentran dentro de las mitocondrias o cloroplastos.
3.2 El citoesqueleto
El citoesqueleto es una red de tres tipos
diferentes de filamentos de proteínas: microtúbulos, filamentos intermedios y
filamentos de actina. Cada tipo está construido a partir de muchos monómeros de
proteínas. El citoesqueleto es un ejemplo sorprendente de interacciones proteína-proteína.
El citoesqueleto se encuentra principalmente en el citosol y también en el
núcleo a lo largo de la membrana nuclear interna. Consideremos primero la
estructura de los filamentos del citoesqueleto y sus funciones en la
construcción y organización de las células. Más adelante, examinaremos cómo
están involucrados en el movimiento celular.
Los microtúbulos: Los microtúbulos son estructuras cilíndricas largas, huecas, de aproximadamente 25 nm de diámetro, compuestas de subunidades proteicas llamadas α- y β-tubulina.

Figura 3.2. Modelo
simplificado de un microtúbulo y las proteínas que lo conforman modeladas como
esferas.
El ensamblaje de tubulina para formar un
microtúbulo da como resultado una estructura con un extremo positivo y un
extremo negativo. Los microtúbulos crecen solo en el extremo positivo, pero
pueden acortarse en el extremo positivo o negativo. Un solo microtúbulo puede
oscilar entre las fases de crecimiento y acortamiento, un fenómeno denominado
inestabilidad dinámica. Este fenómeno es importante en muchas actividades
celulares, incluida la generación de cromosomas durante la división celular.
Los sitios donde se forman los
microtúbulos dentro de una célula varían entre los diferentes tipos de
organismos. Las células animales que no se dividen contienen una estructura
única cerca de su núcleo llamada centrosoma, también llamado centro organizador
de microtúbulos. Dentro del centrosoma se encuentran los centriolos, un par de
estructuras conspicuas dispuestas perpendiculares entre sí. En las células
animales, el crecimiento de microtúbulos generalmente comienza en el centrosoma
de tal manera que el extremo negativo está anclado allí. En contraste, la
mayoría de las células vegetales y muchos protistas carecen de centrosomas y
centriolos. Los microtúbulos se crean en muchos sitios que están diseminados
por una célula vegetal. En las plantas, la membrana nuclear parece funcionar
como un centro organizador de microtúbulos.
Los microtúbulos son importantes para la
forma y organización de las células. Los orgánulos como el aparato de Golgi
están unidos a los microtúbulos. Además, los microtúbulos están involucrados en
la organización y el movimiento de los cromosomas durante la mitosis y en la
orientación de las células durante la división celular.
Filamentos intermedios: Los filamentos intermedios son otra clase de filamento citoesquelético que se encuentra en las células de muchas especies animales, pero no todas.

Figura 3.3. Comparación en
las estructuras de varios componentes del citoesqueleto.
Su nombre se deriva de la observación de
que tienen un diámetro intermedio entre los filamentos de actina y los
microtúbulos. Las proteínas de filamentos intermedios se unen entre sí en una
matriz escalonada para formar una estructura retorcida en forma de cuerda con
un diámetro de aproximadamente 10 nm. Funcionan como fibras que soportan
tensión que ayudan a mantener la forma y rigidez de la célula. Los filamentos
intermedios tienden a ser relativamente permanentes. En comparación, los
microtúbulos y los filamentos de actina se alargan y acortan fácilmente en las
células.
Varios tipos de proteínas se ensamblan en
filamentos intermedios. Las queratinas forman filamentos intermedios en las
células de la piel, intestinales y renales, donde son importantes para la forma
celular y la resistencia mecánica. También son un componente importante del
cabello y las uñas. Además, los filamentos intermedios se encuentran dentro del
núcleo celular. Como se discutirá más adelante en este capítulo, las láminas
nucleares forman una red de filamentos intermedios que recubren la membrana
nuclear interna y proporcionan puntos de anclaje para los poros nucleares.
Filamentos de actina: Los filamentos de actina también se conocen como microfilamentos,
porque son los filamentos del citoesqueleto más delgados. Son fibras largas y
delgadas de aproximadamente 7 nm de diámetro. Al igual que los microtúbulos,
los filamentos de actina tienen extremos positivo y negativo, y son estructuras
muy dinámicas en las que cada hebra crece en el extremo positivo mediante la
adición de monómeros de actina. Este proceso de ensamblaje produce una fibra
compuesta por dos hebras de monómeros de actina que giran en espiral una
alrededor de la otra.
A pesar de su delgadez, los filamentos de
actina juegan un papel clave en la forma y resistencia celular. Aunque los
filamentos de actina se dispersan por todo el citosol, tienden a estar
altamente concentrados cerca de la membrana plasmática. En muchos tipos de
células, los filamentos de actina apoyan la membrana plasmática y proporcionan
forma y resistencia a la célula. Los lados de los filamentos de actina a menudo
están anclados a otras proteínas cerca de la membrana plasmática, lo que
explica por qué los filamentos de actina generalmente se encuentran allí. Los
extremos positivos crecen hacia la membrana plasmática y juegan un papel clave
en la forma y el movimiento de las células.
3.3 Proteínas motoras
Las proteínas motoras son una categoría de
proteínas que usan ATP como fuente de energía para promover varios tipos de
movimientos. Una proteína motora consta de tres dominios: la cabeza, la bisagra
y la cola. La cabeza es el sitio donde el ATP se une y se hidroliza a difosfato
de adenosina (ADP) y fosfato inorgánico (Pi). La unión al ATP y la hidrólisis
causan una curva en la bisagra, lo que resulta en movimiento. La región de la
cola está unida a otras proteínas o a otros tipos de moléculas celulares.
Para promover el movimiento, la región de la cabeza de una proteína motora interactúa con un filamento citoesquelético, como un filamento de actina (. Cuando el ATP se une y se hidroliza, la proteína motora intenta "caminar" a lo largo del filamento. La cabeza de la proteína motora se une inicialmente a un filamento. Para avanzar, la cabeza se separa del filamento, se inclina hacia adelante, se une al filamento y se inclina hacia atrás. Para imaginar cómo funciona esto, considere el acto de caminar e imagine que el suelo es un filamento citoesquelético, su pierna es la cabeza de la proteína motora y su cadera es la bisagra.

Figura 3.4. La quinesina es un ejemplo de proteína motora (YouTube)
Para caminar, levantas la pierna, la
mueves hacia adelante, la colocas en el suelo y luego la empujas hacia atrás
(lo que te impulsa hacia adelante). Esta serie de eventos es análoga a cómo se
mueve una proteína motora a lo largo de un filamento citoesquelético.
Las proteínas motoras pueden causar tres
tipos diferentes de movimientos: movimiento de carga a través de la proteína
motora, movimiento del filamento o flexión del filamento. La región de la cola
de una proteína motora llamada quinesina está unida a una carga, por lo que la
proteína motora mueve la carga de un lugar a otro. Alternativamente, una
proteína motora llamada miosina puede permanecer en su lugar y hacer que el
filamento se mueva. Esto ocurre durante la contracción muscular. Una tercera
posibilidad es que tanto la proteína motora como el filamento están
restringidos en su movimiento debido a la presencia de proteínas de enlace. En
este caso, cuando las proteínas motoras llamadas dineína intentan caminar hacia
el extremo negativo, ejercen una fuerza que hace que los microtúbulos se
doblen.
En ciertos tipos de células, los
microtúbulos y las proteínas motoras facilitan el movimiento de los apéndices
celulares llamados flagelos y cilios (singular, flagelo y cilio). La diferencia
entre los dos es que los flagelos generalmente son más largos que los cilios y
generalmente se encuentran solos o en pares.
Tanto los flagelos como los cilios causan
movimiento al generar curvas que se mueven a lo largo y empujan hacia atrás
contra el fluido circundante. Una célula de esperma genera curvas
alternativamente en cada dirección, que comienzan en la cabeza y se mueven (se
propagan) hacia la punta del flagelo. Alternativamente, un par de flagelos
pueden moverse de manera sincronizada para tirar de un microorganismo a través
del agua (piense en un nadador humano haciendo la brazadas). Ciertas algas unicelulares
nadan de esta manera. En comparación, los cilios son a menudo más cortos que
los flagelos y tienden a cubrir toda o parte de la superficie de una célula.
Protistas como los paramecios pueden tener cientos de cilios adyacentes que
laten de manera coordinada para impulsar el organismo a través del agua.
4. Organelos procariotas el flagelo
|| Regresar al índice ||
Cuando estudié mis cursos básicos de
biología en el colegio e incluso en la universidad, la visión de la célula
procariota que obtuve fue la de un organismo simple sin organelos membranosos y
casi todo el espectáculo se lo llevaba la célula eucariótica. Sin embargo, en
los últimos 10 años ha empezado a emerger una visión renovada de la célula
procariota. Los procariotas poseen un citoesqueleto, aunque más simple que el
de los eucariotas, poseen proteínas homólogas a la actina y a la miosina
llamadas MreB y FtsZ, además de las proteínas filamentosas segregadas por los
flagelos en crecimiento como la flagelina. También se ha empezado a visualizar
la presencia en algunos linajes de un sistema de membranas internas para la
fotosíntesis o la quimiolitotrofía. Adicionalmente otras especies poseen
compartimentos aislados por proteína o polisacáricos. Estos organelos
procarióticos son bastante pequeños, después de toda una bacteria mide casi lo
que mide un organelo eucariótico.
La
mayoría de los procariotas miden entre 1-10 micrómetros, pero pueden variar
desde 0.2-750 micrómetros, siendo la especie más pequeña Mycoplasma genitalium y la más grande Thiomargarita namibiensis. A continuación, realizaremos un
breve estudio por los diferentes organelos procariotas
El flagelo es una estructura semejante a un látigo que emerge de las células procariota y eucariota. La principal función de los flagelos es la locomoción, pero también funciona como un organelo sensorial. Los flagelos se definen por su función más que por su estructura evolutiva, ya que estos pueden tener formas muy diversas. A pesar de lo anterior, los flagelos pueden dividirse de acuerdo a su linaje evolutivo en los flagelos eucariótico, bacteriano y arcaico. Las principales diferencias son las siguientes:

Figura 4.1. Flagelo de una bacteria Gram negativa.
4.1 Los
flagelos bacterianos
son filamentos helicales, cada uno segregado
por una base giratoria que evolucionó en base a otras proteínas giratorias como
las f1f0 ATP sintetasas-hidrolasas que se mueven a favor o en contra de las
manecillas del reloj. El flagelo bacteriano es bastante conocido ya que es el
blanco de los argumentos creacionistas sobre su diseño inteligente o con
propósito único para la motilidad, sin embargo, esto omite que varias partes
por separado pueden funcionar para otras cosas, como por ejemplo órgano
sensorial, jeringa molecular o despojado de casi todas las proteínas
accesorias, simplemente ser un poro secretorio o sintetizar energía (Kåhrström,
2013; Nunes-Alves, 2015).
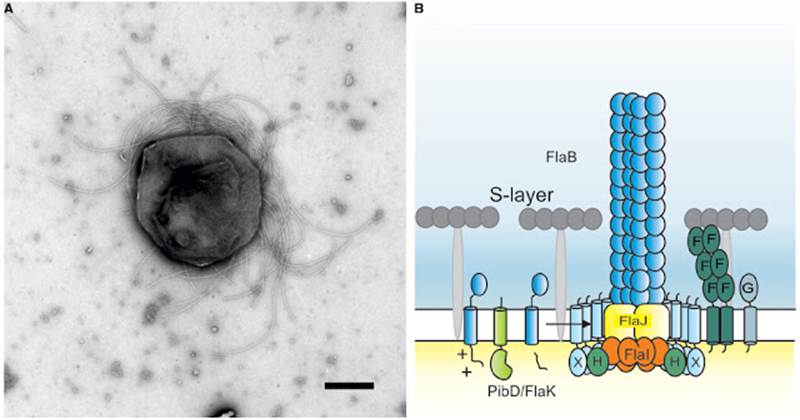
Figura 4.2. Flagelo de una arcaica.
4.2 Los
flagelos arcaicos
No
reciben su nombre porque sean los más viejos sino porque pertenecen al dominio
de las arcaicas, y aunque superficialmente es similar al flagelo de las
bacterias, poseen muchas diferencias ultraestructurales, por lo que en la
actualidad se considera que el flagelo de las arcaicas evolucionó
independientemente al de las bacterias siendo un ejemplo de evolución
convergente y de órganos análogos. Por ejemplo, estos flagelos son energizados
por bombas iónicas como protones o iones sodio(1+) y no directamente por ATP,
además que la proteína que genera el torque no es la F1F0 ATP
sintetaza-hidrolaza sino otra que aún no ha sido identificada. Adicionalmente
los flagelos parecen coordinados a diferencia de las bacterias cuyos flagelos
se mueven independientemente, la coordinación se origina del hecho de que todos
se mueven por la misma presión osmótica (Bardy,
Ng, & Jarrell, 2004; Thomas, Bardy, & Jarrell, 2001).
Otras
estructuras llamadas pili sirven para que una célula con plásmidos pueda
trasferir su plásmido a otra célula o para anclarse en una biopelícula. La
bacteria a la cual se le transfiere el plásmido se transforma generando sus
propios pili y adquiriendo propiedades especiales como la resistencia a los
antibióticos.
5. La membrana celular procariota
Los
muros de una casa, los muros de una caja fuerte o las láminas de un carro
separan al frágil interior de ambientes impredecibles y hostiles. Estas
barreras son duras y rígidas, para poder realizar esta protección. Podríamos
esperar que la barrera que separa lo vivo de lo no vivo fuera una estructura
igual de poderosa, dura y rígida; y aun así, la membrana celular es de hecho
una barrera delgada, frágil y dinámica. La membrana biológica es donde ocurre
la mayoría de reacciones químicas que mantienen a los seres vivos, sean
procariotas o ecuariotas. Por esto es una ventaja para los seres vivos tener
una mayor cantidad de superficie de membranas. Las funciones de las membranas
pueden resumirse como:
5.1 Funciones
de la membrana procariota:
a- Compartimentalizacion
celular: no muy
común en el mundo procariótico, pero si se han reportado especies que poseen
sistema de membranas internas como los tilacoides de las cianobacterias.
b- Andamiaje enzimático: debido a que la mayoría de los
procariotas no poseen membranas internas, las enzimas que necesitan estar
insertadas en membranas se encuentran casi exclusivamente insertadas en la
membrana celular, que no es del todo externa ya que por fuera existen otras
estructuras como membranas secundarias o paredes celulares.
c- Barrera semipermeable
selectiva: en
este caso no es tan importante como en los eucariotas, ya que la pared celular
funciona como un filtro selectivo previo antes de que las sustancias puedan
hacer contacto con la membrana celular.
d- Trasporte de solutos: todas las células deben tener
membranas capaces de intercambiar sustabncias entre el interior y el exterior
para mantenerse vivas. Lo cual incluye la posibilidad de emplear solutos para
la síntesis de energía.
e- Respuestas a señales
externas:
mediante receptores proteínicos insertados en la membrana, este es un aspecto
vital para el desarrollo de biopelículas por medio del sistema de sensibilidad
a la densidad poblacional, cuando el receptor detecta muchas toxinas, activa
una reacion en cascada que actuia sobre el genoma, activando genes que inducen
a la bacteria a cambiar un comportamiento individual a un comportamiento
colectivo.
5.2 Estructura
de la membrana celular:
La
membrana de los seres vivos está compuesta por sustancias químicas semejantes a
la grasa o los aceites llamadas fosfolípidos, pero a diferencia de ellos pueden
mezclarse con el agua. Los fosfolípidos tienen forma alargada, una cabeza que
puede tocar el agua y una cola que es repelida por el agua. Cuando muchos de
estos lípidos están juntos forman espontáneamente una capa doble.
Esta
capa doble se denomina bicapa lipídica. En la cual las cabezas se encuentran en
contacto con el agua externa e interna de la célula, mientras que las colas se
encuentran en medio. A parte de los fosfolípidos otras moléculas semejantes a
las grasas pueden estar presentes como el colesterol. El colesterol afecta la rigidez
de la membrana y por lo tanto su resistencia a los cambios de temperatura.
Entre más colesterol, la membrana es más rígida y soporta mayores temperaturas.
En medio de la bicapa lipídica se encuentran nadando una serie de
moléculas grandes con formas de nudos muy apretados llamadas proteínas de
membrana. Los fosfolipídos de los dos dominios procariotas poseen diferencias
fundamentales:
Fosfolípido de las bacterias y eucariotas: Debido a que las bacterias y las eucariotas son los seres vivos mejores conocidos, por lo general tendemos a explicar a la membrana celular en términos de sus fosfolípidos como si fueran los únicos posibles. Los fosfolipídos de estos dominios se conocen como estéricos debido forman un grupo ester dada la unión de un grupo carboxilo con un grupo hidroxilo de forma tal que el carbono estérico queda con tres enlaces a dos oxígenos, uno de los oxígenos con un doble enlace y otro oxigeno con un enlace simple que se conecta a otra cadena de carbonos (RCOOR).

Figura 5.1. Los
fosfolípidos son dos ácidos grasos unidos a un glicerol y este a un grupo
fosfato.
En la imagen anterior podemos ver el
modelo molecular de un fosfolípido, el cual está compuesto por dos ácidos
carboxílicos largos que se unen a un grupo glicerol por medio de sus grupos
carboxilo. El glicerol recibe a los grupos carboxilo por medio de tres oxígenos
que un grupo hidroxilo que ha perdido su hidrógeno creando un puente estérico.
El glicerol posee por lo tanto dos grupos estéricos para dos ácidos
carboxílicos y un último oxígeno para recibir un grupo fosfato que se orienta
de forma opuesta a las cadenas alifáticas de los grupos carboxilo. De esta
forma se proyecta linealmente una punta con muchos oxígenos que es par y dos
colas alifáticas muy apolares, a mayor longitud de las colas, la apolaridad del
fosfolípido aumenta.
Fosfolípidos de las arcaicas: Existen dos tipos de fosfolípidos en las arcaicas, los fosfolípidos del éter. Un fosfolípido de éter se
forma entre un grupo glicerol y alcoholes 1-terminales de forma tal que solo
hay un oxígeno que sirve de puente entre la cadena alifática larga de la cola y
el glicerol, todo lo demás es igual. Los fosfolípidos de éter se comportan
semejante a los fosfolípidos de éter, sin embargo, las arcaicas poseen otro
tipo de fosfolípido. En algunas arcaicas las dos colas alifáticas es mucho más
extensa y termina en la unión con un azúcar en anillo.

Figura 5.2. Los
fosfolípidos de las arcaicas poseen ramificaciones de grupo metilo.
5.3 Fisiología
de la membrana celular
Debido a
la extensión del presente artículo no ahondaremos en las funciones de la
membrana, tópico que puede ahondarse posteriormente. La membrana celular sirve
como barrera homeostática, definiendo el adentre y el afuera de la célula, y por
lo tanto determinando lo que es nutrición “ingreso de moléculas útiles” de la
excreción “salida de sustancias venenosas”.
El paso
de las sustancias a través de membrana limita la forma de las células
procariotas, así como su tamaño; en los eucariotas condiciona no solo los
rasgos anteriores, también condiciona la forma de tejidos, órganos e inclusive
de todo un ser vivo multicelular. La membrana permite el paso libre de pocas
sustancias, la mayoría debe pasar a través de canales hechos de proteína. Del paso
de sustancias a través de membrana depende que la célula pueda almacenar
nutrientes, transformarlos de forma útil, responder a las condiciones del
ambiente y obtener energía.
6. La pared celular procariota
|| Regresar al índice ||
La
mayoría de las bacterias están rodeadas por una pared hecha de peptidoglicano,
aunque otras células pueden hacer paredes con otros tipos de polisacáridos
complejos. Las membranas celulares de las bacterias les dan forma, pueden ser
duras, flexibles o en ocasiones rígidas. En ausencia de un citoesqueleto más desarrollado,
es la pared celular quien le otorga a la célula procariota la base para tomar
su forma, además que la protege de los cambios osmóticos dentro de un rango de
tolerancia.
La pared
también actúa como un mecanismo de filtrado de materiales antes de llegar a la
membrana. Las paredes celulares también están presentes en las células
eucariotas donde realizan funciones semejantes, pero están hechas con
materiales diferentes. Algunas bacterias como los micoplasmas carecen
totalmente de paredes celulares. Las arcaicas también poseen paredes celulares,
sin embargo, en este caso están hechas de capas de glicoproteínas S,
pseudopeptidoglicano u otros polisacáridos complejos.
6.1 Las
paredes celulares de las bacterias
La pared
celular de los procariotes del tipo eubacteria se divide en dos tipos. Algunas
poseen una pared gruesa y se llaman Gram positivas y otra una pared delgada y
se llaman Gram negativas.

Figura 6.1. La pared celular se muestra en verde y morado. A la izquierda la pared celular de una bacteria grampositiva y a la derecha la pared de una bacteria gramnegativa. La pared está compuesta por una molécula de tipo polipeptido llamada peptidoglicano exclusiva de las bacterias. A las bacterias con paredes gruesas las llamamos Gram positivas y a las de paredes delgadas Gram negativas, debido a que es la pared la que retiene el colorante de Gram (color morado).

Figura 6.2. En este segundo
modelo también podemos apreciar a la izquierda la pared de la célula G+ y a la
derecha la G-.
Las Gram
positivas se ven purpuras y las Gram negativas rosadas al microscopio óptico.
La función de la pared es proteger a la célula y darle una forma definida. El
peptidoglicano de las bacterias es específico de ellas, de hecho, los primeros
antibióticos fueron desarrollados para atacar la formación de la pared
bacteriana, lo cual hace que sus células exploten como globos de fiesta. El
peptidoglicano se encuentra hecho con polisacáridos complejos unidos de forma
cruzada por péptidos inusuales que contienen aminoácidos de giro derecho, lo
cual es raro ya que las proteínas de virtualmente todos los seres vivos se
encuentran hechas con aminoácidos de giro izquierdo
6.2 Pared
celular de las arcaicas
Aunque
no es realmente único, la pared celular de las arcaicas es inusual. Mientras
que el componente principal de la pared de las bacterias es el peptidoglicano,
todas las arcaicas carecen de el, con la sola excepción de los metanógenos. En
ese grupo el peptidoglicano se encuentra modificado de una forma bastante
marcada en comparación con el de las bacterias. En la actualidad se conocen
cuatro tipos de paredes celulares en las arcaicas (König,
1988).
Pseudopeptidoglicano: De forma semejante al
peptidoglicano, se trata de un polímero de cadenas de glicano unidas de forma
cruzada por conecciones cortas de péptidos. Pero a diferencia del
peptidoglicano bacteriano, el azúcar acido N-acetilmuramico es reemplazado por
el ácido N-acetiltalosaminuronico. Adicionalmente los péptidos empleados para
las soldaduras cruzadas emplean péptidos de giro izquierdo (Levógiros) en lugar
de los péptidos dextrógiros (giro derecha) encontrados en las bacterias.
Paredes rígidas: Encontrado en los
clados Methanosarcina y Halococcus, se trata de
una pared dura compuesta por polisacáridos densamente agrupados que
pueden estar sulfatados en el caso de Halococcus. La estructura en este
tipo de pared es compleja y no ha sido investigada totalmente.
Glicoproteinas: Se encuentra en los hipertermófilos, Halobacterium, y algunos metabnógenos. En Halobacterium la pared es una
estructura de proteínas con una gran cantidad se aminoácidos acídicos, por lo
que la estructura en general es fuertemente negativa, evidentemente es una
estructura inestable por sí misma y requiere enormes cantidades de ion sodio
para estabilizarse. Resulta evidente que estos iones son atraídos por la propia
pared y provienen del medio altamente salino en el que estas bacterias
conviven. En consecuencia, si la bacteria es puesta en un ambiente de menor
salinidad su pared celular colapsa por sus propias cargas negativas.
Proteínas: Encontradas en Methanomicrobium
y Desulfurococcus, se trata de
una pared compuesta exclusivamente por proteínas de superficie conocidas como
la capa S (König, 1988).
7. El citoplasma
|| Regresar al índice ||
El
citoplasma de una célula es la parte que se encuentra en el interior de la
membrana celular. Está compuesto por el citosol y el citoesqueleto. El citosol
comprende todas las sustancias de “flotan” junto con el medio en que “flotan”
que es en su mayor parte agua. Las sustancias que flotan son burbujas de
diferentes tipos de grasas o sustancias afines, así como otros compuestos
químicos que pueden mezclarse con el agua como los azucares “carbohidratos” y las
proteínas.
En el
citosol se llevan a cabo muchas reacciones químicas, aunque su principal
función es la de almacenar los materiales necesarios para que la célula pueda
crecer y reproducirse. En los procariótas el genoma se encuentra en flotando en
el citosol en una región denominada nucleoide. El citoesqueleto en los
procariotas no está bien desarrollado, prueba de ello es que no fue reportado
como parte de los procariotes hasta la presente década “2010-2020”. Su
presencia es más importante en los eucariotas donde será analizado de forma
independiente al citoplasma.
Algunas
partes menores del citoplasma son:
7.1 Nucleóide
La palabra nucleóide significa semejante a
un núcleo. Es una región con forma irregular al interior de la célula
procariota que contiene todo o la mayoría del material genético, llamado
genóforo. En contraste con el núcleo verdadero de la célula eucariota el
nucleóide no se encuentra rodeado por una membrana nuclear. El nucleóide
puede verse con claridad por medio del microscopio electrónico de gran aumento,
donde, aunque su apariencia puede ser variable, es claramente visible contra el
citosol. El nucleóide responde compactándose y probablemente protegiendo el
material genético cuando se expone a la célula procariota a condiciones de daño
genético como radiación ultravioleta.
7.2 Glicocalix
Literalmente significa manto de azúcar, se
trata de una red de polisacáradios que se proyectan desde la superficie de la
célula y sirve como un medio de unión con otras bacterias en suspensión,
creando de esta manera microcolonias suspendidas que al unirse a una superficie
inician la generación de una biopelícula. También es generado por algunas
células eucariotas como parte de su matrix extracelular protectora.
7.3 Inclusiones
Las inclusiones citoplasmáticas son
sustancias no vivas que no son capaces de realizar actividades metabólicas y no
se encuentran unidas a las membranas. Las inclusiones son mecanismos de
almacenamiento de nutrientes, productos se secreción o gránulos de pigmento en
fase cristalina.
7.4 Tilacoides
Es un compartimento rodeado por membrana al interior de los cloroplastos y las cianobacterias. Estos son los sitios donde se realizan las reacciones dependientes de la luz de la fotosíntesis. Los tilacoides consisten en una membrana tilacoidal y un lumen interno. Los tilacoides por lo general se organizan en discos apilados verticalmente denominados grana.

Figura 7.1. Los tilacoides son membranas internas.
Los discos de la grana no son
independientes, se conectan por proyecciones de la membrana generando una única
entidad funcional. Los tilacoides son estructuras biológicamente relevantes por
dos razones, la primera y la más obvia es porque allí se realizan las
reacciones dependientes de la luz de la fotosíntesis bacteriana, y la segunda
porque son un sistema de membranas internas que ocurren en células
procarióticas.
7.5 Vacuolas
En efecto no es un error, se han detectado
tres géneros de bacterias sulfurosas filamentosas que lo poseen, siendo
estas: Thioploca, Beggiatoa y Thyomargarita. En
estos géneros el citosol se encuentra reducido y cerca del 40-98% del volume de
la célula es ocupado por la vacuola. Las vacuolas contienen altas
concentraciones de iones nitrato y por lo tanto se piensa que se trata de un
organelo de almacenamiento de nutrientes. Algunas bacterias poseen vacuolas
gaseosas, como en algunas especies de cianobacterias, las cuales ayudan a la
célula a controlar su flotabilidad (Kalanetra, Huston, & Nelson,
2004).
7.6 Vesículas
y tráfico vesicular
A diferencia de lo que sucede en los
eucariotas, el tráfico de vesículas membranosas es un área emergente de estudio
en la biología de los procariótas, la cual es relevante para el sistema de
sensibilidad de densidad poblacional de las bacterias formadoras de películas,
así como para las especies patógenas con un complejo sistema de señalización de
superficie patógeno-anfitrión.
Por más de cuatro décadas, los cultivos de
bacterias gramnegativas reveló la presencia de vesículas rodeadas por membranas
a nivel nanomolecular. La importancia de estas muicrovesículas en la patogénesis
se ha sospechado desde la década de 1970 donde fueron observadas en placa
gingivial dental por medio del microscopio electrónico. En la actualidad se ha
confirmado que las microvesículas son importantes en los sistemas de
colonización de muchas bacterias patógenas como Pseudomonas aeruginosa, Salmonella y Helicbacter pylori (Mashburn‐Warren & Whiteley,
2006).
El descubrimiento de las microvesículas es
especialmente relevante ya que conectan directamente con las hipótesis
evolutivas de los eucariotas, pues ya no se trataría de un rasgo presente y
ausente, sino de un rasgo que se encuentra en todos los tipos celulares con
diferentes grados de sofisticación.
7.7 Nucleolo
Es mejor conocido como el sitio de la
biogénesis (fabricación) de los ribosomas, que es la síntesis de los ribosomas.
El nucléolo también participa en la formación de partículas de reconocimiento
de señales y desempeña un papel en la respuesta celular al estrés.
Tradicionalmente era reconocido como una estructura del núcleo de la célula eucariota,
sin embargo, investigaciones recientes han demostrado su existencia en las arcaicas
(Islas-Morales, Cárdenas, Mosqueira,
Jiménez-García, & Voolstra, 2023).
8. El ribosoma procariota
|| Regresar al índice ||
Es el
responsable por la síntesis de proteínas en todas las células conocidas hasta
el momento.
8.1 Tamaño
Los
ribosomas son un complejo macromelecular “molécula muy, muy, muy grande” cuya
función es tan primordial para los seres vivos que bien pueden ser parte de la
definición misma de estar vivo. Todas las células poseen ribosomas, pero los
virus, viroides y priones no tienen ribosomas y no se considera que estén
vivos. Los ribosomas de los procariotas son más pequeños de los de los
eucariotas, la unidad para medir eso se denomina Svedberg (S) que corresponde a
un coeficiente de centrifugación. El Svedberg es una unidad para medir el
coeficiente de sedimentación de una partícula o macromolécula cuando son
centrifugados en condiciones normales. Esta magnitud tiene dimensiones de
tiempo, de modo que un Svedberg equivale a 10-13 segundos. En otras palabras,
mide cuanto tiempo se tarda una sustancia en una centrífuga para sedimentarse.
En las bacterias el ribosoma completo toma 70S para sedimentarse de forma
total, y cada una de sus dos partes por separado tarda 50S y 30S
respectivamente.
8.2 Anatomía
El
ribosoma está compuesto principalmente por ARN enredado de forma estrecha como
hilo enredado formando un nudo. La forma del nudo está perfectamente
reglamentada y es la que le otorga su función. Al ARN que forma el ribosoma se
lo denomina ARN ribosómico o ribosomal “ARNr”. El centro que realiza la función
del ribosoma o región activa tiene sitios de inserción para el ARNm que viene
del genoma y para otro tipo de ARN que proviene del citoplasma llamado ARN de
transferencia. En general el ribosoma consta de dos unidades que se cierran
entre sí “aplastando” al ARNm y leyéndolo como si fuera una máquina de
escribir. El ribosoma también cuenta con algunas secuencias semejantes a las
proteínas, pero más cortas llamadas péptidos.
8.3 Fisiología
La función de los ribosomas ya sea de procariotas o eucariotas es la de transformar la información del ARNm a proteínas. Para hacerlo las dos subunidades del ribosoma se cierran sobre el ARNm permitiendo que este entre en contacto con su región activa. Una vez allí el código genético es leído por el ARN de transferencia de forma tal que este transfiere una unidad de proteínas llamada aminoácidos.

Figura 8.1. En el modelo
anterior podemos ver el complejo de producción de proteínas. En café el
ribosoma, el rojo el ARNm, en verde el ARNt y en morado los aminoácidos. Cuando
la proteína es muy corta (menos de 100 aminoácidos) se la llama polipéptido.
A medida
que la lectura progresa, el ARN de transferencia no solo transfiere
aminoácidos, también los va pegando de forma tal que se va generando una cadena
muy larga. Cuando la cadena de aminoácidos crece lo suficiente se enreda como
su fuera un nudo de hilos, formando la proteína. Al igual que con el ribosoma,
la función de la proteína depende de la forma que tenga el nudo.
9. Los retículos endoplasmáticos eucariotas
Los
retículos endoplasmáticos son bolsas membranosas muy largas que se encuentran
muy cercanas al núcleo de la célula. La estructura general del RE es la de una
red de membranas denominadas cisternas. Estas estructuras semejantes a sacos
son mantenidos en su posición gracias al citoesqueleto. La membrana
cisternal rodea un espacio interno denominado lumen. El lumen del RE es
continuo con el espacio perinuclear por lo que se puede visualizar al RE como
proyecciones del núcleo y por ende el RE siempre estará asociado al núcleo
celular. El retículo endoplasmático rugoso “RER” se encuentra conectado
directamente al núcleo a través de la membrana nuclear, lo que le permite
acceso rápido a los ARNm que provienen del genoma. El RER se denomina rugoso
porque contiene muchos ribosomas, y la función de los ribosomas es capturar el
ARNm para producir proteínas.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figura 9.1. En esta imagen
de microscopio electrónico podemos ver al núcleo rodeado por las membranas del
retículo endplasmático rugoso.
El retículo endomplasmático liso “REL”
está adyacente al núcleo, pero no está conectado directamente al núcleo, y
tampoco posee ribosomas. Por esta razón no puede crear proteínas, sus funciones
están relacionadas con el metabolismo de las grasas, por lo que se encuentra
asociado a las mitocondrias.
9.1 Retículo
endoplasmático rugoso
La superficie del retículo endoplasmático rugoso RER esta tapizado con los ribosomas que se encargan de la síntesis de proteínas. El punto de unión entre el RER y los ribosomas se denomina traslocón. Los ribosomas no hacen parte integral de la membrana, por lo que pueden unirse o liberarse dependiendo de las condiciones celulares. Un ribosoma solo se acopla al RER una vez que una estructura proteínica se une a un ácido nucleico en el citosol. El ribosoma se acopla de hecho, no por una estructura propia, sino por los primeros 30 aminoácidos que sintetiza al inicio de una síntesis de proteínas. Una vez que estos 30 aminoácidos se acoplan al traslocon, la síntesis de la proteína continua. Pero antes de continuar una peptidasa corta esta primera cola peptídica para permitir que la verdadera proteína pueda sintetizarse.

Figura 9.2. El RER está más
cerca del núcleo y tiene ribosomas, mientras que el REL está más lejos y no los
tiene.
El ribosoma se encuentra en el citosol, y el traslocon funciona
como un canal en el que el ribosoma inyecta la proteína en formación hasta que
termina la síntesis y una nueva peptidasa corta la unión entre el péptido de
unión al traslocon y la mitocondria, de forma tal que se libere el ribosoma, el
péptido de inicio y la proteína del traslocon. El RER es clave para las
siguientes funciones: Fabrica de lisosomas, fábricas de proteínas para
secreción, síntesis de proteínas integrales de las vesículas que les permiten
acoplarse con sus blancos específicos en una cadena de transporte a través del
citoesqueleto, e inicio de la glicosilación de algunos péptidos inmaduros.

Figura 9.3. El RER adquiere
los mensajes del núcleo “mRNA” y emplea los ribosomas para convertir esos
mensajes en polipeptidos, las cuales quedan en su interior.
9.2 Retículo
endplasmático liso
El retículo entoplasmático liso REL posee funciones metabólicas diversas. Sintetiza péptidos, fosfolípidos y esteroides, por lo que en general trabaja con la grasa celular. Las células que segregan estos productos como los testículos, los ovarios y las células de las glándulas cebaseas tiene un REL grande.

Figura 9.4. El retículo
endoplasmático liso es continuo con el rugoso, pero carece de ribosomas.
El REL también se encarga del metabolismo
de los carbohidratos, la desintoxicación de algunos metabolitos secundarios
como el metanol y muchos fármacos. En las células musculares se encarga de
regular la concentración del calcio, valor crucial para la contracción del
sarcómero muscular. El REL está tan modificado en las células musculares que
recibe el nombre de retículo sarcoplasmático. El REL también posee la enzima
glucosa-6-fosfatasa que convierte ña glucosa-6-gosgato en glucosa, un paso
crucial en la gluconeogénesis que permite recuperar los productos de la
fermentación nuevamente en glucosa.
9.3 Plegamiento
de proteínas
El RE sirve para funciones diversas,
incluyendo el plegamiento de las proteínas, su almacenamiento al interior de la
cisterna reticular, el transporte de las proteínas inmaduras por evaginación de
la membrana reticular para formar vesículas y su transporte al aparato de
Golgi.
El plegamiento de las proteínas no solo se
deja a la espontaneidad de las interacciones moleculares de los péptidos, un
grupo de proteínas especializadas llamadas chaperonas se encargan de regular el
plegamiento correcto de los péptidos a proteínas inmaduras. Únicamente las
proteínas plegadas de forma adecuada son exportadas al aparato de Golgi. Si el
número de proteínas no plegadas adecuadamente aumenta, el RE ingresa en un
estado de estrés que puede desencadenar mutaciones, hipoxia o la muerte celular
programada.
10. El aparato de Golgi eucariota
|| Regresar al índice ||
Su forma es semejante a la de los retículos endoplasmáticos, pero se diferencia en que no está cerca de núcleo. El aparato de Golgi es un órgano dinámico, como si se tratara de una terminal de transportes. Comparación entre los resultados experimentales "izquierda, microfotografía" con el modelo teórico esquemático.

{kind=link}
Figura 10.1. El sistema de
membranas internas es dinámico, pues secciones de sistemas grandes como el RER,
el REL, el Golgi, o la membrana celular pueden separarse y viajar en forma de
vesículas a otras regiones, cumpliendo diversas funciones.
En este caso los vehículos son
membranas internas llamadas vesículas, son muy pequeñas y tienen forma
esférica. Las vesículas se forman por separación de una membrana más grande.
Existen muchas rutas desde el RER al aparato de Golgi y viceversa. Desde el
aparato de Golgi a la membrana celular y viceversa. Desde el aparato de Golgi a
otras vesículas. Internamente esta terminal también es capaz de realizar
transformaciones complejas de los materiales provenientes desde el RER y del
exterior. Un ejemplo, combinando una proteína del RER y un carbohidrato del
exterior puede formar glucoproteínas “algunas glucoproteínas se insertan en la
membrana y sirven para identificar la célula, ese es el principio del sistema
de identificación de sangre AB0.
10.1 Descubrimiento
Debido a su gran tamaño y forma distintiva el aparato de Golgi fue uno de los primeros organelos celulares en ser descubiertos y observados con algún detalle. Fue descubierto en 1898 por el fisiólogo italiano Camilo Golgi durante una investigación del sistema nervioso. Cuando lo observó bajo su microscopio Golgi lo nombró como el aparato reticular interno. Algunos cuestionaron su descubrimiento argumentando que no lo vio realmente y que solo se trataba de un artefacto de su microscopio. Con el desarrollo del microscopio moderno en el siglo XX el descubrimiento de Camilo Golgi fue confirmado. El aparato de Golgi fuer acuñado con dicho nombre hasta 1913, antes del cual recibió nombres diversos como ductos de Golgi-Holmgren y aparato de Golgi-Kopsch.
10.2 Localización
Al interior de los eucariotas la
localización del aparato de Golgi difiere. En los mamíferos un solo aparato de
Golgi se ubica generalmente cerca al núcleo de la célula y del centrosoma.
Conexiones tubulares de membrana son responsables de mantener a las cisternas
de Golgi unidas. Pero en general al igual que con el RE la estructura y
localización del aparato de Golgi es dependiente del citoesqueleto. En oros
eucariotas como las levaduras el aparato de Golgi no está unido y forma
vesículas independientes.

{kind=link}
Figura 10.2. Bartolomeo Camillo Emilio Golgi (Corteno Golgi, Italia, 7 de julio de
1843 - Pavía, 21 de enero de 1926) fue un médico y citólogo italiano. Ideó los
métodos de tinción celular a base de cromato de plata, procedimiento que
permitió (tanto a él mismo como a otros investigadores) realizar importantes
descubrimientos, especialmente acerca de las neuronas y su fisiología. Recibió
el Premio Nobel de Medicina (con el español Santiago Ramón y Cajal) en 1906.
10.3 Estructura
En la mayoría de los eucariotas el aparato
de Golgi consiste en una serie de compartimentos consistente en dos redes
principales. La red Golgi cis (RGC) y la res Golgi trans (RGT). La RGC es una
colección de membranas aplanadas y parcialmente fusionadas de discos conocidos
como cisternas que se originan de clusteres de vesículas que se originan del
RE. Cada cisterna se fusiona parcialmente con cuatro a ocho más para
formar un saco, pero en algunos protistas pueden formarse sacos de hasta 16 cisternas.
Estas colecciones de cisternas se rompen en compartimentos cis, medios y trans.
La RGT es la estructura final de la cisterna, desde la cual las proteínas son empaquetadas en vesículas destinadas a ser lisosomas, vesículas secretoras para transportar proteínas integrales de la membrana externa. Existen algunas diferencias en la organización del aparato de Golgi al interior de los eucariotas, por ejemplo, en las levaduras el acoplamiento de las cisternas no se observa y en su lugar permanecen como grandes vesículas independientes. El aparato de Golgi tiende a ser grande y más numeroso en las células que se especializan en la síntesis y secreción de sustancias como las células B secretoras de anticuerpo del sistema inmune.

Figura 10.3. El aparato de
Golgie está hecho de muchas membranas cercanas.
En todos los eucariotas cada saco cis de
ingreso vesicular se contrapone a una cara trans de emisión vesicular. Cada
cara se caracteriza por una micromorfología y una composición proteínica diferentes.
Adicionalmente hay evidencia de que el aparato de Golgi posee rutas
compartimentadas que permiten mantener cadenas de montaje para la modificación
de diferentes proteínas de forma independiente, del mismo modo que una fábrica
de autos puede tener líneas de montaje diferentes para modelos.
10.4 Funciones
De cierta manera la célula puede
visualizarse como una fábrica que se especializa en la fabricación de
proteínas, dichas proteínas se convierten en los trabajadores y la maquinaria
para fabricar más proteínas. En este orden de ideas el aparato de Golgi se
encarga de la modificación y maduración de las proteínas, siendo el equivalente
a una fábrica de autos a las partes finales de la cadena de montaje donde se
pinta y se pone el auto full equipo.
Una vez que el aparato de Golgi ha
finalizado de ajustar las proteínas, las empaqueta en vesículas de emisión, que
funcionan como los camiones de despacho de una fábrica a las zonas de
exhibición. En este caso las zonas de exhibición son la membrana celular o el
exterior de la célula.
Debido a que el aparato de Golgi está
involucrado en la maduración de las proteínas sus funciones estarán
relacionados a procesos bioquímicos de adición de grupos: glicosilación en la
cual se agregan polísacaridos u oligosacáridos; sulfatación en la que se
adicionan grupos sulfato; fosforilación en la que se adicionan grupos fosfato;
y lisis en la que un péptido muy largo se divide en varias proteínas semejantes
independientes.
11. El núcleo celular eucariota
|| Regresar al índice ||
El núcleo celular es el organelo que les da el nombre a las células eucarióticas. El nombre eucariota significa en el contexto científico, células con núcleos verdaderos. La función del núcleo celular es almacenar el genoma. Esto las diferencias de las células procariotas en las que no puede detectarse una membrana interna especializada en almacenar el genoma.

{kind=link}
Figura 11.1. El núcleo es un
organelo especial, pues su membrana es doble, y parte de ella constituye al
propio RER, además posee grandes poros.
Es por esto que células procariotas con
sistema de membranas internas como las cianobacterias se clasifican como
procariotas, pues en el caso de las cianobacterias ningún tilacoide almacena su
genoma. El núcleo celular está compuesto por dos membranas celulares, una
interna y otra externa. Ambas membranas se unen en regiones llamados poros. Es
por esto que es posible afirmar que el retículo endoplasmático rugoso hace
parte estructural de la membrana del núcleo. La membrane externa del núcleo
celular es continua con la membrana del retículo endoplasmático, sin embargo,
aunque están unidos físicamente cada una de las membranas poseen un diferente
contenido de proteínas, lo cual diferencia su funcionamiento.
Al interior de la membrana interna se
encuentra el nucleoplasma el cual contiene al genoma eucariótico. Cuando la
célula se divide una región del núcleo se tiñe densamente y se llama nucleolo,
su función primordial es la de ensamblar nuevos ribosomas. La función del
núcleo es almacenar el genoma, aislándolo del resto de la célula. El núcleo
solo existe cuando la célula está trabajando, es decir en las fases de
crecimiento “G1 y G2” y en la fase de síntesis “S” del ciclo celular. En su
interior NO se almacenan los cromosomas, lo que se almacena es la cromatina ya
sea para replicar el ADN que contiene o para generar ARNm. Debido a que el
núcleo contiene la mayor parte del genoma de la célula, produce mucho ARNm que
es enviad a través de los poros nucleares hacia el retículo endoplasmático
rugoso.
El núcleo es el orgánulo más grande en las
células animales. En las células de mamíferos, el diámetro promedio del núcleo
es de aproximadamente 6 micrómetros (µm), que ocupa aproximadamente el 10% del
volumen celular total. El contenido del núcleo se mantiene en el nucleoplasma
de manera similar al citoplasma en el resto de la célula. El componente fluido
de esto se denomina nucleosol, similar al citosol en el citoplasma. En la
mayoría de los tipos de granulocitos, un glóbulo blanco, el núcleo está lobulado
y puede ser bilobulado, trilobulado o multilobulado.
11.1 Envoltura nuclear
La envoltura nuclear, también conocida como membrana nuclear, consta de dos membranas celulares, una membrana interna y otra externa, dispuestas paralelas entre sí y separadas por 10 a 50 nanómetros (nm). La envoltura nuclear encierra completamente el núcleo y separa el material genético de la célula del citoplasma circundante, sirviendo como barrera para evitar que las macromoléculas se difundan libremente entre el nucleoplasma y el citoplasma. La membrana nuclear externa es continua con la membrana del retículo endoplásmico rugoso (RER), y está tachonada de manera similar con ribosomas. El espacio entre las membranas se llama espacio perinuclear y es continuo con la luz RER.

Figura 11.2. La membrana
nuclear es una estructura semicontínua con el retículo endoplasmático rugoso.
11.2 Los poros nucleares
Proporcionan canales acuosos a través de la envoltura, están compuestos por múltiples proteínas, denominadas colectivamente nucleoporinas. Los poros tienen un peso molecular de aproximadamente 125 millones de daltons y consisten en alrededor de 50 (en levadura) a varios cientos de proteínas (en vertebrados). Los poros tienen un diámetro total de 100 nm; sin embargo, la brecha a través de la cual las moléculas se difunden libremente tiene solo unos 9 nm de ancho, debido a la presencia de sistemas reguladores dentro del centro del poro. Este tamaño permite selectivamente el paso de pequeñas moléculas solubles en agua al tiempo que evita que moléculas más grandes, como ácidos nucleicos y proteínas más grandes, entren o salgan inapropiadamente del núcleo. Estas moléculas grandes deben ser transportadas activamente al núcleo en su lugar. El núcleo de una célula de mamífero típica tendrá alrededor de 3000 a 4000 poros en toda su envoltura, cada uno de los cuales contiene una estructura en forma de anillo simétrica de ocho veces en una posición donde las membranas internas y externas se fusionan. Se une al anillo una estructura llamada canasta nuclear que se extiende hacia el nucleoplasma y una serie de extensiones filamentosas que llegan al citoplasma. Ambas estructuras sirven para mediar la unión a las proteínas de transporte nuclear.

Figura 11.3. Denominado
complejo del poro nuclear, el poro es una estructura compleja que media el paso
de sustancias de manera selectiva.
La mayoría de las proteínas, las
subunidades ribosómicas y algunos ADN se transportan a través de los complejos de
poros en un proceso mediado por una familia de factores de transporte conocidos
como carioferinas. Esas carioferinas que median el movimiento hacia el núcleo
también se llaman importinas, mientras que las que median el movimiento fuera
del núcleo se llaman exportinas. La mayoría de las carioferinas interactúan
directamente con su carga, aunque algunas usan proteínas adaptadoras. Las
hormonas esteroides como el cortisol y la aldosterona, así como otras pequeñas
moléculas solubles en lípidos involucradas en la señalización intercelular,
pueden difundirse a través de la membrana celular y hacia el citoplasma, donde
se unen a las proteínas del receptor nuclear que son traficadas hacia el
núcleo. Allí sirven como factores de transcripción cuando se unen a su ligando;
en ausencia de un ligando, muchos de estos receptores funcionan como
desacetilasas de histonas que reprimen la expresión génica.
11.3 La lámina nuclear
En las células animales, dos redes de
filamentos intermedios proporcionan al núcleo un soporte mecánico: la lámina
nuclear forma una malla organizada en la cara interna de la envoltura, mientras
que se proporciona un soporte menos organizado en la cara citosólica de la
envoltura. Ambos sistemas proporcionan soporte estructural para la envoltura
nuclear y los sitios de anclaje para cromosomas y poros nucleares.
La lámina nuclear está compuesta principalmente de proteínas laminares. Como todas las proteínas, las láminas se sintetizan en el citoplasma y luego se transportan al interior del núcleo, donde se ensamblan antes de incorporarse a la red existente de láminas nucleares. Las láminas que se encuentran en la cara citosólica de la membrana, como emerina y nesprina, se unen al citoesqueleto para proporcionar soporte estructural. Las láminas también se encuentran dentro del nucleoplasma, donde forman otra estructura regular, conocida como velo nucleoplasmático, que es visible mediante microscopía de fluorescencia. La función real del velo no está clara, aunque está excluida del nucleolo y está presente durante la interfase. Las estructuras laminares que forman el velo, como LEM3, se unen a la cromatina e interrumpen su estructura inhiben la transcripción de genes codificadores de proteínas.

Figura 11.4. La lámina
nuclear es un mediador proteínico entre la envoltura y la cromatina.
Al igual que los componentes de otros
filamentos intermedios, el monómero lamina contiene un dominio alfa-helicoidal
utilizado por dos monómeros para enrollarse entre sí, formando una estructura
de dímero llamada bobina enrollada. Dos de estas estructuras de dímero se unen
una al lado de la otra, en una disposición antiparalela, para formar un
tetrámero llamado protofilamento. Ocho de estos protofilamentos forman una
disposición lateral que se tuerce para formar un filamento similar a una
cuerda. Estos filamentos pueden ensamblarse o desmontarse de manera dinámica,
lo que significa que los cambios en la longitud del filamento dependen de las
tasas competitivas de adición y eliminación de filamentos.
Las mutaciones en los genes laminares que
conducen a defectos en el ensamblaje de filamentos causan un grupo de
trastornos genéticos raros conocidos como laminopatías. La laminopatía más
notable es la familia de enfermedades conocidas como progeria, que causa la
aparición de envejecimiento prematuro en sus víctimas. El mecanismo exacto por
el cual los cambios bioquímicos asociados dan lugar al fenotipo envejecido no
se conoce bien (Mounkes & Stewart, 2004).
11.4 Los cromosomas
El núcleo celular contiene la mayoría del
material genético de la célula en forma de múltiples moléculas lineales de ADN
organizadas en estructuras llamadas cromosomas. Cada célula humana contiene
aproximadamente dos metros de ADN. Durante la mayor parte del ciclo celular,
estos se organizan en un complejo de ADN-proteína conocido como cromatina, y
durante la división celular se puede ver que la cromatina forma los cromosomas
bien definidos familiares de un cariotipo. En cambio, una pequeña fracción de
los genes de la célula se encuentra en las mitocondrias.

Figura 11.5. Los cromosomas
son cromatina empacada y duplicada, se forman para poder separar la cromatina
durante la reproducción. Como el ADN está empacado, se encuentra inactivo.
Hay dos tipos de cromatina. La eucromatina
es la forma de ADN menos compacta y contiene genes que la célula expresa con
frecuencia. El otro tipo, la heterocromatina, es la forma más compacta y
contiene ADN que se transcribe con poca frecuencia. Esta estructura se
clasifica además en heterocromatina facultativa, que consiste en genes que se
organizan como heterocromatina solo en ciertos tipos celulares o en ciertas
etapas de desarrollo, y heterocromatina constitutiva que consiste en
componentes estructurales cromosómicos como telómeros y centrómeros. Durante la
interfase, la cromatina se organiza en parches individuales discretos, llamados
territorios cromosómicos. Los genes activos, que generalmente se encuentran en
la región eucromática del cromosoma, tienden a ubicarse hacia el límite del
territorio del cromosoma.
Los anticuerpos contra ciertos tipos de
organización de la cromatina, en particular los nucleosomas, se han asociado
con una serie de enfermedades autoinmunes, como el lupus eritematoso sistémico.
Estos se conocen como anticuerpos antinucleares (ANA) y también se han observado
junto con la esclerosis múltiple como parte de la disfunción general del
sistema inmunitario. Como en el caso de la progeria, el papel desempeñado por
los anticuerpos en la inducción de los síntomas de las enfermedades autoinmunes
no es obvio (Zhang & Zhang, 2015).
11.5 El nucléolo
El nucleolo es la más grande de las
discretas estructuras sin membrana, densamente teñidas, conocidas como cuerpos
nucleares que se encuentran en el núcleo. Se forma alrededor de repeticiones en
tándem de ADNr, ADN que codifica el ARN ribosómico (ARNr). Estas regiones se denominan
regiones organizadoras nucleolares (NOR). Los roles principales del nucleolo
son sintetizar rRNA y ensamblar ribosomas. La cohesión estructural del nucleolo
depende de su actividad, ya que el ensamblaje ribosómico en el nucleolo da como
resultado la asociación transitoria de los componentes nucleolares, facilitando
el ensamblaje ribosómico adicional y, por lo tanto, la asociación adicional.
Este modelo está respaldado por observaciones de que la inactivación del ADNr
resulta en la mezcla de estructuras nucleolares.
En el primer paso del ensamblaje de ribosomas, una proteína llamada ARN polimerasa I transcribe ADNr, que forma un gran precursor de ARNt previo. Esto se divide en las subunidades 5.8S, 18S y 28S rRNA. La transcripción, el procesamiento postranscripcional y el ensamblaje del rRNA se produce en el nucleolo, ayudado por pequeñas moléculas de ARN nucleolar (snoRNA), algunas de las cuales se derivan de intrones empalmados de ARN mensajeros que codifican genes relacionados con la función ribosómica. Las subunidades ribosómicas ensambladas son las estructuras más grandes que pasan a través de los poros nucleares.

Figura 11.6. El nucléolo es
el punto de ensamblaje de los ribosomas.
Cuando se observa bajo el microscopio electrónico,
se puede ver que el nucleolo consta de tres regiones distinguibles: los centros
fibrilares más internos (FC), rodeados por el componente fibrilar denso (DFC)
(que contiene fibrillarina y nucleolina), que a su vez está bordeado por el
componente granular (GC) (que contiene la proteína nucleofosmina). La
transcripción del ADNr se produce en el FC o en el límite de FC-DFC y, por lo
tanto, cuando aumenta la transcripción del ADNr en la célula, se detectan más
FC. La mayor parte de la escisión y modificación de los rRNA se produce en el
DFC, mientras que los últimos pasos que implican el ensamblaje de proteínas en
las subunidades ribosómicas se producen en el GC.
11.6 Otros cuerpos nucleares
Además del nucleolo, el núcleo contiene
varios otros cuerpos nucleares. Estos incluyen los cuerpos de Cajal, los
géminis de los cuerpos de Cajal, la asociación de cariosomas de interfase
polimórfica (PIKA), los cuerpos de leucemia promielocítica (LMP), los
parapeckle y las manchas de empalme. Aunque se sabe poco sobre varios de estos
dominios, son significativos porque muestran que el nucleoplasma no es una
mezcla uniforme, sino que contiene subdominios funcionales organizados.
Otras estructuras subnucleares aparecen
como parte de procesos anormales de la enfermedad. Por ejemplo, se ha informado
de la presencia de pequeñas varillas intranucleares en algunos casos de
miopatía por nemalina. Esta condición generalmente es el resultado de
mutaciones en la actina, y las barras en sí consisten en actina mutante, así
como otras proteínas del citoesqueleto (Mao, Zhang, & Spector, 2011).
12. Las vesículas celulares eucariotas
|| Regresar al índice ||
En biología celular una vesícula se define
como una estructura interna de la célula que consiste en un fluido plasmovesicular
rodeado por una membrana biológica. En la analogía de la fábrica las vesículas
generalmente funcionan como los camiones de carga o las grúas de carga que
transportan los materiales de una línea de montaje a otra, pero en ocasiones
pueden tener otros usos.
Existen
vesículas poco comunes compuestas por una sola lámina de fosfolípidos, estas
vesículas reciben el nombre de liposomas monolamelares y su contenido interno
debe ser exclusivamente de lípidos. La membrana que define la vesícula se
denomina fase lamelar. Las vesículas se
mueven de forma no estocástica, gracias a grúas del citoesqueleto de las mueven
a costo de ATP a través de las fibras, y además poseen proteínas de
reconocimiento de acoplamiento que les permiten reconocer sus blancos y
fusionarse a ellos. Los blancos de las vesículas son diversos y pueden ser
cualquier organelo y otras vesículas. A continuación, estudiaremos algunos
tipos de vesículas
12.1 Vacuolas
Las vacuolas contráctiles “estrellas” fueron descritas por Spallanzani en 1776 en algunos protozoos, aunque fueron malinterpretadas como órganos respiratorios. Dujardin en 1841 denominó estas estrellas como vacuolas. En 1842 Schleiden aplicó el término en la célula vegetal para distinguir el protoplasma vegetal de un espacio independiente y bastante grande. En 1885 Hugo de Vries denominó la membrana vacuolar como tonoplasto, ya que son responsables del tono de la planta.

{kind=link}
Figura 12.1. La vacuola es
el organelo más prominente en la célula vegetal, dado que es su relleno
sanitario.
Una
vacuola es un organelo definido por membrana que está presente en todas las
células vegetales, fúngicas, algunos protistas y algunos animales, inclusive
algunas células bacterianas pueden presentar vacuolas. Las vacuolas son
básicamente compartimentos para almacenar agua, sustancias en solución y
sustancias cristalizadas o en fase sólida.
Las
vacuolas no poseen un tamaño o forma definidas, y su estructura cambia de
acuerdo a las necesidades de la célula. La función e importancia de la vacuola
cambia mucho dependiendo del tipo de célula que se está analizando, aunque
tiene mayor prominencia en las células vegetales, fúngicas y en ciertos
protistas que en los animales y bacterias. Sus funciones son:
Aislamiento: El
aislamiento refiere a la imposibilidad de excretar externamente muchas sustancias
por parte de la célula, así que la vacuola funciona como un pozo séptico o
relleno sanitario. Allí se almacenan los desechos nitrogenados de la excreción
de las proteínas, así como los metabolitos secundarios provenientes de venenos,
fármacos u reacciones internas que salieron de control.
Agua y equilibrio
osmótico: Al contener agua, con diferentes niveles de
electrolitos disueltos como iones salinos y protones, lo cual ayuda a la planta
a regular el equilibrio osmótico entre el citoplasma y la matriz extracelular.
Tono: Mediante la simple presión y
volumen hidráulico el efecto sumatorio de la vacuola le da su rigidez a las
hojas y estructuras vegetales no ligninizadas.
Almacenamiento de
nutrientes: Solo
en las semillas donde las vacuolas se modifican para contener carbohidratos,
lípidos y proteínas.
Defensa: Las sustancias productos del
desecho y aisladas generalmente se modifican secundariamente, generando
metabolitos secundarios con efectos narcóticos o venenosos en los hervívoros.
Adicionalmente, pueden clasificarse dependiendo
del linaje filogenético en:
Vacuolas vegetales: La mayoría de las células vegetales poseen una vacuola única que ocupa
del 30% al 80% del volumen interno de la célula. Cumple todas las funciones
anteriormente descritas.
Vacuolas fúngicas: Realizan funciones semejantes a las vacuolas vegetales, pero son más
pequeñas y puede haber más de una por célula. En las levaduras la vacuola es
una estructura dinámica que puede alterar la forma de la célula.
Vacuolas animales: Las vacuolas animales son más
pequeñas de las vegetales, aunque usualmente son más numerosas, aunque también
existen células animales sin vacuolas. En los animales las vacuolas están
subordinadas al tráfico de vesículas en la ruta de exocitosis y en la ruta de endocitosis.
12.2 Lisosomas
Es un
organelo rodeado por membrana que se encuentra en la mayoría de las células
animales. Se trata de vesículas esféricas que contienen enzimas hidrolíticas
que pueden romper virtualmente cualquier tipo de biomoléculas a excepción de
las membranas, de lo contrario el lisosoma colapsaría y las enzimas quedarían
libres para destruir la célula.
Dicho en
términos simples, un lisosoma es un tipo de vesícula con una composición muy
específica, tanto de las proteínas de membrana como de las proteínas de su
lumen. El pH del lumen es ácido cuando se lo compara con el del citplasma,
llegando a rondar los valores de 4.5-5,0 lo cual lo hace análogo a la función
del estómago. A demás de la degradación de polímeros, los lisosomas están
involucrados en varios procesos celulares incluyendo la secreción de hormonas,
la reparación de la membrana, el metabolismo energético y principalmente en el
sistema inmune.
Los
lisosomas también actúan como un sistema de eliminación de las sustancias que
no son absorbidas por parte de la célula. Un descubrimiento reciente refiere a
la capacidad de los lisosomas de consumir fragmentos celulares desgastados,
proceso denominado autofagia. Los tamaños de los lisosomas pueden variar,
siendo los más grandes 10 veces más voluminosos que los más pequeños. Ellos
fueron aislados y nombrados por el biólogo Belga Christian de Duve, quien
eventualmente recibiría el premio nobel de fisiología y medicina en 1974.
Los
lisosomas son conocidos por contener más de 50 diferentes enzimas líticas, las
cuales son producidas originalmente en el RER, transportadas al aparato de
Golgi. Los lisosomas permanecerán inactivos mientras que su pH interno sea
cercano al 7,0, sin embargo, al fusionarse con vesículas acídicas las enzimas
se activan. La síntesis de los lisosomas está regulada por genes del núcleo.
Mutaciones en los genes de estas enzimas son responsables por más de 30
diferentes enfermedades genéticas humanas, las cuales colectivamente reciben el
nombre de enfermedades de almacenamiento del lisosoma. Estos defectos están
relacionados a desordenes neurodegenerativos, cáncer, enfermedades cardíacas y
enfermedades relacionadas al síndrome de envejecimiento.
12.3 Peroxisomas
Los
peroxisomas son organelos encontrados virtualmente en todas las células
eucarióticas. Se encuentran involucrados en el catabolismo de ácidos grasos de
cadenas muy largas, ácidos grasos de cadena ramificada. Aminoácidos de giro
derecho, poliaminas, reducción de moléculas oxidantes y superoxidantes, así
como la síntesis de plasmógenos, que son otros fosfolípidos críticos para el
funcionamiento normal del cerebro y los pulmones en los mamíferos. También
contienen el 10% del total de la actividad de dos enzimas en la ruta de las
pentosas fosfato, la cual es importante en el metabolismo energético.
Los peroxisomas
fueron aislados e identificados por el biólogo Belga Christian de Duve en 1967
de forma definitiva, aunque originalmente habían sido descritos por el
estudiante de doctorado sueco J. Rhodin en 1954. Una de las principales
funciones de los peroxisomas es el rompimiento de los ácidos grasos de cadenas
muy largas mediante la betaoxidación. En las células animales, los ácidos
grasos de cadenas muy largas son convertidos a ácidos grasos de cadenas de
tamaño medio, los cuales son enviados a la mitocondria para ser introducidos en
la ruta del ciclo de Krebs, convirtiéndose en moléculas de dióxido de carbono y
energía metabólica.
Los
peroxisomas contienen enzimas oxidativas como la catalasa, la oxidasa de ácidos
D-aminos y la oxidasa de ácido úrico. Aunque la última enzima está ausente en
los seres humanos, lo cual explica el surgimiento de la enfermedad conocida
como la gota, generada por la acumulación de ácido úrico. Ciertas enzimas en el
peroxisoma remueven átomos de hidrógeno de sustancias específicas en una
reacción oxidativa empleando radicales libres superóxido lo cual conlleva a la
producción de peróxido de hidrógeno, que es toxico, pero no tanto como un
radical superoxido. El peróxido de hidrógeno luego sirve de sustrato para la
catalasa para oxidar a otras sustancias como fenoles, ácido fórmico,
formaldehido y alcoholes. Este conjunto de reacciones es importante ya que
desintoxica al cuerpo de muchas sustancias que son potencialmente venenosas.
En las
plantas los peroxisomas son aún más eficientes, pues contienen enzimas extra
como la dismutasa de superoxidos, componentes del ciclo del ácido
ascorbico-glutationa, así como deshidrogenasas de NADP de la ruta de las
pentosas fosfato.
12.4 Liposomas
Los
liposomas son vesículas esféricas que pueden contener más de una bicapa
lipídica. Los liposomas pueden emplearse como un vehículo de administración de
nutrientes y drogas farmacéuticas. Los liposomas pueden ser preparados mediante
la disrupción de las membranas. Se trata de vesículas artificiales principalmente.

Figura 12.2. Las vesículas
son partes de organelos mas grandes, y se mueven entre ellos y se fusionan con
ellos, las diferentes funciones no se deben a sus formas, pues todas son
burbujas de grasa al interior de la célula, lo que da sus funciones son las
proteínas que están en sus membranas, y su relación con otras vesículas y el
citoesqueleto que las mueve.
12.5 Fagosoma
Los
fagosomas son vesículas formadas alrededor de una partícula absorbida mediante
la fagocitosis. La vesícula luego puede fusionarse con lisosomas y peroxisomas
para su degradación, sea con el objetivo de obtener nutrientes o para combatir infecciones
de forma activa o pasiva. Una vez que el fagosoma se ha fusionado con los
lisosomas se denomina fagolisosoma.
Algunas bacterias que invaden activamente
el interior de las células a través de los fagosomas son capaces de inactivar
las enzimas de los lisosomas, invertir el flujo de nutrientes desde el
citoplasma hacia el fagosoma e incluso de reproducirse en su interior. Ejemplos
de etsas bacterias son including Mycobacterium
tuberculosis, Mycobacteria avium
paratuberculosis, Coxiella spp., y Rickettsia spp.
12.6 Vesículas
secretoras
Las vesículas secretoras contienen
materiales que deben ser excretados al exterior de la célula. Las células
tienen muchas razones para excretar o secretar materiales. Una razón es la
eliminación de desechos no absorbidos, mientras que en otras ocasiones se trata
de sustancias que permite el normal funcionamiento de las comunidades
celulares, por lo que el contenido excretado puede tratarse de hormonas. Las
vesículas sinápticas se localizan en las terminales presinápticas de las
neuronas, y almacenan neurotransmisores o neurohormonas. Cuando una señal
activa el axón, las vesículas sinápticas se fusionan con la membrana de la
sinapsis liberando su contenido para transmitir la señal a las neuronas vecinas
que también iniciaran un pulso eléctrico y realizaran sus funciones.
Las glándulas endocrinas y exocrinas deben
su función a las vesículas que almacenan, transportan y liberan las sustancias,
ya sean hormonas o fluidos especializados como el moco. Un buen ejemplo de esto
se encuentra en las isletas de Langerhans en el páncreas. Este tejido contiene
muchos tipos de células que se definen en base a qué tipo de hormonas producen
de forma especializada. Las bacterias, las arcaicas y muchos otros eucariotas
parasíticos liberan vesículas que contienen variados y especializados
compuestos tóxicos, y señales buioquímicas para atacar a sus anfitriones e
inhabilitar sus defensas.
12.7 Vesículas
extracelulares
Son producidas por todos los dominios de
la vida. Se trata de vesículas que pueden emitirse al ambiente externo, las
cuales contienen una amplia diversidad de sustancias, y algunas de ellas pueden
contener incluso ADN (Fevrier & Raposo, 2004; Grenier
& Mayrand, 1987; Raposo & Stoorvogel, 2013).
12.8 Otros
tipos de vesículas
Las vesículas gaseosas son empleadas por
las arcaicas, las bacterias y los organismos planctónicos para controlar su
flotabilidad vertical, de forma tal que pueden regular su profundidad de
acuerdo con señales externas como la luz solar. Las vesículas extracelulares en
los animales pueden almacenarse en la matriz extracelular. Algunas de ellas se
especializan en la biomineralización de algunos tipos de matriz extracelular
como el hueso, el cartílago y la dentina (Fevrier & Raposo, 2004; Grenier
& Mayrand, 1987; Raposo & Stoorvogel, 2013).
13. Organelos energéticos y endosimbióticos
|| Regresar al índice ||
Denominamos así a la mitocondria y al
cloroplasto, debido a que están relacionados con los metabolismos:
👉
respiración celular aeróbica con la mitocondria
👉 fotosíntesis
con el cloroplasto.
Adicionalmente ambos organelos parecen
haberse originado de bacterias parasitas o simbióticas que se establecieron
permanentemente en algunas células eucariotas, concepto conocido como la teoría
endosimbiótica.
13.1 La mitocondria
La mitocondria es un orgánulo de doble
membrana que se encuentra en la mayoría de los organismos eucariotas. Sin
embargo, algunas células en algunos organismos multicelulares pueden carecer de
ellas (por ejemplo, glóbulos rojos de mamíferos maduros). Varios organismos
unicelulares, como microsporidios, parabasalidos y diplomónidos, también han
reducido o transformado sus mitocondrias en otras estructuras. Hasta la fecha,
se sabe que solo dos eucariotas, Monocercomonoides y Henneguya salminicola, han perdido por
completo sus mitocondrias. Las mitocondrias generan la mayor parte del
suministro celular de trifosfato de adenosina (ATP), utilizado como fuente de
energía química. Una mitocondria se denomina así la planta de energía de la
célula eucariota.
Las mitocondrias son comúnmente entre 0,75
y 3 μm² en área, pero varían considerablemente en tamaño y estructura. A menos
que estén tinturadas específicamente, no son visibles. Además de suministrar
energía celular, las mitocondrias están involucradas en otras tareas, como la
señalización, la diferenciación celular y la muerte celular, así como también
para mantener el control del ciclo celular y el crecimiento celular. La
biogénesis mitocondrial a su vez se coordina temporalmente con estos procesos
celulares. Las mitocondrias se han implicado en varias enfermedades humanas,
incluyendo trastornos mitocondriales, disfunción cardíaca, insuficiencia
cardíaca y autismo.
El número de mitocondrias en una célula
puede variar ampliamente según el organismo, el tejido y el tipo de célula. Por
ejemplo, los glóbulos rojos no tienen mitocondrias, mientras que las células
del hígado pueden tener más de 2000. El orgánulo se compone de compartimentos
que realizan funciones especializadas. Estos compartimentos o regiones incluyen
la membrana externa, el espacio intermembrana, la membrana interna, las crestas
y la matriz.
Aunque la mayor parte del ADN de una
célula está contenida en el núcleo celular, la mitocondria tiene su propio
genoma independiente que muestra una similitud sustancial con los genomas
bacterianos. Las proteínas mitocondriales (proteínas transcritas a partir del
ADN mitocondrial) varían según el tejido y la especie. En humanos, se
identificaron 615 tipos distintos de proteínas de las mitocondrias cardíacas,
mientras que, en las ratas, se informaron 940 proteínas. Se cree que el
proteoma mitocondrial está regulado dinámicamente.
Una mitocondria tiene una membrana externa
y una membrana interna separadas por una región llamada espacio intermembrana (Figura 13.1). La
membrana interna está altamente invaginada (plegada) para formar proyecciones
llamadas crestas. Las crestas aumentan considerablemente el área de superficie
de la membrana interna, que es el sitio donde se produce el ATP. El
compartimento encerrado por la membrana interna es la matriz mitocondrial.
El papel principal de las mitocondrias es producir ATP. Aunque las mitocondrias producen la mayor parte del ATP de una célula, las mitocondrias no crean energía. Más bien, su función principal es convertir la energía química que se almacena dentro de los enlaces covalentes de las moléculas orgánicas (grasa especialmente, pero también azúcares y proteínas en emergencias) en una forma que las células puedan usar fácilmente. Los enlaces covalentes en azúcares, grasas y aminoácidos almacenan una gran cantidad de energía. La descomposición de estas moléculas en moléculas más simples libera energía que se utiliza para producir ATP. Muchas proteínas en las células vivas usan el ATP como fuente de energía para llevar a cabo sus funciones, como la contracción muscular, la absorción de nutrientes, la división celular y muchos otros procesos celulares.

Figura 13.1. Estructura de una mitocondria.
Esta figura enfatiza la organización de la membrana de una mitocondria, que
tiene una membrana externa e interna. Las invaginaciones de la membrana interna
se llaman crestas. La matriz mitocondrial se encuentra dentro de la membrana
interna. La micrografía es un TEM coloreado.
Las mitocondrias también realizan otras
funciones. Están involucrados en la síntesis, modificación y descomposición de
varios tipos de moléculas celulares. Por ejemplo, la síntesis de ciertas
hormonas requiere enzimas que se encuentran en las mitocondrias. Otro papel
interesante de las mitocondrias es generar calor en células grasas
especializadas conocidas como células grasas marrones. Los grupos de células de
grasa marrón sirven como "almohadillas térmicas" que ayudan a revivir
a los animales en hibernación y a proteger las áreas sensibles de animales
jóvenes del frío.
13.2 El cloroplasto
Los cloroplastos son orgánulos que capturan la energía de la luz y utilizan parte de esa energía para sintetizar moléculas orgánicas como la glucosa. Este proceso, es llamado fotosíntesis. Los cloroplastos se encuentran en casi todas las especies de plantas y algas. La Figura 13.2 muestra la estructura de un cloroplasto típico. Al igual que una mitocondria, un cloroplasto contiene una membrana externa e interna. Un espacio intermembrana se encuentra entre estas dos membranas. Un tercer sistema de membranas, la membrana tilacoidea, forma muchos túbulos aplanados y llenos de líquido que encierran un compartimento único y enrevenado llamado luz tilacoidea. Estos túbulos tienden a apilarse uno encima del otro para formar una estructura llamada granum (plural, grana). El estroma es el compartimento del cloroplasto que está encerrado por la membrana interna pero fuera de la membrana tilacoidea.

Figura 13.2. Estructura de un cloroplasto.
Como una mitocondria, un cloroplasto está encerrado en una membrana doble.
Además, tiene un sistema interno de membrana tilacoide que forma compartimentos
aplanados, que puede considerarse una tercera membrana. Estos compartimientos
se apilan unos sobre otros para formar grana. El estroma se encuentra dentro de
la membrana interna pero fuera de la membrana tilacoidea. Esta micrografía es
un TEM coloreado.
Los cloroplastos son una versión
especializada de orgánulos vegetales que se conocen más generalmente como
plastidios. Todos los plastidios se derivan de proplastidos no especializados.
Los diversos tipos de plastidios se distinguen por sus habilidades sintéticas y
los tipos de pigmentos que contienen. Los cloroplastos, que realizan la
fotosíntesis, contienen el pigmento verde clorofila. La abundante cantidad de
cloroplastos en las hojas de las plantas les da su color verde. Los
cromoplastos, un segundo tipo de plastidio, funcionan para sintetizar y
almacenar los pigmentos amarillos, naranjas y rojos conocidos como
carotenoides. Los cromoplastos dan a muchas frutas y flores sus colores. En otoño,
los cromoplastos también dan a las hojas sus colores amarillo, naranja y rojo.
Un tercer tipo de plastidio, los leucoplastos, generalmente carece de moléculas
de pigmento. Un amiloplastos es un leucoplasto que sintetiza y almacena
almidón. Los amiloplastos son comunes en estructuras subterráneas como raíces y
tubérculos.
13.3 Independencia con respecto
a la célula eucariota
Para apreciar completamente la estructura y organización de las mitocondrias y los cloroplastos, también debemos examinar brevemente sus propiedades genéticas. En 1951, Yasutane Chiba expuso las células vegetales a la mancha de Feulgeno, un colorante específico de ADN, y descubrió que los cloroplastos se tiñeron. Según esta observación, fue el primero en sugerir que los cloroplastos contienen su propio ADN. Los investigadores en las décadas de 1970 y 1980 aislaron el ADN de los cloroplastos y las mitocondrias. Estos estudios revelaron que el ADN de estos orgánulos se parecía a versiones más pequeñas de cromosomas bacterianos.

Figura 13.3. Las mitocondrias se multiplican
por fisión binaria.
Los cromosomas que se encuentran en las
mitocondrias y los cloroplastos se denominan genoma mitocondrial y genoma de
cloroplastos, respectivamente, mientras que los cromosomas que se encuentran en
el núcleo de la célula constituyen el genoma nuclear. Al igual que las
bacterias, los genomas de la mayoría de las mitocondrias y los cloroplastos
están compuestos por un solo cromosoma
circular. En comparación con el genoma nuclear, son muy pequeños.
Por ejemplo, la cantidad de ADN en el genoma nuclear humano (aproximadamente 3
mil millones de pares de bases) es aproximadamente 200,000 veces mayor que el
genoma mitocondrial. En términos de genes, el genoma humano tiene
aproximadamente 22,000 genes diferentes, mientras que el genoma mitocondrial
humano tiene solo unas pocas docenas. Los genomas de cloroplastos tienden a ser
más grandes que los genomas mitocondriales, y tienen un número
correspondientemente mayor de genes. Dependiendo de la especie particular de
planta o alga, un genoma de cloroplasto es aproximadamente 10 veces más grande
que el genoma mitocondrial de las células humanas.
Así como los genomas de las mitocondrias y
los cloroplastos se parecen a los genomas bacterianos, la producción de nuevas
mitocondrias y cloroplastos se asemeja mucho a la división de las células
bacterianas. Al igual que sus contrapartes bacterianas, las mitocondrias y los
cloroplastos aumentan en número por fisión binaria, o se dividen en dos. La Figura 13.3
ilustra el proceso para una mitocondria. El cromosoma mitocondrial, que se
encuentra en una región llamada nucleoide, se duplica y el orgánulo se divide
en dos orgánulos separados. Las divisiones mitocondriales y de cloroplastos son
necesarias para mantener un juego completo de estos orgánulos cuando se produce
el crecimiento celular después de la división celular.
Además, las condiciones ambientales pueden
influir en los tamaños y números de estos orgánulos. Por ejemplo, cuando las
plantas están expuestas a más luz solar, aumenta el número de cloroplastos en
las células de las hojas.
13.4 La teoría endosimbiótica
La observación de que las mitocondrias y
los cloroplastos contienen su propio material genético puede parecer
desconcertante. Quizás se pueda pensar que sería más simple para una célula
eucariota tener todo su material genético en un solo lugar: el núcleo. Los
distintos genomas de las mitocondrias y los cloroplastos se remontan a su
origen evolutivo, que implica una antigua asociación simbiótica.
Una relación simbiótica ocurre cuando dos
especies diferentes viven en contacto directo entre sí. La endosimbiosis
describe una relación simbiótica en la que las especies más pequeñas, el
simbionte, en realidad viven dentro de las especies más grandes. En 1883,
Andreas Schimper propuso que los cloroplastos descendieran de una relación
endosimbiótica mutualista entre las cianobacterias (una bacteria capaz de
fotosíntesis) y las células eucariotas. En 1922, Ivan Wallin también planteó la
hipótesis de un origen endosimbiótico para las mitocondrias.
A pesar de estas ideas interesantes, la cuestión de la endosimbiosis se ignoró en gran medida hasta el descubrimiento de que las mitocondrias y los cloroplastos contienen su propio material genético. En 1970, Lynn Margulis revivió el tema de la endosimbiosis como el origen de las mitocondrias y los cloroplastos en su libro Origin of Eukaryotic Cells. Durante las décadas de 1970 y 1980, el advenimiento de las técnicas de genética molecular permitió a los investigadores analizar genes de mitocondrias, cloroplastos, bacterias y genomas nucleares eucariotas. Los investigadores descubrieron que los genes en las mitocondrias y los cloroplastos son muy similares a los genes bacterianos. Del mismo modo, las mitocondrias y los cloroplastos son sorprendentemente similares en tamaño y forma a ciertas especies bacterianas. Estas observaciones proporcionaron un fuerte apoyo a la teoría de la endosimbiosis, que propone que las mitocondrias y los cloroplastos se originaron a partir de bacterias que se establecieron dentro de una célula eucariota primordial.

Figura 13.4. Una visión
simplificada de la teoría de la endosimbiosis. (a) De acuerdo con este
concepto, las mitocondrias modernas se derivaron de bacterias moradas, también
llamadas proteobacterias α. En el transcurso de la evolución, sus
características cambiaron a las que se encuentran en las mitocondrias de hoy.
(b) Se produjo un fenómeno similar para los cloroplastos, que se derivaron de
las cianobacterias (bacterias azul-verdosas), una bacteria que es capaz de
fotosíntesis.
Durante los próximos 2 mil millones de
años, las características de estas células bacterianas intracelulares cambiaron
gradualmente a las de una mitocondria o cloroplasto.
La simbiosis ocurre porque la relación es
beneficiosa para una o ambas especies. Según la teoría de la endosimbiosis, esta
relación proporcionó a las células eucariotas características celulares útiles.
Los cloroplastos, que se derivaron de las cianobacterias, tienen la capacidad
de llevar a cabo la fotosíntesis. Esto beneficia a las células vegetales al
darles la capacidad de utilizar la energía de la luz solar. En comparación, se
cree que las mitocondrias se derivaron de un tipo diferente de bacteria
conocida como bacteria púrpura, o proteobacterias α. En este caso, la relación
endosimbiótica permitió a las células eucariotas sintetizar mayores cantidades
de ATP. La forma en que la relación hubiera sido beneficiosa para una
cianobacteria o una bacteria púrpura es menos clara, aunque el citosol de una
célula eucariota puede haber proporcionado un entorno estable con un suministro
adecuado de nutrientes.
Durante la evolución de las especies
eucariotas, muchos genes que se encontraron originalmente en los genomas de las
bacterias y cianobacterias púrpuras primordiales se transfirieron de los
orgánulos al núcleo. Esto ha ocurrido muchas veces a lo largo de la evolución,
por lo que las mitocondrias y los cloroplastos modernos han perdido la mayoría
de los genes que aún existen en las bacterias púrpuras y las cianobacterias
actuales. Algunos investigadores especulan que el movimiento de los genes hacia
el núcleo facilita que la célula controle la estructura, función y división de
las mitocondrias y los cloroplastos. En las células modernas, cientos de
proteínas diferentes que forman estos orgánulos están codificadas por genes que
han sido transferidos al núcleo. Estas proteínas se elaboran en el citosol y
luego se incorporan a las mitocondrias o cloroplastos.
14. Otros organelos eucariotas
|| Regresar al índice ||
Las células
eucariotas también poseen organelos que describimos en la sección de los
procariotas, tales como: la membrana celular, la pared celular y los ribosomas.
aunque sus funciones y estructuras generales son esencialmente las mismas, a
continuación plantearemos algunas diferencias.
14.1 Membrana celular eucariota
Las membranas celulares protegen y
organizan las células. Todas las células tienen una membrana plasmática externa
que regula no solo lo que ingresa a la célula, sino también la cantidad de
cualquier sustancia que ingresa. Esencialmente la misma función que cumplen en los procariotas.
Sin embargo,
una de las diferencias notables radica en el hecho de que la membrana celular
eucariota no está tan encajonada en una pared celular. aunque existen linajes
que poseen paredes celulares como hongos y plantas, la célula eucariota
prototípica no está encajonada y por lo tanto puede cambiar su forma. en
esencia podemos decir que la membrana celular de una célula eucariota es mucho
más flexible, pues puede soportar una célula de mayor tamaño y con formas
variables. Por poner un ejemplo de este prototipo de célula eucariota podemos
citar a la humilde ameba o los fagocitos que moran en nuestros cuerpos y nos
defienden contra enfermedades (De Duve &
Pizano, 1995).

Figura 14.1. Las células eucariotas tienen formas muy variadas
debido a la flexibilidad de su membrana celular y también a la gran
especialización de sus citoesqueleto, de esta forma podemos tener células con
formas variantes como la ameba, o células con formas tremendamente
especializadas como la neurona.
La composición lipídica de las membranas
procarióticas y eucarióticas difiere considerablemente. Las membranas
procarióticas tienen lípidos cargados negativamente, a saber,
fosfatidilglicerol, cardiolipina y fosfatidilserina, mientras que las membranas
eucarióticas son ricas en esfingomielina neutra y fosfatidilcolina. Esta parte
estructural diferente imparte funciones diferentes a estas membranas. Las
células eucariotas tienen diferentes compartimentos subcelulares para llevar a
cabo diversas actividades que están confinadas a las membranas celulares de los
procariotas, ya que carecen de compartimentos unidos a la membrana (Yeagle, 2016).
14.2 Paredes celulares de los eucariotas
Las paredes celulares evolucionaron de
forma independiente en muchos grupos, sin embargo los dos grupos de eucariotas más importantes y más famosos
por poseer paredes celulares son sin duda las plantas y los hongos.
Plantas y parientes: Los eucariotas fotosintéticos (los llamados plantas y algas) son
un grupo con paredes celulares de celulosa, donde la pared celular está
estrechamente relacionada con la evolución de la multicelularidad, la
terrestrialización y la vascularización. La celulosa sintasa CesA evolucionó en
cianobacterias y formó parte de Archaeplastida (Reino vegetal) desde la endosimbiosis; los eventos de endosimbiosis secundarios
lo transfirieron (con las proteínas arabinogalactanas) a las algas pardas y los
oomicetos. Posteriormente, las plantas desarrollaron varios genes a partir de
CesA, incluida la familia de proteínas Csl (similar a la celulosa sintasa) y
proteínas Ces adicionales. En combinación con las diversas glicosiltransferasas
(GT), permiten construir estructuras químicas más complejas (Popper et al., 2011).
Las paredes
celulares son significativamente más gruesas que las membranas plasmáticas y
fueron visibles incluso para los primeros microscopistas, incluido Robert
Hooke, quien originalmente identificó las estructuras en una muestra de corcho
y luego acuñó el término células en la década de 1660. El grosor, así como la
composición y organización de las paredes celulares pueden variar
significativamente. Muchas células vegetales tienen tanto una pared celular
primaria, que acomoda a la célula a medida que crece, como una pared celular
secundaria que desarrollan dentro de la pared primaria después de que la célula
deja de crecer. La pared celular primaria es más delgada y flexible que la
pared celular secundaria y, a veces, se mantiene en un estado sin cambios o
ligeramente modificado sin la adición de la pared secundaria, incluso después
de que finaliza el proceso de crecimiento.

Figura 14.2. Las células vegetales poseen una pared celular que se
encuentra de forma externa a sus membranas y contenida en una lamela intermedia
que permite la interacción con otras células, el grosor de la pared es muy
variable pero es una de las estructuras más gruesas de una célula vegetal y se
puede ver con un microscopio óptico simple y primitivo. Sin duda el material
más importante de la para el celular de las plantas es la celulosa.
Los principales componentes químicos de la
pared celular primaria de la planta incluyen la celulosa (en forma de
microfibrillas organizadas; consulte la Figura 1), un carbohidrato complejo
formado por varios miles de moléculas de glucosa unidas de extremo a extremo.
Además, la pared celular contiene dos grupos de polisacáridos ramificados, las
pectinas y los glucanos reticulados. Organizados en una red con las
microfibrillas de celulosa, los glucanos reticulados aumentan la resistencia a
la tracción de la celulosa, mientras que la red coextensiva de pectinas
proporciona a la pared celular la capacidad de resistir la compresión. Además
de estas redes, se puede encontrar una pequeña cantidad de proteína en todas las
paredes celulares primarias de las plantas. Se cree que parte de esta proteína
aumenta la resistencia mecánica y parte consiste en enzimas, que inician
reacciones que forman, remodelan o rompen las redes estructurales de la pared.
Tales cambios en la pared celular dirigidos por enzimas son particularmente
importantes para que la fruta madure y las hojas caigan en otoño.
La pared celular vegetal secundaria, que a
menudo se deposita dentro de la pared celular primaria a medida que la célula
madura, a veces tiene una composición casi idéntica a la de la pared
desarrollada anteriormente. Sin embargo, más comúnmente, se encuentran
sustancias adicionales, especialmente lignina, en la pared secundaria. Lignina
es el nombre general de un grupo de polímeros de alcoholes aromáticos que son
duros y confieren una resistencia considerable a la estructura de la pared
secundaria. La lignina es lo que proporciona las características favorables de
la madera a las células de fibra de los tejidos leñosos y también es común en las
paredes secundarias de los vasos del xilema, que son fundamentales para brindar
soporte estructural a las plantas. La lignina también hace que las paredes
celulares de las plantas sean menos vulnerables al ataque de hongos o
bacterias, al igual que la cutina, la suberina y otros materiales cerosos que a
veces se encuentran en las paredes celulares de las plantas.
Una región especializada asociada con las
paredes celulares de las plantas, y que a veces se considera un componente
adicional de ellas, es la lámina media. Rica en pectinas, la lámina media es
compartida por las células vecinas y las une firmemente. Posicionadas de esta
manera, las células pueden comunicarse entre sí y compartir su contenido a
través de conductos especiales. Denominados plasmodesmos, estos pequeños
pasajes penetran en la lámina media, así como en las paredes celulares
primarias y secundarias, proporcionando vías para el transporte de moléculas
citoplasmáticas de una célula a otra.
Hongos verdaderos: Los hongos usan una pared celular de quitina-glucano-proteína . Comparten
la ruta de síntesis de 1,3-β-glucano con las plantas, utilizando
1,3-beta-glucano sintasas homólogas de la familia GT48 para realizar la tarea,
lo que sugiere que dicha enzima es muy antigua dentro de los eucariotas. Sus
glicoproteínas son ricas en manosa. La pared celular podría haber evolucionado
para impedir las infecciones virales. Las proteínas incrustadas en las paredes
celulares son variables, contenidas en repeticiones en tándem sujetas a
recombinación homóloga. Un escenario alternativo es que los hongos comenzaron
con una pared celular basada en quitina y luego adquirieron las enzimas GT-48
para los 1,3-β-glucanos a través de la transferencia horizontal de genes. La
vía que conduce a la síntesis de 1,6-β-glucano no se conoce lo suficiente en
ninguno de los dos casos. (Farkaš, 2020; Lima, Colombo, &
de Almeida Junior, 2019)

Figura 14.3. La pared celular de los hongos es variable dependiendo
de la especie su grosor y composición poseen variaciones en cuanto a los tipos
de azúcares y su grosor sin embargo la quitina es sin duda el material más
importante.
podemos
dividir la pared celular de los hongos en 2 regiones importantes la región de
mano proteínas y la región quitinosa. La región de manoproteínas es más externa
de la pared celular de los hongos y es la que se encarga de interactuar con el
medio así como con parásitos que los afectan como los virus. La región
quitinosa es más interna y se encarga de interactuar con la membrana celular
verdadera y la región de mano proteínas, esta región intermedia es la que
contiene la quitina y otras sustancias de tipo polisacárido (Garcia-Rubio,
de Oliveira, Rivera, & Trevijano-Contador, 2020).
14.3 Ribosoma eucariota
Los ribosomas son una máquina molecular
grande y compleja que cataliza la síntesis de proteínas, denominada traducción.
El ribosoma selecciona los ARN de transferencia aminoacilados (ARNt) en función
de la secuencia de un ARN mensajero (ARNm) que codifica una proteína y une
covalentemente los aminoácidos en una cadena polipeptídica. Los ribosomas de
todos los organismos comparten un centro catalítico altamente conservado. Sin
embargo, los ribosomas de los eucariotas (animales, plantas, hongos y un gran
número de organismos unicelulares, todos con núcleo) son mucho más grandes que
los ribosomas procariotas (bacterias y arqueas) y están sujetos a vías de
regulación y biogénesis más complejas. Los ribosomas eucariotas también se
conocen como ribosomas 80S, en referencia a sus coeficientes de sedimentación
en unidades Svedberg, porque sedimentan más rápido que los ribosomas
procariotas (70S). Los ribosomas eucariotas tienen dos subunidades desiguales,
denominadas subunidad pequeña (40S) y subunidad grande (60S) según sus
coeficientes de sedimentación. Ambas subunidades contienen docenas de proteínas
ribosómicas dispuestas en un andamio compuesto por ARN ribosómico (ARNr). La
subunidad pequeña controla la complementariedad entre el anticodón de ARNt y el
ARNm, mientras que la subunidad grande cataliza la formación de enlaces
peptídicos (Baßler & Hurt, 2019; Pelletier
& Sonenberg, 2019).
Los ribosomas eucariotas se producen y
ensamblan en el nucléolo. Las proteínas ribosómicas ingresan al nucléolo y se
combinan con las cuatro cadenas de ARNr para crear las dos subunidades
ribosómicas (una pequeña y una grande) que formarán el ribosoma completo. Las
unidades de ribosomas salen del núcleo a través de los poros nucleares y se
unen una vez en el citoplasma con el fin de sintetizar proteínas.
14.4 Flagelo eucariota
Con el objetivo de enfatizar la distinción
entre los flagelos bacterianos y los cilios y flagelos eucariotas, algunos
autores intentaron reemplazar el nombre de estas dos estructuras eucariotas con
"undulipodos" (Lindemann & Lesich, 2021;
Simpson, 2019) o
"cilios”. Sin embargo, el uso discriminatorio de los términos
"cilios" y "flagelos" para eucariotas adoptados en este
artículo sigue siendo común.
El núcleo de un flagelo eucariótico,
conocido como axonema, es un haz de nueve pares fusionados de microtúbulos
conocidos como dobletes que rodean dos microtúbulos centrales únicos
(singletes). Este axonema 9+2 es característico del flagelo eucariota. En la
base de un flagelo eucariótico hay un cuerpo basal, "blefaroplasto" o
cinetosoma, que es el centro organizador de microtúbulos para los microtúbulos
flagelares y tiene una longitud de unos 500 nanómetros. Los cuerpos basales son
estructuralmente idénticos a los centríolos. El flagelo está encerrado dentro
de la membrana plasmática de la célula, de modo que el interior del flagelo es
accesible al citoplasma de la célula.

Figura 14.4. Estructura del flagelo eucariota
Además del axonema y el cuerpo basal, de
morfología relativamente constante, otras estructuras internas del aparato
flagelar son la zona de transición (donde se encuentran el axonema y el cuerpo
basal) y el sistema radicular (estructuras microtubulares o fibrilares que se
extienden desde los cuerpos basales hacia el citoplasma). ), más variables y útiles
como indicadores de relaciones filogenéticas de eucariotas.
Cada uno de los 9 microtúbulos dobles
exteriores extiende un par de brazos de dineína (un brazo "interior"
y otro "exterior") al microtúbulo adyacente; estos producen fuerza a
través de la hidrólisis de ATP. El axonema flagelar también contiene radios
radiales, complejos de polipéptidos que se extienden desde cada uno de los
nueve dobletes de microtúbulos externos hacia el par central, con la "cabeza"
del radio hacia adentro. Se cree que el radio radial está involucrado en la
regulación del movimiento flagelar, aunque aún no se conocen su función exacta
y su método de acción.
Los patrones de latidos regulares de los
cilios y flagelos eucariotas generan movimiento a nivel celular. Los ejemplos
van desde la propulsión de células individuales, como la natación de los
espermatozoides, hasta el transporte de fluidos a lo largo de una capa
estacionaria de células, como en el tracto respiratorio.

Figura 14.5. Patrón de movimiento de un flagelo y un cilio eucarióticos, una distinción tradicional antes de que se
conozcan las estructuras de los dos.
Aunque los cilios y flagelos eucarióticos
son en última instancia lo mismo, a veces se clasifican por su patrón de
movimiento, una tradición anterior a que se conocieran sus estructuras. En el
caso de los flagelos, el movimiento suele ser plano y ondulatorio, mientras que
los cilios móviles suelen realizar un movimiento tridimensional más complicado
con un golpe de potencia y recuperación. Otra forma tradicional de distinción
es por el número de orgánulos 9+2 en la célula.
Referencias
|| Regresar al índice ||
Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K.,
& Walter, P. (2015). Molecular biology of the cell (5th ed.).
Garland Science.
Bardy, S.
L., Ng, S. Y. M., & Jarrell, K. F. (2004). Recent advances in the structure
and assembly of the archaeal flagellum. Journal of Molecular Microbiology
and Biotechnology, 7(1–2), 41–51.
Baßler, J.,
& Hurt, E. (2019). Eukaryotic ribosome assembly. Annual Review of
Biochemistry, 88, 281–306.
Belk, C.,
& Maier, V. B. (2013). Biology Science for Life with physiology.
(Pearson, Ed.) (4th ed.).
Berg, J.
M., Tymoczko, J. L., Gatto, G. J., & Stryer, L. (2015). Biochemistry
(8th ed.). Freeman.
Brooker, R.
J., Widmaier, E. P., Graham, L. E., & Stiling, P. D. (2017). Biology
(4th ed.). McGraw-Hill Education.
Campbell,
M. K., & Farrell, S. O. (2009). Biochemistry (6th ed.). USA: Thomsom
Brooks/Cole.
Cox, M. M.,
Doudna, J. A., & O’Donnell, M. (2012). Molecular biology (1st ed.).
Freeman.
De Duve,
C., & Pizano, M. (1995). Polvo vital: origen y evolución de la vida en
la tierra. Norma, Bogotá.
Farkaš, V.
(2020). The fungal cell wall. Fungal Protoplasts, 3–29.
Fevrier,
B., & Raposo, G. (2004). Exosomes: endosomal-derived vesicles shipping
extracellular messages. Current Opinion in Cell Biology, 16(4),
415–421.
Garcia-Rubio,
R., de Oliveira, H. C., Rivera, J., & Trevijano-Contador, N. (2020). The
fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Frontiers
in Microbiology, 10, 2993.
Grenier,
D., & Mayrand, D. (1987). Functional characterization of extracellular
vesicles produced by Bacteroides gingivalis. Infection and Immunity, 55(1),
111–117.
Hoefnagels,
M. (2015). Biology: concepts and investigations (3rd ed.). McGraw-Hill
New York.
Islas-Morales,
P. F., Cárdenas, A., Mosqueira, M. J., Jiménez-García, L. F., & Voolstra,
C. R. (2023). Ultrastructural and proteomic evidence for the presence of a
putative nucleolus in an Archaeon.
Kåhrström,
C. T. (2013). Biofilms: Flagella function as mechanosensors. Nature Reviews
Microbiology, 11(9), 597.
Kalanetra,
K. M., Huston, S. L., & Nelson, D. C. (2004). Novel, attached,
sulfur-oxidizing bacteria at shallow hydrothermal vents possess vacuoles not
involved in respiratory nitrate accumulation. Applied and Environmental
Microbiology, 70(12), 7487–7496.
Karp, G. C.
(2013). Cell and Molecular Biology, Concepts and Experiments (7th ed.).
USA: Wiley Online Library.
König, H.
(1988). Archaeobacterial cell envelopes. Canadian Journal of Microbiology,
34(4), 395–406.
Lima, S.
L., Colombo, A. L., & de Almeida Junior, J. N. (2019). Fungal cell wall:
emerging antifungals and drug resistance. Frontiers in Microbiology, 10,
2573.
Lindemann,
C. B., & Lesich, K. A. (2021). The many modes of flagellar and ciliary
beating: insights from a physical analysis. Cytoskeleton, 78(2),
36–51.
Mackean, D.
G., & Hayward, D. (2014). Biology (3rd ed.). IGCSE Cambridge.
Mader, S.
S. (2010). Biology (10th ed.). McGraw-Hill Education.
Mader, S.
S., & Windelspecht, M. (2015). Biology (12th ed.). McGraw-Hill
Education.
Mader, S.
S., & Windelspecht, M. (2018). Essentials of biology (5th ed.).
McGraw-Hill Education.
Mao, Y. S.,
Zhang, B., & Spector, D. L. (2011). Biogenesis and function of nuclear
bodies. Trends in Genetics, 27(8), 295–306.
Mashburn‐Warren,
L. M., & Whiteley, M. (2006). Special delivery: vesicle trafficking in
prokaryotes. Molecular Microbiology, 61(4), 839–846.
Mason, K.
A., Losos, J. B., Singer, S. R., & Raven, P. H. (2014). Biology (7th
ed.). McGraw-Hill New York.
Mounkes, L.
C., & Stewart, C. L. (2004). Aging and nuclear organization: lamins and
progeria. Current Opinion in Cell Biology, 16(3), 322–327.
Murray, R.
K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V., & Weil, A.
(2012). Harpers Illustrated Biochemistry (29th ed.). McGraw-Hill
Medical.
Nunes-Alves,
C. (2015). Bacterial evolution: Resurrecting motility. Nature Reviews
Microbiology, 13(4), 187.
Pelletier,
J., & Sonenberg, N. (2019). The organizing principles of eukaryotic
ribosome recruitment. Annual Review of Biochemistry, 88, 307–335.
Pollard, T.
D., Earnshaw, W. C., Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell
Biology (3rd ed.). Elsevier.
Popper, Z.
A., Michel, G., Hervé, C., Domozych, D. S., Willats, W. G. T., Tuohy, M. G., …
Stengel, D. B. (2011). Evolution and diversity of plant cell walls: from algae
to flowering plants. Annual Review of Plant Biology, 62, 567–590.
Raposo, G.,
& Stoorvogel, W. (2013). Extracellular vesicles: exosomes, microvesicles,
and friends. The Journal of Cell Biology, 200(4), 373–383.
Reece, J.
B., Urry, L. A., Wasserman, S. A., Cain, M. L., Minorsky, P. V, & Jackson,
R. B. (2014). Campbell Biology (10th ed.). Pearson Higher Ed.
Sadava, D.,
Berenbaum, M., & Hillis, D. (2014). Life the Science of Biology
(10th ed.). Sinauer & MacMillian.
Simon, E.
J., Dickey, J. L., & Reece, J. B. (2013). Essential biology
(Pearson).
Simpson, G.
G. (2019). Phylogeny: Eukaryotes to Chordates. Evolution’s Clinical
Guidebook: Translating Ancient Genes into Precision Medicine, 173.
Solomon,
E., Martin, C., Martin, D. W., & Berg, L. R. (2014). Biology (10th
ed.). Cengage Learning.
Starr, C.,
Evers, C., & Starr, L. (2013). Biology: Today and Tomorrow With
Physiology (4th ed.). Brooks/Cole.
Taylor, M.
R., Simon, E. J., Dickey, J. L., Hogan, K., & Reece, J. B. (2018). Biology,
Concepts and conections (9th ed.). Boston: Pearson education.
Thomas, N.
A., Bardy, S. L., & Jarrell, K. F. (2001). The archaeal flagellum: a
different kind of prokaryotic motility structure. FEMS Microbiology Reviews,
25(2), 147–174.
Urry, L.
A., Cain, M. L., Minorsky, P. V, Wasserman, S. A., & Reece, J. B. (2017). Campbell
Biology (11th ed.). Pearson.
Weaver, R.
F. (2012). Molecular biology (5th ed.). McGraw-Hill Education.
Yeagle, P.
L. (2016). The membranes of cells. Academic Press.
Zhang, Z.,
& Zhang, R. (2015). Epigenetics in autoimmune diseases: pathogenesis and
prospects for therapy. Autoimmunity Reviews, 14(10), 854–863.
No hay comentarios:
Publicar un comentario