
Índice
Portada

1. Introducción
|| Regresar al Índice ||
El sistema circulatorio es el
sistema de órganos encargado del transporte y distribución de sustancias al
interior de un ser vivo, y como en muchos otros sistemas de órganos, se lo
puede analizar al nivel subcelular, celular, de tejido, órgano y sistema. Los
sistemas circulatorios adicionalmente, se encuentran vinculados a lo que
Christian de Duve menciona como el milieu
interieur (De Duve & Pizano, 1995), o mejor dicho, un fluido interno en el cual las sustancias se
disuelven para su transporte desde un lugar del ser vivo hasta otro. Dado lo
anterior, tenemos la generación de dos mecanismos de transporte, uno aleatorio
y otros no aleatorios. Mucho de lo que estudiamos en los sistemas circulatorios
son adaptaciones de los diferentes seres vivos para que el transporte de
sustancias se realice de manera no aleatoria.
Una de las mayores tendencias de los seres
vivos ha sido la de especializar partes de sus cuerpos para el cumplimiento de
funciones concretas, pero con el inconveniente de hacerse dependientes de otros
sistemas, es necesario que estos componentes especializados se conecten de
algún modo para compartir sustancias. Puede que una parte del cuerpo se
especialice, por ejemplo, en la reproducción, pero para reproducirse es necesario
el aporte de materiales desde los sistemas nutritivos, y la eliminación de
desechos hacia los sistemas de excreción. De cierta manera muchos sistemas de
órganos se tienden a ver como compartimentos estancos aislados del resto, sin
embargo, el sistema circulatorio está allí para recordarnos que todo está
unido, y por tal razón, este sirve como una suerte de resumen de las funciones
de los demás sistemas de órganos. Los sistemas circulatorios cumplen la función
de integrar a los diferentes tejidos, sirviendo como mecanismos de transporte,
o como autopistas, que permiten el funcionamiento simultáneo e integral. Las
sustancias que un sistema circulatorio debe transportar son las siguientes:
👉 Gases:
haciendo parte del sistema respiratorio o de intercambio de gases por medio de
los capilares pulmonares donde se intercambian los gases de importancia
gaseosa.
👉 Nutrientes solubles:
haciendo parte del sistema digestivo por medio de los capilares que rodean a
los intestinos donde se absorben los nutrientes.
👉 Desechos:
haciendo parte del sistema excretor por medio de capilares de la piel y los
capilares especializados de los riñones que rodean al asa de Henle.
👉 Mensajes:
por medio de los capilares que conectan al tejido emisor y receptor de un
mensajero químico.
👉 Células del sistema
inmune: como componentes que fluyen a través del fluido
hemolimfático general o si se da especialización, a través de la linfa y la
sangre.
👉 Células propias del
sistema circulatorio: Especializadas en el transporte de gases,
nutrientes, desechos y mensajeros químicos.
👉 Proteínas solubles:
ya sean estas componentes del sistema inmune “anticuerpos”, del sistema
circulatorio “fibrina” o de transporte de grasas “quilomicrones, lipoproteínas
de alta y de baja densidad”.
Es evidente que al analizar por separado
cada sistema de órganos, hemos tenido que ver secciones del sistema
circulatorio, por ejemplo, el sistema de venas portal que conecta al intestino
con el hígado durante la digestión, los capilares alveolares en la respiración
o el asa de Henle en la excreción. Sin embargo, el objeto del presente capítulo
del sistema circulatorio es la de ser más generales y abarcar incluso al nivel
sub-celular, después de todo una célula también puede llegar a tener
componentes especializados y debe distribuir materiales y desechos.
1.1 William Harvey
William Harvey era un
médico inglés. Nació en Folkestone, Kent, Inglaterra,
el 1 de abril de 1573. Fue el primero en explicar cómo el corazón movía la
sangre a través del cuerpo. Murió el 3 de junio de 1657 en Roehampton. Un hospital
en Ashford, Kent lleva el nombre de Harvey. Fue al The
King's School, Canterbury, luego a Gonville
and Caius College, Cambridge. Luego fue a la
Universidad de Padua, donde se graduó en 1602. Cuando Harvey regresó a
Inglaterra se casó con Elizabeth Browne, la hija del médico real de Isabel I.
Se convirtió en médico en el Hospital St. Bartholomew de Londres desde 1609
hasta 1643.
William Harvey es famoso por su idea de
que el corazón bombea sangre alrededor
del cuerpo. Fue el primero en describir correcta y detalladamente la
circulación y sus propiedades. Algunos hombres sabían del circuito
circulatorio menor del corazón a los pulmones y de regreso, pero su
trabajo se perdió. Una obra de teología del siglo XVI, titulada Cristianismi restitutio, fue escrita por
Miguel Serveto. Después de haber sido quemado en la hoguera por herejía (por
Calvino), su libro fue destruido. Tres copias sobrevivieron. Un siglo y medio
después, mucho después de Harvey, alguien notó que había dicho algo sobre la
sangre que circulaba.

Harvey anunció sus descubrimientos sobre
el sistema circulatorio en 1616 durante una serie de conferencias (sus notas de
conferencias aún existen). Escribió un libro (Exercitatio Anatomica de Motu Cordis et sanguinis in Animalibus) al
respecto en 1628. Dijo que la sangre se
bombea alrededor del cuerpo en un sistema cerrado. El corazón
bombeaba sangre a través del cuerpo antes de regresar al corazón para repetir
el proceso. Esto iba en contra de lo que se creía en ese momento que era
cierto.
El médico romano, Galeno, creía que había
dos sistemas en el cuerpo que se ocupaban de la sangre. En ese momento se
pensaba que la sangre de color rojo oscuro en las venas provenía del hígado y
la sangre de color rojo brillante en las arterias provenía del corazón. Se
creía que el hígado y el corazón producían la sangre y la utilizaban las partes
del cuerpo a las que se bombeaba.
Las ideas de Harvey surgieron de la
disección de cuerpos humanos. Harvey descubrió que el hígado tendría que
producir 540 libras de sangre cada hora para que Galeno tuviera razón. Esto le
mostró que la sangre no se estaba agotando. Estaba siendo reutilizada por el
cuerpo. Dijo que la sangre fluía por el corazón en dos
circuitos, llamados mayor y menor. El circuito menor entre corazón y
pulmones obtiene oxígeno. El circuito mayor entre el corazón y el cuerpo oxigena
los tejidos. Harvey dijo que el corazón era solo una bomba que bombea sangre
por todo el cuerpo, por lo que no era el órgano encargado del raciocinio como
había opinado Aristóteles.
2. Generalidades
|| Regresar al Índice ||
Los sistemas circulatorios transportan un
fluido que a su vez transporta materiales o sustancias químicas, que pueden ser
clasificadas en nutrientes, desechos, mensajeros químicos y xenobióticos. Todos
pueden ser subclasificados en gases solubles o en sustancias no gaseosas
solubles.
2.1 Nutrientes
Son los materiales que necesita la célula
para funcionar y reproducirse en caso de que el tejido requiera crecer o
repararse. Estos materiales provienen en últimas del sistema digestivo, donde
se obtiene la mayoría de sustancias no gaseosas, y de los pulmones, de donde
provienen los gases nutritivos, que para los animales generalmente es el
dioxígeno gaseoso
2.2 Desechos
Los desechos son producidos por las
células como las sustancias finales del metabolismo. Estas sustancias deben
excluirse del fluido interno de la célula o citoplasma, pues al acumularse,
detienen el metabolismo y sin metabolismo no hay vida. Las plantas generalmente
no excretan sino que almacenan los desechos en las vacuolas de forma densa y
los modifican para defenderse. Los animales no almacenan los desechos, sino que
los excretan hacia la MEC por los mecanismos de transporte a través de
membrana. Una vez en la MEC los desechos atraviesan los capilares nuevamente
por los mecanismos de transporte a través de membrana hacia la sangre, y de
allí irán a los órganos excretores. Para los gases de desecho el órgano de salida
es el pulmón, siendo este gas principalmente el dióxido de carbono
2.3 Mensajeros
Los mensajeros químicos sus sustancias que
emplean los seres vivos para coordinar sus células a largas distancias y los
denominamos formalmente como las hormonas. Las hormonas fluyen desde las
células productoras hasta las células blanco por los mismos mecanismos que los
desechos y nutrientes. La sustancia sale de la célula la MEC, de la MEC al capilar,
de allí fluyen hasta el otro capilar donde el proceso se revierte desde el
capilar a la MEC y de allí al tejido blanco.
2.4 Xenobióticos
Ingresan por las mismas rutas que los
nutrientes, pero son claramente toxinas que obstruyen el metabolismo, pero que
el cuerpo confunde por nutrientes, suicidándose en el proceso, los casos
típicos son el plomo, el mercurio y el cromo(6+) (Cuberos, Rodriguez, &
Prieto-Suárez, 2009; Cuberos, Rodriguez, & Prieto, 2009; Forero, Mantilla,
& Martínez, 2009; Hernberg, 2000; Rossi, Gorman, & Wetterhahn, 1988)
3. Mecanismos de transporte a través de membrana
|| Regresar al Índice ||
Como mismos anteriormente, las sustancias
transportadas llegar y salen del sistema circulatorio a través de los
mecanismos de transporte a través de membrana, por lo que es una buena idea
repasar los conceptos asociados a estos procesos.
3.1 Transporte
pasivo
Cuando una sustancia en estado acuoso o disuelto realmente está separada en dos cantidades por una membrana semipermeable a dicha sustancia, esta fluirá libremente de forma tal que las concentraciones a ambos lados tiendan a igualarse. La velocidad de flujo es mayor a medida que la diferencia de concentración –también llamada gradiente –es más alta, y la velocidad aparenta ser cero cuando las concentraciones son las mismas. Solo los nutrientes más pequeños o que son solubles en grasas son capaces de atravesar las membranas de forma pasiva, todo lo demás requiere de un transporte mediado por proteínas para que sea eficaz y eficiente.

Figura 3.1. El transporte
pasivo se debe a la difusión, que es la tendencia natural de las partículas
disueltas a fluir en todas direcciones al azar.
3.2 Transporte
a través de membrana mediado por proteínas
El transporte de sustancias también
involucra trasladar a las malas, ya sea porque se requiere un flujo muy rápido
ya que los tejidos demandan un tipo de sustancia con gran impaciencia; o a que
hay un gas acumulándose rápidamente que resulta tóxico. El transporte activo
emplea proteínas que gastan energía celular, lo que les permite transportar
nutrientes en contra del gradiente de concentración, mientras que el transporte
pasivo facilitado emplea proteínas que aceleran el paso de nutrientes a favor
del gradiente de concentración.
Algunas proteínas empleadas en el
transporte de nutrientes no están ancladas a la membrana, pero si colaboran a
que las concentraciones en un lado de la membrana sea lo suficientemente alto
como para que el proceso osmótico se acelere. A diferencia del intercambio de
gases, la absorción de nutrientes durante los procesos digestivos requieren de
proteínas, y principalmente de transporte activo, por tal razón se dice que la
digestión es un proceso que gasta energía, y que algunos alimentos agotan más
que otros a la hora de digerirlos.
3.3 La
capilaridad
En el sistema circulatorio la capilaridad no se logra exactamente por pliegues sino por ramificaciones que aumentan la superficie de contacto del vaso sanguíneo y a su vez disminuyen el grosor del epitelio sanguíneo, aumentando la tasa de difusión o de transporte de sustancias hasta que se llega al optimo, donde el grosor es de una sola célula.

Figura 3.2. La capilaridad
sanguínea no se logra por pliegues, sino por ramificaciones de los vasos hasta
el punto que los glóbulos rojos solo pueden pasar en fila, uno a la vez.
4. Los fluidos internos, la sangre y otras cosas
|| Regresar al Índice ||
El sistema circulatorio es un medio para
el flujo de materiales disueltos en una entidad líquida, la cual evidentemente
es en su mayoría agua con muchos materiales disueltos. Sin embargo, el nombre
que le damos al fluido interno cambia dependiendo del tipo de ser vivo: sangre,
linfa, hemolinfa, savia, matriz extracelular, entre otros. Por esa razón
emplearemos el concepto de fluido interno como la traducción más cercana de la
idea de milieu interieur, como el
líquido que fluye a través de un sistema circulatorio.
4.1 El citosol y otros fluidos internos
Ya sé que suena muy celular, pero el citosol
es el primer fluido interno de transporte que puede existir, y también el
objetivo de llegada de las sustancias que transportan los demás fluidos. El
citosol o matriz citoplasmática (inglés: cytosol) es el líquido que se localiza
dentro de las células. Constituye la mayoría del fluido intracelular (por sus
siglas en inglés ICF). Está separado por membranas en distintos
compartimientos. Por ejemplo, la matriz mitocondrial separa la mitocondria en
varios apartados.
En las células eucariotas, el citosol se
encuentra dentro de la membrana celular y está incluido en el citoplasma,
citoplasma que también abarca la mitocondria, plastidios, y otros organelos. El
citosol no abarca los fluidos internos ni estructuras de los organelos; el
núcleo celular es independiente. El citosol, es entonces, un líquido de matriz
alrededor de los organelos. En procariotas, la mayoría de las reacciones
químicas del metabolismo toman lugar en el citosol, mientras algunas otras
ocurren en las membranas o en el espacio periplásmico. En eucariotas, si bien,
numerosas rutas metabólicas aún ocurren en el citosol, otras son contenidas
dentro de los organelos.
El citosol es una mezcla compleja de
sustancias disueltas en agua. A pesar de que el agua forma la mayor parte del
citosol, su estructura y propiedades dentro de las células no es bien
comprendida aún. La concentración de iones como el sodio(1+) y potasio(1+) es
diferente en el citosol que en el fluido extracelular MEC; estas diferencias en
los niveles iónicos son importantes en procesos como la regulación osmótica,
señalización de células y la generación de potenciales de acción en células
excitables como las células endocrinas, nerviosas y musculares. El citosol,
también contiene grandes cantidades de macromoléculas, las cuales, pueden
alterar el comportamiento de las moléculas a través de la aglomeración
macromolecular.
A pesar de que alguna vez se llegó a
pensar que el citosol era una solución simple de moléculas, este tiene
múltiples niveles de organización. Dichos niveles incluyen gradientes de
concentración de moléculas de pequeño tamaño, tales como el calcio(2+),
complejos enzimáticos de gran tamaño, los cuales en conjunto llevan a cabo las
rutas metabólicas, y complejos proteicos, tales como los proteosomas y
carboxisomas los cuales delimitan y separan las partes del citosol. Al interior
del núcleo y los organelos se encuentran fluidos homólogos al citosol, con sus
propios componentes, pero igual la sustancia más importante sigue siendo el
agua donde se disuelven las diferentes sustancias.
4.2 Matriz extracelular MEC
Para un ser vivo multicelular muy simple la matriz extracelular es el primer fluido interno, la matriz extracelular (MEC) es el conjunto de materiales extracelulares que forman parte de un tejido. La MEC es un medio de integración fisiológico, de naturaleza bioquímica compleja, en el que están "inmersas" las células. Así la MEC es la sustancia del medio intersticial (intercelular). La MEC es un componente de vida importante. Los animales con células se distinguen por su capacidad de interconectarse una morfogénesis compleja que implica asociaciones celulares cooperativas para formar tejidos. Ahí es donde es importante y distintiva la MEC como componente cohesivo y medio logístico de integración de las diferentes unidades funcionales celulares.

Figura 4.1. La matriz
extracelular o MEC es el medio donde habitan las células en un tejido, y está
compuesto por agua, sustancias disueltas en ellas y sustancias no solubles como
el colágeno, azúcares complejos o polisacáridos, minerales como la
hidroxiapatita “en el hueso” entre otras sustancias.
Toda sustancia que va a tener acceso a una
célula interna debe pasar primero por la MEC, siendo esta un análogo al medio
de cultivo que los científicos emplean para hacer crecer los tejidos de forma
artificial. En seres vivos simples, la matriz extracelular y las células se
encuentran cohesionadas por los componentes no fluidos de dicha matriz, que son
generalmente proteínas fibrosas. En los eucariotas la matriz extracelular está
formada por fibras de azúcares complejos que le otorgan una mayor viscosidad a
la matriz y proteínas de adhesión que mantienen a las células unidas entre sí.
La cantidad, tipo de proteínas, polisacáricos y otras sustancias como minerales
afectará las propiedades de dicha matriz. Algunas matrices son muy fluidas y
especializadas, por ejemplo, la misma sangre que mencionaremos en un apartado
diferente también puede ser considerada como un tipo de tejido cuya MEC es el
plasma o parte fluida de la sangre. Pero otras matrices pueden llegar a
extremos de viscosidad tales que son sólidas, como el hueso en el tejido óseo.
Sin embargo, la mayoría oscila entre un material muy viscoso a un material
gelatinoso.
Las funciones de la MEC son una extensión
de las características físicas de los componentes de la misma, y como
mencionamos anteriormente, son variables dependiendo de la viscosidad del MEC.
Las más generales son:
- Rellenar los espacios entre las células.
- Permitir la compresión y estiramiento de
las células.
- Medio inmediato para expulsar desechos
metabólicos y para la toma de nutrientes, así es, las células excretan y se
nutren del mismo medio la MEC.
- Regeneración de tejidos.
Debido a que la mayoría de las matrices
son viscosas o gelatinosas fluyen poco. El problema no es grave si hablamos de
un ser vivo pequeño compuesto por una cutícula externa, una MEC y pocas células
poco especializadas. Dado que hay pocas células y la MEC es delgada, los desechos
y nutrientes pueden pasar la cutícula por los mecanismos de transporte a través
de membrana y difundirse aleatoriamente por el MEC a velocidades aceptables.
Pero a medida que el ser vivo aumenta, esta viscosidad del MEC hace que la
difusión aleatoria deje de ser un mecanismo viable para la distribución de
sustancias de nutrición o desecho.
Lo anterior impone una restricción básica,
la MEC siempre permanecerá como un fluido viscoso delgado encerrado en un
compartimento concreto al cual deben llegar las sustancias por medio de otro
fluido menos viscoso, que fluya más rápido y permita que la velocidad de
difusión sea más eficiente. Para resolver este dilema fisiológico generalmente
se hace uso de una matriz extra-tisular especializada que interconecte las zonas
de nutrición y excreción con las zonas de funcionamiento. Estas nuevas matrices
extra-tisulares especializadas reciben nombres diferentes, aunque
evolutivamente sean lo mismo que la MEC tisular, solo que menos viscosas, y las
llamamos sangre, hemolinfa, linfa o savia.
4.3 Savia
La savia es el fluido o líquido transportado por los tejidos de conducción de las plantas (xilema o floema). Otros líquidos exudados por las plantas, tales como látex, cerumen, resinas o mucílago, muchas veces son incorrectamente denominados savia. La savia transportada por el xilema (denominada «savia bruta») consiste principalmente en agua, elementos minerales, reguladores de crecimiento y otras sustancias que se hallan en disolución. El transporte de esta savia se produce desde las raíces de la planta hasta las hojas por los tubos leñosos. En el siglo XX ha existido una gran controversia acerca del mecanismo de transporte de la savia bruta en la planta. Actualmente, se considera que toda la evidencia sustenta la teoría de la cohesión-tensión o de émbolo en la que el agua evaporada de la copa crea un efecto de embolo con respecto al agua de las raíces como si fuera una enorme jeringa. La savia elaborada es transportada por el floema de forma basípeta (desde su lugar de formación, hojas y tallos verdes, hacia la raíz) y está compuesta principalmente por agua, azúcares, fitorreguladores y minerales disueltos. El transporte de la savia en el floema se produce desde las fuentes (el lugar donde los carbohidratos se producen y almacenan) hacia los destinos (lugares de la planta donde los carbohidratos se utilizan). La hipótesis de flujo de presión es el mecanismo generalmente aceptado para explicar el transporte floemático.

Figura 4.2. La savia no
tiene color ya que no transporta gases a través de sustancias químicas
coloreadas llamadas pigmentos.
Las plantas representan claramente la
hipótesis evolutiva planteada anteriormente. Las algas unicelulares no
requieren matriz extracelular, mientras que las pluricelulares poseen una
matriz extracelular simple y un tegumento externo que mantiene unido al
conjunto celular. A medida que estas fueron evolucionando se requirió de un
fluido conector y esa fue la savia para los musgos. Sin embargo, sin un sistema
circulatorio la eficiencia de la savia se vio limitada a permitir el
crecimiento de la planta a unos cuantos milímetros. Solo cuando el fluido
conector se encuentra con un sistema de circulación, las propiedades menos
viscosas de dicho fluido les permiten a los nutrientes moverse con mayor
eficacia y en consecuencia el individuo puede crecer mucho.
El fluido de transporte de las plantas es
vascular, conecta las zonas de nutrición con los tejidos que no pueden nutrirse
por sí mismos por medio de tubos hechos de células especializadas, sin embargo,
las células propias del tejido reciben los nutrientes por intermedio de la MEC
que, si entra en contacto con la savia, ya sea bruta o elaborada. La
vascularización es tan especializada que hay conductos diferentes para la savia
con componentes inorgánicos básicos y otros para la savia que transporta el
alimento.
4.4 Hemolinfa
Del griego haima que significa sangre y del latín limpha: agua. Líquido del celoma o hemocele de algunos invertebrados (en especial artrópodos y la mayoría de los moluscos a excepción de la clase cefalópodos que poseen un sistema circulatorio cerrado) que es el líquido homólogo a la sangre de los vertebrados. La hemolinfa es otro tipo de fluido de transporte que conecta la MEC con las zonas de nutrición y excreción, pero a diferencia del de las plantas, la vascularización es limitada, esto implica que la hemolinfa rellena al animal como si se tratara de una piñata.

Figura 4.3. Hemolinfa de
una araña, en los artrópodos la sangre tiende a no tener colores debido a que
el sistema respiratorio es más independiente del circulatorio, a demás son
animales pequeños.
Hay muchísimos tipos de hemolinfa ya que
el único criterio que empleamos para definirla, es el hecho de que sea un
fluido de transporte que no se encuentra encerrado por una tubería “sistema
vascular” completo. Puede ser de diferentes colores o incluso incolora; los
pigmentos suelen proceder de la alimentación o de los procesos metabólicos y no
tienen ninguna función biológica, ya que el transporte de gases es
independiente del aparato circulatorio en los animales que tienen este fluido,
que son los artrópodos. Contiene células sanguíneas de diferentes tipos y
funciones.
4.5 Linfa
En los animales vertebrados la
vascularización es análoga a la de las plantas, tanto así que se generan dos
tipos de conductos para dos tipos de fluidos, aunque evidentemente no los
llamamos savia bruta o elaborada, pues la composición de dichos fluidos es
diferente. Los dos sistemas de conductos vasculares se denominan el sistema
circulatorio linfático y el sistema circulatorio general. La linfa es el fluido
que recorre el sistema circulatorio linfático, siendo principalmente un sistema
para regular la concentración de la MEC, al no recibir eritrocitos no posee
coloración roja. Se produce tras el exceso de líquido que sale de los capilares
sanguíneos al espacio intersticial o intercelular, siendo recogida por los
capilares linfáticos que drenan a vasos linfáticos más gruesos hasta converger
en conductos que se vacían en las venas subclavias.
La linfa recorre el sistema linfático gracias a débiles contracciones de los músculos, de la pulsación de las arterias cercanas y del movimiento de las extremidades. Ahora bien, dijimos que su función es la de regular la concentración del MEC, pero que pasa si por alguna razón, como una obstrucción, ¿el sistema linfático no puede cumplir su objetivo? La consecuencia es que del sistema circulatorio general ingresará agua al MEC y el linfático no la drena, por lo que el MEC se empezará llenar de agua, alterando los delicados equilibrios osmóticos y eléctricos, lo cual impide que la célula se alimente, excrete, esto a nivel celular produce muerte del tejido por acumulación de líquido, proceso denominado edema.

Figura 4.4. La linfa
vertebrada tiende a ser blanca debido a su alto contenido de células del
sistema inmune o células blancas.
Este fluido está compuesto por un líquido
claro pobre en proteínas y rico en lípidos, parecido a la sangre, pero con la
diferencia de que las únicas células que contiene son los glóbulos blancos, que
migran de los capilares y proceden de los ganglios linfáticos, sin contener
glóbulos rojos. También puede contener microorganismos que, al pasar por el
filtro de los ganglios linfáticos, son eliminados. La linfa es menos abundante
que la sangre: se considera que hay aproximadamente 2 litros de linfa, mientras
que el volumen de sangre es de unos 5 litros.
4.6 La sangre
Es el segundo líquido altamente fluido de los vertebrados, y como sus demás contrapartes transportará nutrientes y desechos. La característica más importante de la sangre y que comparte con la hemolinfa es la posesión de pigmentos que incrementan la eficiencia en el transporte de gases metabólicamente importantes como el oxígeno y el dióxido de carbono. En los vertebrados esta sustancia fluye a través del sistema circulatorio general. A parte del pigmento de transporte de gases metabólicos que llamamos hemoglobina y que es de colores rojizos, la sangre también transporta otras sustancias que mejoran la eficacia del transporte de nutrientes.

Figura 4.5. La sangre es
roja debido a que sus células eritrocitarias llevan un pigmento de hierro que
permite transportar gases de importancia metabólica.
Un ejemplo de eso son las lipoproteínas de
alta y baja densidad encargadas de distribuir y recolectar las grasas desde y
hacia el hígado según requiera el metabolismo del organismo. Adicionalmente
también trasporta nutrientes que se disuelven fácilmente en agua y pueden ser
transportado sin necesidad de proteínas de transporte, así como componentes
auxiliares, como células del sistema inmune y materiales para reparar los vasos
sanguíneos en caso de roturas. La mayor parte del presente capítulo de anatomía
comparada se dedicará al estudio de los vasos conductores por donde fluye la
sangre, más que a la sangre misma.
5. La función de transporte de sustancias en unicelulares
|| Regresar al Índice ||
Cuando hablamos de un sistema circulatorio
casi que por definición deberíamos excluir a los seres vivos unicelulares, sin
embargo, debemos recordar que lo que estamos analizando no es solo el sistema
de órganos, sino también el cumplimiento de una función biológica, en este
caso, el transporte de sustancias de un lugar a otro de las células.
Evidentemente a nivel subcelular se repiten temas que tratamos en todos estos
capítulos introductorios, pero ¿qué sucede al nivel de las diferentes células
en seres vivos que no forman tejidos?
5.1 Sistema circulatorio en las bacterias
¿Se puede hablar de un sistema
circulatorio para las bacterias? La sola mención de esta idea podría parecer
risible para muchos, y sin embargo sí, es posible hablarlo, y se puede hablar
tanto en su interior como en su exterior. Al interior de las bacterias el
proceso es principalmente aleatorio por medio de la difusión simple de
sustancias, en la cual con el tiempo una sustancia soluble en agua o
solubilizada a las malas termina por distribuirse homogéneamente en toda la
célula, aunque claro, este proceso es universal y cualquier célula lo tiene, no
solo las bacterias, sin embargo, las bacterias dependen más de esto porque su
transporte de sustancias mediado por vesículas es inexistente o muy limitado. Las
bacterias pueden asumir esto dado que son muy pequeñas, lo cual hace que la
difusión sea un proceso viable. Sin embargo, algunas sustancias no pueden
transportarse únicamente por la difusión simple, y deben ser encerradas en
micro-vesículas para su transporte a las diferentes partes de la célula
bacteriana. Aunque estas micro-vesículas son más pequeñas que sus análogos
eucariotas, se encargan en últimas de la función base, transportar sustancias
de un lugar a otro de la célula (Droleskey, Byrd, Bischoff, Harvey,
& Nisbet, n.d.; Lee et al., 2015; Spiewak & Dutkiewicz, 2008)
Lo más relevante médicamente es el sistema
circulatorio que pueden construir muchas células bacterianas. Ahora, algún
listillo podría preguntar en este punto ¿y no que eran unicelulares? Y la
respuesta es sí, pero no. Aunque clásicamente las bacterias son vistas como
seres unicelulares que no cooperan entre sí, esto se debe a que la
microbiología tiende a estudiarlas en solución, es decir bacterias individuales
flotando en condiciones óptimas para que crezcan rápido, se puedan estudiar
pronto y se rebajen los costos de estudio. Sin embargo, desde hace unas cuantas
décadas los microbiólogos se han dado cuenta de que en condiciones no óptimas
las bacterias se asocian formando los biofilms, biopelóculas o bioplacas. ¿Y
qué tiene que ver una bioplaca con un sistema circulatorio? Vamos por partes,
primero discutiremos un poco de cómo funciona una bioplaca bacteriana (De Beer, Stoodley, &
Lewandowski, 1996; De Beer, Stoodley, Roe, & Lewandowski, 1994; Finlay,
Schultz, Cone, Callow, & Callow, 2013; Kåhrström, 2013; Speziale &
Geoghegan, 2015)
5.2 Anatomía
de las biopeliculas
Las bacterias oscilan entre dos modos de
vida, el de individuos disueltos en el medio acuoso y el de bioplacas. Las
biplacas se forman cuando las condiciones se hacen adversas por medio del
sistema de sensibilidad de densidad poblacional (Speziale & Geoghegan,
2015).
👉 Sistema de sensibilidad a la densidad poblacional: También conocido como el sistema de sensibilidad de quórum, se emplea para coordinar ciertos comportamientos como: la formación de biopelículas, la transformación de una forma comensalista a otra virulenta, o la activación de sistemas de resistencia a antibióticos. El sistema de sensibilidad a la densidad poblacional “quorum sensing” (Costerton, Stewart, & Greenberg, 1999; Miller & Bassler, 2001; Singh et al., 2000) puede ocurrir al interior de individuos de una sola especie, o en individuos de diversas especies de bacterias. Como cualquier sistema de control biológico, el sistema de sensibilidad a la densidad poblacional se basa en la producción de señales químicas que se acopla a receptores específicos de las membranas de las bacterias.

Figura 5.1. A la izquierda
tenemos cuatro bacterias, de dos especies diferentes con baja densidad
poblacional, a la derecha tenemos muchas células apuñadas, lo cual dispara el quorum sensing.
Las bacterias capaces de emplear el
sistema de sensibilidad a la densidad poblacional, producen basalmente una
cantidad mínima de una señal química denominadas autoinductores o feromonas.
Estos mensajeros químicos tienen un efecto exocrino y autocrino. Como el
mensajero tiene un efecto autocrino, si su cantidad aumenta en el medio donde
habita la bacteria, esta se estimula, lo cual significa que hay muchas
bacterias viviendo juntas, es como un mal olor colectivo. Si todas bacterias
producen la cantidad basal la concentración externa aumenta proporcionalmente a
la cantidad de bacterias y en consecuencia el estímulo aumenta en cada bacteria
para producir mayor cantidad del mensajero químico (Costerton et al., 1999;
Miller & Bassler, 2001; Singh et al., 2000).
Cuando la población es pequeña, el
mensajero químico es diluido rápidamente y no es capaz de inducir una respuesta
fisiológica efectiva, tal como ocurre con las hormonas, o con los malos olores.
Sin embargo, cuando la población aumenta a un límite crítico, el mensajero
químico se acumula, generando un cambio en el comportamiento de la población
completa aun cuando estemos hablando de especies unicelulares distintas. A esto
se lo conoce como el sistema de refuerzo positivo, en el cual la mayor cantidad
de población induce un efecto mayor del mensajero químico (Costerton et al.,
1999; Miller & Bassler, 2001; Singh et al., 2000), lo cual también puede
explicar la evolución de la multicelularidad en otros seres vivos de tipo
eucariota.
👉 Las bioplacas o
biopelículas: En una bioplaca
las bacterias se encuentran aisladas del medio externo por medio de la
secreción de una sustancia viscosa y pegajosa que se denomina por homología o
analogía como matriz extracelular MEC (Borlee et al., 2010; Bowen &
Koo, 2011; Hawser, Baillie, & Douglas, 1998).

Figura 5.2. Las bioplacas
son semejantes a los tejidos, ya que no solo es un conjunto de células, sino
que estas están imbuidas en una matriz extracelular que les otorga habilidades
que no tendrían las células por separado, una de esas habilidades es la
resistencia a antibióticos colectiva.
En este sentido, la matriz extracelular
funciona como el fluido interno al interior del cual los mensajeros químicos
son segregados de forma interna, cumpliendo con la definición de la función
endocrina. Las bacterias que generan biopelículas generalmente también poseen en
sistemas de sensibilidad a la densidad poblacional, por lo que segregan
químicos mensajeros que coordinan sus esfuerzos para colonizar un ambiente.
👉 La circulación de nutrientes en una bioplaca: Si fue difícil aceptar que las bacterias no eran tan unicelulares como las habíamos definido desde hace siglos, ha sido aún más aceptar el hecho de que sus conglomerados proteínicos no son una masa simple que permite la difusión molecular. ¿A que nos referimos con esto? Que el MEC bacteriano no es uniforme, sino que está estructurado en una masa gelatinosa y poros fluidos donde el agua puede atravesar, esto aumenta la superficie de intercambio de sustancias. Hay que tener en cuenta que una bioplaca debe desenvolverse como una unidad general, y al hacerse más grande, comienza a ser restringida por el dilema área volumen, a mayor volumen menor área y menos superficie para el intercambio de sustancias, lo cual provoca la posibilidad de que las células del centro de la masa se mueran ya sea por falta de nutrientes, acumulación de toxinas o ambas. Una de las primeras pistas de la existencia de canales en una bioplaca fue el reporte de poros que absorbían partículas de látex, las cuales se fundían más rápido de lo que se esperaría en un MEC viscoso uniformemente distribuido (Drury, Stewart, & Characklis, 1993).

Figura 5.3. Las
biopelículas poseen un sistema de canales de agua que las atraviesan, semejante
al sistema acuífero de las esponjas “algo semejante a los huecos de un queso
gruyere”, lo cual les permite crecer más de lo que sería posible de ser
completamente sólidas.
La microscopía de escaneo focal ha revelado
desde hace ya unas cuantas décadas que las bioplacas no son estructuras
uniformes, sino que en su interior las bacterias se agrupan en nidos celulares
interconectados por una compleja red de canales donde el MEC se hace más fluido
o simplemente desaparece para dar lugar a canales de agua por donde fluyen los
nutrientes y los gases de importancia metabólica, lo cual es precisamente un
análogo de un sistema circulatorio verdadero (De Beer et al., 1996, 1994; Finlay
et al., 2013; Speziale & Geoghegan, 2015; Wilking et al., 2013)
5.3 Transporte de sustancias de los eucariotas
Los eucariotas internamente a parte de la
difusión de sustancias, poseen un mecanismo de transporte especializado mediado
por membranas internas denominado simplemente como el sistema de transporte
vesicular. En ese sentido tanto digestión, excreción, circulación y envió de
mensajeros químicos en últimas dependen de los mismos procesos celulares.
En la Figura 5.4 podemos ver un resumen general del proceso, este esquema sirve para otros sistemas debido a que a nivel celular muchas funciones se realizan por mecanismos conservados evolutivamente. En este caso podemos ver: (1) el retículo endoplasmático rugoso donde se sintetizan enzimas y proteínas, en este caso enzimas digestivas, que luego (2) son trasportadas por vesículas al aparato de Golgi donde (3 y 4) son maduradas en sus formas activas, para (5) ser emitidas al medio externo, o fusionarse con otras vesículas para modificaciones secundarias y ser emitidas al medio externo (5b). Por lo general la digestión enzimática empieza con (7) fagocitosis o endocitosis, la vesícula formada es enviada al aparato de Golgi directamente (7c) o fusionada a una vesícula que transporta las enzimas líticas (7a, 7b), los productos de la digestión se absorben por los mecanismos de transporte a través de membrana y los desechos eliminados (8).

Figura 5.4. Modelo de transporte vesicular
en una célula eucariota, el sistema de membranas internas no es rígido, y
funcionan como bombas de jabón remolcadas por proteónas, estas pueden
fusionarse con otras bombas, pequeñas “vesículas” o grandes “retículos
endoplasmáticos o aparato de Golgi” o con la membrana externa transportando
sustancias en su interior.
Más allá de los procesos individuales,
bajo la perspectiva de un sistema circulatorio lo que nos importa es como el
sistema vesicular funciona como un mecanismo de transporte de sustancias de un
lugar a otro de la célula de forma no aleatoria. Al contrario de la difusión
que es al azar y debe distribuir los materiales homogéneamente, el sistema de
transporte vesicular envía lo que se necesitan, a donde se necesita y en las
cantidades que se necesitan, aunque claro el precio a pagar para ello es una
inversión de energía para formar las vesículas, para fabricar el citoesqueleto
y las proteínas de transporte que son las quinesinas y las dineinas, y
finalmente la energía para que las proteínas de transporte arrastren las
vesículas al lugar donde se necesitan.
Más allá de esto, las células eucariotas
pueden formar conglomerados poco especializados, pero de una naturaleza
semejante a las biopelículas, con un MEC que protege y regula la interacción
con el medio, pero en los eucariotas algunos linajes especializaron más el modo
de vida comunitario para formar tejidos especializados, lo cual requirió el
desarrollo de verdaderos sistemas circulatorios intermediaros entre el medio y
el MEC, sistemas que analizaremos con mayor detalle en las siguientes
secciones.
6. Plantas
|| Regresar al Índice ||
Las plantas verdes abarcan una gran cantidad de formas de vida, desde las algas verdes unicelulares, hasta las plantas con flor. ¿Cuáles de estas poseen sistemas de circulación? Primero definiremos que un verdadero sistema circulatorio es aquel que conecta varios tejidos independientes a través de sus respectivos MEC a los tejidos de obtención de nutrientes, en otras palabras, un sistema circulatorio verdadero se encuentra por fuera del MEC de un tejido concreto. Uno de los parámetros más obvios que aparasen a la hora de requerir un sistema de circulación es simplemente el tamaño, todas las plantas muy pequeñas o lo suficientemente delgadas, en las que no existe especialización de labores entre los tejidos nunca presentan un sistema circulatorio. En esta categoría podemos englobar a las algas verdes unicelulares, a las algas verdes multicelulares, y a los musgos.
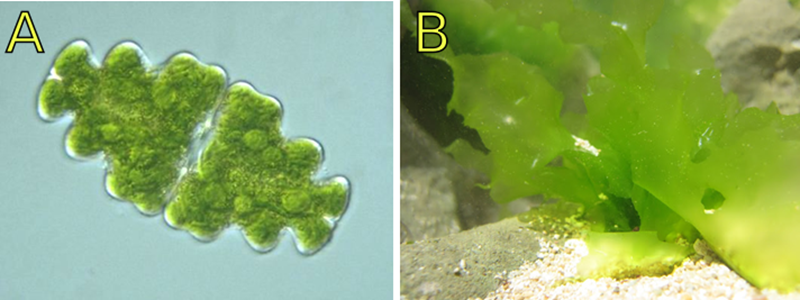
Figura 6.1. (A) Euastrum
sp., (B) Ulva sp.
Las algas son el tipo de plantas más
antiguo y engloba a individuos de una célula y de muchas células. El género Euastrum
sp., es un alga verde unicelular. Aunque sean unicelulares aun las
categorizamos como plantas verdes debido a su estrecha relación genética con
las plantas terrestres más complejas. El género Ulva sp., de algas
verdes a diferencia de otras como Euastrum spp., se caracteriza porque
un solo individuo está compuesto por millones de células, sin embargo, todas
las células de su cuerpo son iguales, y por lo tanto no hay especialización de
funciones o de tejidos. Cada célula se encarga de todo con una eficiencia
mínima pero una independencia máxima.
Los musgos por su parte ya poseen cierta
especialización con estructuras diferenciadas para aferrarse al sustrato y para
hacer la fotosíntesis, sin embargo, son tan minúsculos que la mera capilaridad
de sus tejidos es suficiente para contrarrestar el efecto de la aceleración
gravitacional que impulsa los fluidos hacia el suelo. Desde los helechos hasta las plantas con flor
ya tenemos seres vivos demasiado grandes como para que todos los tejidos se
encarguen de todas las funciones, y por lo tanto se requiere de un sistema de
transporte.
En las plantas terrestres, el sistema de
transporte es vascular, es decir un sistema de transporte cerrado por tubos,
similar al de los vertebrados, los tejidos especializados en el transporte son
denominados xilema y floema. El xilema y el floema se encargan de conducir el
agua y minerales solubles en esta, desde las raíces donde estos materiales son
absorbidos en el suelo por medio de los mecanismos de transporte a través de
membrana, hacia los tallos, las hojas y las demás estructuras vivas de la
planta. También se encargan del transporte de azúcares y otros productos de
alta energía producidos por la fotosíntesis y modificados en las hojas de la
planta, ya sea para mantener vivos a los tejidos del tallo o las raíces, o para
generar estructuras de reserva como en ciertos tipos de raíces modificadas. El
sistema vascular de las plantas es continuo, e integra a las raíces, los tallos
y a las hojas en una sola autopista de transporte, la cual permite a la planta
vascular funcionar como una sola unidad, aunque sus tejidos desempeñen diversas
funciones.
A continuación, enfocaremos este espacio
para la descripción de la anatomía y fisiología del sistema circulatorio de las
plantas traqueofitas denominado floema y xilema.
6.1 Estructura general del bulbo vascular
Por lo general, al revisar un corte
transversal del tejido de una planta, nos encontraremos con que existe una
diferenciación entre los tejidos:
1- Las células externas que sirven de
protección (epidermis o peridermis)
2- Los tejidos basales: fotosintéticos, de almacenamiento y
soporte.
3- El tejido vascular con forma de bulbo central, el cual contiene al xilema, al floema y a otros tejidos accesorios dependiendo del grupo al que la planta pertenece.

Figura 6.2. El bulbo
vascular es la zona donde se agrupan los tejidos floema y xilema.
👉 Eudicotiledóneas: En las plantas herbáceas eudicotiledoneas, los tejidos basales son fotosintéticos siempre que sean verdes. Los bulbos vasculares forman una especie de anillo externo en torno al tejido de soporte o médula. En el modelo anterior podemos apreciar la disposición de los tejidos vasculares en el tallo de una eudicotiledonea (Figura 6.3, izquierda). Se puede ver como los tejidos vasculares conforman una serie de tubos que se disponen radialmente en la parte externa del tallo. En la microfotografía (Figura 6.3, centro) podemos apreciar un corte transversal del tallo de una planta eudicotiledónea, Aquí podemos apreciar con cierta claridad la región vascular o bulbo vascular que contiene al floema y al xilema. En la moicrofotografía (Figura 6.3, derecha) tenemos una ampliación de la primera microfotografía donde podemos apreciar de mejor modo los componentes del bulbo vascular, donde el floema se ubica más cerca del centro, mientras que el xilema se ubica más cerca de la superficie del tallo.

Figura 6.3. Organización
de los tejidos vasculares de las eudicotiledóneas, de izquierda a derecha,
modelo, microfotografía de microscopio óptico u microfotografía de microscopio
electrónico.
👉 Monocotiledóneas: En los tallos de las monocotiledóneas, los bulbos vasculares no forman un anillo, en su lugar se encuentran esparcidos a través de toda el área del corte transversal del tallo, como se representa en la Figura 6.4.

Figura 6.4. Tejido vascular de las
monocotiledóneas, de izquierda a derecha, modelo, microfotografía de
microscopio óptico u microfotografía de microscopio electrónico.
6.2 La circulación en las plantas vasculares
Ahora que hemos visto la estructura
general y la posición de los bulbos vasculares que poseen el xilema y el
floema, pasaremos a ver como se da el transporte de sustancias desde las raíces
hasta las hojas y viceversa, lo cual no es precisamente algo sencillo. Por
ejemplo, los árboles secoya pueden ser literalmente tan grandes como montañas,
llegando a alcanzar más de 110 metros de altura, y subir el agua necesaria para
todas las hojas de la copa que lo mantienen vivo es una verdadera maravilla de
la biología, el agua de las raíces debe ascender entre 85 y 113 metros, ¡TODO
SIN UN CORAZÓN! ¿Cómo lo hacen?
Por otro lado, los productos de la
fotosíntesis, especialmente azucares, deben ser enviados desde las hojas hasta
las raíces, lo cual no es un proceso simple, el azúcar es una molécula compleja
que no se transfiere fácilmente a través de las membranas biológicas, por lo
que se requiere de transporte activo o de un mecanismo análogo a este llamado
translocación.
El transporte a través del xilema del
agua, y el transporte a través del floema de los azucares no se asemeja al
movimiento de los materiales a través del sistema circulatorio cerrado de los
vertebrados, debido a que en las plantas nada circula en un sistema de vasos
capilares interconectados. En otras palabras, en los animales con sistema
circulatorio cerrado, este se asemeja a un circuito cerrado, donde se le puede
dar la vuelta al sistema, al circular cíclicamente. En las plantas la cuestión
es diferente, el agua y los minerales corren en una única dirección (hacia
arriba), mientras que el transporte de azúcares disueltos puede darse en ambas
direcciones axiales (hacia arriba y hacia abajo). Otra de las diferencias
básicas entre los sistemas circulatorios de plantas y animales es que en las
plantas no se tiene un sistema muscular que presione a los vasos o un corazón
que le de potencia extra al flujo de sustancias.
6.3 El xilema
transporta agua y minerales, el modelo de tensión-cohesión
El agua se mueve de manera horizontal a
través de la raíz desde el suelo, atravesando varios tejidos hasta llegar al
xilema. Una vez que se lleva a los canales vasculares del xilema, el agua
asciende a través de una red continua de vasos que componen los elementos del
xilema hasta llegar a las hojas. Los
minerales solubles en agua, como iones de sales viajan por el agua a través de
transporte pasivo, en otras palabras, las plantas no invierten ninguna energía
en el paso de sustancias entre las membranas de los tejidos.
Pero, ¿Cómo es que se da este movimiento?,
la evidencia actual indica que el agua asciende al ser jalada por las células
de xilema. Básicamente nos encontramos ante unan mezcla de fenómenos, dos en
especial, una diferencia de presión y al mecanismo de capilaridad; lo cual se
engloba en el modelo de Tensión-Cohesión de Henry Dixon propuesto en 1896. La
capilaridad es una propiedad de los fluidos que depende de su tensión
superficial la cual, a su vez, depende de la cohesión del líquido y que le
confiere la capacidad de subir o bajar por un tubo capilar (Figura 6.5).
Más importante que la explicación química de la capilaridad, lo que hay que tener en cuenta es que, mientras más delgado sea el tubo, más se elevará el agua de su nivel básico sin importar la fuerza de gravedad y la presión atmosférica, ambas fuerzas empujando o jalando las moléculas hacia o desde el suelo respectivamente. Cuando un líquido sube por un tubo capilar, es debido a que la fuerza intermolecular o cohesión intermolecular entre sus moléculas es menor que la adhesión del líquido con el material del tubo; es decir, es un líquido que moja. El líquido sigue subiendo hasta que la tensión superficial es equilibrada por el peso del líquido que llena el tubo. Éste es el caso del agua, y esta propiedad es la que regula parcialmente su ascenso dentro de las plantas, sin gastar energía para vencer la gravedad. Sin embargo, cuando la cohesión entre las moléculas de un líquido es más potente que la adhesión al capilar, como el caso del mercurio, la tensión superficial hace que el líquido descienda a un nivel inferior y su superficie es convexa.

Figura 6.5. La cohesión del agua a las
paredes de un tubo provoca su ascenso en contra de la gravedad, cuanto más
delgado el tubo, más sube el agua.
Básicamente, entre más delgado es el capilar, más asciende el agua en contra de la gravedad sin necesidad de ejercer ninguna fuerza. Sin embargo, la gravedad no es la única fuerza que empuja las moléculas al suelo, en contra de los intereses de la planta, la otra es la presión, el peso del aire sobre un líquido el cual mantiene su volumen en cierta posición. Cuando la presión disminuye, el volumen ocupado por un líquido aumenta a una temperatura constante. Basados en ese principio, si en la punta del capilar se encuentra una presión menor a la presión atmosférica que enfrenta la planta, esta incrementará su volumen y ascenderá aún más por el capilar.

Figura 6.6. Los árboles
secoya o secuoya “Sequoiadendron giganteum” son árboles colosales, del
tamaño de montañas, levantar el agua requerida para alimentar las hojas de la
copa es un reto biológico.
Esto se logra mediante la evaporación del agua
a través de los estomas (recordemos que en las plantas terrestres, hay una capa
externa llamada cutícula, la cual evita la evaporación del agua, esta solo
puede evaporarse por aberturas microscópicas llamadas estomas). Cuando el agua
se evapora de los estomas, la presión disminuye en la superficie donde estos se
encuentran, es decir especialmente en las hojas, de esta manera, la punta de
las plantas posee una presión menor, mientras que la base posee una presión
mayor, y esta diferencia de presiones hace que el agua sea enviada hacia la
punta ara que las presiones se equilibre. El proceso es semejante a una
jeringa, la evaporación es como cuando el embolo se retira generando un vacío o
disminución de la presión, mientras que la aguja que es como una raíz capilar
permite el flujo de líquidos.
Aunque el modelo de tensión-cohesión fue
propuesto a finales del siglo XIX, no fue hasta finales de la década de 1990 y
principios de la década de los 2000 que se obtuvieron las primeras evidencias
experimentales de que el modelo era en efecto correcto. Sin embargo, es
conveniente hacernos una última pregunta, ¿es suficiente la capilaridad y la
diferencia de presión para explicar el ascenso del agua desde la raíz hasta la
última hoja de los gigantes secoya? Mediante mediciones de la diferencia de
presión por evaporación en las hojas de estos árboles, se ha estimado que sus
vasos capilares pueden hacer elevar el agua hasta una altura no mayor de 130
metros, sin embargo, los arboles más grandes no superan los 110 metros, por lo
que el modelo puede explicar el ascenso del agua aun en estos gigantes, que no
tienen tejido muscular o corazón.
6.4 El floema transporta azúcares
El mecanismo de transporte de azúcar y
otros productos de la fotosíntesis se basa principalmente en el transporte
activo. Como hemos dicho anteriormente, el azúcar, como la sucrosa es una
molécula demasiado compleja como para pasar a través de la membrana por
transporte pasivo, por lo que se requiere del mecanismo de transporte activo.
En otras palabras, para que el azúcar salga de las zonas donde es producido por
medio de la fotosíntesis (especialmente las hojas, pero cualquier zona verde lo
hace) y sea depositado en el floema, se debe invertir energía en el proceso.
El xilema y el floema se encuentran cerca uno
de otro, esto es importante ya que el floema no funcionaría sin el xilema.
Básicamente, algunas moléculas de azúcar serán sacrificadas para generar la
energía suficiente para que el resto pase desde la zona fotosintética al
floema. Una vez allí, el agua del floema al mezclarse con el azúcar genera una
presión osmótica negativa que provoca que parte del agua que venía subiendo por
el xilema se transporte al floema disolviendo parcialmente al azúcar.
La célula verde que contiene cloroplastos
se denomina fuente, ya que produce más azúcar “esferas rojas” del que ella
consume. De este modo, la solución azucarada desciende por el floema hasta los
tejidos donde lo requieren para generar energía o para transformarlo en aceites
u otras sustancias de almacenamiento. En el momento en que el azúcar empieza a
ser absorbido por los tejidos, se genera una presión osmótica positiva que la
empuja hacia afuera del floema y la hace regresar al xilema, donde vuelve a
subir. Cabe anotar que, por el floema también se transportan sustancias como
aceites, aminoácidos, hormonas y hasta virus que infectan a las plantas.
Cabe anotar que en floema puede hacer fluir al azúcar hacia arriba gracias al transporte activo, aunque es en todo caso un proceso lento. La célula receptora se denomina estanco, ya que esta consume más azúcar del que produce, si es que produce azúcar en primer lugar. Las raíces son ejemplos de estancos dado que no reciben luz y no poseen cloroplastos, y por eso no son verdes. Aunque se tienen datos de que este mecanismo general es el correcto, aún quedan muchas incógnitas sobre los detalles de su proceso.

Figura 6.7. Los productos
de la fotosíntesis se translocan activamente desde las células del mesófilo
hasta el floema (1), mientras que el agua fluye por transporte pasivo u ósmosis
(2), el agua atraviesa los filtros libremente, mientras que los productos
fotosintéticos deben atravesar activamente cada uno de los filtros (3)
generalmente hacia abajo, hasta los tejidos no fotosintéticos como frutas y
raíces (4). Los tejidos no fotosintéticos retienen los productos de la
fotosíntesis, y las raíces entregan agua y minerales al xilema (5), la cual
asciende por tensión-cohesión (6) hasta los tejidos fotosintéticos, donde estos
absorben el agua por ósmosis y los minerales por transportes mediados por
proteínas (7).
7. Generalidades del sistema circulatorio de los animales
|| Regresar al Índice ||
En cooperación con el sistema
respiratorio, el sistema circulatorio se encarga de distribuir los gases
metabólicos entre los tejidos y los órganos de intercambio de gases con el
ambiente. Sin embargo, esa no es su única función, virtualmente todo debe pasar
a través del sistema circulatorio para llegar desde el lugar donde se fabrica o
donde se obtiene a los lugares donde se requiere. A demás de lo anterior, el
sistema circulatorio posee otras funciones importantes, por ejemplo, se encarga
de ajustar la presión hacia y desde el interior del cuerpo. La sangre
transporta calor desde el interior del cuerpo hacia el exterior para disiparlo
(como si fuera una máquina térmica normal). Alternativamente, algunos animales
incapaces de controlar su temperatura interna o que simplemente no lo hacen
para no gastar energía de los alimentos pueden obtenerla del calor del medio
ambiente, la sangre en ese caso transporta el calor desde el ambiente hasta el
interior.
Básicamente, la función básica de
cualquier sistema circulatorio es el transporte. El transporte de materiales
desde un lugar a otro en el interior del organismo, y depende de los movimientos
y de la difusión de las sustancias en los fluidos internos del animal. Los
nutrientes, los gases, y los desechos metabólicos son generalmente
transportados en una mezcla en solución, anclados a otros compuestos solubles o
en el interior de células especializados para el transporte de sustancias.
Cualquier sistema de fluidos móviles que reduzca la distancia de difusión que
los compuestos químicos deben atravesar en el interior del cuerpo de un animal
es descrito como un sistema circulatorio, sin importar su origen embriológico,
su origen genético o su diseño anatómico y fisiológico.
La naturaleza del sistema circulatorio se
relaciona de manera directa con el amaño, complejidad y estilo de vida del
animal en cuestión. Por lo general, los fluidos circulatorios con ambientes
acuosos extracelulares y que se encuentran en constante movimiento producidos
por las células del animal. Sin embargo, existen algunos ejemplos en los que la
función del sistema circulatorio es realizada de manera parcial por otros mecanismos.
En todos los animales, los tejidos que fluyen intracelularmente juegan un rol
crítico como un medio de transporte. Aun cuando existen mecanismos complejos
para el bombeo, el tejido fluido sigue siendo necesario para distribuir los
materiales a las células que los requieren, y con ello mantener el proceso de
homeostasis. El tejido fluido se denomina sangre o hemolinfa y puede o no
portar células especializadas en el transporte de sustancias. Los glóbulos
rojos o eritrocitos son un ejemplo de células que transportan en su interior
materiales necesarios para mantener el metabolismo marchando, en este caso
transportan principalmente oxígeno y dióxido de carbono.
Más allá de los mecanismos poco
especializados para el proceso de circulación en los animales, existen dos
diseños principales por medio de los cuales los seres vivos llevan a cabo el
proceso de circulación. Estos son, la circulación abierta y la circulación
cerrada.
7.1 El sistema circulatorio abierto
Los sistemas circulatorios abiertos están asociados con una reducción del celoma en el adulto, incluyendo una perdida secundaria de la mayor parte del peritoneo a través de los órganos y en el interior de la superficie del animal. El sistema circulatorio abierto, por lo general incluye un corazón bien definido como el mecanismo de bombeo del fluido principal, así como varios vasos sanguíneos, cámaras o ventrículos pobremente definidos. El nivel de complejidad de dichos sistemas dependerá principalmente del tamaño, complejidad y hasta cierto punto actividad física del animal.

Figura 7.1. El sistema
circulatorio de los artrópodos es abierto, en la sangre fluye libre bañando la
cavidad interna o celoma y sus órganos, el vaso sanguíneo funciona como corazón
y le da movimiento a la sangre.
En el sistema circulatorio abierto, los
órganos se encuentran "flotando" en hemolinfa, y el cuerpo del animal
se encuentra "lleno" de esta, a diferencia de los animales con
sistemas circulatorios cerrados donde existe una cavidad vacía o celoma. El
corazón simplemente genera una corriente que le da movimiento a la hemolinfa,
pero la circulación es completamente caótica, generándose una mezcla poco
controlada.
A pesar de las antes nombradas semejanzas
con el sistema circulatorio humano por todos conocido y que es cerrado, el
sistema circulatorio abierto se denomina de tal forma porque en cierto punto
los vasos sanguíneos se abren a la cavidad principal del animal vaciando su
contenido allí, en otras palabras, nunca se forma un circuito.
La hemolinfa fluye libremente a través de
la cavidad interna del animal llamada hemoceloma, bañando con su flujo a los
órganos internos y a la pared externa del tracto gastrointestinal. El fluido
circulatorio en este caso es denominado hemolinfa, mientras que la cavidad
general en este caso será denominada hemoceloma. Los sistemas circulatorios
abiertos son típicos de los artrópodos y los moluscos no cefalópodos, y por lo
tanto estos animales son denominados de manera parafilética como hemocelomados.
Solo porque el sistema circulatorio abierto parezca algo simplote en
comparación con el sistema circulatorio cerrado, no debe ser visto como mal
diseñado o (y este es mi favorito) “menos evolucionado” o inclusivo
ineficiente.
De hecho, en muchos grupos de animales,
ese tipo de aparato circulatorio ha asumido una gran variedad de funciones más
allá de la circulación. Por ejemplo, en los bivalvos y gastrópodos, el
hemoceloma funciona como un esqueleto hidrostático (la rigidez del cuerpo de
estos animales funciona como un esqueleto hidráulico) para la locomoción y otro
tipo de actividades. En los artrópodos acuáticos. El esqueleto hidrostático
mantiene al animal con su forma mientras cambian de piel. Mientras que, en los
artrópodos terrestres, donde el transporte de oxígeno hacia el interior de los
tejidos es llevado a cabo por el sistema de tráqueas, el hemoceloma se encarga
principalmente de la termorregulación. En muchas arañas, el movimiento de las
patas se realiza por la presión de la hemolinfa (como gatos hidráulicos), y en
este caso el sistema circulatorio asume una función muscular.
En ocasiones el hemoceloma no es completo,
sino que se encuentra parcialmente separado en secciones por tabiques
conectados entre sí por puntos de acceso. En estos casos los compartimentos
individuales son denominados senos “no es mi culpa que tenga doble sentido”. En
caso de que los compartimentos sean aún más pequeños y aislados, serán
denominados lagunas.
7.2 El
sistema circulatorio cerrado
El sistema circulatorio cerrado se basa en la separación de la sangre de los líquidos que interconectan los tejidos, e incluso, desecando grandes áreas del cuerpo que quedaran completamente vacías (celoma). En el sistema circulatorio cerrado, la sangre no se encuentra regada por el celoma, si no contenida en un enorme órgano continuo que recibe el nombre colectivo de sistema circulatorio. El sistema circulatorio es continuo y forma un gran circuito. Las únicas zonas válidas para el intercambio de gases son los vasos capilares.

Figura 7.2. Sistema
circulatorio simplificado de los peces
De este modo emergerán dos fluidos, un
estático y que permanece alimentando y uniendo a los tejidos denominado linfa,
el cual extra muy relacionado con el sistema inmune. El segundo fluido es
altamente móvil, y fluye a través de todo el circuito circulatorio
distribuyendo materiales y gases metabólicos al cual denominamos sangre. El
intercambio de sustancias entre el circuito circulatorio y los tejidos se da en
zonas especializadas denominados vasos capilares, pero una vez los vasos se
hacen más grandes les es imposible llevar a cabo el intercambio de sustancias.
Más adelante veremos muchos más detalles de este sistema circulatorio ya que
este es el que poseen los vertebrados como los seres humanos.
7.3 Importancia de los corazones
Los sistemas circulatorios, abiertos o
cerrados, generalmente poseen mecanismos estructurales para el bombeo de la
sangre o la hemolinfa y así mantener los niveles de presión adecuados. Más allá
de la influencia del movimiento corporal, los corazones pueden clasificarse de
este modo. Vesículas contráctiles (anélidos), corazones ostiados (artrópodos),
y corazones de cámara (moluscos y vertebrados). Los corazones también se pueden
clasificar de acuerdo al mecanismo por medio del cual se inicia la contracción.
Los corazones miogénicos como los de los moluscos y los vertebrados poseen
mecanismos que inician la contracción desde el interior del órgano; contrario a
los corazones neurogénicos que inician la contracción por estructuras neuronales
exteriores al corazón.
La presión sanguínea y las velocidades de
flujo están íntimamente asociadas no solo a la actividad de bombeo mecánico
sino también a los diámetros de los vasos sanguíneos. Energéticamente hablando,
cuesta más mantener un flujo a través de una tubería delgada que por una
tubería grande. Este costo es minimizado en los animales manteniendo los vasos
sanguíneos más pequeños en lugares muy específicos dedicados únicamente para el
intercambio de gases, y dependiendo donde sea posible de vasos sanguíneos
grandes menos costosos energéticamente. Un detalle extra que explica la
arquitectura de los capilares es el siguiente, cuando el radio de un vaso
sanguíneo disminuye la presión aumenta y con ello la velocidad de movimiento de
la sangre, pero si el vaso sanguíneo se divide en varios, el volumen total de
todos los vasos sanguíneos pequeños es equiparable al de un vaso sangre, lo que
mantiene la presión y la velocidad de la sangre en niveles normales. Es por
esto que los capilares sanguíneos se ramifican pareciéndose a una maraña de
raíces interconectadas entre sí.
8. Esponjas y cnidarios
|| Regresar al Índice ||
Como ya se ha mencionado anteriormente, la
estructura física de las esponjas es muy simple, siendo una amalgama de células
pobremente especializadas sin generar verdaderos tejidos. Por lo anterior no
podemos hablar ni de órganos ni de sistemas en el sentido clásico de la
palabra. Sin embargo, la estructura corporal más compleja que poseen también
define la mayoría de sus funciones y se denomina el sistema acuífero.
Las células de la superficie externa de una esponja forman el pinacodermo y se llaman pinacocitos. La mayoría de las superficies internas comprenden el coanodermo y están compuestas de células flageladas llamadas coanocitos. Ambas capas son de una sola célula de espesor. Entre estas dos láminas celulares delgadas se encuentra el mesohilo, que puede ser muy delgado en algunas esponjas simples, o masivo y grueso en especies más grandes. El pinacodermo está perforado por pequeños orificios llamados poros dérmicos u ostia (singular, ostium), dependiendo de si la abertura está rodeada por varias células o una célula, respectivamente. El agua es arrastrada a través de estas aberturas y es conducida a través del coanodermo por los flagelos de los coanocitos. Los coanocitos bombean grandes volúmenes de agua a través del cuerpo de la esponja a presiones muy bajas, estableciendo el sistema de corriente de agua (sistema acuífero).
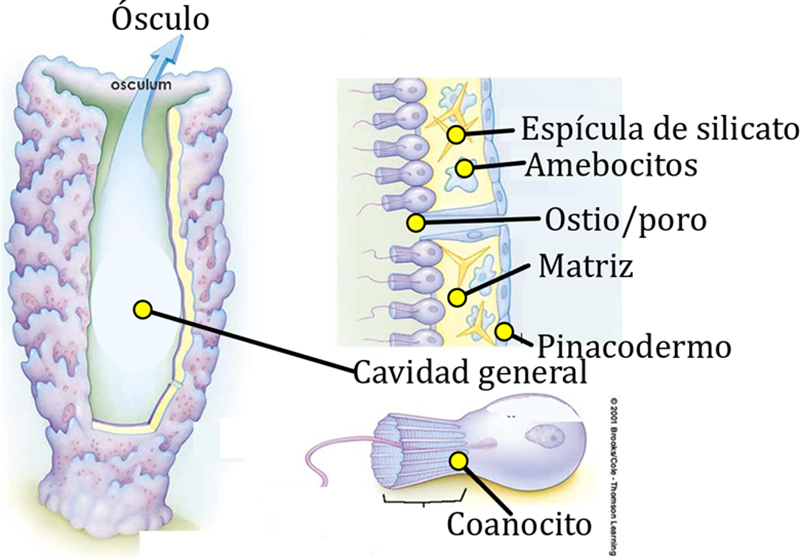
Figura 8.1. Los canales del
ostio en las esponjas generan un sistema acuífero que puede ser más complejo en
algunas especies, generando un laberinto por donde fluye el agua con gases
metabólicos, nutrientes y desechos.
Una cutícula, o capa de colágeno, puede
cubrir (o incluso reemplazar) el pinacodermo en algunas especies. El
pinacodermo en sí mismo puede ser una hoja externa simple, pero por lo general
también cubre algunas de las cavidades internas del sistema acuífero donde no
se producen coanocitos. Las células de pinacodermo que recubren los canales
internos se llaman endopinacocitos. El coanodermo también puede ser simple y
continuo, o plegado y subdividido de varias maneras creando un sistema acuífero
complejo con canales laberínticos. El mesohilo varía en grosor y desempeña
funciones vitales en la digestión, la producción de gametos, la secreción del
esqueleto y el transporte de nutrientes y productos de desecho por parte de
células ameboides especiales. El mesohilo incluye una mesoglea coloidal no
celular en la que están incrustadas fibras de colágeno, espículas y diversas
células; como tal, es realmente un tipo de mesénquima. Un gran número de tipos
de células se pueden encontrar en el mesohilo. La mayoría de estas celdas
pueden cambiar de un tipo a otro según sea necesario; pero algunos se
diferencian irreversiblemente, en células reproductivas o del esqueleto.
Durante el crecimiento, el pinacodermo y
el coanodermo son cada uno de una sola célula de espesor. Al aumentar su
plegamiento a medida que aumenta el volumen de mesohilo, estas capas mantienen
una relación de área de superficie a volumen suficiente para mantener un
intercambio adecuado de nutrientes y desechos en todo el individuo. El
coanodermo grueso de una célula puede seguir siendo simple y continuo (la
condición asconoide), o puede plegarse (la condición siconoide), o puede
subdividirse en gran medida en cámaras flageladas separadas (la condición
leuconoide).
8.1 Acuífero
asconoide
La condición de asconoide se encuentra en algunas esponjas calcáreas radiales simétricas de adultos (por ejemplo, Clathrina, Leucosolenia) y en la etapa de crecimiento temprano (Olynthus) de esponjas calcáreas recién asentadas. Las esponjas asconoides rara vez exceden los 10 cm de altura y permanecen como unidades tubulares simples en forma de jarrón.

Figura 8.2. Arquitectura de
una esponja asconoide.
Las paredes delgadas encierran una cavidad
central llamada atrio (= cavidad general de la esponja), que se abre hacia el
exterior a través de un solo osculum. El pinacodermo de las esponjas asconoides
y siconoides muy simples tiene células especializadas llamadas porocitos.
Durante la embriogenia, cada porocito se alarga y rueda para formar un tubo
cilíndrico. El porocito se extiende por todo el camino a través del
pinacodermo, el mesohilo delgado y el coanodermo hacia el atrio, emergiendo entre
los coanocitos adyacentes. La abertura externa del canal del porocito se llama
ostium o poro incurrente. El coanodermo es una capa simple y desplegada de
coanocitos que recubren todo el atrio. El agua que se mueve a través de una
esponja asconoide fluye a través de las siguientes estructuras: ostium → esponjoceloma
(sobre el coanodermo) → osculo.
8.2 Acuífero
sinconoide
El plegado simple del pinacodermo y el coanodermo produce la condición de siconoide, dentro de la cual son posibles varios niveles de complejidad. A medida que aumenta la complejidad, el mesohilo puede espesarse y parecer tener dos capas. La "región cortical" externa, o corteza, a menudo contiene elementos esqueléticos que son diferentes de los que se encuentran en la porción interior del mesohilo. En esas esponjas con una corteza, las aberturas incurrentes están revestidas por varias células (no formadas por un solo porocito) y se denominan poros dérmicos.

Figura 8.3. Arquitectura de
una esponja sinconoide.
de siconoide, los coanocitos se restringen
a cámaras específicas o divertículos de la aurícula llamados cámaras de
coanocitos (o cámaras flageladas o canales radiales). Cada cámara de coanocitos
se abre hacia el atrio por una gran abertura llamada apopilo. Las esponjas
siconoides con una corteza gruesa poseen un sistema de canales o canales
incurrentes que conducen desde los poros dérmicos a través del mesohilo hasta
las cámaras de los coanocitos. Las aberturas de estos canales a las cámaras de
los coanocitos se llaman prosopilos. En una esponja siconoide tan compleja, el
agua que se mueve desde la superficie hacia el cuerpo fluye a lo largo de la
siguiente ruta: poro incurrente (dérmico) → canal incurrente → prosopilo → cámara de
coanocitos → apopilo → atrio → osculo. La construcción de siconoide se encuentra en muchas
esponjas de calcáreas (por ejemplo, Scypha, también conocida como Sycon).
Algunas esponjas siconoides parecen radialmente simétricas, pero su
organización interna compleja es en gran parte asimétrica.
8.3 Acuífero
leuconoide
La condición leuconoide se produce mediante un plegamiento adicional del coanodermo y un mayor engrosamiento del mesohilo mediante el crecimiento cortical. Estas modificaciones están acompañadas por la subdivisión de las superficies flageladas en cámaras de coanocitos ovaladas discretas. En la condición leuconoide, se encuentra un aumento en el número y una disminución en el tamaño de las cámaras de los coanocitos, que generalmente se agrupan en grupos en el mesoilo engrosado. El atrio se reduce a una serie de canales excurrentes (o canales de exhalado) que transportan agua desde las cámaras de los coanocitos hasta el oscula. El flujo de agua a través de una esponja leuconoide es: poro dérmico → canal incurrente → prosopilo → cámara de coanocitos → apopilo → canal excurrente → osculum.

Figura 8.4. Arquitectura de
una esponja leuconoide.
La organización leuconoide es típica de la mayoría de las esponjas calcáreas y de
todos los miembros de la Demospongiae.
Es importante darse cuenta de que el caudal no es uniforme a través de las
diversas partes del sistema acuífero. Funcionalmente, es fundamental que el
agua se mueva muy lentamente sobre el coanodermo, lo que permite el tiempo para
el intercambio de nutrientes, gases y desechos entre el agua y los coanocitos.
Los cambios en la velocidad del flujo de agua a través de esta plomería son una
función de los diámetros efectivos acumulados de los canales, a través de los
cuales se mueve el agua. La velocidad del flujo de agua disminuye a medida que
aumenta el diámetro de la sección transversal; así, en una esponja, las
velocidades son más bajas sobre el coanodermo. Además, el agua que sale del
ósculo debe ser transportada lo suficientemente lejos para evitar que sea
reciclado por la esponja. En entornos de turbulencia, corrientes o acción de
las olas relativamente altas, este reciclaje potencial de desechos no es un
problema. Sin embargo, las esponjas que residen en aguas relativamente
tranquilas dependen del mantenimiento de altas velocidades de flujo de agua a
través de los ósculos (o en formas corporales modificadas) para empujar el agua
de desecho lo suficientemente lejos de la esponja para evitar su retorno. En
esponjas complejas, las diferencias en velocidad son dramáticas. La regulación
de la velocidad de flujo también se facilita en algunas esponjas, en parte, por
la actividad de las células ameboides (llamadas células centrales) que residen
cerca de las apoplas de las cámaras de los coanocitos. Estas células pueden
ralentizar o acelerar la salida del agua de las cámaras al cambiar de forma y
posición a través del apopilo.
El
reconocimiento de los diversos niveles de organización y complejidad entre los
poríferos permite describir rápida y simplemente el plan anatómico básico de
una esponja. Sin embargo, hay muy poca evidencia
de que el plan de ascoides sea necesariamente el más primitivo o que todos los
linajes de esponjas hayan pasado por estos tres niveles de complejidad durante
su evolución. Tampoco todas las esponjas pasan por estas tres etapas de
desarrollo. Además, las gradaciones de y los intermedios entre los tres planes
básicos son comunes. No obstante, entre las esponjas adultas, las
organizaciones más simples (asconoide y siconoide) ocurren solo en la clase Calcarea,
que se cree que es la clase más primitiva de poríferos vivos. Además, las
esponjas calcáreas de la condición leuconoide pasan a través de las etapas
asconoide y siconoide a medida que crecen, y solo en esta clase ocurren los
tres planes del cuerpo organizacional.
8.4 Cnidarios
Los cnidarios sufren de los mismos contratiempos que un gusano plano, pero a diferencia de este su cavidad interna o gastrodermo puede ser alimentado con un fluido circulante gracias al movimiento de su manto y tentáculos, nos referimos al agua.

Figura 8.5. Ya sean pólipos
o medusas, los cnidarios son lo bastante delgados y poco activos como para no
requerir un sistema circulatorio verdadero.
Los cnidarios pueden carecer de un sistema
circulatorio verdadero y depender de los mecanismos de transporte a través de
membrana para alimentar sus tejidos internos, mientras que la cavidad
gastrovascular dependerá del flujo de agua fresa desde la boca/ano del animal.
Debido a que los tejidos de los cnidarios son delgados y a que muchas veces son
muy pequeños, las únicas zonas que realmente requieren de un fluido circulante
es el gastrodermo, zona que es alimentada contantemente con agua fresca. El
resto del animal transporte sus nutrientes de un tejido a otro por mecanismos
de transporte a través de membrana similar a lo que ocurre en los gusanos
planos.
9. Platelmintos y nematodos
9.1 Platelmintos
En todos los animales, los fluidos intercelulares juegan un rol crítico como medio de transporte. Aun cuando existen sistemas complejos para la circulación de fluidos sanguíneos, el fluido tisular (linfa) sigue siendo necesario para transportar los materiales disueltos hasta las células.

Figura 9.1. Anatomía de una
planaria. Al ser animales pequeños, pueden realizar su transferencia de
sustancias no requiere conductos especializados, el sistema digestivo y
excretor se encargan de lo demás.
Los gusanos platelmintos son generalmente
muy pequeños (por algo son planos) por lo que los materiales, aunque no poseen
conductos especializados para ellos, pueden atravesar las capas celulares
mediante los mecanismos de transporte a través de membrana: transporte pasivo
(gases y excreción) y transporte activo (nutrientes). En algunos animales como
los gusanos planos no existen cámaras especializadas para los fluidos del
cuerpo más allá que los espacios intercelulares, donde los materiales deben
atravesar célula a célula hasta llegar a donde son necesarios en lugar de ser
canalizados por una tubería.
Los gusanos planos alcanzan sus máximos
tamaños en ambientes ricos en nutrientes como el agua o "en el caso de los
parásitos" en ambientes ricos en nutrientes fácilmente absorbibles como el
intestino delgado o el sistema circulatorio de otros animales. Esto hace que el
transporte de materiales sea lento y costoso energéticamente hablando, lo que
mantiene a los gusanos planos a tamaños muy pequeños, donde la cantidad de
capas celulares es lo suficientemente pequeña como para que los materiales
atraviesen dichas capas sin quedar agotados.
9.2 Nematodos
Los animales blastocelomados (cavidad interna hueca que diferente de la cavidad gastrointestinal y de las cavidades propias de los órganos) utilizan el fluido del pseudoceloma llamado hemolinfa como un órgano circulatorio, básicamente lo podemos definir como un sistema circulatorio abierto, aunque carece de corazones.

Figura 9.2. El líquido que
rellena el pseudoceloma se encarga de las funciones de transporte interno.
La propulsión en este caso es ocasionada
por los movimientos propios del cuerpo del animal. En los blastocelomados como
los gusanos redondos y los rotíferos a parte del mero fluido del pseudoceloma
se encuentras células diluidas especializadas para el transporte de sustancias,
aunque sus funciones específicas aún no han sido propiamente estudiadas.
10. Anélidos 1, Poliquetos
|| Regresar al Índice ||
Los gusanos segmentados o anélidos pueden
poseer tanto sistemas circulatorios abiertos como cerrados, aunque lo ms común
es que sean cerrados. En las especies que no poseen tabiques que separen los segmentos
generalmente poseen un sistema circulatorio cerrado en el celoma, el cual
contiene células disueltas llamadas celomocitos. Sin embargo, debido a la
diversidad estructural de estos gusanos, es conveniente dividir la discusión de
su sistema circulatorio en sus grupos más importantes. El sistema circulatorio
basal de los anélidos es un sistema circulatorio cerrado análogo al de los
vertebrados, aunque sin corazones no se puede aplicar el concepto de vena o
arteria y en su lugar simplemente llamaremos a sus conductos como vasos
sanguíneos.
Tendemos a pensar en los gusanos como un
conjunto homogéneo, sin embargo, los anélidos se caracterizan por tener tamaños
mayores, y ser más complejos estructuralmente, y dicha complejidad se encuentra
asociada a un sistema circulatorio más eficiente. La eficiencia del sistema
circulatorio se debe al problema clásico de la superficie, el volumen y la
especialización, solo algunas partes del cuerpo obtienen nutrientes, ya sea de
los alimentos o de los gases atmosféricos, por lo que dichos nutrientes deben
viajar desde donde son obtenidos hacia donde son requeridos.
La forma más sencilla de entender el sistema circulatorio de los poliquetos es estudiarlo en conjunción con el sistema respiratorio, así que es probable que repita algunas cosas que ya se han mencionado en el respectivo sistema. El sistema circulatorio de los poliquetos es principalmente cerrado o altamente vascularizado. Estos vasos sanguíneos se organizan de manera semejante a los vertebrados, con canales gruesos para transporte pesado y capilares con paredes de una célula en los que se realiza el intercambio de sustancias.

Figura 10.1. Modelo del
sistema circulatorio alrededor de un segmento y su queto en Nereis.
Estos capilares pueden distribuirse en
toda la piel en aquellas especies que respiran cutáneamente o agruparse
densamente en los quetos o filamentos que funcionan como branquias. También de
forma análoga a los vertebrados, los poliquetos han desarrollado pigmentos que
transportan de forma más eficiente los gases de importancia metabólica.
En el modelo anterior podemos ver la
estructura de intercambio de gases de Nereis
El movimiento de la sangre a través del
sistema circulatorio en Nereis depende principalmente de la acción
muscular de las paredes del animal, así como de capas de musculo que rodean los
vasos sanguíneos. No existen corazones u órganos analogables pulsantes. La
sangre oxigenada de las branquias retorna hacia el vaso sanguíneo dorsal que se
dirige a la cabeza, de modo tal que la sangre más “fresca” de gases nutritivos
pasa primero por el sistema digestivo y por el ganglio cerebral.
10.1 Variación
en el sistema circulatorio de los poliquetos
Existen muchas variaciones a este esquema
básico de sistema circulatorio cerrado, y solo mencionaremos unos pocos para
mostrar la diversidad de estos seres vivos. Algunos poliquetos pueden presentar
un sistema circulatorio cerrado reducido o perdido, retornando a un sistema
circulatorio abierto. En algunos casos como en el género Syllidae spp. la
reducción del sistema circulatorio cerrado puede ser causada probablemente por
una reducción en el tamaño máximo del adulto.
Los gliceridos por el contrario son
grandes y han perdido su sistema circulatorio cerrado abriéndolo al celoma, por
lo que son animales de sistema circulatorio abierto. Estos animales poseen
células especializadas para el transporte de oxígeno, con una proteína de
transporte muy común, la hemoglobina, por lo que su hemolinfa es de color rojo.
Cada segmento de los glicéridos está imperfectamente separado, lo cual le
permite a la hemolinfa fluir por todo el animal, mientras que la función de
intercambio de gases es realizada por la piel.
La pérdida del sistema circulatorio no es
la única tendencia evolutiva, otros géneros han experimentado una ruta opuesta,
y en lugar de simplificar o perder el sistema circulatorio cerrado lo que han
hecho es hacerlo más complejo. En los terebelidos, sabelidos y serpilidos el
vaso sanguíneo dorsal se encuentra reemplazado por un espacio voluminoso lleno
de sangre denominado seno viceral. Usualmente el vaso dorsal continua
anteriormente formando en la cola un anillo que la conecta con el vaso ventral.
En los serpulidos los vasos que forman el anillo están equipados con válvulas
que evitan el flujo reverso de sangre de regreso al vaso sanguíneo dorsal.
10.2 Corazones
Algunas estructuras pulsantes han
evolucionado en algunas especies de poliquetos. Estas son especialmente bien
desarrolladas en especies sésiles que se encuentran protegidas por tubos duros,
y que exponen sus filamentos para respirar o cazar. Debido a que no se mueven
deben desarrollar estructuras accesorias que mantengan la sangre en
movimiento. Estas estructuras son
denominadas por analogía como corazones, aunque este título no tiene relación
filogenética con los corazones de los vertebrados.
Los corazones de los poliquetos son poco
más que porciones con una mayor capa muscular y un poco más grandes que el
resto del conducto sanguíneo. En los quetopteridos el vaso dorsal cumple esta
función. Los terebelidos poseen una estación de bombeo en la base de las
branquias que sirve para mantener la presión sanguínea al interior de los
capilares de las branquias. La posición y forma de estos corazones es variable
entre los diferentes linajes de poliquetos.
10.3 Pigmentos
respiratorios
Los pigmentos respiratorios son moléculas
con color que se enlazan a los gases de importancia metabólica, dioxígeno para
la forma nutritiva y dióxido de carbono para la forma tóxica. En general la
mayoría de los pigmentos respiratorios de los poliquetos son variaciones de la
hemoglobina, aunque la clorocruorina es común en algunas familias de sabelidos
y serpulidos, así como la hemeritrina en los magelonidos. Algunos poliquetos
pueden tener más de un pigmento como en algunos serpulidos que mezclan la
hemoglobina con la clorocruorina.
Los pigmentos pueden encontrarse disueltos
directamente en la sangre, en el fluido celómico, en ambos o encerrado en
células especializadas para evitar problemas osmóticos provocados por tener
demasiadas proteínas disueltas en la sangre. Los tipos de pigmentos reflejan
los tipos de estilo de vida de los gusanos que los poseen, por ejemplo algunos
gusanos que viven en zonas de intermareas experimentan condiciones cambiantes
de solubilidad de oxígeno, por lo que tener pigmentos diferentes para
condiciones diferentes es una ventaja en ese tipo de ecosistemas en particular.
11. Anélidos 2, Oligoquetos e hidrudineos
|| Regresar al Índice ||
El sistema circulatorio de los
oligoquetos, cuya especie tipo es la lombriz de tierra, es semejante al de los
poliquetos, siendo fundamentalmente un sistema circulatorio cerrado con
presencia de pigmentos respiratorios. Las diferencias entre los sistemas
respiratorios de los oligoquetos con respecto a los poliquetos se fundamentan
esencialmente en el hecho de que tratamos ahora con animales que habitan la
tierra seca o ambientes de intermareas tendientes a tener una menor
disponibilidad de agua.
En la lombriz de tierra el sistema circulatorio es cerrado y consiste inicialmente en tres vasos sanguíneos principales que discurren longitudinalmente desde la cabeza hasta la cola, generando segmentos como anillos precisamente en cada uno de los segmentos del animal, después de todo el nombre común de los anélidos es gusanos segmentados. El más grande de estos tres conductos es el vaso sanguíneo dorsal, que al igual que los semicorazones en algunos poliquetos, presenta una pared muscular más desarrollada, por lo que genera gran parte de la presión sanguínea que mantiene la sangre de los anélidos en movimiento. Suspendida en el mesenterio justo por debajo de las vísceras se encuentra el vaso sanguíneo ventral. El tercero de estos cabales se encuentra por debajo de la médula nerviosa y es denominado vaso sanguíneo subneural.

Figura 11.1. Sistema
circulatorio de la lombriz de tierra.
A partir de los vasos segmentales que
forman los anillos se proyectan vasos menores hacia la pared corporal o hacia
las vísceras, los cuales forman el sistema capilar de alimentación.
Generalmente el intercambio de sangre entre los vaso sanguíneos dorsales y
ventrales ocurre de forma independiente en cada segmento. La mayoría de los
oligoquetos también poseen de dos a cinco pares de vasos sanguíneos grandes,
musculares que rodean la faringe para el transporte de sangre desde la región
dorsal a la región ventral. Estos vasos sanguíneos ayudan a propeler la sangre
actuando como “corazones” secundarios manteniendo la presión sanguínea. Al
igual que el vaso sanguíneo principal dorsal, estos vasos faríngeos están
equipados con válvulas para evitar el flujo recurso de la sangre al ejecutar la
contracción del vaso sanguíneo.
La mayoría de los oligoquetos poseen
hemoglobina disuelta directamente en el plasma; los miembros de las familias
Naididae carecen de hemoglobina. Estos organismos presentan formas
rudimentarias de un sistema inmune asociado al sistema circulatorio en forma de
células amiboideas que patrullan los fluidos circulatorios.
En las sanguijuelas o hirudineos el
espacio celómico se ve reducido por una invasión de tejido del cual puede
derivar un sistema circulatorio secundario con vasos sanguíneos y lagunas de
almacenamiento relativamente complejas en comparación del sistema circulatorio
original heredado del ancestro anélido. Incluso en algunos linajes el sistema
circulatorio del tejido celómico ha reemplazado parcial o totalmente al sistema
cerrado original.
12. Moluscos
|| Regresar al Índice ||
A pesar de que los moluscos son
protostomos celomados, su celoma se encuentra fuertemente reducido. La principal
cámara corporal corresponde a un espacio lleno de líquido de transporte, por lo
que recibe el nombre de hemoceloma y en consecuencia nos encontramos ante un
sistema circulatorio abierto. La cavidad celómicas recibe en consecuencia el
nombre de hemoceloma. El hemoceloma puede proyectarse en senos e inclusive en
una compleja red de vasos que asemejan a un sistema circulatorio cerrado. El
grado de semejanza con un sistema circulatorio cerrado varía mucho, pudiéndose
presentar corazones.

Figura 12.1. Los moluscos como los
cefalópodos poseen un sistema circulatorio mixto, con regiones cerradas y
abiertas.
La hemolinfa de los moluscos contiene
varias células especializadas, incluyendo amebocitos con parte de un sistema
inmune rudimentario. La hemolinfa de los moluscos también posee un pigmento
respiratorio, pero no es la hemoglobina basada en el hierro a la que estamos
acostumbrados, por el contrario, se trata de una molécula basada en el cobre
denomninada hemocianina. Muchos moluscos pueden emplear hemoglobina o
mioglobina.
Por lo general se presenta un corazón en
posición dorsal al interior de una cámara pericardíaca, y se estructura con un
par de atrios (más comúnmente denominados aurículas en el idioma español) y un solo
ventrículo. En los monoplacóforos y los nautilos hay dos pares de ventrículos
independientes. Los ventrículos reciben la hemolinfa oxigenada desde las
branquias (denominadas ctenidios) o desde la aorta cefálica. La arteria
anterior se ramifica y eventualmente se abre en varios senos al interior de los
tejidos que son bañados en la hemolinfa oxigenada. Esto significa que el
intercambio de sustancias entre el MEC tisular y la hemolinfa se da casi
directamente sin la intervención de un sistema de capilares. Los senos
sanguíneos simplemente abren a los tejidos, y luego drenan para llegar a las
venas retornantes al corazón.
En el esquema del sistema circulatorio de
los moluscos mostrando en la Figura 12.1 se
puede observar que una vez que los vasos sanguíneos principales llegan a un
sistema de tejidos para el intercambio de sustancias, en lugar de ramificarse
en un sistema de capilares, simplemente se decanta en una cavidad hemocelómica
independiente, que baña el tejido que va a ser alimentado y luego es drenado
por la siguiente parte del sistema circulatorio. En este sentido podemos
afirmar que el sistema circulatorio de los moluscos, aunque se denomine
abierto, en realidad es su propio tipo intermedio entre el sistema abierto y el
sistema cerrado. Los moluscos pueden tener incluso una tendencia al cerramiento
con la aparición de sistemas capilares como ocurre en los cefalópodos.
Los cefalópodos dependen de una alta
presión sanguínea para mantener sus activos modos de vida, y, en consecuencia,
a parte del cerramiento secundario de sus sistemas circulatorios, también
experimentaban la aparición de corazones auxiliares secundarios denominados
corazones branquiales, que incrementan la presión sanguínea de la hemolinfa que
regresa a los ctenidios “branquias”.
13. Artrópodos
|| Regresar al Índice ||
Los artrópodos poseen un sistema
circulatorio abierto. Sin embargo, de manera semejante a lo que ocurre con los
moluscos, los vasos sanguíneos de los artrópodos pueden variar de tamaño, desde
muy pequeños, hasta cubrir la casi totalidad del cuerpo del artrópodo. Las
funciones del sistema circulatorio también pueden variar grandemente, por
ejemplo, en los quelicerados y los crustáceos, la sangre transporta el oxígeno
ya que sus mecanismos de obtención de aire son las branquias o el sistema de
pulmones en libro. Los insectos por su parte no transportan gases disueltos por
la sangre, en lugar de ello, los tejidos realizan intercambio de gases
directamente en el aparato respiratorio sin mediación del circulatorio
Los artrópodos terrestres no arácnidos
poseen un sistema respiratorio de tráqueas corporales que permite el
intercambio de gases virtualmente en cualquier parte del cuerpo, lo que le
permite a la hemolinfa refrescarse con oxígeno por todas partes o como
mencionamos anteriormente, los tejidos pueden recibir el oxígeno casi
directamente a través de superficies húmedas.
13.1 Estructura
del vaso principal dorsal
El sistema circulatorio de los artrópodos
es abierto y consta de un sistema de vasos que no se cierra, si no que se abren
y depositan el fluido hemolinfa en la cavidad general del animal denominado
hemoceloma. La entrada a la arteria es en la parte trasera del animal y en una
serie de orificios llamados ostium dispuestos a lo largo de esta arteria
principal, para luego salir por la parte delantera. La arteria principal
también es el corazón cubierto en consecuencia con varias capas musculares.
Generalmente la aorta termina en uno o más sacos contráctiles denominados sacos
frontales aórticos que actúan como más corazones accesorios. Estos sacos ayudan
a impulsar la sangre a través de canales arteriales que van hacia las antenas.
Por lo general los canales arteriales no se capilarizan, sino que se encuentran
abiertos expeliendo la hemolinfa a un saco llamado seno que es una delgada
proyección del hemoceloma, de forma tal que la hemolinfa rellena el centro de
la antena.
13.2 Diafragmas
En el abdomen se encuentra una hoja muscular denominado tabique dorsal, septo dorsal o diafragma dorsal, que se encuentra sosteniendo al vaso sanguíneo principal. Esta placa forma un espacio abierto que divide parcialmente el hemoceloma en la cavidad principal ventral y una cavidad dorsal denominada seno pericárdico. Adicionalmente también puede presentarse un tabique ventral muscular que separa parcialmente la posición de la médula nerviosa más ventralmente del hemoceloma. De esta forma el hemoceloma de los insectos está dividido en tres zonas pero aun así la hemolinfa fluye por los tres, siendo estos el seno pericárdico en el dorso, el seno visceral en el centro y el seno perineural en el vientre. Estos senos pulsan, pero cumplen diversas funciones. La primera es ayudar a dar más movimiento en la hemolinfa en los tres senos, y la otra es anclar músculos de patas y alas.

Figura 13.1. Modelo del
sistema circulatorio de un insecto.
13.3 Patrón de
circulación
El patrón básico de flujo de la hemolinfa
es el siguiente: la hemolinfa fluye a través del corazón dorsal, especialmente
en la parte del abdomen donde es más fuerte la capa muscular, siendo propelida
hacia la aorta. El flujo reverso de hemolinfa es impedido debido a que los
ostios se cierran. La sangre sale por varios orificios de la aorta cerca de la
cabeza o incluso en las antenas, y la presión hemolínfática hace que esta sea
impulsada hacia atrás rellenando las tres cavidades en que se divide el
hemoceloma: seno pericárdico, seno visceral y seno perineural. La hemolinfa que
se desvía hacia el seno pericárdico ingresa por los ostios al corazón, mientras
que los tabiques abdominales evitan que la hemolinfa se estanque en la zona
ventral y visceral.
13.4 Contacto
con el sistema respiratorio
El sistema de tráqueas penetra en todos
los senos e incluso en antenas, patas y alas, por lo que la hemolinfa siempre
se encuentra intercambiando gases metabólicos. De allí que no deban pasar por
“lugares especiales” para que esto ocurra. Ahora bien, el problema emerge
cuando el artrópodo no puede intercambiar gases metabólicos de importancia por
todas partes, sino que se encuentra limitado a órganos concretos de intercambio
de gases como las branquias o los pulmones en libro.
13.5 Las branquias, pulmones en libro y alas
En los crustáceos con branquias el sistema circulatorio de impulso sanguíneo ubicado dorsalmente se encuentra reflejado por un sistema de vasos sanguíneos de disposición ventral que se encargan de retomar la hemolinfa baja en oxígeno. Estos vasos sanguíneos conducen la hemolinfa desde el hemoceloma hacia las branquias, donde la hemolinfa se refresca con oxígeno y de allí es dirigida a los vasos sanguíneos ventrales, siendo una organización semejante a la que ocurre en los moluscos, aunque claro está, con un hemoceloma más grande.

Figura 13.2. Modelo del sistema circulatorio de un crustáceo.

Figura 13.3. Modelo del ala
de un insecto.
Los capilares branquiales también nos revelan una pista de la evolución de las alas. Ya hemos discutido que las alas de los insectos evolucionaron de las branquias de los artrópodos (Clark-Hachtel & Tomoyasu, 2016; Haug, Haug, & Garwood, 2014; Pass, Tögel, Krenn, & Paululat, 2015; Whitfield, 2016), por lo que no sorprende que las alas y las branquias son de las pocas partes del cuerpo de un artrópodo que es alimentado por medio de un sistema de vasos sanguíneos capilares en lugar de ser alimentadas directamente por el hemoceloma. Estos vasos sanguíneos son de cierta forma formados por el propio hemocelóma, pero al ser estrechos y canalizados los podemos clasificar como vasos sanguíneos, aunque separados de la arteria principal. En la base del hemoceloma que conecta a las venas de las alas se encuentran un par de órganos pulsantes que funcionan como corazones accesorios denomiandos corazones mesotoracicos.

Figura 13.4. Modelo del
sistema circulatorio de una araña.
Los pulmones en libro de los arácnidos
también plantean ciertos problemas para la organización del sistema circulatorio.
Los arácnidos con sistema de pulmones en libro tienen una organización
particular, la sangre oxigenada fluye por medio del vaso sanguíneo dorsal
típico de los artrópodos, sin embargo, los ostios de la arteria dorsal abren a
un seno hemocelómico especializado que encierra hemolinfa que proviene desde
los pulmones en libro. Este seno hemocelómico especializado y separado del
resto del hemoceloma es denominado vena pulmonar.
Los arácnidos también presentan otros
sistemas accesorios, por ejemplo, poseen capilares o precapilares que
transportan la hemolinfa a la pared corporal, y además poseen también un
sistema de tráqueas que apoya a los pulmones en libro. Algunos quelicerados
también presentan hemocianina como pigmento respiratorio.
14. Algunos deuterostomos
|| Regresar al Índice ||
14.1 Equinodermos
El transporte interno de sustancias en los equinodermos es realizado principalmente en el celoma peri-visceral que es el espacio extra al interior del cuerpo, el sistema vascular de agua conocido como pie de agua y por el sistema hemático. Todos los sistemas se derivan del celoma. Los fluidos se mueven a través de estos sistemas principalmente por la acción de cilios y en algunos casos por presión muscular.

Figura 14.1. Sistema
acuífero de la estrella de mar.
El sistema hemático es un complejo de
canales y espacios encerrados principalmente por canales celómicos denominados
senos perihemáticos. El sistema se encuentra mejor desarrollado en los
holoturios “pepinos de mar” y en los crinoides, en los cuales los canales
forman un flexo semejante a una red. En otros grupos el sistema se organiza
radialmente y generalmente discurre en paralelo a los elementos del sistema de
flujo de agua. En estos casos el sistema hemático consiste en un anillo
hemático oral y aboral cada uno con extensiones radiales. Los dos anillos están
conectados por el seno axial.
Al interior del seno axial existe un
núcleo de tejido esponjoso denominado glándula axial. La glándula axial
aparentemente es responsable de producir algunos celomocitos. El seno axial se
encuentra abierto a través de poros hacia el canal rocoso siendo la fuente de
fluidos del sistema de transporte de agua. Los canales hemáticos radiales del
anillo aboral se extienden hasta las gónadas. Otros canales radiales aparecen
desde el anillo radial y se encuentran asociados con las hileras de los pies.
Estos canales se almacenan al interior de un espacio perihemático denominado
seno hiponeural. Un tercer anillo hemático denominado anillo gástrico se
encuentra en muchos equinodermos incluyendo a las estrellas de mar y se
encuentra asociado con el sistema digestivo.
El sistema hemático obtiene el movimiento
de la sangre a través de cilios. En las estrellas de mar y erizos de mar se
encuentra un seno axial extra reforzado con tejido muscular denominado saco
dorsal, el cual se encuentra cerca de la unión con el anillo hemático aboral.
El saco dorsal palpita, aparentemente apoyando el movimiento de fluido
sanguíneo a través de los canales y senos hemáticos. La fisiología del sistema
hemático aún no se encuentra del todo comprendida, pero probablemente ayuda a
distribuir nutrientes absorbidos desde el sistema digestivo.
Lamentablemente los modelos del sistema
circulatorio de las estrellas de mar no son tan intuitivos como en otros
organismos, ya sea porque al encontrarse tan asociados al sistema digestivo es difícil
distinguir uno del otro, o que del mismo modo se encuentran extremadamente
asociados al sistema de transporte de agua (imagen anterior), el cual se
encarga de dar presión hidráulica a los podios, especialmente en las estrellas
de mar. Por tal razón, especialmente para las estrellas de mar el sistema de
transporte de agua puede ser una representación aceptable del sistema
circulatorio.
14.2 Tunicados
Los tunicados como los demás cordados poseen un notocordio al inicio de su desarrollo biológico, pero a diferencia de los vertebrados, en los cuales el notocordio se vuelve un sistema de vertebras cerrados, en los tunicados esta estructura se pierde paulatinamente.

Figura 14.2. Corazón "si es que puede recibir ese nombre" de los tunicados, es tubular.

Figura 14.3. Circuitos circulatorios de
varios animales.
Los tunicados son animales muy
interesantes en términos de la biología del desarrollo y la teoría evolutiva
debido a los grandes cambios que experimentan, las larvas son móviles y se
asemejan a un cefalocordado, mientras que los adultos son animales fijos que se
alimentan por filtración. Aunque al inicio de su desarrollo poseen un celoma
este también se pierde durante su desarrollo. Su corazón es tubular y con una
sola cámara, no posee válvulas y si presión es causada por movimientos
peristálticos típicos de un sistema con tejido muscular liso. No posee
endocardio y su epicardio puede llegar a envolver a otros órganos.
Los tunicados poseen un sistema
circulatorio abierto, aunque con una gran cantidad de vasos sanguíneos que se
extienden casi por todo su cuerpo. La sangre o hemolinfa es interesante debido
a una gran cantidad de características. Por ejemplo, la sangre no posee
hemoglobina a pesar de que su genoma contiene cerca de 4 genes para ella.
También poseen el gen de la hemocianina, pero esta proteína no es usada para el
transporte de oxígeno en este tipo de animales.
En la Figura 14.3 se
pueden ver los circuitos circulatorios de varios animales, de arriba hacia
abajo, el primero es el circuito circulatorio de los tunicados el cual es
abierto, pero como se muestra en la figura, los vasos sanguíneos se proyectan
casi hasta completarlo de manera cerrada. Los siguientes dos intermedios son
circuitos circulatorios de cefalocordados, mientras que el inferior es el
circuito circulatorio de un vertebrado con un corazón verdaderamente
diferenciable. Otro detalle interesante de la sangre o hemolinfa de los
tunicados es su tendencia a contener altas concentraciones de metales pesados
de transición como el vanadio, junto con proteínas asociadas a este que lo
hacen soluble. Otro metal presente en altas concentraciones es el litio. La
concentración de vanadio en un tunicado puede ser 1000 veces mayor a la
presente en el agua que lo rodea. Por lo general el vanadio será depositado en
la túnica, posiblemente para hacerla más dura.
14.3 Cefalocordados
El sistema circulatorio de los cefalocordados es extremadamente similar al de los vertebrados terrestres, y no es de extrañar, ya que estos animales son lo más cercano fisiológicamente a lo que se esperaría que fuera el ancestro común de todos los vertebrados y los cordados.

Figura 14.4. Modelo del circuito circulatorio
de un cefalocordado típico (anfioxus).
A pesar de lo que se muestra en la Figura 14.4, las
branquias en sí mismas no son las que se encargan del intercambio de gases, si
no, la piel externa. Algunos autores no reconocen al vaso sanguíneo ensanchado
como un corazón real, aunque si lo fuese seguiría siendo más simple que el de
los peces al ser unicameral. La sangre no posee células sanguíneas y tampoco
expresa la hemoglobina a pesar de tener los genes para ello.
A pesar de que se señala como un corazón,
el vaso sanguíneo que propulsa la sangre es en sí mismo un mero vaso sanguíneo
ensanchado con una sola cámara, es tan simple que algunos autores ni siquiera
lo reconocen como tal, señalándolo como un mero vaso sanguíneo más ancho y
contráctil.
15. Venas, arterias y capilares en los vertebrados
|| Regresar al Índice ||
El sistema circulatorio de los vertebrados es básicamente un sistema de tubos interconectados entre sí, sin aberturas de ningún tipo a la cavidad general o celoma. En el interior de los tubos se encuentra un fluido circulante denominado sangre. El sistema circulatorio de los vertebrados se lo denomina como cerrado, debido a la carencia de aberturas a la cavidad general o a los tejidos, de este modo los fluidos se separan completamente, uno para los tejidos que los mantienen alimentados, y la sangre en sí que nunca sale del sistema circulatorio.

Figura 15.1. Los vertebrados
poseen un sistema circulatorio cerrado.
Los vertebrados siempre presentan un
corazón, aunque rudimentos evolutivos de este están presentes en los parientes
más cercanos de los vertebrados, los cefalocordados. La función del corazón es
dar impulso a la sangre, aunque los vasos sanguíneos pueden contraerse y
dilatarse de modo tal que pueden impulsar de manera secundaria a la sangre a
todo lo largo de su recorrido. El sistema circulatorio es integrante, es decir,
es un sistema que permite el intercambio de sustancias entre todos los demás
sistemas del cuerpo y, por lo tanto, su función primordial es el transporte de
sustancias. En este sentido es primordial para completar las funciones de
digestión y respiración organísmica, así como la de excreción e incluso la
función inmune.
15.1 Partes del sistema circulatorio
A pesar de la variación de tamaño, existen
tres tipos principales de vasos sanguíneos: las arterias, las venas y los
capilares.
La definición de vena y arteria dependerá del corazón y no del nivel de oxigenación de la sangre, si la sangre deja el corazón, cualquier vaso que transporta sangre que sale del corazón recibe el nombre de arteria independientemente de que esa sangre esté desoxigenada u oxigenada, de hecho la característica relevante es que la sangre lleva una alta presión; del mismo modo todo vaso sanguíneo que transporta sangre que ingresa al corazón recibirá el nombre de vena independientemente de si transporta sangre oxigenada u desoxigenada, siendo la característica importante una baja presión, aunque la presión es mucho más baja en la vena sistémica “Cava” que en la vena pulmonar.

Figura 15.2. Diagrama
esquemático del sistema circulatorio de un vertebrado pulmonado, con los
principales vasos sanguíneos.
15.2 Venas y
arterias
Sin embargo, lo anterior solo es cierto para el circuito circulatorio mayor que lleva la sangre a los capilares sistémicos. Por convención, la referencia entre la nomenclatura de arteria y vena es el corazón, la sangre que deja el corazón le dará el nombre de arteria al vaso que la transporta; mientras que la sangre que ingresa al corazón le dará el nombre de vena al vaso de la transporta. De este modo, las arterias del circuito circulatorio menor “en los vertebrados con respiración pulmonar” llevan sangre desoxigenada mientras que las venas del circuito circulatorio menor llevan sangre oxigenada al corazón respectivamente. Los vasos sanguíneos y los diferentes nombres que reciben, en este esquema la coloración y los nombres se hacen en referencia al circuito sistémico, donde las arterias son rojas y las venas azules; puesto que en el circuito pulmonar la coloración se invierte.

Figura 15.3. Las venas y
arterias sistémicas se unen en capilares sistémicos que dejan pasar materiales
a los tejidos por los mecanismos de transporte a través de membrana.
Las arterias y las venas poseen paredes
tubulares organizadas en tres capas que encierran al epitelio interno. Estas
capas reciben el nombre de túnica íntima, túnica media y túnica adventicia. La
túnica intima se encuentra en contacto con el epitelio interno o lumen del vaso
sanguíneo. La túnica adventicia es la parte externa de los vasos sanguíneos más
grandes, y la túnica media se encuentra como su nombre lo dice se encuentra en
medio de las dos. La túnica media en ocasiones contiene tejido muscular liso,
el cual le otorga una capacidad contráctil al vaso (vasodilatación y
vasoconstricción), en otras palabras, pueden generar movimientos peristálticos
lo que le otorga un impulso extra a la sangre, independiente al impulso inicial
del corazón. Las venas tienen por lo general válvulas de un solo sentido que
impide que la sangre que contiene gases tóxicos disueltos regrese, mientras
que, las arterias no poseen estas válvulas.
15.3 Estructura
de las venas y las arterias
En el modelo anterior podemos apreciar la variación del grosor de las paredes de los vasos sanguíneos a medida que se van transformando en capilares. La pared de los capilares es la más delgada, consistiendo tan solo en una capa de células epiteliales. A medida que los vasos sanguíneos se acercan al contacto íntimo con los tejidos, su radio vascular disminuye, así como el grosor de su pared, tomando el nombre de arteriolas y vénulas respectivamente. En los vasos muy pequeños, la túnica adventicia es muy delgada, mientras que la túnica media está compuesta casi exclusivamente por musculo liso; por lo que las vénulas y las arteriolas son muy similares en términos de estructura.

Figura 15.4. El grosor de las venas y arterias va disminuyendo capa a capa hasta llegar a ser capilares, donde ambos se unen.

Figura 15.5. La sangre es un
tejido móvil que transporta materiales como lipoproteínas que transportan
grasas y células como los glóbulos rojos o blancos, partes de células como las
plaquetas y proteínas solubles.
Una vez que las paredes se adelgazan al
máximo, los gases, nutrientes, desechos, iones y el calor pueden fluir a través
del epitelio de los capilares. Para facilitar este paso de sustancias, los
epitelios de los capilares son muy delgados, alrededor de una célula de grosor
y no más, lo que implica que la mayoría de las sustancias deben atravesar tan
solo dos membranas.
15.4 Arterias
Las arterias son todos aquellos vasos
sanguíneos que transporta sangre que deja el corazón sin importar si esta es
sangre oxigenada o desoxigenada. La estructura de las arterias varía con el
tamaño. Las arterias más grandes poseen una considerable cantidad de fibras
elásticas en sus paredes, mientras que las arterias más pequeñas carecen de
estas fibras elásticas. Las diferencias estructurales se dan debido a la
diferencia funcional entre las arterias grandes y las arterias pequeñas. La
función de las arterias, como ya se recalcó en la definición, es la de
transportar la sangre que deja el corazón a los diferentes tejidos, lo que
implica que sus paredes deben absorber los pulsantes incrementos de presión
cuando el corazón late, y esa fuerza debe ser soportada, absorbida y canalizada
por la pared de la arteria.
Las fibras elásticas les permiten a estas
arterias soportar estos incrementos rítmicos de presión sin romperse o
explotar, y de hecho, les permite transmitir la energía que transporta la
sangre a todo lo largo del cuerpo. Esta energía puede sentirse en las arterias
del cuello o de la muñeca izquierda al tocarlas con una presión adecuada y es
lo que se conoce como pulso. Esta transmisión de energía a través de las fibras
elásticas le permite al corazón usar menos energía, para hacer circular la
sangre, lo que en últimas mantiene una presión arterial relativamente baja, lo
cual es muy importante debido a que, las arterias pequeñas carecen de fibras
elásticas. Si las arterias grandes no transmitieran la energía, el corazón
debería latir con más fuerza para impulsar la sangre, lo que implicaría que la
sangre que llega a las arteriolas lo haría con mucha presión. Al carecer de
fibras elásticas las arteriolas se pueden romper o explotar causando una serie
de enfermedades relacionadas con el rompimiento de vasos sanguíneos como un
derrame cerebral.
El taponamiento arterial, causado por
placas de sangre coagulada, o por grasa acumulada en el epitelio interno de la
arteria incrementa la presión sanguínea lo que puede provocar rompimientos de
los capilares sanguíneos a los cuales esa arteria alimenta. Los rompimientos de
los vasos sanguíneos capilares ocurrirán en cualquier lugar donde existan
capilares sanguíneos, lo cual se traduce en todo el cuerpo, sin embargo, el
rompimiento en el cerebro es particularmente peligroso debido a la gran
sensibilidad del cerebro al incremento de la presión y a la carencia de
nutrientes.
15.5 La arterioesclerosis
Es un término general utilizado en medicina humana y veterinaria, que se refiere a un endurecimiento de arterias de mediano y gran calibre. La arteriosclerosis por lo general causa estrechamiento (estenosis) de las arterias que puede progresar hasta la oclusión del vaso impidiendo el flujo de la sangre por la arteria así afectada.

Figura 15.6. La
arteriosclerosis es una enfermedad arterial y de los capilares que se genera
cuando la LDL “transportador de grasa de baja densidad” deja grasa a medio
camino de los tejidos, obstruyendo el flujo de sangre. Cuando aumenta la
presión, parte del epitelio de la arteria se rompe generando una reacción
rápida de coagulación que genera un infarto.
Los factores de riesgo más comunes son los
siguientes: hipertensión tanto los valores sistólicos como los diastólicos
influyen a la elevación del riesgo acompañado de hipercolesterolemia.
Gradualmente existe a mayor edad una mayor tensión arterial debido a la pérdida
de elasticidad de los vasos. El consumo de cigarrillos, aumenta la presión
debido a la afección de la microvasculatura generalizada y también por la
predisposición a la arterosclerosis que presentan, aumento de homocisteína en
plasma, factores relacionados con la hemostasia y trombosis, y por supuesto los
antecedentes familiares. Otros posibles factores de riesgo son niveles elevados
de la proteína C reactiva (PCR) en la sangre que puede aumentar el riesgo de
aterosclerosis y de ataques al corazón; altos niveles de PCR son una prueba de
inflamación en el cuerpo que es la respuesta del organismo a lesiones o
infecciones. El daño en la parte interna de las paredes de las arterias parece
desencadenar la inflamación y ayudar al crecimiento de la placa.
Las personas con bajos niveles de PCR
pueden tener aterosclerosis a una tasa más lenta que cuando se tienen niveles
elevados de PCR y la investigación está en curso para establecer si la
reducción de la inflamación y la disminución de los niveles de PCR también
puede reducir el riesgo de la aterosclerosis al igual que las hormonas
sexuales, los estrógenos son protectores de la aterosclerosis y por ende las
mujeres son más afectadas después de la menopausia.
No existe tratamiento médico alguno
demostrado para la arteriosclerosis pese a ser el fármaco probablemente más
buscado por la industria farmacéutica, vamos que todos los CEO “título de los
directores ejecutivos de las empresas más importantes del planeta, son la gente
que en verdad importa en términos económicos” se mueren o de cáncer o de un
ataque al corazón. El tratamiento farmacológico (antihiperlipidémicos,
antiagregantes o anticoagulantes) sirve para disminuir sus causas o sus
consecuencias. El tratamiento quirúrgico es muy resolutivo en la cardiopatía
isquémica y también en otras localizaciones. El tratamiento profiláctico
consiste en evitar los factores predisponentes de la enfermedad y a las
complicaciones de ésta: obesidad, hipertensión, sedentarismo, hiperglucemia,
hipercolesterolemia, tabaquismo, etc. Para ello lo ideal es practicar ejercicio
suave, una dieta equilibrada como la mediterránea, baja en grasas, técnicas de
relajación para evitar el estrés, dejar de fumar, etc.
Investigadores en China encontraron que el
hongo reishi mejora el flujo sanguíneo y baja el consumo de oxígeno del músculo
cardíaco. Resultados similares fueron también encontrados por científicos
japoneses, quienes evidenciaron que el reishi contiene ácido ganodérico, el
cual reduce la presión sanguínea y el colesterol e inhibe la agregación
plaquetaria, la cual puede conducir a ataque cardíaco y otros problemas
circulatorios (Berger et al., 2004; Meletis & Barker, 2005; Mizuno, Sakai,
& Chihara, 1995).
15.6 Venas
Las venas son vasos de recolección, donde
la sangre fluye con una presión baja a una velocidad mínima. Por esta razón,
cerca del 70% del total de la sangre circulante reside en las venas. Esto
implica que, para cumplir con las necesidades metabólicas normales, el cuerpo
solo requiere de un 30% de su sangre distribuyendo oxígeno y nutrientes, aunque
en momentos de necesidad y debido a la adecuada señalización hormonal, la
sangre de las venas p0uede ser liberada parcialmente al lado arterial, para que
existe una mayor presión y velocidad, lo cual a su vez potencia las capacidades
metabólicas del individuo, obviamente esto solo puede suceder durante momentos
de extrema necesidad, dado que si la presión es alta todo el tiempo se corre el
riesgo de dañar el sistema de capilares ocasionando infartos localizados o
incluso un accidente cerebrovascular mortal.
Debido a las bajas o negativas presiones
en la zona venosa del sistema circulatorio, las venas están equipadas con válvulas
de paso de un solo sentido, que funcionan como un torniquete, lo que hace que
la sangre venosa solo pueda avanzar en un solo sentido de manera independiente
a la poca presión sanguínea que puedan tener.
Para contraponerse a la presión negativa y
al obvio hecho de que el corazón no puede hacer mucho, las venas dependerán de
los cambios de presión ocasionadas en las zonas donde se encuentra una alta
masa muscular, que le impregna movimiento y presión nuevos al vaso sanguíneo,
en otras palabras, más que el corazón es el sistema muscular y los movimientos
del resto del cuerpo lo que permite el flujo de sangre en el lado venoso del
circuito cardiovascular. De esta manera, aunque lenta la sangre venosa logra
llegar al corazón. La sangre de los órganos no musculares como los pulmones o
el corazón dependerán de las fuerzas remanentes o simplemente de la gravedad.
Las venas varicosas se ocasionan en puntos donde la presión negativa de la sangre es muy grande y logran vencer a las válvulas de una sola vía de las venas, generalmente en las zonas donde la gravedad es más permanente. Cuando se está mucho tiempo de pie, las venas de las piernas son las que soportan más este problema, lo que provoca o un vencimiento de la válvula o un reforzamiento de toda la vena lo que genera la varice.

Figura 15.7. Al dañarse la
válvula de la vena, el tejido a su alrededor se engrosa para elevar la sangre
por peristalsis.
15.7 Tipos de
varices
Además de resultar antiestéticas, las
varices pueden ocasionar diversos síntomas y dar lugar a complicaciones. Lo
mejor, sobre todo si tienes factores de riesgo, es tomar medidas para prevenir
su aparición.
👉 Varices Grado I o
varículas: En esta etapa, se ven en algunos sitios y a través de la piel, las
venas finas de color violáceo. A veces pueden tener forma estrellada, y se
denominan arañas vasculares. Por lo general son únicamente un problema de tipo
estético pero, en determinadas ocasiones, pueden producir sensación de pesadez
y cansancio en las piernas.
👉 Varices Grado II: Las venas se van haciendo más
visibles y empiezan a notarse los primeros síntomas como: Pesadez y cansancio
en las piernas; Dolor; Calambres; Hormigueos; Sensación de calor o picores y
escozores.
👉 Varices Grado III: Las venas están más dilatadas y
tortuosas. Los síntomas van aumentando progresivamente, y aparece hinchazón y
edemas y cambios de coloración en la piel.
👉 Varices Grado IV: Aparecen zonas eczematosas y
úlceras. Las úlceras son difíciles de tratar y pueden infectarse con facilidad.
👉 Complicaciones de las venas
varicosas: Debe tenerse en cuenta que algunos procesos que se describen en el
apartado de síntomas, aparecen después de un largo proceso de evolución sin
tratamiento (Grados III y IV), por lo que pueden considerarse como
complicaciones. Así pues, podemos considerar como complicaciones de las
varices:
👉 Cambios en la piel: Aparte de la aparición de manchas
originadas por el depósito de glóbulos rojos, cuando la acumulación de estos y
de líquidos aumenta, se producen alteraciones en la piel y en los tejidos que
están bajo la misma (alteraciones tróficas), debido a que los nutrientes no
llegan correctamente. Como consecuencia de estas alteraciones, la piel se
endurece, presenta un aspecto seco y escamoso, con cambios de color
generalizados (ya no son manchas más o menos aisladas) e intensos picores.
👉 Úlceras varicosas: Con mucha frecuencia aparecen como
consecuencia del rascado, debido a los picores por las alteraciones de la piel.
Se presentan sobre todo en la cara interna de los tobillos y son dolorosas, bastante
difíciles de curar, y se infectan con facilidad.
👉 Hemorragias: Se producen por la ruptura de las
venas varicosas, cuya pared está muy debilitada, al igual que la piel, saliendo
la sangre al exterior. Pueden deberse al rascado, por un traumatismo que puede
ser mínimo, o incluso producirse de forma espontánea, pudiendo ocurrir mientras
el paciente duerme, lo que puede aumentar su gravedad si este no se da cuenta.
La sangre suele ser oscura y fluye continuamente, sin borbotones, por ser
sangre venosa. Como todas las hemorragias, son muy escandalosas, pero no suelen
tener demasiada importancia si se tratan adecuadamente; lo que debe hacerse es
elevar la pierna y aplicar un vendaje compresivo y, por supuesto, acudir al
médico para que pueda establecer un tratamiento definitivo, que evite su
reaparición.
👉 Flebitis superficial: Es la inflamación de una vena
varicosa y no debe confundirse con la flebitis profunda, que aparece en situaciones muy distintas y puede tener
graves repercusiones. Se observa la vena varicosa como un cordón dilatado y
endurecido, y hay una zona inflamada a su alrededor, que está caliente, enrojecida y dolorosa.
👉 Infecciones: Las lesiones producidas por el
rascado, así como las úlceras varicosas, e incluso la propia fragilidad de la
piel, pueden facilitar la entrada a organismos oportunistas, y dar lugar a
infecciones que será necesario tratar adecuada e insistentemente, pues suelen
ser bastante rebeldes al tratamiento. La zona infectada es dolorosa, y se verá
inflamada, enrojecida y caliente, pudiendo haber supuración.
👉 Trombosis de las
venas varicosas: Ocurre por la formación de un coágulo en el interior de las venas
varicosas y, con frecuencia, se da conjuntamente con la flebitis, pues esta
puede facilitar la aparición de trombos, y el trombo suele provoca la
inflamación de la vena. Por este motivo, sus síntomas son muy similares.
Normalmente no suele plantear consecuencias graves, salvo que el trombo se
produzca en una zona en que pueda desprenderse y alcanzar la circulación
profunda.
👉 Embolismo pulmonar: Es una complicación muy poco
frecuente en las varices, pues para que ocurra tendría que desprenderse un
trombo y, como se ha señalado, alcanzar la circulación profunda, llegar al
corazón y, desde este, a las arterias pulmonares.
15.8 Microcirculación y sistema capilar
La función del sistema capilar es permitir
el intercambio de sustancias entre un tejido y el sistema circulatorio, como
gases metabólicos, nutrientes, iones salinos y productos de desecho.
Es el componente específico que le da
significado al sistema cardiovascular, el cual soporta las funciones
metabólicas de las células es la microcirculación, si el sistema circulatorio
es un conjunto de pasillos, entonces los capilares serían las puertas de
acceso. La microcirculación está compuesta por tres elementos básicos, las
arterias pequeñas o arteriolas, las venas pequeñas o vénulas y el sistema de
capilares en sí mismo. La sangre que fluye al sistema de capilares es
controlada por esfínteres hechos de musculo liso. A estos esfínteres se los denomina
esfínteres precapilares, son pequeños anillos que restringen la entrada de
sangre al sistema capilar.
Las paredes de las arteriolas, las vénulas
poseen pequeñas capas de musculo liso. Estas capas de musculo liso se
encuentran conectadas al sistema endocrino y nervioso, y de esta manera se
controla cuanto oxígeno y nutrientes son distribuidos a un determinado tejido
en un momento específico. De esta manera, la distribución se ajusta para
concordar con las necesidades metabólicas de un tejido en un momento
específico.
Los capilares también son empleados como
un disipador de calor, cuando el animal se encuentra en movimiento se genera
calor debido a la ineficiencia natural de cualquier sistema físico en
movimiento (si, la entropía nunca falla) cuando se produce mucho calor en el
interior de los tejidos este puede generar una disminución en la eficiencia de
los tejidos e incluso causar daños en estos. Para evitar el desastre térmico,
la sangre captura calor a través de los capilares de los tejidos y envían ese
calor a los capilares de la piel, donde es irradiada al medio externo.
Este detalle puede verse particularmente e
animales con piel sin pelo de color claro, como en algunos seres humanos, los
cuales experimentan un enrojecimiento de la piel, cuando hacen ejercicio. Un
detalle interesante a cerca del sistema capilar es que, aunque individualmente
su volumen es un verdadero chiste “le tomaría horas a una pequeña gota de
sangre atravesar un solo vaso capilar”, tomados de manera colectiva su volumen
es suficiente para vaciar todo el sistema circulatorio mayor, dejando al
corazón y a las arterias principales como la carótida completamente secas. Esto
no ocurre gracias a los esfínteres precapilares, que permiten el paso selectivo
de sangre a algunos capilares y a otros no.

Figura 15.8. Los capilares
son la razón de existencia del sistema circulatorio, son la zona de embarque y
desembarque de nutrientes y desechos, dependiendo del lugar donde nos
encontremos.
Cuando dos tejidos diferentes requieren
sangre para realizar sus actividades metabólicas estos compiten ya que, como
mencionamos antes, no hay suficiente sangre en el cuerpo para llenar todos los
capilares a la vez. Un ejemplo sencillo de esto es cuando se hace demasiado
ejercicio al mismo tiempo que se come un alimento muy difícil de digerir. En el momento en que ambos tejidos abren sus
capilares a la vez, la sangre del sistema circulatorio mayor es absorbida por
estos y la presión sanguina baja mucho, lo que puede causar isquemia “una falla
localizada de la distribución de sangre”.
15.9 Hemodinámica de la circulación
Los patrones de flujo y presión de la sangre circulante, a través de los vasos sanguíneos constituye la hemodinámica de la circulación. Las presiones del sistema circulatorio asociados a los lados arterial y venoso son notablemente diferentes. Cuando los ventrículos del corazón se contraen, el pico de fuerza produce la presión sistólica. La presión diastólica constituye el momento de menor presión en los vasos sanguíneos, el cual se alcanza entre cada latido del corazón. La presión diastólica resulta de la fuerza mantenida por el empuje arterial.
Figura 15.9. Diástole y
sístole (YouTube)
La presión sanguínea se expresa de manera
que represente los valores de ambos momentos sístole/diástole. Por ejemplo, en
adultos jóvenes del género masculino la presión sanguínea promedia el siguiente
valor 120/80 donde la presión sistólica es la primera en enunciarse y la que
presenta el valor más alto. Si las arterias empiezan a presentar síntomas de
enfermedad o degeneración “como perdida de elasticidad o acumulación de agentes
obstructores” la presión sanguínea se eleva.
Unas de las mayores presiones sanguíneas
que pueden registrarse en los vertebrados pertenecen a las jirafas, promediando
un valor de 260/160 al nivel del corazón. Tal presión es necesaria para que la
sangre pueda ascender en contra de la gravedad a través del largo cuello del
animal. Una vez que la sangre ingresa al corazón, la presión habrá disminuido a
120/70, un valor comparable al de los humanos.
En la mayoría de los vertebrados la
presión sanguina disminuye una vez que la sangre deja el corazón. La baja en la presión se debe a dos factores,
la fricción de la sangre con las paredes de la cara interna del vaso sanguíneo
y a la perdida de área de los vasos sanguíneos una vez se acercan a los
capilares. El flujo que cualquier líquido o fluido a través de un tubo hueco es
resistido por la fricción de las partículas del líquido contra la pared del
mismo.
El momento de mayor pérdida de presión
ocurre en los capilares, los cuales carecen además de tejido elástico, lo que
implica que, no solo es el punto de menor presión, si no que este DEBE SER el
punto de menor presión, de lo contrario puede ocurrir un derrame de un vaso
sanguíneo y ocasionar un infarto localizado. Una vez que la sangre deja los
capilares para ingresar a las venas, la sangre sigue con una presión baja o
incluso negativa, es decir con una tendencia a devolverse debido a la poca
fuerza que posee. Esta tendencia problemática debe ser resuelta por los vasos
sanguíneos venosos.
16. Organización del sistema circulatorio
|| Regresar al Índice ||
Antes de comenzar con esta sección, se le
aconseja al lector ya haber leído la sección del sistema respiratorio llamada “Relación
en entre sistema respiratorio y circulatorio”.
16.1 Los vasos sanguíneos principales
Los vasos sanguíneos son tan diversos,
como los órganos a los que estas suplen con nutrientes. Sin embargo, la
diversidad en los cordados, están basadas en modificaciones a un plano general
de organización heredado desde el último ancestro común de todos los cordados.
Debido a que, generalmente el sistema de vasos sanguíneos se encuentra
intensamente modificado en muchos de los organismos tipo que son analizados en
la actualidad como animales de referencia, el plano básico solo es visible con
claridad en unos pocos cordados, que han retenido un patrón más básico y
gener

Figura 16.1. Modelo del sistema circulatorio de los cefalocordados.

Figura 16.2. Modelo del
sistema circulatorio de un pez estrictamente acuático.
Como lo mencionamos en temas anteriores, a
pesar de que el corazón en sí de los cefalocordados no es homologable al de los
demás vertebrados, su sistema circulatorio sí que lo es Figura 16.1,
basándose en una arteria aorta donde la sangre fluye hacia los arcos aórticos,
donde pasan a la aorta dorsal, donde la sangre avanza desde los arcos a la
cola. A lo largo del camino existen múltiples derivaciones que alimentan los
diferentes órganos. Este es, en términos generales el plano característico del
sistema circulatorio de todos los cordados, excepto en los tunicados.
El sistema circulatorio de un tiburón en
comparación con el del cefalocordado no existen grandes modificaciones, y tal
vez el más grande desarrollo está en el corazón y en las derivaciones a los
nuevos órganos que aparecen en los grupos más recientes de cordados. Por lo
general, la sangre que sale del corazón, ingresa a una arteria principal no
dividida en su base llamada aorta ventral, y avanza en dirección de la cabeza
hasta llegar a la faringe. En este punto, generalmente la aorta ventral genera
una serie de derivaciones que forman unos arcos alrededor dela faringe,
denominados arcos aórticos, mientras que la última derivación genera las
carótidas externas, que se encargan de llevar sangre al cerebro.
Los arcos aórticos, pasan en medio de los
arcos branquiales, y en los animales con branquias funcionales, son
precisamente estos arcos aórticos los que se expanden, aplanan y forman los
capilares generando lo que llamamos branquias. Después de pasar por los arcos
branquiales, los arcos aórticos se unen a una arteria principal en la zona
dorsal del animal, por lo que se la denomina aorta dorsal. Saliendo de la punta
más frontal de la aorta dorsal se genera otro par de arterias carótidas,
denominadas carótidas internas, las cuales llevan sangre hacia la cabeza,
generalmente penetrando en la cavidad craneana para suplementar al cerebro con
sangre. Sin embargo, esta es la excepción, la aorta dorsal por sí misma, en su
mayoría lleva sangre desde la zona anterior “cerca a la cabeza” hacia la zona
posterior “hacia la cola”. Cerca del nivel del hígado, los dos vasos sanguíneos
paralelos de la aorta se unen para formar una sola aorta, la cual distribuye la
sangre hacia la parte posterior y eventualmente extendiéndose hasta la cola y
la región caudal. A lo largo del camino, la aorta dorsal da nacimiento a una
serie de arterias parietales hacia la pared corporal local, así como a una
enorme cantidad de arterias principales, generalmente especulares “en parejas
simétricas” hacia algunos tejidos y órganos.
El par de arterias subclavas suplen a los
apéndices anteriores (aletas frontales o apéndices frontales) y usualmente se
originan directamente de la aorta dorsal. El par de arterias iliacas suplen a
los apéndices posteriores (aletas pélvicas si las hay, apéndices pélvicos),
estas también se derivan directamente de la aorta dorsal. Las arterias
genitales (ováricas o espermáticas) llevan sangre hacia los órganos
reproductivos. El par de arterias renales suplen de sangre a los riñones, son
vasos sanguíneos muy grandes que se derivan de la aorta dorsal. Esto asegura
que los riñones reciben sangre muy rápido en el circuito arterial, mientras que
la presión sanguínea es aun relativamente alta, un factor en el movimiento de
la sangre “hemodinámica” que facilita la labor de filtrado de metabolitos
tóxicos.
Típicamente, los vertebrados poseen tres
derivaciones de la arteria aorta dorsal que no van en parejas, estas son la
arteria celiaca que suple de sangre al hígado, el bazo, el estómago y parte de
los intestinos. La arteria mesentérica anterior que suple de sangre a la
mayoría del intestino delgado. Y finalmente la arteria mesentérica posterior
que suple de sangre al intestino grueso. En los vertebrados actuales que
descienden de linajes muy antiguos, la sangre regresa al corazón lo hace por
varias venas prominentes. La vena
cardinal común también llamada sinus, es la vena más grande que recibe la
sangre retornante de la vena cardinal anterior (precardinal) y de la vena
cardinal posterior (postcardinal). Estas drenan las parte anterior (cabeza y
tronco) y posterior (cola), respectivamente.
Venas tributarias de los apéndices
superiores usualmente descargan en la ven cardinal común por medio de la vena
subclava. Las venas de la región lateral del cuerpo, y de los apéndices
posteriores también descargan su contenido en la vena cardinal común vía la
vena lateral abdominal. El sistema de venas porta, es una ruta vascular que
comienza en un sistema capilar y termina en otra red capilar, sin que la vena
atraviese en algún momento el corazón branquial (o cualquier corazón). Esta
aclaración la hacemos porque los peces sin mandíbula poseen corazones accesorios
justo en las venas porta del intestino/hígado diferentes al corazón principal,
pero los demás vertebrados no lo poseen.
16.2 Sistemas portal
La vena porta hepática fluye desde la red capilar del intestino delgado hacia el hígado, y puede o no poseer un corazón accesorio en medio. La función de la vena porta hepática es la de transportar los nutrientes absorbidos por el intestino delgado hacia el hígado, para almacenamiento y procesamiento de muchos productos finales de la digestión.

Figura 16.3. Normalmente las
venas y arterias se conectan por un capilar, pero en ocasiones hay dos
capilares conectados entre sí, los vasos que conectan a dos capilares sin
corazón en medio son portales, los cuales, dependiendo de la estructura misma
del vaso serán venosos o arteriales.
16.3 Sistema
portal hepático
La vena porta hepática se forma cuando la arteria que alimenta a los intestinos se transforma en su respectiva vena, esta conecta con los capilares del hígado y posteriormente si avanza hacia el corazón.
Figura 16.4. El sistema
portal hepático son venas que fluyen desde los capilares intestinales a los
capilares hepáticos sin pasar por un corazón.
Es notorio el fenómeno de disminución de
presión al transformarse el vaso sanguíneo en vea, sin embargo, de todos los
craneados, solo los más ancestrales como los mixinos o las lampreas poseen un
corazón porta que eleva la presión y permite el paso de sangre a través del
segundo sistema capilar. La razón por la cual los demás vertebrados no requieren
un segundo corazón tal vez se deba a que corazones branquiales son más
potentes.
El sistema porta hepático conecta el
hígado con todas las superficies donde puede absorberse nutrientes, ya sabemos
que yeyuno y el duodeno son las zonas más importantes para este fenómeno, pero,
aunque sea leve, en prácticamente cualquier superficie de tracto
gastrointestinal puede realizarse absorción de nutrientes.
16.4 Sistema
portal renal
El segundo sistema de venas porta, es la
vena porta renal. El sistema de vena porta renal comienza en los capilares de
la cola y las extremidades pelvianas, y se reúne en una pareja de venas, las
cuales descargan su contenido en los riñones. La función del sistema de venas
porta renales no ha sido bien entendido. Sin embargo, debido a que transporta
sangre caudal directamente hacia los pulmones, algunos autores han sugerido,
que estas venas proveen una ruta directa para el filtrado de desechos
metabólicos producidos por el tejido muscular caudal. Otros autores han
sugerido que es un mecanismo para disminuir la presión sanguínea de la sangre
que regresa del área caudal.
bien la función de filtrado de un riñón
ocurre en la zona arterial del circuito y requiere de una alta presión para
filtrar, el riñón también requiere de una zona de baja presión para reabsorber
ciertos nutrientes y especialmente agua. El sistema de venas porta renal, al
disminuir la presión del área venosa, permite dicha zona de baja presión. El
sistema de venas porta renal se encuentra presente en todas las clases de
vertebrados excepto en los mamíferos. Aunque los mamíferos carecen del sistema
de vena porta renal, el riñón mamífero aún posee una zona de baja presión, el
cual puede ser la contraparte de los mamíferos al sistema de venas porta de los
riñones, el cual permite la recuperación de agua de la orina en formación.
16.5 Sistema
circulatorio menor y mayor
Las modificaciones filogenéticas en este
patrón básico de venas y arterias están altamente correlacionadas, con cambios
funcionales, y adaptaciones fisiológicas al entorno. La circulación en un pez estrictamente
acuático posee un solo circuito, avanza por el corazón, pasa por las branquias,
avanza hacia los capilares y regresa al corazón. Este circuito es el clásico
que siempre se enseña, pero en la naturaleza existe una amplia variedad de
otros circuitos circulatorios.
Durante la transición desde la vida acuática a la vida terrestre, las branquias dan paso a los pulmones, esto se acompaña con una alteración en una gran cantidad de vasos sanguíneos, primeramente, una derivación de algunos arcos aórticos hacia la vejiga natatoria, y luego, una derivación venosa desde a vejiga natatoria hacia el corazón directamente. El sistema circulatorio de los vertebrados terrestres generalmente posee dos circuitos, el mayor o sistémico homólogo al de los peces solo que sin branquias "generalmente" y el menor o pulmonar.

Figura 16.5. Esquema sobresimplificado de un pez que no es estrictamente acuático, con un corazón lineal.

Figura 16.6. Esquema
sobresimplificado de la arquitectura de los corazones y los circuitos
circulatorios de peces, anfibios y reptiles. Estos esquemas fueron populares a
mediados del siglo XX cuando muchos paleontólogos y fisiólogos aún se apegaban
a la teoría ortogenesista de la evolución, la cual planteaba una progresión
lineal de estructuras, la realidad es mucho más compleja.
A estas dos derivaciones las conocemos
como vena pulmonar y arteria pulmonar. En algunos peces, y en la mayoría de los
tetrápodos, las venas cardinales se hacen menos importantes en la recolección
de sangre sistémica hacia el corazón. En reemplazo se desarrollan de manera más
prominente la vena precava (vena cava posterior) que recupera la sangre de la
parte posterior del cuerpo (cola, apéndices inferiores y viseras) y la vena
precava (vena cava anterior) que se encarga de drenar la parte anterior del
cuerpo (tronco posterior y cabeza).
16.6 Los vasos sanguíneos venosos
Las venas principales que retornan la
sangre hacia el corazón son complejas y altamente variables. En cada uno de los
grupos, las venas componen pocos sistemas funcionales que emergen
embriológicamente, de lo que parece ser un patrón de desarrollo común. En los
vertebrados que poseen un circuito circulatorio doble, uno mayor y otro menor,
tenemos un sistema venoso desde el sistema de órganos y tejidos, y otro que
regresa de los pulmones. Aun dentro del sistema de venas que regresa de los
órganos existen unos subtipos de venas denominados porta, los cuales quedan en
medio de dos sistemas capilares. El sistema venoso sistémico representa todos
los vasos sanguíneos que al emerger de un capilar regresan al corazón de manera
desoxigenada. Este sistema de vasos es el que más se asocia con los colores,
llevado a las expresiones algo imprecisas de sangre venosa “desoxigenada” y
sangre arterial “oxigenada”, tales expresiones son solo aplicables para los
vasos sanguíneos venosos sistémicos, pues como hemos definido y como veremos
posteriormente, el sistema circulatorio menor posee un “cambio de colores”.
Temprano durante el desarrollo, existen
tres grupos grandes de venas en pareja: las venas vitelinas que lo conectan a
las membranas alimentarias; las venas cardinales del cuerpo mismo del embrión;
y las venas abdominales laterales de la región pélvica. El par de venas
vitelinas están entre los primeros vasos sanguíneos en desarrollarse en el
embrión. Emergen desde las membranas embrionarias hacia el embrión en
desarrollo. En ese momento giran hacia la parte anterior “hacia la cabeza”,
avanzando por las vísceras y luego ingresando al sinus venosus del corazón en desarrollo “y recordamos que el sinus venosus se
manifiesta en TODOS los corazones así sea en el embrión”.
El primordio del hígado crece en las venas
vitelinas. Los cordones hepáticos se desarrollan de manera asociada a las venas
hepáticas como sinusoides hepáticos. Poco a poco con el desarrollo de los
sinusoides en el hígado, las secciones de vasos sanguíneos adquieren el nombre
de vena hepática. Las venas cardinales concluyen en las venas cardinales
anteriores, las cuales drenan sangre desde la región de la cabeza, y las venas
cardinales posteriores, que drenan la sangre desde los órganos y los tejidos
del resto del cuerpo.
Ambos pares de venas cardinales se unen
poco antes de llegar al corazón para formar las cortas venas cardinales
comunes, las cuales ingresan al sinus
venosus. Las venas cardinales
anteriores consisten en una gran cantidad de vasos sanguíneos que se unen poco
a poco desde el cerebro, el cráneo, el cuello, los músculos y la piel. Las
venas cardinales posteriores se desarrollan principalmente como vasos
sanguíneos de los riñones aun en desarrollo. Las venas abdominales laterales
están presentes en peces, pero estas son generalmente absorbidas o no se
desarrollan en los tetrápodos. En los peces cada vena se une a la vena iliaca,
desde las aletas pelvianas y fluyen hacia delante por las paredes laterales del
cuerpo del pez. Al nivel del hombro, las venas iliacas se unen a la vena
braquial, y poco después se transforma en la vena subclava, la cual poco
después ingresa a la vena cardinal común.
Sin embargo, en los tetrápodos, las venas
subclavas regresan de manera independiente al corazón, y las venas laterales
abdominales ingresan al hígado. En los anfibios, las venas laterales abdominales
se unen en una vena media común, la vena abdominal ventral, la cual fluye a
través de la pared del celoma ventral de animal. En los cocodrilos, las aves y
los mamíferos, las venas abdominales se encuentran ausentes. Los desarrollos
venosos posteriores son mucho más complejos que los vistos en la zona arterial,
conllevando al desarrollo de sistemas venosos asimétricos muy complejos. Muchos
peces poseen órganos de reparación accesorios, sin embargo, los vasos
sanguíneos recientemente oxigenados reingresan a la circulación general antes
de ingresar al corazón a través de las venas cardinales. De entre los peces más
cercanos a los tetrápodos y los dipnoos, los polipteriformes, a pesar de poseer
una derivación pulmonar desde su sexto arco aórtico tal como en los peces
pulmonados y los demás tetrápodos, en ellos la vena retornante desde la vejiga
natatoria no reingresa directamente al corazón, sino que, también, se une a la
vena cardinal común.
Es en los dipnoi donde la vena retornante desde el corazón reingresa
directamente al corazón, y no al sinus venosus, si no de manera directa al ventrículo. En los
dipnoos que poseen pulmones en pareja, son dos venas las que emergen, las
cuales se unen poco antes de ingresar al corazón. Embriológicamente, la vena pulmonar
no emerge por la conversión de una vena preexistente. En su lugar una gran
cantidad de vasos sanguíneos pequeños se originan de manera independiente para
drenar los pequeños bulbos pulmonares embrionarios. Poco después, los vasos
sanguíneos se fusionan en las venas pulmonares que ingresan al ventrículo del
corazón.
17. Los arcos aórticos en peces
|| Regresar al Índice ||
El numero ancestral de arcos aórticos y arcos branquiales a través de los cuales corren los primeros es materia aun de debate. Algunos peces ostrocodermos poseían 10 pares de arcos branquiales, y presumiblemente una igual cantidad de aórticos. Los ostrocodermos fueron peces sin mandíbulas acorazados, muy relacionados con los placodermos. Actualmente, los parientes vivos más cercaos a ellos son las lampreas y los mixinos, aunque es debatible. El número de pares de arcos aórticos varía mucho en los cordados vivos. Las lampreas poseen 8 arcos, los mixinos poseen 15. Algunas especies de tiburones poseen de 10 a 12.

Figura 17.1. Los peces dipnoi y los peces con vejigas
natatorias posen una arteria branquial eferente en el 6 arco "el más
lejano a la cabeza" que se divide en dos, una va al órgano
respiratorio/flotador y es homólogo a la arteria pulmonar, el otro conecta con
el sistema circulatorio sistémico, aunque no aparece en los adultos,
embrionalmente también aparece, incluso en los humanos, son tan homólogos que
se los denomina con el mismo nombre, ductus
arteriosus "ducto arterial".
Sin embargo, solo 6 aparecen de manera rutinaria durante el desarrollo embrionario en la mayoría de los peces con mandíbula “incluyendo en este grupo a los vertebrados terrestres”. De acuerdo con lo anterior, 6 será el número de arcos que usualmente se toma como canónico, debido a la evidencia embrionaria. Por lo general estos se designan mediante numerales romanos.

Figura 17.2. Diagrama muestra los arcos aórticos de una lamprea.

Figura 17.3. Arcos aórticos
generales que normalmente permanecen en los peces con mandíbula, se numeran en
romano desde el más lejano del corazón I hasta el más cercano al corazón VI.
La variación filogenética entre los arcos aórticos puede ser intrincada. Aunque, la referencia a un patrón básico de 6 arcos es una simplificación bastante útil. Sin embargo, la literatura a cerca del nombramiento de cada arco aórtico es confusa, y cada autor puede ponerle un número diferente a un arco diferente dependiendo de sus gustos. Otros autores insisten en usar números hasta el X, en reconocimiento de que este sea el número de arcos verdaderamente ancestral. Otros abandonan el esfuerzo de adjudicar homologías, ignoran los arcos que se pierden filogenéticamente y simplemente numeran los arcos tal como los encuentran en el adulto “lo que implica que también ignoran los posibles arcos que se generan de manera transitoria durante el desarrollo embrionario”. Sin embargo, en esta serie de artículos hemos tomado, y seguiremos tomando la opinión de Kardong y colaboradores (2012), tomando 6 pares para representar el patrón embrionario básico.

Figura 17.4. Los arcos
aórticos de un embrión de tiburón: muestra los primordios de 6 arcos, sin
embargo, el arco I no se forma completamente, y en su lugar cierta sección del
arco I se conecta al arco II. La zona eferente de los arcos "rojas"
se bifurcan alrededor del arco branquial para formar bucles que en el adulto
interconectan todas las branquias, llamados canales postremáticos y
pretremáticos.
17.1 Los peces
cartilaginosos
Rápidamente después de que la aorta ventral sale del corazón, esta genera una serie de derivaciones, los arcos aórticos, los cuales, al llegar a los arcos branquiales se subdividen generando la red capilar que permite el intercambio de gases. La parte del arco que conecta desde la aorta hasta la branquia se denomina arteria aferente, mientras que la que transporta la sangre desde la red capilar a la arteria dorsal se denomina arterias eferentes. La red capilar, en medio de las arterias, rodean la zona cartilaginosa de las branquias.

Figura 17.5. Los arcos
branquiales detallados de un tiburón. Se observa que el canal aferente para el
espiráculo no se forma del arco branquial I si no del III. Recalcamos que el embrión
posee un arco aórtico II diferenciado, mientras que en el adulto el arco II también
se combina parcialmente al arco III a través del bucle tremático.
En los peces cartilaginosos, el primer
arco branquial se reduce mucho, pero no se pierde, formando un orificio
pequeño, algunas veces redondo, denominado espiráculos. Durante el desarrollo
embrionario, la zona ventral del primer arco aórtico, que se esperaría alimenta
al primer arco branquial no se desarrolla. Aunque el espiráculo aún mantiene en
los adultos una red capilar, esta proviene no de la aorta dorsal, si no de otro
arco branquial ya con sangre oxigenada. Por esto se piensa que el espiráculo
tiene un rol respiratorio nulo, pero es empleado para alguna función, dada su
necesidad de una red capilar propia, tal vez como un órgano sensorial.
Lo anterior implica que en los peces
cartilaginosos empieza a existir una tendencia a la perdida de los arcos
branquiales frontales. Otro detalle interesante, la arteria coronaria se deriva
directamente de los arcos aórticos oxigenados. Los arcos aórticos remanentes
(del II al VI) forman derivaciones que finalmente terminan con la formación de
bucles, que funcionan como una red capilar entre las branquias. Aunque la
arteria carótida externa emerge en el embrión en desarrollo, desde la punta de
la aorta ventral, esta se desvía posteriormente a los bucles branquiales,
llevando sangre oxigenada finalmente hacia la mandíbula. La arteria carótida
interna que suple al cerebro con sangre oxigenada desde el arco branquial II
obtiene la sangre desde la arteria branquial eferente.
17.2 Los peces
óseos
La tendencia que se ve en los cartilaginosos también se ve en los peces óseos, donde el primer arco branquial desaparece en la mayoría de las especies completamente, o en algunas formas un espiráculo como en los tiburones. En la mayoría de los peces con órganos de respiración accesorios, la sangre oxigenada obtenida desde estos órganos, ingresa de manera directa a la circulación sistémica venosa antes de llegar al corazón, incrementando los niveles promedio de oxigeno de la sangre que regresa al corazón.

Figura 17.6. En la mayoría
de los peces óseos, se forman cuatro pares de arcos aórticos (III, IV, V y VI).
Ellos se asocian a las branquias mediante 5 arcos branquiales. En varias especies
la primera hendidura de la faringe persiste como un pequeño espiráculo, sin
embargo, en la mayoría es solo una estructura juvenil transitoria.
17.3 Los peces
pulmonados
En los peces pulmonados, la sangre oxigenada que sale de los pulmones a través de la vena pulmonar ingresa directamente al ventrículo del corazón. Filogenéticamente, los peces pulmonados serían los exponentes más antiguos de la tendencia a la separación de corrientes sanguíneas, es decir, de la formación de un segundo circuito circulatorio, denominado circulación menor o circulación pulmonar.

Figura 17.7. Arcos aorticos
en vista lateral derecha de un pez pulmonado adulto del género Protopterus. En
este caso tenemos un estado de respiración branquial, cuando el pez no respira
a través de su pulmón, en ese caso posee el esfínter de la vena pulmonar
cerrada. En los peces pulmonados, así como ocurre en la mayoría de los peces
óseos, la primera hendidura branquial se reduce a un espiráculo sin función
respiratoria, o simplemente desaparece sin dejar rastro. Este se encuentra
asociado al arco aórtico I, el cual también se encuentra reducido. En el pez
pulmonado australiano, Neoceratodus, las 5 hendiduras faríngeas remanentes
se abren a branquias completamente funcionales, las cuales son alimentadas por
cuatro arcos aórticos (III, IV, V y VI).
Cuando el agua posee poco oxígeno, las
branquias no oxigenan la sangre, en tal caso el ducto arterial se cierra y se
abre la arteria pulmonar se abre para alimentar los pulmones. En el pez
pulmonado africano, Protopterus, la funcionalidad de las branquias se
encuentra reducida. Lo formación de los capilares de los arcos III y IV no está
presente “lo que implica que las branquias correspondientes no funcionan”. En
todos los peces pulmonados, el vaso sanguíneo eferente del arco aórtico (VI) el
cual se encuentra más cercano al corazón, genera una derivación denominada
arteria pulmonar, pero mantiene su conexión a la aorta dorsal “tal como ocurre
con los embriones de los mamíferos”. A esta derivación la denominaremos ducto
arterial “ductus arteriosus”.
18. Los arcos aórticos en los vertebrados terrestres
18.1 Las
salamandras
Como ya vimos en los artículos de los corazones, los anfibios fueron considerados como poseedores de un sistema circulatorio ineficiente, debido a que su ventrículo no cuenta con tabiques y a que solo tienen una arteria, por la cual sale tanto sangre oxigenada como desoxigenada, invitando a la conclusión de que ambos flujos de sangre se mezclaban. No solo eso, de hecho, la anatomía de los arcos branquiales “si ellos también tienen” también invitaba a esta conclusión. En los anfibios como las salamandras los primeros dos arcos branquiales (I y II) desaparecen muy temprano durante el desarrollo embrionario.

Figura 18.1. Arcos aórticos
en una vista lateral de un renacuajo de salamandra. Podemos observar que es
prácticamente el mismo del de un pez dipnoi.
De hecho, los peces alevines de los dipnoi
también presentan un pulmón en desarrollo no funcional y branquias externas.
Los patrones de los arcos restantes difieren mucho, dependiendo de si hablamos
de la etapa de renacuajo, que posee branquias completamente funcionales, o de
si hablamos del adulto generalmente con pulmones funcionales (o que depende de
la piel o ambos). En la mayoría de los renacuajos de salamandra, los tres
siguientes arcos aórticos (III, IV y V) portan branquias externas, mientras que
el último arco aórtico (VI) genera directamente la arteria pulmonar. Sin
embargo, la biología es la ciencia de las excepciones, y en este caso no es la
excepción. En la salamandra neotenica Necturus,
en la cual el arco aórtico desaparece, manteniéndose solo su sección dorsal
formando la base de la arteria pulmonar.
La sección de la aorta ventral entre los
arcos III y IV se transforma en la arteria carótida común, la cual alimenta a
la carótida externa y la carótida interna. Los siguientes dos arcos (IV y V),
conforman los vasos sanguíneos sistémicos principales que se unen a la aorta
dorsal. El arco aórtico final (VI) también se une a la aorta dorsal, su última
y corta sección forma el ducto arterial.
Poco antes de unirse a la aorta dorsal, el
sexto arco aórtico genera la derivación que genera la arteria pulmonar, la cual
a su vez se divide en pequeñas derivaciones que atraviesan la zona baja de la
boca, la faringe y el esófago antes de ingresar realmente a los pulmones. En
las salamandras sin pulmones, la arteria pulmonar, si esta persiste, se conecta
a la piel del cuello y la espalda en lugar de a los pulmones.
18.2 Anuros
“ranas y sapos”
En los anuros (ranas y sapos), el renacuajo posee generalmente branquias internas, las cuales residen en los últimos cuatro arcos aórticos (III, IV, V y VI), y la arteria pulmonar embrionaria es igual al del renacuajo de una salamandra. Durante la metamorfosis, las branquias se pierden completamente, junto con el ducto carótido V. Los arcos aórticos persistentes (III, IV y VI) se expanden para suplir de sangre a la cabeza, el tronco y los circuitos pulmonares respectivamente. El tercer arco aórtico, junto con la sección asociada de la aorta dorsal anterior se transforma en la arteria carótida interna. La porción anterior de la aorta ventral se convierte en la carótida externa.

Figura 18.2. Los arcos aórticos en el adulto son muy diferentes al del renacuajo, comenzando por la pérdida de los capilares branquiales y la activación del pulmón. Se pierde una sección de la aorta dorsal entre los arcos aórticos III y IV llamado ducto carótido. En la mayoría de las especies de salamandras, las branquias externas se pierden durante la metamorfosis, pero los arcos aórticos son retenidos como los principales vasos sanguíneos sistémicos. La sección de la aorta dorsal que conecta los arcos aórticos III y IV, se denomina ducto carótido, usualmente se cierra durante la metamorfosis. Esto fuerza las carótidas llenarse con sangre de una derivación de la aorta ventral.

Figura 18.3. Los arcos aórticos
de los anuros en una vista lateral de un anuro "rana o sapo" en su
estado adulto. El estado larval o de renacuajo es muy similar al de la
salamandra, solo que las branquias internas. La principal novedad con respecto
al adulto de la salamandra es que el ducto arterial se desvía ya no conecta a
la aorta si no a la piel, recibiendo el nuevo nombre de arteria cutánea. Las carótidas interna y externa se derivan
ambas de la carótida común, la sección de la aorta ventral entre los arcos III
y IV. Un cuerpo carótido puede ser encontrado usualmente en la base de la
carótida interna. El siguiente arco aórtico expandido es el arco (IV) se une
con la aorta dorsal, convirtiéndose en la mayor arteria sistémica que suple al
cuerpo. El último arco aórtico (VI) pierde su conexión a la aorta dorsal debido
a que el ducto arterial se cierra “algo muy similar a lo que ocurre a un feto
humano durante el nacimiento, en el que también su respectivo ducto arterial se
cierra”. Sin embargo, a diferencia de los mamíferos, en los anfibios, el ducto
arterial no se retrase, en su lugar se desvía transformándose en la cutánea.
Como ya hemos visto, el sexto arco
arterial forma una derivación en Y, un lado va hacia la aorta dorsal y la otra
hacia el pulmón o vejiga natatoria. Y lo mismo sucede con las ranas, sin
embargo, en el adulto la primera derivación va ahora hacia la piel, mientras
que la segunda derivación permanece hacia los pulmones, pero su desarrollo e
importancia se hace más marcada. El igual que ocurrió con los peces dipnoi,
donde existía un tabique parcial en el corazón tanto a la altura de la aurícula
y el ventrículo, así como una única arteria de salida, los fisiólogos
originales pensaron que el sistema circulatorio de los anfibios era poco
eficiente debido a que el ventrículo no estaba tabicado y a que también existía
solo una única arteria de salida.
Al haber un solo ventrículo y una única
arteria, se concluyó que, las sangres oxigenada y desoxigenada deberían
mezclarse de manera obligatoria. Sin embargo, no fue hasta que se realizaron
los trabajos de fisiología experimental en los que se demostró que de hecho, la
mezcla de sangres es muy poca, y que en su mayoría, los flujos de sangre
oxigenada y desoxigenada permanecen independientes gracias a que la aorta posee
una serie de pliegues en espiral que separan las sangres; el como el único
ventrículo mantiene la independencia de los flujos de sangre es un verdadero
misterio, aunque se ha dicho que las trabéculas “pliegues del endocardio en la
sección del cono ventricular” juegan un rol importante.
18.3 Reptiles
Comenzando por los reptiles, pero representado en las aves y los mamíferos, los arcos aórticos empiezan a presentar una tendencia a la asimetría durante el desarrollo hacia el adulto.

Figura 18.4. Los arcos
aórticos en una vista lateral de un reptil. La característica principal que
observamos es la división de la aorta dorsal en dos arterias de salida, el arco
pulmonar que es homólogo al arco aórtico VI, y dos aortas una derecha y otra
izquierda. A parte de ellos solo permanecen los arcos aórticos III y IV. La
base del arco aórtico izquierdo “derivado de la aorta ventral”, junto con el
lado izquierdo del arco aórtico IV y la sección curva de la aorta dorsal en la
que continua el flujo de sangre constituyen en conjunto el arco aórtico
izquierdo. El arco sistémico derecho comprende los mismos elementos, pero al
lado derecho: la base del arco aórtico derecho derivado de la aorta ventral, el
lado izquierdo del arco aórtico IV y la sección curva de la aorta dorsal
derecha. Los arcos sistémicos se unen detrás del corazón para formar la aorta
dorsal común. El arco aórtico derecho tiende a ser el más prominente,
principalmente debido a los vasos sanguinos adicionales que este suple.
Los arcos aórticos III, IV y VI persisten
en los reptiles, pero la mayoría de los cambios se centran en expansiones y
modificaciones en el arco aórtico IV. Tal vez la modificación más significativa
el sistema arterial en los reptiles es la subdivisión de la aorta ventral.
Durante el desarrollo embrionario, la aorta ventral se divide para formar las
bases de tres arterias separadas que salen del corazón. El arco aórtico
izquierdo, el arco aórtico derecho (ambos unidos por un puente anatómico
llamado Foramen de Panizza), y el tronco pulmonar. El tronco pulmonar incorpora
las bases de ambos lados el arco aórtico VI. El tronco pulmonar se une ahora al
arco VI, el cual, como de manera tradicional tiene las conexiones a los
pulmones en el adulto. De esta manera, los brazos del arco VI se convierten en
el arco pulmonar, una arteria en Y conecta al corazón con los dos pulmones.
Por ejemplo, las arterias carótidas, que
se originaban de la aorta ventral en los grupos de vertebrados más antiguos,
emergen en los reptiles del arco sistémico derecho. La sangre que pasa a través
del arco sistémico derecho debe fluir por el cuerpo o ingresar a la carótida
para suplir de sangre a la cabeza. En la mayoría de los reptiles, las arterias
subclavas se derivan de la aorta dorsal, pero en algunos reptiles, estas se
derivan de los arcos sistémicos. Estas modificaciones de los arcos aórticos en
los reptiles producen un circuito pulmonar y dos circuitos sistémicos, cada uno
surgiendo de manera independiente del corazón.
18.4 Las aves
En las aves, el arco sistémico derecho se hace predominante temprano durante el desarrollo embrionario, mientras que el izquierdo nunca se desarrolla de manera completa.

Figura 18.5. Las carótidas
comunes pueden derivarse del arco sistémico derecho de manera independiente, o
emerger como una única arteria carótida y después hacer una derivación en Y. En
algunos reptiles, como por ejemplos en las tortugas existe un vaso sanguíneo
grande, pero algo corto, denominado arteria braquiocefálica; sin embargo, en
las aves sirve como en vaso sanguíneo más importante en la zona anterior en
muchas aves. La arteria braquiocefálica también se deriva del arco aórtico
derecho. Más allá de esta unión, el arco sistémico se curva posteriormente para
suplir con sangre al resto del cuerpo. En las aves, tal como en los reptiles,
los arcos pulmonares se forman de la base de ambos lados del arco aórtico VI, y
ambos brazos suplen a los pulmones.
Tal como en los reptiles, el arco aórtico
derecho surge de la combinación de tres vasos sanguíneos, la base del arco
aórtico derecho derivado de la aorta ventral, el lado derecho del arco aórtico
IV, y la sección curva de la aorta dorsal. Las carótidas emergen generalmente
de los mismos componentes de los arcos aórticos como en los reptiles (el arco
aórtico III y partes de la aorta ventral y dorsal), y se derivan del arco
aórtico derecho. Sin embargo, el par de arterias suclavas que alimentan las
alas emergen de las carótidas internas y no de la aorta dorsal. Las carótidas
comunes y las arterias subclavas suplen la cabeza y las extremidades frontales
respectivamente.
18.5 Mamíferos
En los mamíferos, se forman los 6 arcos aórticos canónicos, pero solo tres persisten durante el desarrollo, experimentando modificaciones sucesivas. Al final, estos tres arcos aórticos formaran, las arterias carótidas, el arco pulmonar y el arco sistémico. Las arterías carótidas y las arterias pulmonares son ensambladas de los mismos componentes sobre los cuales se ensamblan sus homólogos en los reptiles. Las carótidas en los mamíferos se forman a partir de ambos lados del arco aórtico III y partes de la aorta ventral y dorsal. El arco pulmonar se forma de las bases de ambos lados del arco aórtico VI “nuevamente la ruta canónica establecida desde los peces pulmonados”.

Figura 18.6. El arco
sistémico emerge de manera embrionario únicamente del brazo izquierdo del arco
aórtico (IV) y el lado izquierdo de la aorta dorsal, mientras que el lado
derecho no se genera. Nuevamente un tema que ya se intuía en los reptiles y las
aves “aunque en el otro lado”. De esta manera podemos decir que el sistema
circulatorio sistémico en los mamíferos depende solo del lado izquierdo del
arco aórtico IV. Las carótidas comunes pueden compartir un origen
braquiocefálico o derivarse de manera independiente en diferentes puntos del
arco aórtico. Las otras diferencias notables en los mamíferos es la formación
de las arterias subclavas. La arteria subclava izquierda emerge del arco
sistémico izquierdo. La arteria subclava derecha emerge el brazo derecho del
arco aórtico IV, partes de la aorta dorsal derecha y las arterias que crecen de
manera adjunta hacia la extremidad derecha.
19. Los corazones
|| Regresar al Índice ||
El corazón es una bomba que se encarga de
imprimirle movimiento a la sangre a través de los vasos sanguíneos,
especialmente en el lado arterial del sistema circulatorio, pero también le
imprime parte del movimiento a la sangre en el lado venoso por aspiración
“mediante la creación de una presión de succión cuando la sangre abandona la
aurícula o aurículas”. Si el corazón incrementa su ritmo de bombeo, a este
estado se lo conoce como taquicardia, y los valores de la presión sanguínea pueden
incrementarse hasta 5 veces. Si el ritmo del corazón decrece, a este estado se
lo denomina bradicardia, y los valores para la presión sanguínea pueden
disminuir marcadamente. Por ejemplo, las tortugas marinas que se sumergen
pueden disminuir su potencia cardíaca hasta 1/50 de su valor habitual.
En ciertos animales, el corazón también
sirve como un canal que separa a la sangre oxigenada que proviene de los
pulmones, de la desoxigenada que proviene de las venas sistémicas, evitando que
las sangres se mezclen y disminuya la tensión diferencial, lo que en últimas
disminuiría la eficiencia del animal. A diferencia de muchos órganos que vienen
en parejas, el corazón es un órgano único que se ubica en la posición que
debería tener uno de los lóbulos del pulmón izquierdo. En esta sección nos
enfocaremos exclusivamente en los corazones de los vertebrados, aunque
hablaremos algo de los corazones de los cefalocordados, y de cómo pudo haber
evolucionado desde un ancestro sin corazón.
19.1¿Qué es un corazón?
La palabra corazón se utiliza para denotar
a bombas divididas en recamaras internas que se encargan de impulsar a la
sangre circulante a través del circuito circulatorio. Sin embargo, la cuestión
en la anatomía comparada no es tan simple, si bien el modelo biológico al cual
estamos más acostumbrados somos nosotros mismos, donde solo existe un corazón,
en diferentes tipos de vertebrados podemos visualizar la presencia de más
bombas. De este modo, y para los propósitos de este escrito, describiremos como
corazón a cualquier bomba de sangre dividida en cámaras recubierta con tejido
muscular cardíaco. Aun así, también nos encontraremos generalmente con que, el
corazón principal al que estamos acostumbrados recibirá la denominación de
corazón branquial, aunque el animal no posea branquias, esto se debe a que
anatómicamente los corazones se encuentran en la misma zona homologa a la de
los peces, donde el corazón está justo antes de las branquias u arcos
branquiales no funcionales.
19.2 El corazón básico de los vertebrados tiene
cuatro cámaras en serie
Embriológicamente, el corazón de los vertebrados consiste en cuatro cámaras lineales o en serie, estas cámaras se denominan: sinus venosus, auricula “atrium”, ventrículo y el bulbus cordis. La sangre ingresa de la circulación sistémica sin oxígeno al sinus venosus sin oxígeno, y sale por el bulbus cordis también sin oxígeno a la aorta. Se trata de un corazón con forma de tubo al inicio, y alrededor de este, se posiciona una gran cantidad de tejido muscular cardíaco. Posteriormente y dependiendo de la especie, el tubo sufre modificaciones, giros y enroscamientos que dan lugar a las múltiples fisonomías de los corazones de los vertebrados.

Figura 19.1. Estructura del corazón
básico de los vertebrados.
Por lo general, el sinus venosus recoge sangre de dos diferentes tipos de venas, las venas cardinales comunes, que portan la sangre recolectada del sistema circulatorio sistémico, y la vena hepática que regresa del hígado cargada de nutrientes. A pesar de lo anterior, existe una nomenclatura muy laxa a cerca de las cámaras de los corazones de los vertebrados, y de manera clásica solo se denomina cámaras a aquellas divisiones entre las aurículas y los ventrículos, de este modo clásicamente se describe al corazón de los peces como de dos cámaras, aun cuando pueden tener más.

Figura 19.2. El modelo
anterior es un esquema simplificado del corazón básico de los vertebrados (SV)
Sin embargo, el bulbus cordis es solo una
estructura embrional transitoria, cuando se empieza a madurar el ventrículo se
abre paso hacia la aorta de manera directa, por lo que el corazón de los peces
debería ser denominado como de tres cámaras “sinus venosus, auricula y ventrículo”
sin embargo solo se toman dos cámaras “aurícula y ventrículo” para la
nomenclatura en muchos libros de texto. Esto se debe a que la nomenclatura de
las cámaras fue originalmente tomada con respecto al corazón humano, donde el sinus venosus y el bulbus
cordis desaparecen durante el curso del desarrollo embrionario.
👉 Ciclo cardíaco general, diástole auricular y sístole ventricular: Durante el ciclo cardíaco siempre se dan dos movimientos simultáneos en cada mitad del corazón. Durante la diástole auricular se genera un efecto sifón que jala la sangre desde las venas hasta el sinus venosus y la aurícula, estando cerrada la válvula que conduce al ventrículo. Al mismo tiempo el ventrículo y el bulbus cordis realizan su sístole con la cual se empuja la sangre hacia la aorta. En esta fase, la válvula auriculoventricular está cerrada para evitar que la sangre se devuelva a la aurícula y salga toda hacia la aorta.


Figura 19.3. La sístole es
la presión y la diástole es la dilatación.
👉 Ciclo cardíaco general, sístole auricular y diástole ventricular: Durante la sístole auricular, las válvulas del sinus venosus
se cierran para evitar que la sangre se devuelva a las venas y de este modo
fluya completamente a la aurícula y al bulbus cordis. La válvula
auriculoventricular se abre debido a la diástole ventricular lo que permite el
flujo de la sangre desde la aurícula. En este punto la sangre se mueve por un
efecto doble, primero por el empuje de la aurícula y el sinus venosus y segundo por el
efecto succión del ventrículo y el bulbus cordis al expandirse.
En el corazón básico, el sinus venosus
y la aurícula experimentan la diástole cuando el ventrículo y el bulbus cordis
experimentan la sístole en el siguiente paso del ciclo cardíaco, el sinus venosus
y la aurícula experimentan la sístole cuando el ventrículo y el bulbus cordis
experimentan la diástole. Ambas sístole y diástole le imprimen movimiento a la
sangre, la sístole por empuje mecánico mediante una presión positiva, y la
diástole mediaste una succión hacia la cámara vacía o presión negativa.
19.3 ¿Cuántas cámaras tiene un corazón?
En este punto, la confusión puede llegar a ser tremenda, en la tradición escolar en la que me eduque, se nos decía que el corazón de los peces tenía dos cámaras, el de los anfibios tres y el de las aves y mamíferos cuatro, siempre asociando el incremento en la cantidad de cámaras con un mejoramiento hacia la perfección que simbolizábamos los seres humanos.

Figura 19.4. El modelo
anterior resalta con claridad este error conceptual, la tradición con la cual
me eduqué, donde de manera incorrecta se describe al corazón de los peces como
de dos cámaras en serie, cuando en realidad son más. Después fui a la
universidad… pero otras personas no, y se quedaron con esa idea. Una de las
cuestiones básicas, es que, en anatomía comparada, los ventrículos y las
aurículas estarán acompañadas por otras dos cámaras, que pueden estar o no
dependiendo del vertebrado que estemos analizando.
De este modo, la tradición de la anatomía comparada (“aunque no nuestra tradición escolar” señala que el corazón básico de los vertebrados posee cuatro cámaras, sinus venosus, aurícula, ventrículo y bulbus cordis “o conus arteriosus, la razón del cambio de nombres de la última cámara la discutiremos luego” (Bourne, 1980; Fange, 1972; Kardong, 2011, 2014; Randall, 1970; Romer & Parsons, 1962).

Figura 19.5. Los modelos
anteriores representan corazones de algunos peces de linajes primitivos,
"arriba" el de los mixinos y "abajo" el de las lampreas,
ambos revelan arquitectura en serie de los corazones de los peces, con cuatro
cámaras.
Sin embargo, la asignación del estado de
la cámara en muchas ocasiones puede llegar a ser complejo, hasta el punto de
tener confusión acerca del conteo de cámaras en una especie dada. Los peces
mixinos “cuyo corazón cubriremos en detalle en próximos artículos” posee el
corazón de cuatro cámaras básico de los vertebrados en serie y con forma de S.
Sin embargo, aunque las primeras tres cámaras, el sinus venosus, la aurícula y el
ventrículo son fácilmente señalables (Kardong 2002; Pough et al 2002; Randall 1968)
el conus arteriosus puede ser muy
rudimentario y difícil de señalar (Wright et al 1984). Por otro lado, las
lampreas poseen las cuatro cámaras en serie, todas bien definidas (Fange 1972).
Por otra parte, los lagartos de tipo varano, se dice que poseen unos
ventrículos septados de tal forma, que se generan cinco cámaras, dos atrios y
tres ventrículos (Burggren, 1988; Burggren y Johansen, 1982).
Aunque, en las reconstrucciones
tridimensionales de los corazones de los embriones de los ratones muestran un sinus venosus
pobremente desarrollado aunque el miocardio jamás forma una autentica cámara
(Moorman y Christoffels, 2003b), lo cual indica que solo las dos aurículas y
los dos ventrículos son anatómicamente funcionales en los ratones (Christoffels
et al 2000). Las dificultades al determinar la identidad de las cámaras
cardíacas emergen durante la evolución del corazón de los vertebrados. El
sistema circulatorio es uno de los sistemas más flexibles que existen, por lo
que alunas cámaras pueden reducirse, otras pueden unirse, otras avanzar
borrando del paso a las del frente, dividirse en su interior por tabiques
nuevos.
A pesar de ello, eso ha evitado de los
anatomistas y fisiólogos comparativos intenten clasificaras de acuerdo a su
anatomía (Bourne 1980; Fange 1972; Kardong 2002; Randall 1970; Romer 1962) o a
su fisiología (Christoffels et al 2000). Por otro lado, Simões-Costa y
colaboradores (2005) los clasifican de acuerdo al tipo de flujo, entrante o
saliente de la sangre, y es eso lo que veremos a continuación.
19.4 La
alimentación del corazón y los infartos
Como cualquier otro músculo, o mejor, más que cualquier otro musculo, “ya que el musculo del corazón “miocardio” nunca descansa”, requiere de elevadas cantidades de oxígeno y nutrientes, así como la necesidad de eliminar una enorme cantidad de desechos y dióxido de carbono para soportar su metabolismo, y mantenerlo en movimiento. Si el miocardio llegara a agotarse, se genera una isquemia, que es la muerte de su tejido, lo que provoca arritmias o en últimas un infarto general.

Figura
19.6. La
pared interna del miocardio, especialmente la del ventrículo, genera unos conos
que se proyectan al interior denominados trabéculas. La textura resultante luce
esponjosa.
En muchos peces y en los tetrápodos más primitivos, esta demanda es
solucionada mediante un intercambio directo de sustancias entre el miocardio y
la sangre que pasa por su lumen “la cavidad interna”. La pared interna del
miocardio, especialmente la del ventrículo, genera unos conos que se proyectan
al interior denominados trabéculas. La textura resultante luce esponjosa. Las
trabéculas "esa cosa esponjosa de adentro" son estructuras que pueden
verse en una amplia gama de animales vertebrados, mientras que en los animales
más simples apoyan a la alimentación del miocardio, mientras que, en
vertebrados superiores o más derivados, evitan el reflujo de sangre hacia las
aurículas.
Cuando este proceso se hace inefectivo los animales emplean las
arterias coronarias, las cuales atraviesan la pared del corazón, usualmente en
la parte más externa del miocardio. Usualmente se encuentran bien desarrolladas
en los peces elasmobranquios, los cocodrilos, las aves y los mamíferos, en
donde estas suplen la mayor parte de las necesidades del miocardio.
19.5 La fisiología del corazón, ingreso y salida de
sangre
Generalmente, cuando pensamos en el corazón, imaginamos una estructura que funciona toda del mismo modo, como si fuera una sola unidad, al latir imaginamos que todo el corazón se expande al mismo tiempo, y al contraerse, lo imaginamos contraerse todo al mismo tiempo.
Figura 19.7. Arterias
coronarias que alimentan al miocardio de manera externa.
Sin embargo, esta aparente simplicidad puede, y es muy engañosa. Esto se debe a que el corazón en realidad es un ensamblaje de elementos diferenciados, los cuales deben hacer dos cosas: recibir “succionar desde las venas” sangre (flujo entrante) y empujar la sangre por las/la arteria(s) (flujo saliente). Esta dicotomía en el funcionamiento del corazón se expresa de muchas formas, a nivel anatómico, fisiológico, de desarrollo y genético. Los compartimentos para el flujo entrante y el flujo saliente son distinguibles el corazón de cámaras lineales de los peces, así como el de los anfibios y amniotas. De este modo, todos incluido el corazón humano, tienen una estructura en forma de S, donde se tienen a las caparas de flujo entrante “sinus venosus y/o aurículas” en un nivel superior al que se encuentran as caparas de flujo saliente “bulbus cordis y/o ventrículos”.
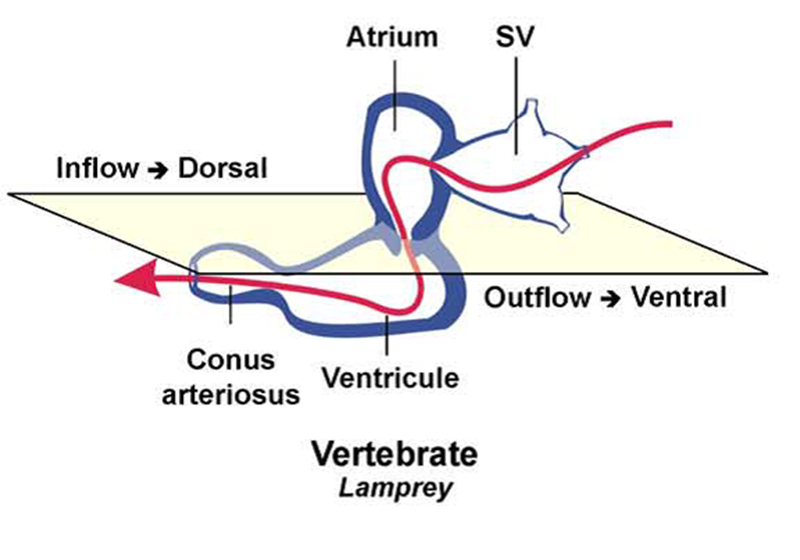
Figura 19.8. Corazón en forma de S de los
peces sin mandíbula.
El corazón de la lamprea “Figura 19.8” es
un perfecto ejemplo de corazón en S, donde el flujo entrante al mismo tiempo en
que rebota "en la imagen contra la pared izquierda de la aurícula cae, evitando que se dé un reflujo hacia el sinus venosus
y a las venas. Esta es una adaptación eminentemente funcional para evitar el
reflujo de sangre producto de su choque contra las paredes del corazón, si el
corazón fuera lineal de manera perfecta, cuando la sangre chocase contra la
pared o la válvula cerrada, debido a la tercera ley de Newton, la sangre
rebotaría, creando un reflujo que enviaría sangre de regreso hacia las venas,
creando problemas circulatorios. Sin embargo, resulta curioso que las bombas
“corazones” de los cefalocordados y urocordados si son lineales (Moller y
Philpott 1973), aunque en estos animales, la presión sanguínea es tan baja, que
el rebote seria mínimo, por lo que no requerirían la estructura en forma de S.
Todos los corazones de los animales
vertebrados "que incluye de momento a los mixinos o hagfish aunque no
tienen vertebras" poseen una organización de cámaras por niveles para
evitar los reflujos. Tal como nos lo muestra el corazón de un caracol, si las
cámaras están de forma lineal perfecta, se crea un contraflujo lineal debido a
la tercera ley de Newton. Para evita
esto, los corazones de los vertebrados están en forma de S, lo cual crea una
caída natural, cuando la sangre golpea la pared del corazón y rebota, ya no
fluye contra la vena de ingreso, sino contra otra pared, pues ya ha caído un
poco debido a la gravedad, lo cual evita o reduce el riesgo de contracorrientes
que disminuirían la eficiencia del circuito circulatorio.
El movimiento general del corazón también se divide en dos tipos, la diástole, que es la expansión de una cavidad generalmente succiona líquido de una vena, por lo cual es un movimiento que apoya al flujo entrante, mientras que la sístole expulsa la sangre, empujándola hacia otra cavidad o hacia una arteria. La distinción en el desarrollo entre las funciones de flujo entrante y saliente es aparente en los mapas de destino de tejidos, donde las células precursoras ocupan diferentes secciones en el embrión y en el primordio del corazón. Incluso a nivel molecular, existen genes que se activan para uno u otro tipo de flujo, función y estructura (Bruneau 2002; Moorman y Christoffels 2003a; Xavier-Neto et al 2001; Christoffels et al 2000).
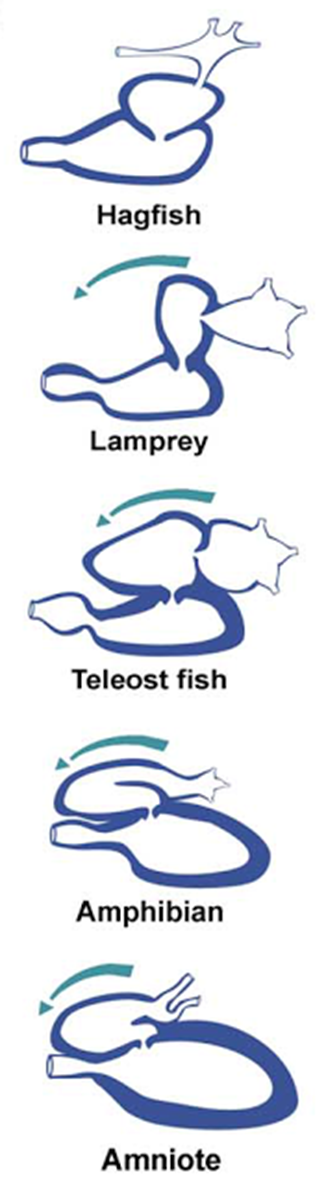
Figura 19.9. Vista lateral
de los corazones de los vertebrados.
Estos descubrimientos son evidencia
persuasiva de que la segmentación del corazón refleja un principio
morfogenético subyacente que divide a las bombas circulatorias más avanzadas en
unidades especializadas para los flujos saliente y entrante de la sangre. En
los vertebrados, este plan morfogenético incluye cambios en la posición
relativa de las unidades. Por lo tanto, los corazones de los vertebrados poseen
un nivel de sofisticación mecánica que mejora de manera significativa la
circulación sanguínea. Usando este criterio (Simoes-Costa 2005) la aparente
diversidad de corazones, se vuelve una variación pequeña de un tema repetitivo
de unidades de recepción y salida de sangre.
19.6 Anatomía y fisiología general de los corazones
En los peces las arterias coronarias se derivan arcos que se derivan de las branquias, los cuales cargan sangre recientemente oxigenada. Una vez se da el intercambio de gases a través de los capilares del miocardio, las venas coronarias ingresan en el sinus venosus. En adición a las válvulas conales, el miocardio desarrolla una serie de válvulas entre las cámaras. Las válvulas sinoauriculares se forman entre el sinus venosus y la aurícula u la válvula auriculoventricular se forma entre la aurícula y el ventrículo (es homologa a las válvulas bicúspide y tricúspide de los mamíferos, aves y cocodrilianos).

Figura 19.10. El corazón de
los caracoles escargot es lineal y posee el problema teórico que se anticipa si
la válvula de salida está al mismo nivel del de la válvula de entrada, la
sangre rebota disminuyendo la eficiencia del corazón. En las lampreas y demás vertebrados,
este problema se soluciona dejando las válvulas en diferentes niveles cada una,
las venosas arriba, la auriculoventricular en medio y la arterial abajo.
Durante el flujo normal de sangre, las
válvulas se abren, cuando la cámara experimenta la diástole succionando la
sangre a su interior, y se cierran cuando la cámara experimenta la sístole,
para evitar el flujo retrogrado de sangre cuando la cámara presiona. A su vez,
el corazón yace en una cavidad llamada cavidad de pericardio, la cual está delineada
por una membrana de tejido epitelial “el pericardio”. En muchos peces, la
cavidad pericárdica yace en un marco de hueso o cartílago, formando un
compartimiento semirrígido que mantiene al corazón en su posición. La
contracción secuencial de las cámaras del corazón mueve la sangre desde una
cámara a la siguiente y finalmente hacia la aorta ventral.
Como ya hablamos en otra oportunidad, la
presión de las venas depende de la succión del corazón y de los músculos
adyacentes a las venas, así como de las válvulas que evitan que la sangre en
las venas se devuelva, esto se debe a que cuando la sangre pasa por los
capilares pierde casi la totalidad de la fuerza que le impartió la sístole del
ventrículo, y al salir deberá moverse por la succión/diástole de la aurícula y
de los músculos adyacentes.
19.7 Clasificación y nomenclatura de los corazones
usada más comúnmente
Las aves y los mamíferos tienen corazones
de cuatro cámaras, pero de las cuatro cámaras del corazón básico de los peces,
solo dos persisten como los compartimientos de recepción y emisión de sangre,
la aurícula y el ventrículo. Por otra parte, el corazón de las aves y los
mamíferos funcionan más como dos corazones separados por una membrana. A pesar
de que el corazón de las aves y los mamíferos son ambos derivados evolutivos de
tetrápodos ancestrales, ellos se desarrollaron de manera independiente de
diferentes ancestros, lo que se denomina como evolución paralela.
Filogenéticamente, los anfibios y reptiles
vivientes permanecen entre los tetrápodos derivados y los peces, una posición
que ha seducido a muchos a creer que estos son ejemplos de intermediarios en lo
que debió ser la evolución de los corazones de aves y mamíferos. Aunque
ciertamente, la ruta evolutiva de los corazones de las aves y los mamíferos
pasó a través de los tetrápodos ancestrales, se debe recordar que los anfibios
y reptiles modernos están tan alejados en el tiempo de dichos tetrápodos como
los mamíferos y las aves.
Inclusive, la diversidad de los sistemas
circulatorios y de los corazones de los anfibios y los reptiles modernos es
mucho más grande de lo que aprendemos generalmente en la escuela, para citar
dos ejemplos: en los anfibios nos enseñan el corazón tipo de las ranas que es
tricameral, sin embargo, no recordamos la advertencia, o no se nos dice que,
existen otros corazones en los anfibios, por ejemplo, los del genero de
salamandras Siren spp., que es de cuatro cámaras. Otro ejemplo, esta vez en los
reptiles, los cuales son generalmente olvidados en nuestras clases de anatomía
en el colegio, la diversidad es grande, por ejemplo, los varanos monitores
poseen corazones con 5 cámaras.
19.8 La
contracción y el musculo cardíaco
La contracción es una propiedad intrínseca del musculo cardíaco. Las células individuales muestran inclusive una contracción rítmica si son aisladas fuera del cuerpo en un medio de cultivo adecuado. Las células cardíacas tienden a moverse de manera sincronizada, pero la contracción del miocardio completo usualmente comienza para los peces, en una región específica en el sinus venosus/aurícula denominada marcapaso o nódulo sinoauricular, posteriormente disparándose a través de un sistema conductor nervioso a través del ventrículo y otras regiones contráctiles. Adicional al nódulo sinoauricular existe un segundo nódulo denominado auriculoventricular en la pared del corazón.

Figura 19.11. La
clasificación y la nomenclatura más usada para los corazones se resume en esta
imagen, la cual, a pesar de dar una idea general e introductoria posee tantos
errores que harían sonrojar a cualquier anatomista comparativo de vertebrados.
Para empezar, se representa al corazón de los peces como de dos cámaras,
pasando al anfibio de tres y al mamífero de cuatro, como si hubiera una
secuencia evolutiva/platónica de incremento en la cantidad de cámaras, lo cual
es totalmente incorrecto. Los peces poseen cuatro cámaras lineales, mientras
que los mamíferos las tienen en paralelo "siendo las cuatro de los
mamíferos derivadas de solo dos del corazón de los peces". Existen otros
errores, pero lo discutiremos en artículos futuros.
En la ventrícula (atrium) izquierda se encuentra el marcapaso natural o nódulo sinoauricular, que es donde se inicia el latido del corazón, los marcapasos artificiales tratan de solventar problemas que pueden surgir en esta área del corazón. El nódulo auriculoventricular consiste en fibras de Purkinje, fibras similares a las neuronales, las cuales realmente son fibras musculares modificadas. Las fibras de Purkinje salen del nódulo auriculoventricular, dividiéndose en racimos izquierdos y derechos, los cuales viajan en el tabique interventricular hasta la punta del corazón, para luego devolverse a través de su respectivo lado del corazón. La velocidad a la cual el corazón late esta inicialmente bajo la influencia del sistema nervioso y el sistema endocrino. El ritmo cardiaco también responde a la velocidad de llenado de la sangre.

Figura 19.12. Anatomía
general del corazón de los mamíferos donde se indica por medio de un corte, sus
cámaras paralelas (YouTube)
(YouTube) (YouTube).
Palpitar normal del corazón, el sonido
doble se debe a que la sístole auricular y la sístole ventricular no son
eventos simultáneos, se dan de manera rítmica, primero una y luego la otra.
Durante el ejercicio, la cantidad de
sangre venosa que regresa al corazón se incrementa en parte debido al
incremento de la presión impartida por los músculos “como ya hemos dicho, las
venas son independientes a la sístole del corazón, por lo que para que la
sangre se mueva dependen de la presión de la diástole que es menor, y por lo
tanto dependen de la presión impartida por los músculos”. Cuando una mayor cantidad de sangre llega al
corazón, este se estira, y como si fuera un caucho, esto hace que almacene más
energía, por lo que el siguiente latido es mucho más potente. A este fenómeno
se le denomina reflejo de Frank-Starling.
19.9 El
corazón de los tunicados
De acuerdo a numerosos estudios, los tunicados son los animales más cercanamente emparentados a los vertebrados terrestres, aunque eso es verdad en términos genéticos. Sin embargo, su curso evolutivo no fue paralelo al de los vertebrados y cefalocordados, en lugar de ello regresaron a un modo de vida sésil en el adulto, aunque ciertas estructuras, como por ejemplo el corazón (Simoes-Costa 2005). Como ya lo mencionamos en una oportunidad anterior, el sistema circulatorio de los tunicados es abierto, el cual consiste en una bomba ventral unida a vasos sanguíneos endoteliales muy extendidos, en realidad la apertura es bastante pequeña en comparación con la extensión de los vasos (Satoh 1994). En los tunicados la bomba “pseudo-corazón” consiste de un tubo con una capa simple de tejido muscular cardiaco rodeado de un celoma y un pericardio. Es un corazón con una sola cámara, sin válvulas internas o una polaridad distintiva (Simoes-Costa 2005).

Figura 19.13. Sistema
circulatorio de un tunicado.
El sistema circulatorio de un tunicado es
bastante similar al de un cordado, sin embargo, se abre en la vesícula
gástrica, lo cual en los cordados es asumido por una red capilar. Por lo demás
posee un corazón que se ubica justo antes de los arcos branquiales. A pesar de
esta semejanza fundamental, existían otras diferencias que hicieron pensar por
muchos años que este corazón no era homólogo al de los cordados, hasta que se
encontró evidencia genética de lo opuesto. Sin embargo, en los estadios de
desarrollo iniciales existe una distinción del eje dorsal y ventral, y en
muchos tunicados la bomba adquiere una forma de V. Una de las habilidades más
extrañas del pseudo-corazón de los tunicados es la capacidad de revertir la
perístalsis, de modo tal que el flujo de sangre puede revertirse (Simoes-Costa
2005).Por mucho tiempo se pensó que el corazón de los tunicados no era homólogo
al corazón de los cefalocordados y craneados debido a esta y otras diferencias
anatómicas, sin embargo, análisis genéticos han demostrado que muchos de los
genes que regulan la construcción de todos esos órganos son los mismos, lo cual
revela su naturaleza fundamentalmente homóloga (Simoes-Costa 2005).
19.10 El corazón de anfioxos
Filogenéticamente, el corazón probablemente comenzó como un vaso sanguíneo de tipo venoso que se ensanchó en el interior de alguna zona del sistema cardiovascular. Evidencia de esto es que los vasos sanguíneos venosos poseen válvulas de cierre hermético unidireccionales, tal como las que tiene el corazón. Embriológicamente, todos los corazones comienzan como un tubo “homologo a un vaso sanguíneo venoso” sobre el cual se posicionan los primordios del tejido muscular cardíaco que con el tiempo generan el miocardio. En los seres humanos este estadio de tubo se genera cerca de la tercera semana de gestación. En términos comparativos en cierto punto del desarrollo, todos los corazones de los vertebrados son en esencia indistinguibles morfológicamente (Fishman and Chien, 1997). Si nos enfocamos en anfioxos nos encontramos con un caso interesante, ya que este animal posee una gran cantidad de vasos sanguíneos contráctiles.

Figura 19.14. El sistema
circulatorio de anfioxos.
Otros autores (Fishman y Chien, 1997;
Fishman y Olson, 1997; Moorman y Christoffels, 2003a) han discutido las
diferencias entre el corazón de los craneados y la hipotética bomba de sangre
del ancestro común de los craneados y cefalocordados “el ancestro común de
todos los cordados”. Usando el corazón de anfioxos como una referencia, se
pueden identificar novedades evolutivas como la formación de tabiques y
válvulas, las cuales dan origen a las cámaras; así como al endocardio,
epicardio, circulación coronaria “en esta aun algunos peces como los mixinos no
la poseen”, capa interna de endotelio y sistema eléctrico. Ninguna de estas
características es observada en anfioxos, por lo cual existe mérito en asumir
que, el corazón del ancestro común de todos los cordados tendría un
pseudo-corazón muy simple, similar al de anfioxos.
Sin embargo, hay que ser precavidos, el
anfioxo moderno ha tenido tanto tiempo para evolucionar como el corazón
penta-cameral de un varano o el corazón tetra-cameral de un ave o un mamífero.
Debido a varias características del modo de vida del anfioxos moderno como la carencia
de aletas “como en paleobranchiostoma”, y una vida sedentaria; además de la
carencia de ciertas características que si están presente en los urocordado
“tunicados” se puede sugerir que el anfioxos moderno es una forma relativamente
derivada y simplificada de los cefalocordados más primitivos y cercanos al
ancestro común de todos los cordados (Blieck, 1992; Hirakow, 1985; Kozmik et
al., 1999; Mallatt and Chen, 2003; Randall and Davie, 1980; Conway Morris,
2000). Muchas características del sistema circulatorio de anfioxos son
rudimentarias. Tales características son compartidas con deuterostomos basales
como los gusanos hemicordados y los equinodermos holotúridos, e inclusive
protostomos como los anélidos, foronidos lofoforonidos y equiuranidos (Brusca and
Brusca, 2003)
La circulación en anfioxos es cerrada,
mostrando las siguientes características:
- Vasos sanguíneos ventrales y dorsales
- Canales vasculares agrupados alrededor
del intestino “¿porta?” y la faringe “¿branquias?”
- Carencia de una cubierta endotelial
continua
- Múltiples vasos sanguíneos engrosados y
contráctiles “pseudo-corazones”
Las múltiples bombas de anfioxos no son
fácilmente homologables al corazón de los craneados (Carter, 1967; Jefferies,
1986; Moller and Philpott, 1973; Rahr, 1979; Randall and Davie, 1980). Los
pseudo-corazones de anfioxos son vasos sanguíneos peristálticos que se
encuentran en las venas hepáticas y subintestinales, así como en la arteria
endostiliar. Fisiológicamente, todos, uno o ninguno de estos pseudo-corazones
es homologo al corazón de los craneados, sin embargo, en esos términos es
virtualmente imposible detectar cual es la respuesta verdadera, por lo que solo
resta los patrones de expresión genética durante el desarrollo, aquel vaso
sanguíneo que comparta los mismos genes y el mismo patrón de desarrollo con el
corazón de los craneados será sin lugar a dudas el pseudo-corazón homólogo.
En base a los análisis de Holland y
colaboradores (2003) se ha determinado que el vaso sanguíneo que primero se
desarrolla es el subintestinal, y que de hecho este le da origen a todos los
demás vasos sanguíneos contráctiles que se presentan en la red hepática. Estas
evidencias nos indican que probablemente la bomba subintestinal es el homologo
al pseudo-corazón de los tunicados y al corazón de los craneados. Otro detalle
es la naturaleza del sinus venosus en anfioxos, el cual se coloca en la
confluencia de las venas principales “hepática y cardinal”, aunque su estado
como una cámara cardíaca es controversial, más aún porque no late (Simoes-Costa
et al. 2005). Existe aún mucha
controversia sobre la naturaleza del sinus venosus de anfioxos y sobre si
realmente es homólogo al de los craneados.
Los análisis del sistema circulatorio de
los tunicados y de los cefalocordados indican características comunes también
compartidas por los carneados. Todos los cordados colocan su bomba de empuje
principal justo antes de los arcos branquiales de modo tal que la sangre que
fluye a través de ellos sale empujada hacia las branquias. Sin embargo, en los
tunicados, la perístalsis se puede invertir, así que con excepción de ellos,
todos los cordados poseen un circuito circulatorio que se extiende por la zona
dorsal y la zona ventral, la sangre que fluye por el dorso va desde la cabeza
hasta la cola, y la que fluye por el vientre va desde la cola a la cabeza.
Esto contrasta con los anélidos y otros
animales semejantes e que la dirección es inversa (Brusca y Brusca, 2003). Aun
con la versatilidad en el flujo de sangre, los tunicados se asemejan a los craneados
en que su bomba principal se ubica justo en frente de la convergencia de las
venas principales y justo detrás de los arcos branquiales. Otra similaridad es
que la bomba de sangre principal se encuentra resguardada de una cobertura
especial o pericardio (Percy y Potter, 1991; Schmidt-Nielsen, 1990). Aunque, la
circulación en los tunicados es abierta más que cerrada.
Aunque la circulación de los
cefalocordados es cerrada, y la red de vasos sanguíneos es fácilmente
homologable a los vasos sanguíneos principales de los craneados, el problema
que enfrentamos es, que el empuje no está centralizado, en lugar de poseer una
sola bomba de empuje posterior a la convergencia de las venas principales y
anterior a los arcos branquiales, posee una serie de bombas distribuidas
principalmente en la red porta/hepática y en la zona intestinal. (Moller y
Philpott, 1973; Rahr, 1979). Aun así, basándose en la estructura del cuerpo
indica que anfioxos se asemeja mucho a los deutorostomos ancestrales como
pikaia (Hirakow, 1985; Rahr, 1981). En resumen, ambos, los tunicados y los
cefalocordados poseen ciertos caracteres que se comparten con los craneados y
otros que no, pero ninguno representa perfectamente una idea consistente con lo
que debería ser el plano original del sistema circulatorio de los vertebrados,
aunque, en muchos sentidos son mucho más simples.
20. El pensamiento darwiniano y platónico
|| Regresar al Índice ||
La historia del pensamiento evolutivo ha
sido un camina largo y tortuoso, y no es que, después de que la publicación de
articulo original de Darwin y Wallace todos los biólogos pasaran de un
pensamiento, digamos que no creacionista, pero si evidentemente platónico a un
pensamiento evolutivo de corte darwiniano. Ha sido un proceso álgido, lento y
gradual; tanto así que muchas afirmaciones que se presentan como darwinistas en
realidad son platónicas. Una de esas ideas platónicas disfrazadas de
darwinistas, y que influyeron, influyen, y péseme decir, influirán aun por
décadas al entendimiento básico del sistema cardiovascular de los vertebrados
es la idea de “la perfección”.
La perfección es una idea platónica, al igual que la superioridad; en la escala natura expuesta por platón en el Timeo, en que la variabilidad de la vida se expresa como una escalera, en la que el hombre se encuentra en la punta como la criatura más perfecta, solo siendo excedido por los dioses o por Dios y los ángeles “dependía de la cosmovisión”. De este modo, los animales inmediatamente más parecidos a nosotros descendían ciertos escalones en esta escala natural. De lo anterior se desprendía que los mamíferos eran superiores a las aves, estas a los reptiles, estos a los anfibios, estos a los peces, estos a los invertebrados, estos a las plantas y estos a los minerales. Como siempre una idea fuertemente platónica. En el estudio de los corazones, a mí, y a los estudiantes de mi generación durante el bachiller se nos inculcó esta idea platónica gracias a los libros de texto, donde se decía que el corazón óptimo, el más perfecto, era el corazón de cuatro cámaras de los mamíferos y las aves. El siguiente era el corazón de los reptiles y anfibios con tres cámaras y por ultimo era el corazón de los peces de dos cámaras.

Figura 20.1. La escala
natural, expresada por Platón en el Timeo y después abrazada por el
cristianismo ha sido una de las ideas más influyentes en el entendimiento de la
biología a lo largo de casi más de dos milenios. Peor aún es que su iconografía
impregna aun el imaginario colectivo, de hecho, expresiones como "la
cúspide de la cadena evolutiva", son en realidad ideas de platónicas y no
evolutivas.
Errores por todas partes,
· los peces tienen más de dos cámaras en sus corazones
· los anfibios pueden separar sus flujos sanguíneos, debido a la
diferencia de densidad de las sangres con y sin oxígeno.
· solo muestran tres opciones, y la diversidad de los sistemas
cardíacos
· da la falsa impresión de progreso, donde a mas cámaras más
"perfecto" es el corazón, una idea platónica que nos cuesta dejar
atrás, aun en imágenes tan difundidas como esta.
Más allá del feo descache de representar
los corazones como no son “como en los peces” de omitir las ambigüedades “peces
pulmonados” o la diversidad “corazón cocodriliano”, detalles del funcionamiento
“como en las semi-cámaras del corazón de los lagartos” que es adjudicable a la
transposición didáctica, debemos hablar de una tendencia que si afectó a la
ciencia real. Y es que muchas afirmaciones que se consagraron darwinistas aun
cargaban de un rezago platónico. Para los fisiólogos y anatomistas originales,
que aun cargaban con una carga fuertemente platónica, resultó tentador medir el
desempeño de los corazones de los vertebrados, en como su sistema
cardiovascular se asemejaba a las necesidades del corazón de los mamíferos, que
era después de todo el corazón de los humanos, el pináculo de la “evolución”
(resaltar que pináculo de la evolución es paradójicamente una idea
anti-darwiniana y más bien platónica)
Por ejemplo, los tabiques cardíacos
parciales han sido denominados como “incompletos” en comparación a las
divisiones anatómicas completas del corazón de los mamíferos. Los corazones y
arcos aórticos de vertebrados “inferiores” fueron interpretados como diseños
“imperfectos” debido a que el diseño mamífero era considerado óptimo
“nuevamente una idea platónica sobre el ideal de pináculo, y perfección”. Tal
como hemos visto en artículos anteriores, los peces pulmonados como los
anfibios y reptiles no cocodrilianos se pensó originalmente que ocurría una
mezcla significativa de sangre oxigenada y desoxigenada. Obviamente una premisa
como esa es fácil llegar a la conclusión de que estos animales no podrían
sostener actividades metabólicas muy exigentes debido a la disminución de la
concentración de oxigeno durante la mezcla de sangre. Ciertamente la mezcla de
sangres sería un diseño problemático; sin embargo, el trabajo experimental,
tanto en los peces pulmonados como en los reptiles cocodrilianos ha llevado a
concluir que eso no sucede en la realidad. Las ideas de evolución y progreso
están tan inextricablemente integradas, que pareciera que cuando se critica una
se critica la otra, o que si se disiente de una se diciente de la otra. Nada
más falso, progreso y evolución no son solo ideas que no se deben mezclar, son
de hecho ideas contrarias; el progreso tiene un sentido lineal; la evolución
no, su sentido es de una amplificación de diversidad en muchas, muchísimas
ramas, especializada en nichos diferentes, pero todas aptas para las
condiciones específicas en las que se desarrollan.
Para los que estén familiarizados con la
obra de Sagan sabrán que este despreciaba el enfoque platónico del
conocimiento, el cual favorece solo las ideas y desprecia la realidad sensible.
Este servidor la desprecia aún más por el hecho de que por más que lidiamos su
influencia nos alcanza de maneras tan inadvertidas y que peor aún son
calificadas de afirmaciones basadas en la teoría de la evolución. Lo que observamos al revisar el sistema
cardíaco de los vertebrados es precisamente esta segunda idea, una idea no
progresiva, no lineal; donde cada estructura fisiológica actual es igual de
apta a todas las demás que han sobrevivido hasta el día de hoy. Por ejemplo, la
desviación cardíaca en los reptiles ayuda a prevenir el edema “perdida de
energía y oxígeno, con reingreso del peligroso dióxido de carbono, cuando la
sangre completa el circuito circulatorio pulmonar cuando este no contiene
oxigeno suficiente” solo es posible gracias a que
· el corazón no posee el tabique interventricular o
· los arcos arteriales poseen un punto de contacto. Ambos aspectos
que desde el punto de vista de un mamífero son o imperfectos o
subdesarrollados.
De lo anterior traducimos, que, ningún
corazón moderno debería verse como un intermedio evolutivo, todos han poseído
el mismo tiempo para evolucionar, y dada la gran variabilidad del sistema
circulatorio resulta muy complejo tratar de señalar los caracteres ancestrales,
pero en algunos es posible como el sinus venosus y el bulbus cordis.
21. Sistema circulatorio de los peces agnatostomos
|| Regresar al Índice ||
Los peces abarcan la mayor variabilidad
dentro de los vertebrados, después de todo son los más viejos, en consecuencia,
poseen la gama más diversa de arquitecturas del sistema circulatorio como vimos
en capítulos anteriores. Ahora nos enfocaremos en la estructura de sus
corazones, la cual compagina con la diversidad de sus sistemas circulatorios
generales.
21.1 Arquitectura del sistema circulatorio de los
peces
La mayoría de los peces que mencionamos aquí posee una arquitectura simple con un solo circuito circulatorio que discurre del corazón a los capilares respiratorios en las branquias, de allí a los capilares sistémicos de los órganos y de allí de regreso al corazón.

Figura 21.1. Los peces
estrictamente acuáticos poseen un sistema circulatorio simple, con branquias y
capilares sistémicos.
El esquema anterior muera las zonas de
intercambio de gases, y principalmente, el hecho de que los corazones de los
peces generalmente no lidian con el problema de impulsar sangres con contenido
gaseoso diferente, lo cual altera también sus densidades.
21.2 Mixinos
El corazón de los peces mixinos Figura 21.2 no
posee la cuarta válvula por lo cual lo definiremos como un corazón de tres
cámaras, sinus
venosus que recolecta la sangre venosa, la aurícula en medio y el
ventrículo encargado de despedir la sangre hacia la única arteria que está
conectada al corazón, la aorta. Sin embargo, su sistema cardiovascular posee
ciertas sorpresas. Como todos los corazones de los vertebrados, el corazón de
los mixinos permanece en la parte anterior del animal “frente” y dispuesto
hacia la zona ventral; está compuesto por un musculo cardiaco “miocardio”, y se
encuentra conectado a las venas principales del cuerpo “la vena hepática y la
vena cardina” las cuales confluyen en una cámara llamada sinus venosus.
De manera clara incluye tres cámaras en serie, el sinus venosus que recolecta la sangre venosa de la vena hepática y la vena cardinal; luego vienen la aurícula “atrium” y el ventrículo. El bulbus cordis es muy pequeño en estos peces, y existe gran debate si el corazón de estos peces seria tri-cameral asumiendo que el bulbus cordis está completamente reducido, o de si es tetra-cameral. Sin embargo, el consenso actual es que falta la cuarta cámara y por lo tanto el corazón en los mixinos es de tres cámaras en serie.

Figura 21.2. Modelo del corazón de cuatro
cámaras lineales.
El circuito circulatorio de un mixino “en detalle Figura 21.3” mostrado de manera muy esquemática las esferas negras son las bombas de sangre, las dos más grandes son corazones con musculo cardíaco y las pequeñas son bombas con musculo liso. El corazón homologo al resto de los peces es el corazón branquial que está justo antes de los arcos branquiales "gills".

Figura 21.3. Los mixinos se caracterizan por
poseer más de un corazón.
El sinus venosus tiene conexión a
tres venas, dos cardinales que recolectan la sangre de los tejidos sistémicos y
una hepática que regresa del hígado con la mayoría de los nutrientes. El flujo
de sangre en estos sistemas circulatorios va desde la cola a las branquias en
la zona ventral y desde las branquias hacia el resto del sistema en la zona
dorsal. Esto quiere decir que cuando la sangre ingresa al corazón en la zona
ventral, esta es emitida hacia los arcos branquiales, los cuales conectan los
vasos dorsales con los ventrales. No existen redes nerviosas especializadas que
accionen el latido del corazón de los mixinos, y, por lo tanto, el movimiento
del corazón depende del rebote elástico del corazón cuando se llena de sangre
“reflejo de Frank-Starling”, que genera una contracción.
El corazón de la vena porta en los mixinos
tiene derecho a denominarse como tal debido a la presencia de musculo cardíaco
en sus paredes. Aunque en secciones pasadas describimos al corazón de los
cefalocordados como de varias bombas no homologables al corazón de los
craneados, en los mixinos también se da el detalle de la presencia de corazones
secundarios. De este modo tenemos la presencia de un corazón branquial, el cual
es homologo a los demás corazones de los vertebrados, y de corazones accesorios
o de bombas de sangre accesorias debido a que “al igual que con los
cefalocordados” estas bombas de sangre carecen de un músculo cardíaco.
Existente tres tipos de corazones accesorios en los mixinos. Los corazones
cardinales que se encuentran en las venas cardinales y que impulsan la sangre
resolviendo en parte el problema de la baja presión sanguínea en la zona venosa
del circuito circulatorio de los vertebrados.
Los corazones caudales se ubican en la
cola y cumplen una función similar a la del corazón cardinal, impulsar a la
sangre venosa de baja presión al salir de la red capilar. El corazón porta es
especial, esto se debe a que la sangre en el sistema porta atraviesa dos redes
capilares la primera al ingresar a los intestinos, allí se generan capilares y
la sangre pierde presión, luego la sangre sale de allí por el sistema porta al
hígado donde debe pasar por una segunda red capilar. El corazón porta se
encuentra entre esas dos redes capilares para incrementar la presión de la vena
porta. A diferencia de los demás corazones accesorios, el corazón porta si
posee musculo cardíaco, aunque son los mixinos los únicos peces en poseer este
órgano entre los vertebrados “aunque los cefalocordados también lo poseen”.
21.3 Lampreas
El corazón de las lampreas incluye al
igual que con las mixinos tres cámaras fácilmente identificables, sinus venosus,
aurícula y ventrículo con las mismas funciones y conexiones, pero en contraste
con los mixinos su corazón posee estrías en la parte posterior del ventrículo,
lo que desemboca en una pequeña “cámara”, lo que recibe el nombre genérico de bulbus cordis
en las lampreas posee el nombre especifico de bulbo arterial, lo cual implica
precisamente eso, un bulbo en la base de la arteria aorta.

Figura 21.4. Al igual de en
los cefalocordados y en los agnatos “sin mandíbulas” como los mixinos, las
lampreas poseen un corazón principal y varias bombas de sangre accesorias. Sin
embargo, nos enfocaremos en el corazón principal homólogo al resto de los
corazones de los vertebrados, el corazón branquial que se encuentra justo antes
de los arcos branquiales. En este caso si digo vertebrado, porque a diferencia
de lo que ocurre en los cefalocordados y en los mixinos, en las lampreas el
notocordio se desarrolla rodeando a la medula, para luego segmentarse en discos
que protegen al sistema nervioso conocido como vertebras.
El corazón de las lampreas, a diferencia del corazón de los mixinos, la parte final del ventrículo presenta pliegues que guían la sangre en su curso a la aorta, el desarrollo posterior de estas válvulas es de especial importancia en los peces dipnoi. Las paredes del bulbo arterial carecen de músculo cardíaco pero posee una buena cantidad de fibras de musculo liso organizadas longitudinalmente y circunferencialmente. Válvulas de una sola vía “como las de las venas” separan las cámaras previniendo el flujo retrogrado de sangre, lo cual sumado a la forma en S del corazón hace que el flujo de sangre sea unidireccional idealmente.

Figura 21.5. Corazón en S de
las lampreas.
22. Sistema circulatorio de los peces gnatostomos
|| Regresar al Índice ||
22.1 Cartilaginosos
Los corazones de los condrictios “peces
cartilaginosos” y de los teleósteos “peces óseos” poseen cuatro cámaras
básicas, el sinus
venosus, la aurícula, el ventrículo y el cono arterial/bulbo
arterial. Todos con válvulas de un solo sentido como en las venas.

Figura 22.1. Los tiburones no poseen vejigas natatorias, por lo que nos sirven para mostrar la arquitectura de un sistema circulatorio básico de los vertebrados.

Figura 22.2. El corazón en S
de los peces óseos estrictamente acuáticos.
Aquí explicaremos cierta nomenclatura confusa, el nombre genérico bulbus cordis lo usamos para referirnos a la cuarta cámara que se ubica de manera posterior al ventrículo, sin embargo, este a su vez recibe nombres específicos diferentes dependiendo de la presencia o ausencia de cierto tipo de tejido muscular en las paredes de la cámara.

Figura 22.3. El ciclo
cardíaco de los peces. (a) finaliza la diástole auricular las cámaras se
distienden al máximo llenándose la aurícula y el sinus
venosus de sangre. (b) sístole auricular y diástole
ventricular, la sangre sale de la aurícula y llena el ventrículo. (c) comienza
nuevamente la diástole auricular donde el sinus
venosus succiona sangre de las venas mientras que la
aurícula aún está vacía y también la sístole ventricular con el cerrado de la
válvula auriculoventricular. (d) sístole ventricular completa, la sangre sale
del ventrículo hacia la aorta mientras que continua la diástole auricular
comenzando a llenar la aurícula.
22.2 Peses
óseos no pulmonados
El corazón de los peces óseos no dipnoos,
si le pusieron una tarea de dibujar el corazón de un pez, el modelo anterior es
probablemente el que usted está buscando, después de todo en la actualidad la
mayoría de los peces son óseos con circuitos circulatorios simples. Si la pared de la cámara posee musculo
cardíaco se la denomina cono arterial, y este se contrae junto con el
ventrículo durante la sístole para empujar la sangre hacia la aorta, y se
expande junto con el ventrículo durante la sístole para capturar sangre desde
la aurícula. Si por el contrario la pared posee musculo liso, lo llamamos bulbo
arterial y no se contrae. En los conductos encontramos el cono arterial,
mientras que en los teleósteos encontramos ambos tipos de cámara, el bulbo
arterial y el cono arterial. Ambos corazones poseen pliegues en la parte
posterior de sus ventrículos, ya sea en el cono arterial o en el bulbo
arterial, muy importantes cuando estamos tratando con peces que poseen
mecanismos accesorios de respiración a parte de las branquias. Esto se verá de
manera patente tanto en los dipnoos como en los anfibios.
Muchos peces óseos poseen algo llamado vejiga natatoria, lo cual modifica el esquema general del circuito circulatorio además de generar complicaciones a la hora de representar el contenido de gases de la sangre.
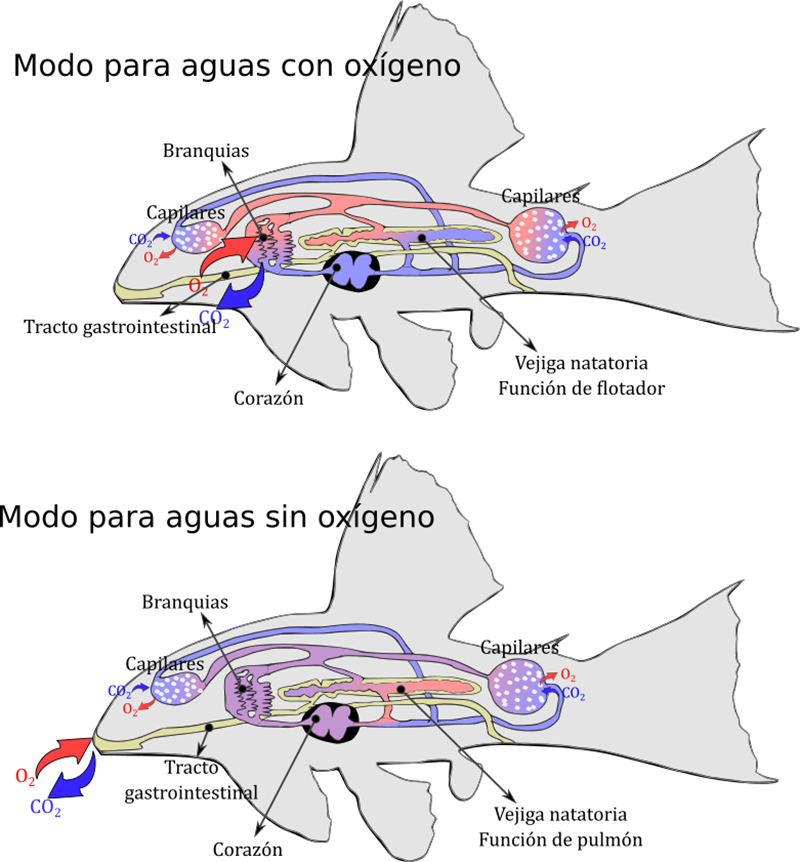

Figura 22.4. Arquitectura de un sistema
circulatorio de un pez que posee una vejiga natatoria/pulmón en la cual se
desvían gases metabólicos según requiera.
Algunos peces han perdido esta capacidad facultativa y han especializado
la desviación y la bolsa únicamente como vejiga natatoria o únicamente como
pulmón.
En la Figura 22.4 se
representa a la vejiga natatoria en su modo de flotador. Muchos peces óseos
poseen una derivación desde las branquias hacia una bolsa en la cual hay un
flujo reverso de oxígeno desde la sangre hacia el aire contenido en la vejiga
aumentando la flotabilidad del animal. El aire puede perderse debido a que esta
bolsa de gas se deriva a su vez del sistema digestivo y el aire se puede perder
por el tracto digestivo hacia la boca. Sin embargo, algunos peces pueden hacer
el truco reverso, el primero es simplemente acercarse a la superficie y tomar
bocanadas de aire para no sacrificar el oxígeno que ya corre por sus arterias.
En otros el flujo de gases en la vejiga puede ir en retorno, es decir el
oxígeno puede ir desde la vejiga hacia la sangre, y de allí al corazón. Este
truco solo se usa cuando la sangre que viene de las branquias es pobre en
oxígeno debido a un estancamiento del agua, en estos casos en que el agua
pierde oxígeno y las branquias dejan de funcionar, poder tomar oxígeno del aire
gaseoso es una ventaja evolutiva primordial.
23. Sistema circulatorio en peces pulmonados
|| Regresar al Índice ||
Los peces pulmonados representan una alternativa más especializada de la vejiga natatoria, aunque esta se manifiesta primordialmente en el adulto, Cuando son alevines algunos géneros como Protopterus respiran por branquias externas, las cuales pierden parcialmente a medida que llegan al adulto y sus pulmones maduran. Estos peces literalmente generan pulmones, por lo que el flujo de oxígeno es unidireccional, desde el aire a los pulmones a la sangre.

Figura 23.1. Los peces
verdaderamente pulmonados emplean vasos sanguíneos degenerados reclutados desde
los arcos branquiales para generar el circuito respiratorio menor a los
pulmones. Este proceso es ontogénico y solo madura durante la metamorfosis, a
demás las demás branquias se pierden al madurar el pulmón.
El corazón posee una apariencia externa en
forma de S semejante a la de los anfibios urodelos “salamandras” (Burggreen y
Johansen 1986) el cual tiene tabiques que separan las partes derecha e
izquierda, sin embargo, a pesar de la creencia común, el corazón de los dipnoi no tiene cuatro cámaras reales.
La septación “la barrera que separa las partes izquierda y derecha de las
aurículas y ventrículos” es parcial, y en Neoceratodus
las de las aurículas apenas si son visibles.

Figura 23.2. Corazón de Protopterus.
Corazón del pez pulmonado Protopterus “Figura 23.2”,
aquí se observan septos o tabiques parciales que separan las aurículas y
ventrículos del corazón. Aunque la septación es parcial, funcionalmente cumple
su propósito de mantener las corrientes de sangre oxigenada y desoxigenada
separadas entre sí. En los anfibios, la separación del atrio se hace permanente
y las de los ventrículos desparecen. (Hirakow 1971). Las separaciones de los
lados derecho e izquierdo de las aurículas y ventrículos llamadas septos son
una estructura bastante flexible, por ejemplo, el septum (separación, o
tabique) del ventrículo de Neoceratodus es mucho menos notoria que en Protopterus (Burggreen y Johansen 1986).
La mayoría de los anfibios posee un corazón de tres cámaras “Figura 23.3”, en la que está presente el tabique del atrio (aurícula), pero no del ventrículo. Podría pensarse que se trata de un retroceso hacia el "ideal" cuatro cámaras completamente separadas de los amniotas, pero más bien se trata de una rama paralela al corazón de cuatro cámaras, de hecho, existen anfibios cuyo corazón se asemeja más al de los dipnoi con cuatro cámaras parcialmente separadas.

Figura 23.3. El corazón de los anfibios.
En otras palabras, la antigua visión de
progreso de corazones de dos cámaras (peces) a tres cámaras (anfibios) a cuatro
cámaras (amniotes) es solo un producto de nuestra vieja tendencia a pensar en
la evolución de manea lineal donde también confunden el proceso evolutivo con
progreso. Sin embargo, antes de hacer un argumento, vale la pena mencionar a
otro integrante de esta historia, aunque es un anfibio, se llama Siren lacertina. S lacertina se diferencia de otros anfibios en que posee una
septación parcial tanto en ventrículo como en aurícula (Putnam, 1977), ergo, no
todos los anfibios tienen tres cámaras en sus corazones, y, es más, podemos
establecer que el carácter de tres cámaras en una rama paralela del proceso
evolutivo que conllevó al corazón totalmente septado de los amniotes (reptiles
modernos, aves y mamíferos).
El corazón de los dipnoos pose cuatro
cámaras principales, típicas de un pez teleósteo Figura 23.2, un sinus venosus
relativamente empequeñecido que recibe sangre desoxigenada de la vena cardinal
y la vena hepática, una aurícula que posee ahora una nueva conexión a la vena
pulmonar, la aurícula, el cono arterial el cual posee una adaptación
interesante que comparte con los anfibios no sirénidos el cual desemboca en una
única arteria, la aorta. A pesar de que los tabiques interauriculares e inter
ventriculares no son completos, resulta notable que la sangre desoxigenada
tiende a fluir por el lado izquierdo mientras que la oxigenada por el lado
derecho sin mezclarse en el corazón.
El corazón de un pez pulmonado se
caracteriza por la presencia de tabiques parciales que separan los lados
derecho e izquierdo de su corazón. Ahora
sin embargo, la anatomía del corazón de los dipnoi
posee un detalle aparentemente molesto, solo tiene una condenada arteria de
salida Figura 23.2, lo
que implicaría que a pesar de la separación de la sangre en el corazón gracias
a los tabiques parciales, esta se perdería al mezclarse la sangre en el cono arterial y en la aorta. La válvula
espiral, una serie de pliegues en el cono arterial, ayuda a mantener la
separación de los tipos de sangre por lo menos hasta llegar a los primeros
arcos branquiales, donde la sangre ya oxigenada tendera a tomar arcos
diferentes a la desoxigenada. Las válvulas espirales son derivaciones
especializadas de las válvulas canales encontradas ya en el corazón de peces
sin mandíbula como las lampreas, y consisten en dos pliegues del endocardio
ambos pliegues quedan opuestos y de frente, pero no se tocan, es decir no
forman un tabique.

Figura 23.4. Neoceratodus posee branquias y pulmones en el adulto, por lo que este tipo de sistema circulatorio emplea simultáneamente dos órganos respiratorios.
El cono arterial posee una rotación interna de 270° los cuales hacen desembocar a los pliegues en una espiral doble en la zona del lumen. Debido a que el cono arterial se encuentra de manera directa al ventrículo, la sangre oxigenada tiende a entrar por el canal en espiral izquierdo, mientras que la desoxigenada tiende a entrar por el canal espiral derecho. Lo interesante del mecanismo es que aun cuando ambos flujos ingresan a la aorta, la separación parcial es suficiente para enfocar a los tipos de sangre en arcos branquiales diferentes previniendo así la mezcla de la sangre, y por lo tanto manteniendo un sistema circulatorio eficiente con pulmones y branquias funcionando en paralelo.

Figura 23.5. En peces como Protopterus
los dos órganos ventilatorios no funcionan al tiempo, los alevines emplean
branquias externas, y sin pulmones desarrollados su circuito circulatorio es
semejante al de peces como tiburones, pero a medida que el pulmón madura y su
sistema de vasos sanguíneos crece este va asumiendo las funciones de las
branquias que se van perdiendo.
Cuando un pez pulmonado llega a la superficie para “chupar aire” en sus pulmones, e flujo de sangre a los pulmones se incrementa. Cuando esta sangre oxigenada regresa al corazón y sale por la aorta tiende a entra a los arcos branquiales III y IV, los cuales carecen de branquias, fluyendo de manera directa a la aorta dorsal, y de allí al sistema circulatorio sistémico. La sangre desoxigenada que regresa de la vena hepática y la vena cardinal y pasa por el corazón, tiende a entrar a los arcos branquiales V y VI, los cuales poseen tanto branquias como derivaciones a los pulmones. Si se van a los pulmones completan un circuito circulatorio menor, pero si siguen por las branquias desembocan en la aorta dorsal al sistema circulatorio sistémico. A pesar de la perdida de las branquias de los arcos branquiales II y IV los dipnoos pueden respirar bastante bien con las branquias restantes sin necesidad de recurrir a sus pulmones mientras que el agua se encuentre adecuadamente oxigenada. Sin embargo, cuando los niveles de oxígeno disminuyen, la difusión de este gas a las branquias declina significativamente, pero el aire posee una buena cantidad de oxígeno, en ese caso el pez chupa aire atmosférico lo cual provoca que la sangre de los arcos V y VI dirija su sangre hacia los pulmones y no hacia la aorta dorsal. La pérdida de las branquias de los arcos III y IV posee un significado adaptativo, pues si permanecieran, la sangre oxigenada de los pulmones podría perder su oxigeno con respecto al agua debido al gradiente de concentración, (en ese caso la sangre portaría más oxigeno que el agua y el flujo por transporte pasivo siempre tiende a igualar las concentraciones).

Figura 23.6. Tanto en los
teleósteos con vejigas natatorias como en los dipnoi tenemos 6 arcos branquiales, se enumeran del I al VI, donde
el I "no mostrada en ambas imágenes" es el más cercano a la cabeza
mientras que el VI el más cercano al corazón. En los teleósteos con vejiga
natatoria, todos los 5 arcos poseen branquias funcionales. Las branquias V y VI en ambos tipos de peces
poseen derivaciones ya sea a la vejiga natatoria o a los pulmones, sin embargo,
en los teleósteos todos los arcos tienen branquias funcionales, (Gill), mientras que en los dipnoos las
branquias III y IV ya no existen, permitiendo que la sangre oxigenada pase de
largo hacia la aorta dorsal.
Por otra parte, así como existen
diferencias en la extensión de los tabiques que separan los lados de los
corazones entre los diferentes peces pulmonados, así también hay diferencias en
la funcionalidad de pulmones y branquias El pez pulmonado australiano por
ejemplo posee pulmones poco desarrollados con respecto a sus branquias, por lo
que depende más de la respiración branquial que de la pulmonar. El pez
pulmonado africano es el inverso, dependiendo más de la respiración pulmonar
que de la branquial. Por lo anterior, recalco, la sangre en los peces
pulmonados no se mezcla a pesar de tener una sola aorta.
24. Sistema circulatorio de los anfibios
|| Regresar al Índice ||
Bueno, antes de empezar con asuntos de
anatomía y fisiología debemos aclarar algunos conceptos básicos. Por alguna
razón la gente confunde el corazón tipo que se enseña en un curso de biología
básico con un corazón platónico, ideal que deben tener todos los seres vivos
que tienen esa misma descripción. Tal fenómeno ocurre con el corazón de los
anfibios. Sin embargo los anfibios pueden presentar dos tipos de corazones, el
corazón simplificado tipológico que describiremos con detalle y que se denomina
de tres cámaras debido a la desaparición del tabique ventricular, y el segundo
encontrado principalmente en el género Siren
que es de cuatro cámaras, el cual es más cercano a la forma ancestral que
conectaría a los ancestros de los peces pulmonados con tabiques parciales pero
con cuatro cámaras al fin y al cabo, los anfibios con corazones de cuatro cámaras parciales o
totales y el corazón de los reptiles cocodrileanos con cuatro cámaras bien formadas. Debido a que el funcionamiento
del sistema y el corazón de Siren es
similar en principio al de los corazones de los peces pulmonados, nos
enfocaremos en lo siguiente al corazón de tres cámaras más común de los
anfibios.
24.1 Anatomía y fisiología
Por mucho tiempo, especialmente entre los libros de biología básica escolar "y yo crecí con ellos" se transmitió la idea arcaica y errada que la sangre de mezclaba en el ventrículo, sin embargo, esa idea es falsa.

Figura 24.1. Vista
simplificada del sistema circulatorio de los anfibios.
Al igual que en los dipnoos, la sangre que entra en el cono arterial aún está separada en corriente desoxigenada y corriente oxigenada. Por alguna razón la gente confunde el corazón tipo que se enseña en un curso de biología básico con un corazón platónico, ideal que deben tener todos los seres vivos que tienen esa misma descripción. Tal fenómeno ocurre con el corazón de los anfibios. El corazón tipo que comúnmente se enseña “y que describiremos en detalle a continuación” es un corazón cuyo ventrículo no posee tabiques, pero el ventrículo si, tradicionalmente se le denomina corazón de tres cámaras.

Figura 24.2. El corazón de los anfibios
muestra una mezcla de las cámaras lineales o en S de los peces y la separación
de los flujos de sangre, esta separación es mucho más marcada en el género Siren
spp., que si tiene tabiques separadores.
Esta representación en forma de S es algo
engañosa Figura 24.2,
aunque nos permite ver las 5 cámaras que tiene el corazón de un anuro, sinus venosus,
dos aurículas, un ventrículo y un cono arterial. Si señores, no es un corazón
de tres cámaras. Por otro lado, en otras representaciones más realistas, el
cono arterial y la aorta se elevan tocando la pared externa de los ventrículos
como su dejáramos colgando al corazón desde todos los vasos sanguíneos, lo cual
deja al ventrículo como ápice inferior. Este esquema no muestra las trabéculas.
El problema es que ese corazón pertenece solo a un grupo de anfibios modernos
los anuros “sin colas” o ranas, sapos y afines. Por esto recalco, en esta
sección hablaremos del corazón de sapos y ranas y no del corazón de las
salamandras. ¿Por qué hacer esta aclaración?, simplemente porque no todos los anfibios tienen corazón de rana.
Los anfibios confían para su intercambio
de gases con el medio externo en una diversidad de mecanismos al igual que los
peces teleósteos, pudiendo realizar la ventilación completamente por la piel
(Salamandras plethodontidas), en branquias (la mayoría de los renacuajos y en
el adulto del axolote), o en pulmones (la mayoría de ranas, sapos y salamandras
hasta cierto grado). Debido a la
diversidad de fuentes de sangre oxigenada y desoxigenada, la forma de los
corazones varia.
Generalmente, en los anfibios con pulmones
funcionales el corazón se compone por un sinus venosus; la aurícula
tabicada conectada a una vena pulmonar; el ventrículo que carece de tabiques; y
un cono arterial similar al de los peces pulmonados, el cual presenta una serie
de pliegues que mantiene la separación “válvula espiral” entre la sangre oxigenada
y desoxigenada. Excepto por las salamandras del genero Siren spp., los anuros son únicos con
respecto al resto vertebrados con respiración pulmonar, ya que aún los dipnoos
poseen tabiques en el ventrículo para separar las corrientes de sangre oxigenada
y desoxigenada.
Las válvulas semilunares se encuentran en la base del cono evitando que exista un flujo de sangre retrogrado durante la diástole ventricular. Al igual que ocurre con los dipnoi, posterior a la válvula semilunar y en el interior del cono arterial se encuentran una serie de pliegues que giran a medida que avanzan formando una espiral doble, que separa los flujos de sangre. A pesar de que en los adultos de los anfibios hablar de arcos branquiales es algo complejo, el proceso es algo similar al que sucede en los dipnoos, la sangre que va separada por la espiral toma arcos diferentes, ya sea para los pulmones o para el sistema. Los arcos por donde se va la sangre sin oxígeno serian homólogos a la arteria pulmonar, mientras que los arcos que van al sistema serian homólogos a las derivaciones de la aorta. Todo muy bien y homologo entre dipnoos anfibios y cocodrilianos, pero hay un detalle, ¿Cómo es que un ventrículo no tabicado puede mantener los flujos de sangre separados?
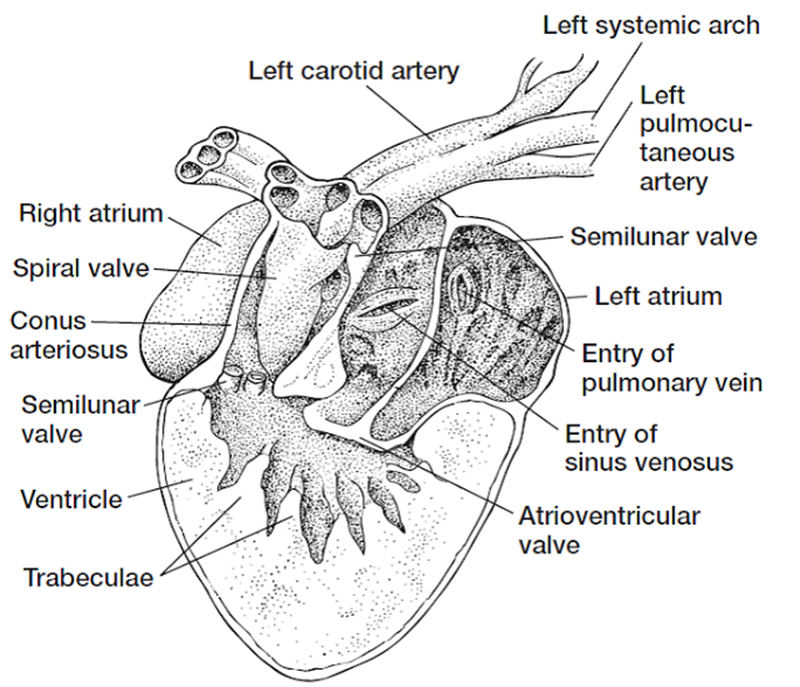
Figura 24.3. Una
representación más realista del corazón de los anuros, con el ventrículo como
cono inferior y todos los vasos sanguíneos en la parte superior como las puntas
de una corona. El cono arterial forma la base de la cual se desprende la arteria
pulmonar y la aorta. Interesante es que, en esta vista, el sinus venosus
queda atrás de toda la imagen, y por eso nunca lo vemos en las imágenes
clásicas de corazones. Tal como lo hemos mencionado en otros artículos, existe
diversidad en la morfología y fisiología de los corazones de los anfibios, el
mejor ejemplo es el género de salamandras Siren, las
cuales poseen tanto ventrículo como aurícula completamente tabicados recordando
al corazón de cuatro cámaras de los reptiles cocodrilianos.
Tal habilidad es sin duda algo
sorprendente, más aún cuando los fisiólogos y anatomistas originales pensaban
que las sangres se mezclaban, cosa que se ha demostrado como falsa. Aunque aún
sigue siendo un problema peliagudo en fisiología animal, se ha hipotetizado que
las trabéculas del ventrículo sirven como almacenes para un tipo de sangre,
mientras que el otro llena el centro del corazón, y de esta manera el
ventrículo mantiene separados ambos flujos, aunque esto solo funcionaria si,
los dos tipos de sangre tienen propiedades tan diferentes que fluyen de manera
separada aun cuando entran en contacto.
24.2 Efecto de la metamorfosis en el sistema
circulatorio de los anfibios
Sistema circulatorio anterior de un renacuajo de salamandra Figura 24.4-A, es muy similar al de los dipnoos en cuanto a la presencia de arcos branquiales funcionales mezclados con arcos branquiales sin branquias. En todo caso se observa claramente cómo, la arteria pulmonar (Pa) es un resultado evolutivo de un arco branquial con una derivación a los pulmones. El sistema circulatorio anterior de una salamandra adulta Figura 24.4-B, se observa la perdida de las branquias en los arcos, dejando entonces como arterias sistémicas. La arteria pulmonar sigue siendo una derivación del arco VI que también está conectado a la aorta dorsal (Da).

Figura 24.4. La metamorfosis altera
radicalmente la anatomía del sistema circulatorio sobre la marcha estos
procesos se dan mientras el individuo está activo y luchando por su existencia.
En A tenemos un renacuajo de salamandra, en el cual no se ha completado el
circuito circulatorio menor, el pulmón no está conectado al corazón, en B
podemos ver que después de la metamorfosis el circuito se completa. En C
tenemos el sistema circulatorio de una rana adulta. Otro aspecto importante es
que la metamorfosis provoca la degeneración de la mayoría de los arcos
branquiales y sus vasos sanguíneos, excepto aquellos reclutados en la
circulación del adulto.
Sistema circulatorio anterior de un una
rana Figura 24.4-C,
al igual que en las salamandras, desaparecen las branquias de los arcos, pero
en este caso el arco VI no se conecta a la aorta dorsal (Da) si no que posee
una derivación a la piel (Pc) a parte de la derivación a la arteria pulmonar
(Pa). En resumen, a pesar de que hay un solo ventrículo los flujos de sangre no
se mezclan y tienden a ingresar secciones diferentes del cono arterial. La
razón de ello aún permanece en debate, pero se cree que las trabéculas
"pliegues en el borde inferior del ventrículo" juegan un papel
importante.
25. Sistema circulatorio de los reptiles no cocodrileanos
|| Regresar al Índice ||
Los reptiles se caracterizan por haber
evolucionado originalmente para sobrevivir en ambientes más puramente
terrestres, por lo cual presentan adaptaciones más relacionadas con la respiración
de aire atmosférico que los anfibios. El corazón de los anfibios tiene una
capacidad para soportar un mayor volumen de sangre, así como para generar una
mayor presión sanguínea. Lo anterior les permite a los reptiles poseer
metabolismos más poderosos que sustentan estilos de vida más activos. A medida
que la variabilidad de los corazones ha empezado a entenderse de una mejor
manera, la antigua visión antropocentrista de ver a los corazones reptilianos
como incompletos con respecto al corazón de los mamíferos como una noción
falsa. Tal como establece la moderna Teoría Sintética de la Evolución, todo ser
vivo que ha sobrevivido hasta la actualidad se debe considerar como totalmente
evolucionado.
Lo anterior también posee implicaciones
sobre el anticuado concepto de fósil viviente, puede que algunos seres vivos
posean caracteres ancestrales, pero eso no implica que toda su fisiología sea
ancestral, aun cuando posean algunos caracteres que se han conservado
evolutivamente, generalmente, poseen otros caracteres que han cambiado o se han
diversificado a lo largo de las eras geológicas. Aun así, por cuestiones de
espacio o tiempo, los dos corazones que se han enseñado de manera clásica en
los reptiles son el corazón de los quelonios y el corazón de los reptiles
cocodrilianos, aunque también veremos algo del corazón de los varanos.
25.1 El corazón de las tortugas y lagartijas
En estos reptiles, el sinus venosus
está reducido en comparación con el presente en los anfibios, pero aún sigue
siendo una cámara conspicua y retiene la función de recibir la sangre de las
principales venas retornantes del sistema y del hígado. Al igual que en los
peces, el marca-pasos se encuentra posicionado en el sinus venosus, por lo que es en
esta cámara en la que se origina el latido. La aurícula se encuentra
completamente dividida por un tabique grueso en dos lados, derecho e izquierdo,
mientras que unas válvulas prominentes homólogas a las válvulas bicúspide y
tricúspide, encargadas de evitar el flujo retrogrado de sangre desde el
ventrículo. Sin embargo, entre los amniotas empieza a verse una característica
general y es la reducción se ciertas cámaras.
Estrictamente hablando, el ventrículo posee una sola cámara verdadera, y así es como se representa en los esquemas básicos, dando la impresión de que es un corazón similar al de los anfibios, pero desde luego esta es una visión algo simplista de la realidad. El ventrículo en realidad posee tres compartimentos interconectados, la razón por la cual no los denominamos cámaras reales es la ausencia de válvulas entre las conexiones. Estos tres compartimientos se denominan cavum venosum “cavidad venosa”, cavum pulmonale “cavidad pulmonar” ambos conectados por un tabique muscular con la abertura apropiada; y el cavum arteriusum “cavidad arterial”. La cavidad arterial está conectada a la cavidad venosa por el canal interventricular. Si consideráramos a las cavidades del ventrículo como cámaras reales, y contando al sinus venosus, el corazón de estos reptiles seria de seis cámaras. El ciclo cardíaco es algo complejo de explicar visualizando la imagen real del corazón, por lo que utilizaremos un esquema para poder conceptualizarlo más fácilmente.

Figura 25.1. Figura
relativamente precisa del corazón de una tortuga, debido a que la cavidad
pulmonar taparía a la cavidad arterial en este punto de vista, hay dos
imágenes, la de arriba muestra la cavidad pulmonar, y la de abajo la cavidad
arterial. En el corazón de este tipo el
cono arteria ancestral de los anfibios y peces teleósteos solo aparece como una
estructura transitoria durante el desarrollo embrional, rápidamente
dividiéndose hasta la base, formando la base del tronco arterial, finalmente
genera la base de tres arterias principales, la arteria pulmonar y las arterias
sistémicas derecha e izquierda. La forma en que las arterias salientes de la
corona del corazón se conectan al resto del sistema circulatorio de la parte posterior
del animal varía mucho, dependiendo del tipo de reptil.
En primera instancia, debemos entender que los reptiles al carecer de branquias deben tener adaptaciones especiales si desean sumergirse. Eso lo logran alterando el funcionamiento de sus corazones, por lo que diferenciaremos la fisiología del corazón en el modo normal que describiremos en el presente artículo, y el modo de desviación o modo de inmersión que describiremos en el siguiente artículo.

Figura 25.2. Modelo
simplificado del corazón de tortugas y lagartijas.
25.2 Modo
normal
Durante el modo normal, el corazón funciona en paralelo, la sangre llega al sinus venosus y a la aurícula derecha por medio de la diástole ventricular, al mismo tiempo se llena la aurícula derecha por el mismo movimiento.

Figura 25.3. Ciclo cardíaco
normal o de superficie.
De las auriculas la sangre pasa al único
ventrículo, pero no se mezclan, ya que el ventrículo este compartimentado, la
sangre venosa ingresa a la cavidad venosa, mientras que la sangre arterial
ingresa a la cavidad arterial en espera.
Durante la diástole auricular la sangre es expulsada al ventrículo y
aquí es donde la cosa se nos complica. Lo que va a diferenciar los modos del
corazón es la fuerza con la que late un lado del ventrículo con respecto a la
otra, esto hará que la sangre desoxigenada siga dos rutas diferentes. Durante
el modo normal, la sangre desoxigenada llena la cavidad venosa, mientras que la
sangre oxigenada llena la cavidad arterial. Este es el primer paso.
La sangre venosa pasa a la cavidad
pulmonar y de allí a la arteria pulmonar. La sangre arterial llena entonces la
cavidad venosa donde antes estaba la sangre venosa, y de allí es enviada a las
arterias sistémicas. El corazón late de
modo tal que, manda la sangre desoxigenada a la cavidad pulmonar, mientras que
la sangre oxigenada llena la cavidad venosa. Desde allí la sangre desoxigenada
es enviada por la arteria pulmonar, mientras que la sangre oxigenada es enviada
por los arcos sistémicos al resto del cuerpo. Nuevamente, LA SANGRE NO SE
MEZCLA.
25.3 Modo de
inmersión
La desviación sanguínea es un fenómeno fisiológico que caracteriza a los reptiles con vidas acuáticas muy marcadas como las tortugas. Cuando una tortuga se sumerge, su sistema circulatorio entra en un estrés directo, ya que los pulmones no pueden realizar intercambio de gases con el agua. Cuando los pulmones se ven incapacitados para realizar intercambio de gases, la presión parcial de oxígeno en su interior disminuye rápidamente hasta igualar la presión parcial en el sistema circulatorio, por lo que, en tal momento, es de poca utilidad para una tortuga hacer que su sangre circule por todo el circuito circulatorio menor. Al mismo tiempo, si utiliza toda la sangre para fluir únicamente por el circuito sistémico puede remover más eficientemente los metabolitos tóxicos como el dióxido de carbono generados por las operaciones metabólicas normales, lo cual alarga el tiempo en que su fisiología puede operar sin una fuente de oxigeno rica. Para lograr este objetivo, las tortugas pueden realizar la desviación de izquierda a derecha, en la cual la sangre no circula por el circuito circulatorio menor de los pulmones.

Figura 25.4. Ciclo
circulatorio durante la inmersión en el agua.
Durante el modo normal, la sangre
desoxigenada ingresa por las venas al sinus venosus, luego pasa a la
aurícula e ingresa posteriormente al ventrículo. En el ventrículo pasa por la
cavidad venosa y luego a la cavidad pulmonar, desde donde es enviada por la
arteria pulmonar a los pulmones. El
circuito menor se completa cuando la sangre regresa de los pulmones por la vena
pulmonar a la aurícula izquierda, de allí para al ventrículo. En el ventrículo
ingresa a la cavidad arterial, y de allí a la cavidad venosa después de que
esta se vacía de sangre desoxigenada. Una vez en la cavidad venosa, la sangre
oxigenada es enviada a las arterias sistémicas y de allí al resto del cuerpo.
Durante el modo inmersión hay una
alteración en la ruta de la sangre. Todo es igual desde la entrada al sinus venosus hasta
que la sangre llega al ventrículo. Pero la diferencia está en la ruta interna
del ventrículo. En este caso cuando la sangre se encuentra en la cavidad
venosa, en lugar de ser enviada a la cavidad pulmonar, es enviada directamente
a las arterias sistémicas, cosa posible ya que la cavidad venosa está conectada
a las arterias sistémicas.
26. Sistema circulatorio de los reptiles cocodrileanos
|| Regresar al Índice ||
El corazón de los reptiles cocodrilianos es realmente interesante al presentar una mezcla de caracteres, entre el corazón de los demás reptiles; y el corazón de aves y mamíferos. Al igual que sucede en los demás reptiles, el cono arterial forma la base de tres arterias “estilo reptil” y no dos “estilo aves y mamíferos”. La base de las arterias está custodiada por válvulas lunares, tal como ocurre en la mayoría de los vertebrados. Como siempre las válvulas permiten el flujo de sangre solo en una dirección. El sinus venosus aún se encuentra presente, aunque más reducido aun, su función se mantiene como la primera cámara en recibir la sangre proveniente de las venas sistémicas.

Figura 26.1. Modelo del
corazón de un reptil cocodrileano.
Las aurículas y los ventrículos sin
embargo se asemejan más a los corazones de las aves y mamíferos, en el sentido
de estar completamente divididos en tabiques; lo cual separa a la aurícula en
dos cámaras fisiológicamente independientes, la aurícula derecha e izquierda.
Lo mismo sucede con el ventrículo, que a diferencia de los demás reptiles
también se subdivide completamente por otro tabique, lo cual separa
fisiológicamente al ventrículo en un lado derecho e izquierdo. Esta división
hace que el corazón funcione en realidad como dos corazones en paralelo, donde
el lado derecho se encarga de succionar sangre del sistema y empujar hacia los
pulmones, mientras que el lado izquierdo del corazón se encarga de succionar
sangre de los pulmones y luego empujarla al sistema. El ventrículo derecho sin
embargo posee una característica interesante, posee dos arterias salientes, la
primera es bastante obvia, la arteria pulmonar, la cual empuja sangre desde el
ventrículo derecho “que está desoxigenada” a los pulmones. Sin embargo, lo que
es remarcablemente extraño, es la presencia de una segunda arteria, que es
sistémica, la cual envía sangre al sistema.
26.1 Modo
normal
Otro aspecto interesante es que esta arteria está conectada por un puente a la otra arteria sistémica que sale del ventrículo izquierdo. A este puente lo denominaremos foramen de Panizza. Esta adaptación sirve para realizar la desviación de sangre cuando el reptil se sumerge, algo parecido a lo que sucede con las tortugas. Al igual que sucede con las tortugas, el corazón de los cocodrilos presenta un cambio de fisiología según esté en el modo normal o en el modo inmersión. En el modo normal es cuando el animal se encuentra en tierra respirando aire atmosférico, en tal caso su sangre debe completar los dos circuitos circulatorios, el sistémico o mayor y el pulmonar o menor. En el modo normal, la sangre ingresa por las venas al sinus venosus y de allí a la aurícula derecha. De allí se dirige al ventrículo derecho. Cuando el ventrículo derecho hace su sístole, la sangre es enviada por la arteria pulmonar. Ahora, si están poniendo atención, se preguntarán por que la sangre no ingresa por ambas arterias.

Figura 26.2. Ciclo
circulatorio en estado de normalidad.
Cuando la sangre del ventrículo izquierdo
ingresa a la arteria izquierda, esta fluye tanto al exterior como hacia la otra
arteria gracias al puente que conecta a ambas arterias, de allí la sangre se
dirige al exterior, pero también hacia el ventrículo derecho, en ese momento la
válvula semilunar se cierra clausurando el conducto por esa arteria. La razón de ello es el puente de las arterias
sistémicas. Ambos ventrículos hacen su sístole al mismo tiempo, pero el lado
izquierdo es más potente en el modo normal, esto hace que la sangre del
ventrículo izquierdo pase por la arteria sistémica izquierda a mayor presión;
la presión de transmite por el puente entre las arterias, lo que hace que se
cierre la válvula de la arteria derecha. Como la arteria derecha está cerrada,
la sangre no tiene más opción que salir por la arteria pulmonar. Luego, la
sangre pasa por los pulmones, regresa al corazón por la aurícula izquierda, de
allí pasa al ventrículo izquierdo; ingresa a la base de la arteria izquierda,
la cual desvía sangre a la derecha por medio del puente arterial o foramen de
Panizza, lo cual hace que la sangre salga finalmente por ambas arterias al
sistema.
26.2 Modo de
inmersión
Cuando un cocodrilo se sumerge por un
periodo de tiempo considerable, su corazón ingresa en el modo inmersión.
Reflejos inconscientes hace que se cierren una serie de válvulas y esfínteres a
todo lo largo y en la base de la arteria pulmonar clausurando esa ruta. Al
clausurarse esta ruta, la presión sistólica en el ventrículo derecho se eleva,
haciendo que la sangre desoxigenada salga más rápido por la arteria derecha,
luego pasando por el puente y regresando a la base de la arteria izquierda, en
ese momento la válvula se cierra, clausurando la salida al ventrículo
izquierdo.
En el modo inmersión, la sangre sistémica se recicla directamente hacia el sistema, lo cual alarga el periodo de tiempo en que el reptil puede permanecer activo sin respirar nuevamente. De ese modo, la sangre desoxigenada no completa el circuito circulatorio pulmonar, y es enviada de manera directa al sistema. Como en las tortugas originalmente se ha pensado como una adaptación a la inmersión para ahorrar energía y mejorar la eficiencia de retiro de metabolitos tóxicos de la sangre. Sin embargo, aunque la desviación cardíaca se estudió como un mecanismo asociado a la inmersión, se ha demostrado que muchos reptiles lo utilizan en otras circunstancias. Debemos recordar que si algo caracteriza a la vida reptiliana es que es una vida de ahorradores por naturaleza. En lugar de usar energía propia para calentar sus tejidos, los reptiles dejan que el Sol lo haga por ellos; mientras no hay nada que hacer, ellos se quedan quietos recibiendo Sol o escapando de él. Si no necesitan gastar energía en algo, buscan ahorrar lo más posible y allí es donde entra la desviación cardíaca.

Figura 26.3. El foramen de
Panizza desvía la sangre venosa de regreso al sistema circulatorio mayor.
Cuando un reptil descansa no necesita
mucha energía, de lo cual se sigue que requiere muy poco oxígeno. Si requiere
muy poco oxígeno, por lo que puede respirar a intervalos muy largos, e
inclusive puede ingresar en el modo inmersión, en el cual no necesita respirar,
un mecanismo que permite precisamente ahorra la energía de enviar la sangre por
el circuito circulatorio pulmonar cuando los pulmones están sin oxígeno. El
tiempo entre cada aspiración y llenado de pulmones varia con el modo de vida;
por ejemplo, reptiles no acuáticos mantienen la fisiología de desviación, no
para hacer inmersión, si no para hacer hibernación o estivación. Como el modo
inmersión originalmente permite un ahorro de energía cuando los pulmones no se
llenan, este puede ser empleado cuando el animal entra en reposo para hibernar.
Sin embargo, existen otras posibles aplicaciones. La desviación cardiaca juega
roles en la regulación del contenido de dióxido de carbono en la sangre; el
cual no solo es un metabolito toxico, también puede ser empleado como un
acidificante de la sangre, y también como materia prima para la síntesis de
ácidos, y purinas “componentes del ADN y el ARN”.
La sangre oxigenada y más alcalina se
emplea para estimular a los osteocitos y acelerar el crecimiento óseo. En otras
palabras, la desviación cardiaca puede emplearse para regular a otros sistemas
del cuerpo de los reptiles, especialmente aquellos que se ven afectados por las
temperaturas. Generalmente las funciones gástricas y de crecimiento se ven
afectadas por la temperatura, y la desviación permite regular eso. Sin embargo,
hay muy pocos estudios aun sobre las otras posibles utilidades del modo
inmersión, ya que, de toda la diversidad de reptiles, solo se conoce la
fisiología de algunos cuantos.
27. Sistema circulatorio de aves y mamíferos
|| Regresar al Índice ||
El corazón de las aves y mamíferos representa un interesante caso de evolución paralela/convergente, en la que ambas estructuras se desarrollaron a partir de un ancestro amniote diferente, para generar una estructura similar. En ambos casos del corazón original de cuatro cámaras en serie, dos de ellas se pierden o reducen casi completamente. El sinus venosus se reduce a un pequeño bulbo donde se encuentra el marcapaso. Tanto en aves como en mamíferos, el cono arterial se genera durante las estepas embrionarias, pero luego es dividido, formando las bases de dos arterias, la arteria pulmonar que se conecta al ventrículo derecho, y la arteria aorta general que se conecta al ventrículo izquierdo.

Figura 27.1. El diagrama
circulatorio de aves y mamíferos posee dos circuitos, el mayor o a los
sistemas y el menor o a los pulmones. A diferencia de los reptiles, estos no
poseen normalmente desviaciones, por lo que las adaptaciones a la inmersión se
realizan de otras formas cuando es necesaria.
Estructuralmente hablando, el corazón de
las aves es anatómica y fisiológicamente similar al de los mamíferos, pero
embrionalmente no se genera de una misma manera. Las diferencias principales
están en los vasos sanguíneos principales de la corona, especialmente las
arterias pulmonares. En los mamíferos, la zona reducida de lo que fuera el sinus venosus
posee una gran cantidad de fibras de Purkinje formando el nódulo sino atrial.
El nódulo sino atrial aun retiene la función de marcapaso iniciando el impulso
eléctrico que coordina la contracción del corazón. En cuanto a las dos cámaras
restantes, estas se encuentran anatómicamente subdivididas por tabiques
completos en izquierdo y derecho, esto sumado al circuito circulatorio menor,
hace que el corazón de cuatro cámaras de aves y mamíferos funcione en paralelo,
como si fueran dos corazones independientes que se mueven al unísono.
27.1 El
bloqueo de la desviación de inmersión
Aunque estructuralmente similares, los corazones de las aves y los mamíferos se originaron de manera independiente, aunque de un ancestro común con corazón tetracameral, lo cual los hace un ejemplo de evolución paralela. Esta diferencia es reflejada por su desarrollo embrionario, donde el modo de aparición del tabique interauricular y del tabique interventricular se da de maneras diferentes en cada grupo. A pesar de ello, fisiológicamente, ambos funcionan de manera similar, al funcionar como dos corazones que integran dos circuitos circulatorios, el pulmonar y el sistémico. A diferencia de lo que ocurre en los reptiles, ninguno es capaz de realizar la desviación cardiaca, debido a las necesidades metabólicas de ambos grupos, ya que da la casualidad que ambos son grupos de animales endotérmicos. La desviación cardiaca es solo operativa en animales que pueden mantener sistemas metabólicos de niveles más bajos.

Figura 27.2. Aunque
estructuralmente semejantes, los corazones de mamíferos y aves son analogías
paralelas evolutivas que abandonaron el foramen de Panniza y otras desviaciones
cardíacas propias de amniotes como los reptiles.
27.2 El ciclo
cardíaco en los mamíferos
En los mamíferos, el ciclo cardíaco
comienza con la entrada de la sangre a las aurículas provenientes de las venas
cava, la superior es homologa completamente a las venas cardinales de los peces
en el sentido de que carga solo sangre sistémica. La vena cava inferior por
otro lado carga no solo sangre sistémica si no toda la descarga de nutrientes
provenientes del hígado. Ambas venas descargan su contenido en la aurícula
derecha, mientras que la vena pulmonar descarga sangre oxigenada en la aurícula
derecha, este evento se da durante la diástole auricular de manera simultánea y
paralela. Casi al mismo tiempo el ventrículo se encuentra en diástole, por lo
que las válvulas bicúspide y tricúspide se encuentran cerradas en ese instante.
Durante el siguiente paso del ciclo
cardíaco las aurículas experimentan su
sístole, empujando la sangre, debido a que las válvulas de las venas se
cierran, y a que las válvulas bicúspide y tricúspide se abren la sangre no
tiene más opción que ir a los
ventrículos, quienes de manera simultáneas experimentan su diástole, esto hace
que la sangre se mueva por una combinación de fuerzas, la que genera el empuje
de los aurículas por la contracción, y por el vacío de succión del ventrículo
al expedirse.
Cuando el ventrículo se abre, las válvulas
semilunares de las arterias pulmonares y aorta se cierran, permitiendo que los
ventrículos se llenen completamente de sangre, hasta que reinicia el primer
paso del ciclo cardíaco, nuevamente los ventrículos experimentan sus sístoles,
cerrando las válvulas bicúspide y tricúspide, mientras las válvulas semilunares
se cierran, enviado la sangre a las arterias, mientras que de manera casi
simultánea, las aurículas inician su diástole reiniciando el ciclo.
27.3 La inmersión en mamíferos y aves
La capacidad de tener vidas bajo el agua no se restringe a los anfibios y los reptiles, algunos mamíferos pasan periodos de tiempo considerables bajo el agua, donde el caso más extremo sean quizás los cachalotes, estas inmensas ballenas se sumergen hasta profundidades que son problemáticas incluso para los submarinos de inmersión profunda, allí realizan tareas pesadas como la cacería del calamar gigante, para luego regresar a la superficie, todo con una bocanada de aire. Los corazones de los mamíferos y las aves están fisiológicamente obstaculizados para realizar la desviación cardíaca propia de los reptiles. En otras palabras, las aves y los mamíferos naturalmente se encuentran bloqueados por un diseño no muy útil para las actividades bajo el agua.

Figura 27.3. Dado que los
mamíferos y aves no pueden reciclar su sangre sistémica por desviación
cardíaca, dependen de una hemoglobina enriquecida, su sangre es su tanque de
oxígeno. Después de la sangre el musculo es el segundo reservorio, gracias a
una mioglobina enriquecida.
Cuando se sumergen, el oxígeno en sus
pulmones se consume rápidamente, y rápidamente se vuelve poco ventajoso para el
corazón empujar la sangre por todo el circuito circulatorio menor en términos
de volumen sanguíneo perdido y energía empleada en vano. Sin embargo, los
corazones de las aves y mamíferos se encuentran tabicados, por lo que es
imposible generar la desviación cardiaca. Los ajustes deben ocurrir por otros
medios. Cuando un tetrápodo se sumerge ocurren tres cambios fisiológicos en el
sistema circulatorio. Primero que todo se genera bradicardia, es decir el ritmo
cardiaco se hace más lento, haciendo que el ritmo metabólico se haga más
relajado, lo cual de manera indirecta reduce tanto la cantidad de sangre que
fluye por el circuito circulatorio menor como la cantidad de oxigeno que es
consumido de los pulmones. Adicionalmente el cuerpo comienza a tomar oxígeno
almacenado en la sangre y en los músculos, y dado que los mamíferos solo tienen
acceso al oxígeno del aire, básicamente eso les debe durar. A mayor adaptación
a la inmersión los mamíferos dependen más del oxígeno muscular y sanguíneo que
el que se encuentra en los pulmones.
En segunda instancia, se incrementa el
ritmo del metabolismo anaeróbico “sip, las aves y los mamíferos también tienen
rutas anaeróbicas de respiración” en las fibras del musculo esquelético. En
última instancia, se abren una gran cantidad de vasos sanguíneos capilares, que
desvían mucho del flujo sanguíneo hacia los órganos vitales “en temas
anteriores también hablamos que el volumen total de los vasos sanguíneos
capilares es superior al de los vasos sanguíneos principales”. Al mismo tiempo
los capilares evitan que la sangre sea llevada a los pulmones, al musculo esquelético
“que es la mayor parte del musculo corporal” el cual funciona anaeróbicamente
en ese instante y al intestino.
En general, un ave y un mamífero hacen lo
mejor que pueden con una fisiología no muy apta cuando se encuentran bajo el
agua, adaptando las funciones secundarias de ciertos órganos como los vasos
sanguíneos capilares, para prolongar su estadía sumergidos. Cuando las aves y
los mamíferos se sumergen, existe muy poco oxigeno disponible, eso sumado a la
presión circundante hace que los pulmones colapsen “queden exprimidos como
pasas”. Aunque una gran cantidad de oxigeno queda almacenada inicialmente como
mioglobina, rápidamente es consumida. Por esta razón los tejidos echan mano de
rutas metabólicas no consumidoras de oxígeno “aunque consumen muchísima
energía” y al mismo tiempo, se hace más lento el pulso. Ninguna de esas
estrategias es única de las aves y los mamíferos, son adaptaciones secundarias
convergentes.
27.4 El embrión del pollo y su corazón

Figura 27.4. El desarrollo
embrionario es una de las evidencias para señalar que el corazón no es más que
una enorme vena superdesarrollada, pues incluso sus válvulas son semejantes en
forma y función a las que poseen las venas.
Los embriones y los individuos adultos viven en ambientes muy diferentes, por lo que no es de sorprender que los sistemas circulatorios atraviesen fases de desarrollo y fisiología, diferenciados. Debido a que el musculo cardíaco es completamente involuntario en sus pulsaciones, una vez se forma comenzará a moverse de manera rítmica, por lo que el corazón naciente comenzará a latir a los pocos días de iniciada su formación

Figura 27.5. Los vasos
sanguíneos que salen del corazón embrionario forman ramas que rodean a los
arcos faríngeos, algunos de estos vasos se degradan, mientras que las
hendiduras faríngeas se fusionan para formar la faringe y la tráquea.
El corazón de un pollo no posee tabiques
que separan los lados derecho e izquierdo, solo posee una vena de salida y otra
de entrada. También presenta transitoriamente un sinus venosus y un tronco
arterial homólogo al cono arterial de los peces. La arteria aorta que sale se
divide en arcos que rodean a la faringe en desarrollo. En las aves, el corazón
naciente consiste en una bomba con un solo ventrículo sin divisiones, que tiene
las mismas demandas hemodinámicas que el corazón de un pez adulto. En ambos, el
pez adulto y el embrión de un pollo, los tejidos de intercambio de gases y los
tejidos sistémicos se encuentran en serie. La demanda de sangre e completa por
un corazón en serie en un único circuito circulatorio “como en los peces”. Este
es un ejemplo más de adaptaciones estructurales que responden a necesidades
similares.
Los arcos aórticos rodean a la faringe en
desarrollo, la cual posee aberturas, como similares a los arcos branquiales que
presentan los peces. El corazón embrionario debe ser visto tanto como una etapa
de desarrollo transitoria, pero también como una etapa funcionalmente activa en
todo el sentido de la palabra. La mayoría de los vertebrados afrontan cambios
en el sistema circulatorio para acomodarse a los cambios y exigencias
fisiológicas que ocurren durante y después del nacimiento (o rotura del huevo).
Estos cambios son mejor conocidos en los mamíferos placentarios, grupo al cual
pertenecen los seres humanos.
27.5 El corazón fetal en los mamíferos placentarios
En los mamíferos euterios (todos los mamíferos menos monotremas y marsupiales) el feto depende exclusivamente de la placenta materna, la cual le proporciona todos los nutrientes, incluyendo el oxígeno. Una sola vena umbilical lleva sangre oxigenada desde la placenta hacia el hígado del feto, allí, la mitad ingresa al hígado y la otra mitad lo rodea ingresando directamente en la vena hepática. La sangre en la vena hepática se une a una gran cantidad de sangre que regresa por la vena precava y postcava hacia la aurícula derecha tal como en el corazón de un adulto “claro, después de que se forman los tabiques interauriculares”. Sin embargo, la mayoría de la sangre nunca llega a los pulmones en desarrollo debido a dos características anatómicas.

Figura 27.6. Esquema que
muestra el circuito circulatorio del feto de un mamífero. El foramen ovale
es la válvula que se encuentra en medio del tabique interauricular, el cual
permite comunicar sangre oxigenada directamente del ventrículo izquierdo al
derecho. La sangre desoxigenada avanza por la arteria pulmonar, pero no avanza
más que en un 10% a los pulmones, el resto sigue por la derivación al sistema
circulatorio sistémico uniéndose a él por el ducto arterial.
La primera es que el tabique auricular no
se forma completamente, y en su lugar deja una abertura, por lo que las
aurículas derecha e izquierda comparten sangre sin que este pase por el
circuito circulatorio pulmonar. A demás del orificio, también se genera una
válvula de una sola vía que evita el flujo retrógrado de sangre desde la
aurícula derecha a la aurícula izquierda.
Por si fuera poco, y al igual que sucede
con los peces pulmonados, la arteria pulmonar posee una derivación hacia el
circuito circulatorio sistémico, y la sangre en su mayoría fluye hacia el
sistema y no hacia los pulmones. Después de completar su viaje por el sistema
de capilares, la sangre ale del feto por las arterias umbilicales, las cuales
se derivan de la arteria iliaca. Cerca del final de la gestación, el feto
mamífero/humano posee un sistema circulatorio complejo y muy especializado. De
manera fisiológica, el corazón de los mamíferos nos demuestra una vez más una
propiedad física de las sangres con y sin oxígeno, por alguna razón “¡No Se
Mezclan!” aun cuando tienen la oportunidad de hacerlo.
En este caso la oportunidad se da cuando
la sangre oxigenada de la placenta ingresa a las venas cava, donde se mezcla
con sangre desoxigenada, ambos flujos ingresan a la aurícula derecha, pero allí
se separan. La sangre oxigenada tiende a ingresa a la aurícula izquierda a
través de la cavidad del tabique interauricular incompleto, desde allí la
sangre oxigenada completa el resto del circuito circulatorio mayor como en un
corazón adulto.
La sangre que sale del ventrículo izquierdo nutre especialmente al cerebro en desarrollo. Cuando los humanos nacen, ocurren una gran cantidad de cambios de manera casi simultánea. A medida que el tejido fetal y de la madre se separan durante el proceso de nacimiento, la circulación placentaria termina. Solo entonces los pulmones que hasta entonces habían permanecido inactivos comienzan a funcionar. Cuando se incrementa la presión de oxigeno de la sangre que retorna de los pulmones se activan reflejos que cierran la derivación de la arteria pulmonar a sistema. Sin embargo, deben pasar unas cuantas semanas hasta que el orificio del tabique interauricular sea sellado por una combinación de tejido fibroso y muscular. Sin embargo, la válvula que se encuentra en el orificio del tabique no desaparece de manera definitiva hasta que el infante humano posee ya un año de edad.

Figura 27.7. El corazón
embrionario con el foramen ovale. (YouTube).
El conducto y la válvula que permiten el
paso de sangre desde la aurícula fetal derecha a la aurícula fetal izquierda recibe
el nombre de foramen ovale. Resulta interesante, sin embargo, que esta pequeña
adaptación falla en desarrollarse en alrededor de un tercio de los humanos de
manera completa, lo cual es un gran porcentaje. La razón por la cual no va un
tercio de la población al médico por problemas cardíacos es que aun con un
tabique parcial “¿a qué corazones me recuerda eso?” la separación se logra
gracias a presiones diferenciales entre las aurículas y a la probada tendencia
de las sangres oxigenada y desoxigenada a no mezclarse aun cuando se les da la
oportunidad más forzada. Lo anterior conlleva a una falta de síntomas clínicos.
Muchas características anatómicas del corazón del feto mamífero recuerdan de
una u otra forma a peculiaridades de corazones y sistemas circulatorios de
otros vertebrados, que tanto es solo una convergencia debido a presiones
similares, y que tanto se debe a un plan de desarrollo genético común excede
mis conocimientos actuales, sin embargo, una cosa es segura, como todo lo que
es forjado por selección natural, esto no es una coincidencia.
27.6 Conclusiones a cerca de la evolución de los
corazones vertebrados
Resulta increíblemente difícil trazar un
modelo evolutivo para los corazones de los vertebrados, especialmente en las
primeras etapas. Un detalle es muy evidente cuando tomamos en cuenta el
desarrollo embrionario y fetal, y es que, el corazón, así como el resto del
sistema circulatorio es un órgano muy plástico, y proteico en la definición
griega de la palabra, cambia y se adapta con mucha facilidad.
Por lo anterior es complicado diferenciar
entre características ancestrales y especializaciones específicas entre los
diferentes corazones de los vertebrados. Aquí tal vez más que en cualquier otro
sistema de órganos tenemos solo puntitas, de un árbol, del cual es muy difícil
inferir las ramas. Tal vez, la única ayuda que tenemos es el desarrollo
embrionario y la genética, de la cual, si podemos entablar relaciones, en
primera instancia el corazón comenzaría como un vaso sanguíneo al cual se rodea
con fibras de tejido muscular cardiaco y un pericardio, como en los tunicados y
en los embriones de los demás vertebrados. Al mismo tiempo, los primeros
vertebrados también poseían corazones accesorios tal como se ha visto en los
cefalocordados, los mixinos y las lampreas.
Sin embargo, los corazones accesorios
pierden importancia en favor de una circulación centralizada en un único punto.
Después de ello, el corazón embrional empieza a desarrollar cámaras y a generar
nuevos vasos sanguíneos, y dependiendo del grupo, las diferentes
especializaciones para el modo de vida único de cada grupo. Es una descripción
muy general, pero tiene su razón principal en el hecho de que no existe un
corazón básico sobre el cual podamos trabajar en muchos grupos, es un problema particularmente
insidioso en los reptiles y en los cordados basales.
28. Sistema circulatorio linfático
|| Regresar al Índice ||
El sistema linfático es el compañero del sistema circulatorio. Ayuda a que los fluidos retornen al sistema circulatorio general y está enfocado en varias funciones especializadas, dentro de las cuales la más relevante es la función inmune. Estructuralmente hablando existen dos componentes del sistema linfático: los canales linfáticos y el tejido linfático. Si hablamos de analogías, el sistema linfático tiene varios, desde ser la carretera del sistema inmune a ser simplemente el drenaje por donde sale el exceso de agua de los tejidos.

Figura 28.1. El sistema
circulatorio linfático es paralelo al general, sirve para transportar al
sistema inmune y para regular el agua.
28.1 Linfa
El fluido que es transportado por el
sistema linfático se denomina linfa. En su mayoría está constituido por agua
obtenida de la matriz extracelular de los tejidos y algunas sustancias
disueltas como electrolitos, proteínas, pero sin células rojas por lo que es
incapaz su tasa de transporte de gases metabólicamente importantes como el
oxígeno molecular y el dióxido de carbono es baja, sin embargo, transporta
otras células. Debido a la naturaleza del tubo linfático terminal, la
composición de la linfa es la misma que la del fluido intersticial que está en
contacto con los tejidos.
28.2 Los canales linfáticos en los vertebrados
Colectivamente los canales linfáticos
constituyen un sistema tubular ciego que recircula el fluido desde los tejidos
de regreso al sistema circulatorio general. Las paredes de los canales
linfáticos son similares a los de las venas del sistema circulatorio general en
el sentido de que poseen válvulas de cerrado unidireccionales semejantes a las
que se encuentran en el corazón. Se dice
que son canales ciegos debido a que sus puntas penetran en los tejidos, pero no
se conectan al sistema circulatorio general.
En términos básicos la función del sistema circulatorio linfático es regular el contenido de agua en la matriz extracelular de los tejidos, esto se debe a que durante la circulación por los capilares parte del agua del plasma ingresa a los tejidos y las vénulas solo pueden recobrar el 90% de esta. El 10% restante permanece en los tejidos diluyendo la matriz extracelular lo cual causa problemas osmóticos en los tejidos, al haber más agua las células pierden nutrientes por presión osmótica, además ingresan más agua de lo normal lo que conlleva a que se inflen como globos explotando. Si esta situación se generaliza en todo un tejido entonces este colapsa, fenómeno denominado edema. El sistema circulatorio linfático recupera ese 10% de agua desde la matriz extracelular y lo retorna nuevamente al sistema circulatorio general.

Figura 28.2. Los capilares
linfáticos están en estrecha relación con los capilares sanguíneos, esto les
permite a los glóbulos blancos atravesar las paredes entre uno y otro según sea
necesario para controlar las infecciones.
La presión de flujo en los canales
linfáticos es baja, pero no tan baja, pues de lo contrario los canales
absorberían exceso de fluido. Los canales principales del sistema linfático
recogen la linfa que transporta el agua recolectada desde los tejidos gracias a
la red capilar. Al igual que el sistema circulatorio general los canales
linfáticos se ramifican subsecuentemente hasta que el grosor de sus paredes es
el de una sola célula, lo cual favorece el transporte a través de la membrana
del agua, los electrolitos, las proteínas y otras células. Una vez que los
capilares recogen el material retornar a canales más y más grandes y depositan
en agua en las venas precava y postcava.
Debido a que el sistema linfático solo
apoya la función de la circulación general su presencia no se condensa en
canales mayores, por lo que su presencia siempre es la de una red de canales en
cada sector del cuerpo. Las principales redes de drenaje linfático son: la red
linfática yugular que drenan los tejidos de la cabeza y el cuello, la red
linfática subclava que drena los apéndices anteriores, la red linfática lumbar
que drena los apéndices posteriores, y la red linfática torácica que drena el
tronco, las vísceras y la cola si está presente.
La presión del fluido de los canales
linfáticos no proviene del corazón de forma directa, ya que el sistema
linfático es independiente y se comunica con la circulación general por los
mecanismos de transporte a través de membrana. En consecuencia, se crea el
problema de generar presión. Las soluciones para este problema son variadas,
pero siguen el mismo tema de la circulación general, por ejemplo, creando
corazones linfáticos que emplean musculo estriado en lugar de musculo cardiaco
para generar potencia, emplear el movimiento muscular general para presionar
los canales linfáticos o asociarse íntimamente con las arterias principales de
modo que el impulso del corazón se transmita indirectamente a los canales
principales del sistema circulatorio linfático.
En el ser humano los canales linfáticos
tienen una arquitectura segmentada que se referencia como bulbos, cada bulbo
inicia y termina en una cisterna que abre y cierra unidireccionalmente como la
de las venas, asegurando que no exista flujo retrogrado hacia el ciego de los
canales, lo cual los haría explotar conllevando a un edema. El sistema capta la
linfa por medio de transporte pasivo a través de membrana, del siguiente modo.
Cuando el último segmento está lleno de linfa capturada desde el tejido, la
válvula de salida al siguiente segmento se abre, dado que el segmento está
vacío se genera una presión de vacío que absorbe la linfa, dejando al segmento
ciego vacío y cerrando la válvula. En la siguiente etapa el segmento ciego
vacío captura fluido de la matriz extracelular del tejido por medio de los
mecanismos de transporte pasivo y osmosis, llenándose de agua y partículas en
igual concentración que la matriz hasta que la presión osmótica se iguala a
ambos lados de la membrana del canal linfático terminal. En la siguiente etapa
se vuelve a repetir la primera, haciendo del sistema linfático un canal
palpitante semejante al venoso, pero que requiere del apoyo de la presión
muscular para funcionar adecuadamente.
28.3 Tejido
linfático en los vertebrados
El sistema linfático también incluye el
tejido paralinfático e inmune, que es una colección de tejido conectivo y
células libres. Las células libres abarcan a las células del sistema inmune o
leucocitos, conocidas popularmente como los glóbulos blancos debido a que
forman una fase blanquecina durante la centrifugación de sangre total en medio
de la fase de plasma y la fase eritrocitaria en el fondo. El tejido linfático
puede encontrarse casi en cualquier parte del cuerpo en forma de un tejido de
distribución difuso, ya sea en placas o encapsulado en los nódulos linfáticos.
Un nódulo linfático es una colección de tejido linfático encapsulado en una barrera de tejido fibroso. Los nódulos linfáticos se localizan al interior de los canales linfáticos a lo largo de la ruta de la linfa que retorna al sistema circulatorio general. Esta ruta asegura que la linfa penetra a través del tejido linfático como si fuera un filtro, de forma tal que las células y partículas al interior de la linfa sean expuestas a las células del sistema inmune. Los nódulos linfáticos se encuentran en los mamíferos y algunas aves acuáticas pero están ausentes en los demás vertebrados, convirtiéndolos en un ejemplo de carácter análogo. En los reptiles lo que sucede es la dilatación de los vasos linfáticos denominados cisternas linfáticas o sacos linfáticos.

Figura 28.3. Modelo
esquemático de la relación entre el sistema circulatorio linfático y el sistema
circulatorio general. Los bulbos intermedios se denominan los nódulos
linfáticos, lugares donde se producen las células del sistema inmune en el
adulto.
28.4 Fisiología del sistema linfático
El sistema circulatorio linfático sirve
como un sistema de apoyo al sistema circulatorio venoso, absorbiendo el exceso
de fluido que etapa de los capilares al MEC de los diferentes tejidos para
evitar los edemas. También se encarga de la absorción de lípidos del sistema
digestivo. El problema del edema plantea una situación anatómica complicada,
por un lado, el sistema general arterial posee de por si una alta presión, esta
presión favorece un flujo osmótico hacia el MEC del tejido en lugar se
proseguir hacia las venas por su baja presión. El sistema linfático al estar
aislado de la presión arterial recibe de manera osmóticamente favorable el
exceso de fluido desde el MEC, pero debe decantar dicho fluido en algún punto
del sistema circulatorio general donde la presión sanguínea sea baja.
En la mayoría de los peces el sistema general venoso es la puerta de acceso para el fluido linfático, especialmente en las venas que están a punto de hacer contacto con el corazón, donde el efecto de aspiradora del corazón contribuye a que los fluidos venosos y linfáticos sean jalados a la fuerza, en lugar de ser impulsados por una presión sanguínea que ya casi no poseen en ese punto.

Figura 28.4. Modelo de un
nódulo linfático, el cual no solo está conectado por los vasos linfáticos, sino
también por vasos del sistema circulatorio general, esto para enviar las
señales de alerta más fácil y para ser alimentados.
Sin embargo, con la evolución de los
pulmones, la sangre venosa retornante desde los pulmones se encuentra bajo una
alta presión. La división de la aurícula en dos cámaras independientes permitió
la separación de las presiones sanguíneas retornantes, una alta, desde los
pulmones y otra baja desde las venas del sistema circulatorio venoso general.
Por tal razón para que los pulmones pudieran afianzarse evolutivamente de
manera permanente era necesario la aparición del tabique auricular que permitió
establecer el conducto de baja presión, por el cual el fluido linfático podía
acceder. El tejido linfático se encuentra íntimamente ligado al conjunto de
órganos y tejidos conocidos generalmente como el sistema inmune, encargado de
la remoción y destrucción de materiales extraños foráneos como bacterias,
virus, protozoos, toxinas, células cancerosas e incluso xenobióticos. Sin
embargo, los detalles de este sistema los veremos en su capítulo propio.
29. Sistema circulatorio humano
|| Regresar al Índice ||
Habiendo discutido en detalle muchos elementos del sistema circulatorio de los vertebrados, es conveniente no caer en la redundancia. Los seres humanos poseemos un sistema circulatorio mamífero particularmente típico.

Figura 29.1. Principales vasos sanguíneos del sistema circulatorio humano.

Figura
29.2. Aunque aparentemente complejo, el
sistema consta de un circuito menor o pulmonar y el circuito mayor o sistémico.
29.1 Sistema
circulatorio general y pulmonar
Los componentes principales del sistema
cardiovascular humano son el corazón, la sangre, los vasos sanguíneos y los
capilares, así como el sistema circulatorio linfático. En él están incluidos:
la circulación pulmonar, un recorrido a través de los pulmones, donde se
oxigena la sangre; y la circulación sistémica, el recorrido por el cuerpo para
proporcionar sangre oxigenada. Un adulto promedio contiene aproximadamente 4,7
a 5,7 litros de sangre, lo que representa aproximadamente el 7% de su peso
corporal total. La sangre se compone de plasma, glóbulos rojos, glóbulos
blancos y plaquetas. También el sistema digestivo funciona con el circulatorio
para proporcionar los nutrientes que el sistema necesita para mantener el
bombeo del corazón.
Estructuralmente hablando, el sistema se
divide en dos, la circulación mayor que va del corazón a los tejidos generales
y de regreso al corazón; mientras que la circulación menor va del corazón a los
pulmones y de regreso al corazón, sin órganos accesorios para el intercambio de
gases.
29.2 El
corazón humano
Es el órgano principal del aparato circulatorio. En los animales vertebrados, incluyendo el hombre, es un músculo hueco que funciona como una bomba aspirante e impelente que impulsa la sangre a través de las arterias para distribuirla por todo el cuerpo. El corazón humano tiene el tamaño de un puño y un peso de 300 gramos que equivale al 0.40% del peso corporal, está situado en el centro de la cavidad torácica flanqueado a ambos lados por los pulmones.

Figura 29.3. Anatomía
detallada del corazón humano.
Los pulmones son asimétricos debido a la
presencia del corazón, siendo el pulmón izquierdo alrededor de un 33% más
pequeño que su contraparte derecha, eso se debe a que el espacio
correspondiente al tercio faltante es dedicado al corazón, por lo que este se
ubica solo un poco a la izquierda del pecho, demasiado a la izquierda o hacia
arriba simplemente es el pulmón izquierdo “¡piénselo si desean matar
vampiros!”. Su funcionamiento es idéntico al de cualquier corazón mamífero que
ya tratamos en secciones anteriores.
30. Las microcirculaciones capilares
|| Regresar al Índice ||
La circulación de la sangre a todos los
órganos posee características especiales que se encuentran adaptadas para
suplir necesidades concretas de las funciones especializadas de cada órgano. En
este subsección examinaremos las características únicas de la circulación
sanguínea en el corazón, el cerebro, el intestino delgado, el hígado, el
músculo esquelético, la piel, el pulmón, el riñón y el feto.
30.1 Circulación
coronaria
La circulación coronaria provee un flujo de sangre al corazón, lo cual puede parecer estúpido ya que el corazón maneja volúmenes importantes de oxígeno. El problema radica en el hecho de que el endotelio del corazón que están en contacto con la sangre en su interior aísla la sangre allí contenida de transportar gases metabólicos al miocardio, y en consecuencia este debe ser alimentado hacia afuera por medio de las arterias coronarias.

Figura 30.1. Sistema
circulatorio coronario.
El objetivo de la circulación coronaria
más concretamente no es el corazón como un todo, sino el músculo que rodea al
corazón y que le permite contraerse, al cual denominamos miocardio. El
miocardio es el único músculo que nunca descansa realmente, por lo que siempre
debe consumir oxigeno aun en condiciones de no ejercicio corporal. Es más, en
estas condiciones basales, el miocardio consume tanto o más oxígeno que una
masa igual de músculo estriado realizan un ejercicio vigoroso. Y lo más
importante, este consumo basal dete multiplicarse por dos, tres o hasta cinco
veces a medida que el cuerpo ingresa en un ejercicio vigoroso.
Aunque parezca paradójico, el órgano que
más demanda oxígeno es también uno de los que tiene mayores restricciones para
su obtención debido a su propia fisiología. El flujo de sangre a través del
miocardio ocurre principalmente durante la diástole ya que las fibras están
relajadas y esto permite el acceso de la sangre, por el contrario, durante la
sístole las fibras se tensan y la sangre es expulsada de los capilares del
miocardio. La dependencia cardiaca de recibir la mayoría de su oxigeno durante
la diástole no presenta efectos deletéreos obvios en el flujo de sangre
coronaria total aun durante el ejercicio vigoroso, sin embargo, las personas
con arterias coronarias comprometidas por arteriosclerosis, un aumento de
tiempo en el cual el corazón está comprimido puede afectar las células del
miocardio con principios de isquemia.
Algunos mamíferos como los perros poseen
una red de conexiones microvasculares de respaldo entre las arterias
epicardiacas principales, y se denominan canales colaterales. Un bloqueo total
de una de las arterias del epicardio no crea necesariamente un infarto debido a
que la sangre puede ser desviada a los canales colaterales. Los seres humanos
no somos tan afortunados debido a que hemos perdido casi todos nuestros canales
colaterales en el curso de la evolución, así que el bloqueo de una de las
arterias principales es casi una sentencia de muerte. Sin embargo, si el
individuo sobrevive a uno o más eventos de hipoxia en el miocardio, los canales
colaterales crecen para crear una desviación coronaria natural que ayuda a
mantener al individuo activo por un poco más de tiempo.
30.2 Circulación
cerebral
En el ser humano el principal órgano vital es el cerebro. De hecho. en la actualidad el órgano que se emplea como estándar para determinar la muerte biológica es el cerebro, debido a que aun cuando el corazón se detenga, este puede reanimarse, pero si el cerebro para no puede reactivarse de nuevo. La mayor causa de heridas cerebrales es la hipoxia causada por accidentes vasculares en el cuello o en el cerebro, debido a la oclusión de las capilares debido a procesos arterioscleróticos o aneurismas que ocurren como resultado del taponamiento de los conductos.

Figura 30.2. Sistema
circulatorio del cerebro y el rostro.
La microcirculación cerebral tiene muchas
características en común con la microcirculación coronaria. El cerebro y el
corazón poseen altísimas tasas metabólicas, extraen enormes cantidades de
oxígeno de la sangre y poseen una habilidad muy limitada de conducir rutas
metabólicas anaeróbicas en situaciones de estrés por falta de oxígeno. Un
aspecto interesante de la fisiología sanguínea del cerebro es que las regiones
evolutivamente antiguas poseen una capacidad para regular la presión sanguínea
y sobrevivir fácilmente a situaciones de hipoxia, asegurando que los órganos
vitales mantengan su funcionamiento, sin embargo esta habilidad se puede
paulatinamente a medida que se llega a las regiones evolutivamente más
recientes, hasta llegar al corteza que tiene poca o nula habilidad para
regularse, y en consecuencia la conciencia que allí reside es un sensible a la
falta de oxígeno.
Otra característica de la circulación
cerebral es que los vasos sanguíneos son capaces de adaptarse a condiciones de
alta o baja presión sanguínea siempre y cuando dichos cambios sean graduales,
un par de semanas son suficientes como para que el cerebro se adapte y opere
con normalidad, sin embargo, el cerebro es muy sensible a los cambios de
presión instantáneos o rápidos, esto se debe a que los vasos sanguíneos del
cerebro son inmunes a las hormonas vasodilatadoras y vasoconstrictoras.
30.3 Circulación
intestinal
El intestino delgado completa la digestión química de los alimentos y absorbe la mayoría de los nutrientes para el sostenimiento del resto del organismo. El consumo basal de sangre del intestino delgado es del 20% y el uso de oxígeno también es de un 20%, lo cual representa un quinto de toda la función cardiaca. Cuando se consume alimento en gran cantidad ambos valores se elevan a un 40%, lo cual implica que la irrigación sanguínea durante la actividad intestinal requiere de mucha energía.

Figura 30.3. Vasos
sanguíneos que rodean al intestino delgado.
Si se realiza ejercicio intenso justo
después de haber comido, el flujo sanguíneo a través del intestino disminuye a
la mitad de lo normal, esto retrasa la absorción de los nutrientes, pero no
causa ningún efecto colateral grave. Una vez que el estrés energético al cual
se somete el cuerpo termina, el flujo de sangre al intestino retorna. Esta
prelación es evolutiva, ya que en el ambiente un animal debe cuidar su
supervivencia, y si el alimento ya se encuentra en su intestino bien puede
esperar a que la situación de peligro pase, para poder seguir con su proceso de
digestión.
La vascularización del intestino delgado
es compleja. Las arterias y venas intestinales penetran la pared muscular y
forman una red capilar en la submucosa. Las capas musculares reciben pequeñas
arteriolas de plexo submucoso, otras arteriolas pequeñas continúan individualmente
hacia la submucosa profunda alrededor de las glándulas y la mucosa de los
vellos. Toda la red capilar de la submucosa puede regular el flujo de sangre
que fluye a través de ellas.
Debido a que el intestino se encuentra en
contacto casi directo con los nutrientes, la sangre que este obtiene solo se
requiere por su contenido de oxígeno y porque es el mecanismo para disponer de
sus desechos metabólicos. Esto hace que cuando el intestino se encuentra en la
condición basal metabólica posea poca o nula capacidad para autorregular su
flujo sanguíneo, mientras que cuando la sangre fluye a través de él, el
intestino es capaz de regular su flujo. La mucosa intestinal recibe entre el
60% y el 70% del volumen total de sangre que se dirige al intestino en total.
Este volumen de sangre puede exceder el consumo basal del cerebro y el corazón,
lo cual explica porque después de una comida fuerte el cuerpo se siente lento y
con sueño, toda la energía se la está siendo consumida por el intestino.
30.4 Circulación
hepática
La circulación hepática irriga uno de los órganos más grandes del cuerpo, el hígado. El hígado se encarga principalmente de mantener la composición sanguínea del plasma sanguíneo. Por ejemplo, todas las proteínas del plasma son producidas por el hígado, y el hígado se encarga además de almacenar y dispensar glucosa a partir del glucógeno en inventario. El hígado también filtra la sangre de eritrocitos viejos, bacterias y metabolismos secundarios internos o externos.

Figura 30.4. Modelo del
sistema circulatorio del hígado.
El hígado humano tiene un flujo de sangre
importante, pero no tan destacable siendo apenas un cuarto de lo que subsume el
corazón en su estado base. La sangre proviene tanto de la arteria hepática que
ya ha pasado por el estómago, el intestino delgado, el páncreas, el vaso y
algunas porciones del intestino grueso, así como del sistema de venas portal
que regresan del intestino delgado. La sangre del sistema portal representa
entre un 67% a un 80% del total del volumen sanguíneo que ingresa al hígado. La
regulación de la cantidad de sangre que ingresa al hígado es un proceso
dinámico. La sangre arterial hepática aumenta o disminuye en función de la
cantidad de sangre que ingresa desde el sistema portal, manteniendo al hígado
con una homeostasis en su volumen sanguíneo. Este mecanismo se denomina como
tampón arterial hepático, y puede compensar hasta una disminución del 25% de la
sangre proveniente del sistema portal. El mecanismo exacto que permite realizar
esto se encuentra aún en investigación, pero se asume que es un mecanismo
hormonal.
Justo después de una comida fuerte y de
que el intestino hubiera absorbido los nutrientes, estos son almacenados en la
sangre y conducidos a partir del sistema portal al hígado. Durante este evento
la presión sanguínea depende principalmente de los capilares intestinales y de
las propias venas porta, más que de la regulación hormonal propia del hígado.
30.5 Circulación
del músculo esquelético
La microcirculación del musculo esquelético involucra al tejido con mayor masa del cuerpo. Cerca del 30% al 40% de la masa de un humano adulto. El consumo de oxígeno por parte del músculo esquelético en el estado base es bajo, pero debido a su masa representativa, su consumo total representa uno de los objetivos principales para la distribución del gas de oxígeno a lo largo del cuerpo. El flujo de sangre a través del músculo esquelético puede aumentar 10 veces, 20 veces o más durante la máxima vasodilatación asociada con el ejercicio aeróbico de alto rendimiento.

Figura 30.5.
Microcirculación del músculo y la piel.
Durante este instante el metabolismo de
demanda de oxígeno y nutrientes y su respectiva emisión de dióxido de carbono y
desechos aumenta de forma comparable. Durante este momento el flujo sanguíneo a
través de los músculos puede llegar a tres o más veces el consumo cardiaco
basal, aunque evidentemente en ese instante el corazón también se encuentra en
un estado de mayor consumo. Toda esta sangre se obtiene del intestino, el
estómago y el hígado que se vacían casi completamente.
30.6 Circulación
cutánea
La estructura vascular capilar cambia
dependiendo de la ubicación. En todas las áreas, sin embargo, se pueden
distinguir tres secciones. La primera sección se encuentra en la dermis
profunda, allí se extienden vénulas y arteriolas principales que envían y
recolectan sangre desde las capas más superficiales. La segunda sección se
encuentra en dermis superficial donde se genera una primera red capilar que
alimenta a las células con nutrientes y oxígeno. La tercera sección más externa
se encuentra en la epidermis, allí los capilares forman arcos de radiación, los
cuales se encargan precisamente de radiar la energía sobrante en forma de
radiación electromagnética infrarroja, y su análogo más sencillo es la del
radiador de un carro. En este sentido la sangre cumple con la función de
enfriamiento del cuerpo a través de la piel.
Debido a que el agua tiene una capacidad
calorífica mucho mayor que la del aire, si la piel está mojada el proceso de
radiación de calor es más efectivo, por tal razón las zonas que se calientan
más fácil se encuentran asociadas a glándulas exocrinas que emiten sudor. El
contenido de agua aumenta según la necesidad de radiar calor, o aumenta el contenido
de grasa según la zona está más relacionada con los caracteres sexuales
secundarios. Este patrón experimenta cambios en las manos, pies, orejas, nariz
y algunas áreas del rostro donde la red capilar intermedia proliferan bastante,
en contraste con las extremidades y el torso donde es limitada. A una
temperatura fresca de unos 25°C y baja humedad los vasos sanguíneos se
encuentran significativamente contraídos por medio de la emisión de
noradrenalina por parte de los nervios simpáticos que inervan todas las áreas
de la circulación.
La piel es el órgano más grande y
representa del 10% al 15% de toda la masa corporal, siendo la interfase del
cuerpo con el ambiente externo. La principal función de la piel es la
protección del cuerpo del ambiente externo, y su función al estar combinada con
el sistema circulatorio es la regulación del calor corporal, sea este producido
internamente, o que el ambiente lo emita o lo absorba. Cuando no es necesario
radiar calor los vasos sanguíneos de los capilares se mantienen cerrados,
evitando que la sangre fluya cerca de la superficie, lo cual aísla el calor del
medio externo. Por el contrario, si se necesita radiar calor, los capilares se
abren permitiendo que la sangre fluya cerca de la piel.
La piel como tal tiene una de las tasas
metabólicas más bajas de todos los órganos y requiere relativamente poca sangre
para mantener sus condiciones basales. En consecuencia, a pesar de su masa
relativamente extensa, su metabolismo base no plantea mayores requerimientos en
su demanda de sangre. Sin embargo, en climas cálidos y húmedos la demanda de
emitir el calor hace que la piel seas un reservorio importante de la sangre, lo
cual va en detrimento de otros sistemas de órganos.
30.7 Circulación
pulmonar
La microcirculación pulmonar tiene pocos detalles del lado de la sangre, sin embargo, hay que recordar que estos capilares no están en contacto con un tejido interno, sino con los alveolos que están en contacto con un ambiente semi-externo.

Figura 30.6. Vasos sanguíneos
de los pulmones.
Todos sabemos que el intercambio de gases
con un fluido depende del área de intercambio, a mayor área mayor intercambio
de gases. Adicionalmente la superficie de los alveolos debe estar húmeda para
que el gas en fase gaseosa se disuelva a una fase disuelta donde si puede hacer
el transporte pasivo a los capilares sanguíneos. Esto provoca una situación
delicada al alveolo, este debe estar húmedo, pues de lo contrario no hay flujo
de gases, pero solo a un mínimo necesario, si aumenta el fluido la tasa de
intercambio de gases entre el fluido pulmonar y el aire pulmonar disminuye,
proceso denominado edema pulmonar.
30.8 Circulación
renal
En cuanto a la microcirculación renal, sus detalles están relacionados a la estructura anatómica denominada el asa de Henle. En resumen, se trata de un intercambio de sustancias muy complejo ya que el plasma debe intercambiar desechos y nutrientes con la orina en formación en varios ciclos antes de que se forme la orina definitiva con los desechos.

Figura 30.7. Modelo del
sistema circulatorio renal.
30.9 Circulación
fetal
El desarrollo del feto humano depende de
que reciba nutrientes y de que pueda eliminar los desechos, cosa que no puede
hacer por sí mismo, aproximadamente al séptimo día de la concepción la blástula
que sale de la trompa de Falopio debe encontrar su camino hacia el endometrio.
Cuando el embrión entra en contacto con el endometrio, parte de su estructura
formará al feto, pero otra parte que es la más externa formará una estructura
de sostenimiento e interacción con la madre, las membranas embrionarias, de las
cuales debemos subrayar a la placenta.
La placenta fetal humana libera hormonas que permiten el desarrollo de un sistema circulatorio embrionario sostenido a partir de dos arterias umbilicales, que se desprenden de las arterias iliacas internas, y una sola vena umbilical. En el sistema cardiovascular fetal el orden de vena/arteria y sus funciones se encuentran invertidas. La vena umbilical transporta la sangre oxigenada y los nutrientes hacia el feto desde el cuerpo de la madre, mientras que las arterias se encargan de drenar dióxido de carbono y desechos metabólicos.

Figura 30.8. La placenta
media el intercambio de nutrientes y desechos entre la madre y el feto, pero
impide que las sangres se mezclen.
Resulta curioso que, a pesar de la enorme
transferencia de nutrientes y desechos entre la madre y el feto, no se
compartan de forma significativa los elementos celulares, en otras palabras,
hay muy poco intercambio de glóbulos rojos y glóbulos blancos entre la madre y
el feto. Esta aparente paradoja es posible debido a que la sangre maternal y
fetal se mantienen casi completamente separadas gracias al villus
placentario. El villus placentario es un espacio de separación
entre la circulación del feto y la madre, los capilares de ambos permiten el
flujo de sustancias a través del villus, pero mantiene aislado el
flujo de células.
Los capilares insertados en el villus
placentario se denominan vasculares placentarios, y la complejidad de su
ramificación depende de la concentración de nutrientes y/o desechos en ambos
lados de la relación madre feto. Si los nutrientes disminuyen del lado de la
madre, la complejidad de su vascular placentario aumenta, incrementando de este
modo el área de intercambio de sustancias. Esto permite regular el equilibrio
de sustancias entre madre y feto manteniendo la homeostasis.
Durante el desarrollo fetal, el tejido
fetal invade y causa una degeneración parcial del endometrio materno. El
resultado después de unas 10 a 16 semanas de gestación es un espacio entre el villus y el tejido materno vivo que se
encuentra lleno se sangre. En lugar de capilares solo hay un espacio cavernoso
lleno de sangre estancada. Para el final de la gestación el feto representa el
consumo de un órgano de alta demanda, tomando entre el 15% al 25% de la sangre
que sale del corazón en el estado base, por esta razón una madre gestante no
puede realizar ejercicios demandantes al final de su gestación. En comparación
la placenta representa la mitad de la emisión sanguínea del corazón del feto.
31. El sistema circulatorio y el ejercicio
|| Regresar al Índice ||
Tu corazón (conocido con frecuencia como
el miocardio, lo que significa músculo del corazón) es una bomba muscular de
cuatro cámaras del tamaño de tu puño y se localiza ligeramente hacia la
izquierda del centro de tu pecho. Su trabajo es bombear sangre a tu cuerpo.
Cuando haces ejercicio, tu ritmo cardíaco puede aumentar de un promedio de
descanso de 72 latidos por minuto, a 200 o más dependiendo de tu condición
física y edad. Conforme adquieras más condición, tu corazón se vuelve más
fuerte y tu ritmo cardíaco en descanso disminuirá. Las personas que están en
forma, con frecuencia tienen ritmos cardíacos bajos en reposo; este ritmo en
atletas altamente entrenados puede ser tan bajo como 30 latidos por minuto.
Tu cuerpo usa los vasos sanguíneos para
transportar la sangre a todo tu cuerpo. Las arterias toman la sangre del
corazón; las venas la regresan a él y los capilares dejan y recogen la sangre
de tus músculos y pulmones. Cuando te ejercitas, la hormona adrenalina provoca
que tus vasos sanguíneos se expandan de manera que un volumen de sangre mayor
al normal pueda pasar mediante ellos. Esto se llama vasodilatación, una
respuesta a corto plazo del cuerpo al ejercicio, y es una de las razones por la
que tus vasos sanguíneos pueden volverse más prominentes durante el ejercicio.
Como respuesta a largo plazo, tu cuerpo establece nuevos capilares para que el
oxígeno pueda entregarse mejor, y se pueda remover más dióxido de carbono de
tus músculos en movimiento.
Cuando te ejercitas, la sangre se desvía
de órganos no esenciales, como aquellos involucrados con tu sistema digestivo y
reproductivo, y hacia tus músculos. Esto se llama acumulación de sangre y
asegura que los músculos que están trabajando obtengan tanto oxígeno como
necesitan. Una vez que has terminado tu ejercicio extenuante, es importante
motivar a que esa sangre acumulada se mueva de los músculos y regrese a la
circulación general, lo que comúnmente se logra realizando un enfriamiento que
consista en ejercicio cardiovascular ligero y estiramientos. La sangre
acumulada que se queda en los músculos se relaciona con la aparición de dolor
después del ejercicio.
Tu sangre contiene tres tipos diferentes
de células: las células blancas que combaten las infecciones, plaquetas que
ayudan a la coagulación de la sangre y las células rojas que transportan el
oxígeno. Dichas células están suspendidas en un líquido llamada plasma, que es
principalmente agua. Cuando te ejercitas, las células sanguíneas se saturan con
oxígeno en su esfuerzo para asegurar que cantidades suficientes de este
componente estén disponibles para tus músculos. Como beneficio a largo plazo
del ejercicio, la cantidad de células rojas aumentan conforme a adquieras más
condición, y así eres más hábil para transportar mayores cantidades de oxígeno
en tu cuerpo.
Cuando se ejercita el tiempo suficiente, a un nivel aeróbico el hígado aumenta la producción de la lipoproteína de alta densidad o HDL. Esta lipoproteína se encarga de recolectar la grasa dispersa en los vasos sanguíneos y retornarla al hígado, donde será redistribuida por medio de la lipoproteína de baja densidad a los músculos para que estos puedan tener energía para moverse. Es importante hacer ejercicio de modo tal que la HDL siempre esté en altas concentraciones y la grasa no se acumule en las arterias y en los vasos capilares, especialmente en los capilares coronarios o en los capilares cerebrales, donde pueden desembocar en la arteriosclerosis, con la consecuencia final de un infarto fulminante al miocardio o un infarto cerebral.
Referencias bibliográficas
|| Regresar al Índice ||
Alberts, B.,
Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P.
(2015). Molecular biology of the cell (5a ed.). Garland Science.
Belk, C., & Maier, V. B.
(2013). Biology Science for Life with physiology. (Pearson, Ed.) (4a
ed.).
Berta, A., Sumich, J. L.,
& Kovacs, K. M. (2015). Marine mammals: evolutionary biology.
Academic Press.
Borlee, B. R., Goldman, A. D.,
Murakami, K., Samudrala, R., Wozniak, D. J., & Parsek, M. R. (2010).
Pseudomonas aeruginosa uses a cyclic‐di‐GMP‐regulated adhesin to reinforce the
biofilm extracellular matrix. Molecular microbiology, 75(4),
827–842.
Bowen, W. H., & Koo, H.
(2011). Biology of Streptococcus mutans-derived glucosyltransferases: role in
extracellular matrix formation of cariogenic biofilms. Caries research, 45(1),
69–86.
Brusca, R., Brusca, G. J.,
& Haver, N. J. (2003). Invertebrates (2a ed.). Sinauer Associates.
Campbell, M. K., &
Farrell, S. O. (2009). Biochemistry (6a ed.). USA: Thomsom Brooks/Cole.
Costerton, J. W., Stewart, P.
S., & Greenberg, E. P. (1999). Bacterial biofilms: a common cause of
persistent infections. Science, 284(5418), 1318–1322.
Cox, M. M., Doudna, J. A.,
& O’Donnell, M. (2012). Molecular biology (1a ed.). Freeman.
Cuberos, E., Rodriguez, A. I., & Prieto-Suárez, E.
(2009). Niveles de cromo y alteraciones de salud en una población expuesta a
las actividades de curtiembres en Bogotá, Colombia. Revista de Salud Pública,
11(2), 278–289.
Cuberos, E., Rodriguez, A. I., & Prieto, E.
(2009). Chromium levels and their relationship with alterations in the health of
tannery workers living and working in Bogotá, Colombia. Revista de Salud Pública, 11(2), 278–289.
De Beer, D., Stoodley, P., & Lewandowski, Z.
(1996). Liquid flow and mass transport in heterogeneous biofilms. Water
Research, 30(11), 2761–2765.
De Beer, D., Stoodley, P.,
Roe, F., & Lewandowski, Z. (1994). Effects of biofilm structures on oxygen
distribution and mass transport. Biotechnology and bioengineering, 43(11), 1131–1138.
De Duve, C., & Pizano, M. (1995). Polvo vital:
origen y evolución de la vida en la tierra. Norma, Bogotá.
Droleskey, R., Byrd, J.,
Bischoff, K., Harvey, R., & Nisbet, D. (s/f). Probiotics and Their Use in
Food Animal Production: a Novel Explanation for Antibacterial Properties. Proceedings
of the American Oil Chemists’ Society.
Drury, W. J., Stewart, P. S.,
& Characklis, W. G. (1993). Transport of 1‐μm latex particles in Pseudomonas aeruginosa biofilms. Biotechnology
and bioengineering, 42(1), 111–117.
Finlay, J. A., Schultz, M. P.,
Cone, G., Callow, M. E., & Callow, J. A. (2013). A novel biofilm channel
for evaluating the adhesion of diatoms to non-biocidal coatings. Biofouling, 29(4), 401–411.
Forero, A. R., Mantilla, J. F. G., & Martínez, R.
S. (2009). Accumulation of lead, chromium, and cadmium in muscle of capitán
(Eremophilus mutisii), a catfish from the Bogota River basin. Archives of
environmental contamination and toxicology, 57(2), 359–365.
Goodenough, J., & McGuire,
B. (2012). Biology of Humans, Concepts, Applications and Issues (4a
ed.). San Francisco: Pearson, Benjamin Cummings.
Hawser, S. P., Baillie, G. S.,
& Douglas, L. J. (1998). Production of extracellular matrix by Candida
albicans biofilms. Journal of medical microbiology, 47(3),
253–256.
Hernberg, S. (2000). Lead
poisoning in a historical perspective. American journal of industrial
medicine, 38(3), 244–254.
Hoefnagels, M. (2015). Biology:
concepts and investigations (3a ed.). McGraw-Hill New York.
Kåhrström, C. T. (2013).
Biofilms: Flagella function as mechanosensors. Nature Reviews Microbiology,
11(9), 597.
Kardong, K. V. (2011). Vertebrates,
comparative anatomy, function, evolution (6a ed.). McGraw-Hill New York.
Kardong, K. V. (2014). Vertebrates:
Comparative Anatomy, Function, Evolution (7a ed.). McGraw-Hill Education.
Karp, G. C. (2013). Cell
and Molecular Biology, Concepts and Experiments (7a ed.). USA: Wiley Online
Library.
Lee, S.-M., Kwon, H. Y., Im,
J.-H., Baek, J. H., Kang, J.-S., & Lee, J.-S. (2015). Identification of
Outer Membrane Vesicles Derived from Orientia tsutsugamushi. Journal of
Korean medical science, 30(7), 866–870.
Mackean, D. G., & Hayward,
D. (2014). Biology (3a ed.). IGCSE Cambridge.
Mader, S. S. (2010). Biology
(10a ed.). McGraw-Hill Education.
Mader, S. S., &
Windelspecht, M. (2015). Biology (12a ed.). McGraw-Hill Education.
Mader, S. S., &
Windelspecht, M. (2018). Essentials of biology (5a ed.). McGraw-Hill
Education.
Marieb, E. N., Wilhelm, P. B.,
& Mallatt, J. (2017). Human Anatomy (8a ed.). Pearson.
Mason, K. A., Losos, J. B.,
Singer, S. R., & Raven, P. H. (2014). Biology (7a ed.). McGraw-Hill
New York.
Miller, M. B., & Bassler,
B. L. (2001). Quorum sensing in bacteria. Annual Reviews in Microbiology,
55(1), 165–199.
Murray, R. K., Bender, D. A.,
Botham, K. M., Kennelly, P. J., Rodwell, V., & Weil, A. (2012). Harpers
Illustrated Biochemistry (29a ed.). McGraw-Hill Medical.
Pollard, T. D., Earnshaw, W.
C., Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell Biology
(3a ed.). Elsevier.
Reece, J. B., Urry, L. A.,
Wasserman, S. A., Cain, M. L., Minorsky, P. V, & Jackson, R. B. (2014). Campbell
Biology (10a ed.). Pearson Higher Ed.
Rossi, S. C., Gorman, N.,
& Wetterhahn, K. E. (1988). Mitochondrial reduction of the carcinogen
chromate: formation of chromium (V). Chemical research in toxicology, 1(2),
101–107.
Sadava, D., Berenbaum, M.,
& Hillis, D. (2014). Life the Science of Biology (10a ed.). Sinauer
& MacMillian.
Sadler, T. W. (2012). Medical
Embryology. (C. Taylor, Ed.) (12a ed.). Baltimore: Lippincott Williams
& Wilkins.
Shier, D., Butler, J., &
Lewis, R. (2015). Hole’s Human Anatomy & Physiology (14a ed.).
McGraw-Hill Education.
Simon, E. J., Dickey, J. L.,
& Reece, J. B. (2013). Essential biology (Pearson).
Singh, P. K., Schaefer, A. L.,
Parsek, M. R., Moninger, T. O., Welsh, M. J., & Greenberg, E. P. (2000).
Quorum-sensing signals indicate that cystic fibrosis lungs are infected with
bacterial biofilms. Nature, 407(6805), 762–764.
Solomon, E., Martin, C.,
Martin, D. W., & Berg, L. R. (2014). Biology (10a ed.). Cengage
Learning.
Speziale, P., & Geoghegan,
J. A. (2015). Biofilm formation by staphylococci and streptococci: structural,
functional, and regulatory aspects and implications for pathogenesis. Frontiers
in cellular and infection microbiology, 5.
Spiewak, R., & Dutkiewicz,
J. (2008). In vitro study of pro-inflammatory and anti-tumour properties of
microvesicles from bacterial cell wall of Pantoea agglomerans. Ann Agric
Environ Med, 15(1), 153–161.
Starr, C., Evers, C., &
Starr, L. (2013). Biology: Today and Tomorrow With Physiology (4a ed.).
Brooks/Cole.
Taylor, M. R., Simon, E. J.,
Dickey, J. L., Hogan, K., & Reece, J. B. (2018). Biology, Concepts and
conections (9a ed.). Boston: Pearson education.
Weaver, R. F. (2012). Molecular
biology (5a ed.). McGraw-Hill Education.
Wilking, J. N., Zaburdaev, V., De Volder, M., Losick, R., Brenner, M. P., & Weitz, D. A. (2013). Liquid transport facilitated by channels in Bacillus subtilis biofilms. Proceedings of the National Academy of Sciences, 110(3), 848–852.
No hay comentarios:
Publicar un comentario