
Índice
Índice
Portada

Portada
1. Introducción
1. Introducción
Desde que era niño, el tema que más me fascinó en la biología sin duda alguna fue la evolución, las eras geológicas y la existencia de criaturas imponentes y maravillosas que ahora no vemos. Tal vez eso marcó mi vocación de cierta forma. El problema es que la instrucción que recibí sobre el asunto fue algo frustrante y no es que mi profesor de bachillerato no la enseñase, tal vez es que de alguna forma magistral fue capaz de evadir toda controversia posible.

Figura 1.1. Los animales. Fue en esta enciclopedia "tomo de los Animales" donde por vez primera tuve contacto con la teoría de la evolución, sin tener demasiado contacto con el nombre de Darwin. Las ilustraciones son simplemente preciosas y el contenido textual es muy bueno aun para los estándares actuales.
La enseñanza de las ciencias tiende a tener ese defecto, instruir sobre datos, fechas, apellidos y teorías, pero no sobre los debates, las controversias, la competencia por el crédito académico, en fin, el trabajo que realmente se realiza al interior de las comunidades científicas. Aunque la palabra controversia no es una palabra que asociamos a la labor científica y puede sonar incluso hasta relativista, desde el punto de vista de ciertas corrientes filosóficas no positivistas de la ciencia es todo lo contrario, es decir gracias a la existencia de la controversia al interior de las comunidades científicas es que podemos tener la seguridad de un trabajo científico adecuado y honesto. Después de todo, la ciencia se basa en la capacidad de dudar de todo lo posible para buscar respuestas adecuadas, pero siempre en movimiento, siempre mejorables.
No soy tan arrogante como para pensar que puedo hacerlo mejor, pero al menos haré el intento. En este capítulo estará diseñado para ser aplicada en una unidad didáctica que debo realizar sobre el tema de la teoría de la evolución y estará fuertemente influenciada por la traducción que realicé hace ya un par de años de este artículo de revisión “(Kutschera & Niklas, 2004) The modern theory of biological evolution: an expanded síntesis. Naturwissenschaften 91:255–276. Aunque para los conceptos expandidos intentaré emplear fuentes más recientes. Sin embargo, también cabe resaltar otras fuentes como el libro de (Futuyma, 2005) "Evolution" así como diversos artículos relacionados con reconstrucciones históricas de diversos conceptos. Otras fuentes tal vez sean en base al espíritu e intención de S. J Gould “1941-2002” y también a la perspectiva de T. S. Kuhn “1922-1996” en el sentido de un adecuado manejo histórico de los problemas, su contexto y la necesidad de combatir los mitos al interior de la enseñanza de las ciencias, los cuales, aunque parcialmente cómodos resultan inconvenientes a la hora de una adecuada enseñanza o divulgación de este tema tan importante de las ciencias de la naturaleza.
1.1 Sir Ronald Aylmer Fisher (17 de febrero de 1890 - 29 de julio de 1962)
Fue un erudito, matemático, estadístico, genetista y académico británico. Por su trabajo en estadística, ha sido descrito como "un genio que casi sin ayuda creó las bases de la ciencia estadística moderna" y "la figura más importante de las estadísticas del siglo XX". En genética, su trabajo utilizó las matemáticas para combinar la genética mendeliana y la selección natural; esto contribuyó al resurgimiento del darwinismo en la revisión de principios del siglo XX de la teoría de la evolución conocida como síntesis moderna. Por sus contribuciones a la biología, Fisher ha sido llamado "el mayor de los sucesores de Darwin".

Fisher tenía fuertes opiniones sobre la raza y la eugenesia, insistiendo en las diferencias raciales; sin embargo, la evidencia sugiere que, aunque era claramente un eugenista, no apoyaba el racismo científico, y derivaciones de sus propias técnicas de ANOVA han llevado a la conclusión de que la especie humana no puede dividirse en razas biológicas. Fue profesor de Eugenesia Galton en el University College de Londres y editor de Annals of Eugenics.
Desde 1919 en adelante, trabajó en la Estación Experimental Rothamsted durante 14 años, allí, analizó sus inmensos datos de experimentos de cultivos desde la década de 1840, y desarrolló el análisis de varianza (ANOVA). Allí estableció su reputación en los años siguientes como bioestadístico.
Es conocido como uno de los tres principales fundadores de la genética de poblaciones. Esbozó el principio de Fisher, las teorías de la selección sexual de la hipótesis del hijo fugitivo y sexy de Fisher. Sus contribuciones a la estadística incluyen promover el método de máxima verosimilitud y derivar las propiedades de los estimadores de máxima verosimilitud, la inferencia fiducial, la derivación de varias distribuciones de muestreo, los principios fundamentales del diseño de experimentos y mucho más.
2. Generalidades de las teoría evolutivas
2. Generalidades de las teoría evolutivas
Cuando creamos una unidad didáctica sobre la teoría de la evolución es casi que obligatoria la mención de Darwin y del debate en el siglo XIX, en ocasiones ponemos la lectura de El Origen de las Especies y otros fragmentos de libros de texto, sin embargo, es posible realizar conexiones mucho más ricas. Seríamos afortunados si en el mismo ciclo escolar también se nos presenta la oportunidad de enseñar genética.
En mi caso, cuando me encontraba revisando los contenidos para la creación de la genética postmendeliana, en la que hay que hablar del ligamiento al sexo, los tipos de sangre, la codominancia y la meiosis surgió un patrón crucial. Cuando los temas se dan desconectados de sus autores y su desarrollo histórico es relativamente fácil separar a la genética postmendeliana del desarrollo de la teoría de la evolución en el siglo XX, pero cuando se indaga sobre el contexto histórico emerge un patrón, la genética postmendeliana y el desarrollo de la teoría de la evolución en el siglo XX son una misma historia. Es por esta razón que durante el desarrollo de temáticas primero me limité a todo el siglo XIX e inicios del XX; genética clásica y dinámica de poblaciones. He de retomar la genética postmendeliana junto con la teoría de la evolución en el siglo XX como lo que es, un solo cuerpo de conceptos.
2.1 La evolución como teoría
Sea cual sea la versión de ciencia manejada por los científicos una cosa es clara, la teoría científica es el sistema explicativo de mayor nivel al interior de las ciencias de la naturaleza.

Figura 2.1. Doctor David Goodstein. Es un físico y educador estadounidense. Desde 1988 hasta 2007 sirvió como Vice-rector del Instituto de Tecnología de California, donde también es un profesor de física y física aplicada.
De hecho, desde los acercamientos no positivistas la teoría científica y el consenso de la comunidad científica tienen un mayor nivel de importancia que la misma información, que los datos. Un ejemplo de esto puede verse al final del capítulo 49 del documental El Universo Mecánico y Mas Allá donde el profesor Goodstein “1939-vivo” advierte que no existe modelo o teoría perfectos, nuestras propuestas abstractas siempre estarán en conflictos con alguna parte de la información, con algún dato (Blinn & Goodstein, 1985). En tal caso los científicos deben preguntarse que está mal, ¿será un error experimental?, ¿algún presupuesto o simplificación necesaria durante los cálculos? O ¿será el indicio clave que permita generar una nueva teoría? Las teorías en las ciencias de la naturaleza son sistemas abstractos que no se prueban como un conjunto, son contrastadas contra la naturaleza por medio de intermediarios llamados hipótesis.
De la teoría emerge la hipótesis como consecuencia, estas hipótesis son contrastadas contra los fenómenos de los experimentos los cuales son interpretados por los instrumentos y ciertos presupuestos teóricos donde la regla más básica es el mantenimiento de un sistema de razonamientos lógicos. En cualquier caso, diremos que la teoría de la evolución es tan teórica como la teoría atómica, la teoría del electromagnetismo, o la teoría celular. De hecho, si comparamos la teoría de la evolución con la teoría atómica, la propuesta de Darwin seria análoga a la propuesta de Dalton, es decir, la teoría de la evolución actual es tan cercana a Darwin como lo es el modelo del átomo moderno a la propuesta de Dalton. Un aspecto final extraído de la propuesta de Kuhn, una ciencia sin teoría difícilmente puede ingresar al periodo de ciencia normal, es decir, una ciencia sin teoría no puede unificar un proyecto de investigación estable y no puede crecer.
2.2 La evolución en la escuela
“La verdad de nuestra fe se vuelve objeto de ridículo entre los infieles si cualquier católico, no dotado con el aprendizaje científico necesario, presenta como dogma lo que el escrutinio científico demuestra falso”.
Santo Tomás de Aquino.
El aspecto controversial de la teoría de la evolución es sin duda la razón por la cual el tema se hace más apasionante entre el público en general que otras historias de la ciencia como la teoría celular, los modelos atómicos o la mecánica clásica. ¿Qué tiene de especial la teoría de la evolución para ser tan controversial? Durante la edad de la ilustración las ciencias de la naturaleza comenzaron a realizar afirmaciones cada vez más osadas sobre el modo en que funcionaban diversos aspectos de la naturaleza que por tradición habían sido explicados por medio del mito y la religión. Esto ha decantado desde nuestra perspectiva en una especie de conflicto entre la religión, específicamente las diversas ramas del cristianismo con las ciencias a tal punto que muchos estudiantes de secundaria tienen dificultades con procesar el hecho de que muchos científicos brillantes son y fueron devotos religiosos e inclusive sacerdotes en total ejercicio como Gregor Mendel “1822-1884”, Francisco Ayala “1934-vivo” o Georges Lamaitre “1894-1966”.

Figura 2.2. Tomas de Aquino, Santo y Doctor en Filosofía Teológica de la iglesia católica.
Existe la percepción de que la teoría de la evolución niega a Dios, en palabras del profesor Douglas Futuyma la estrategia sobre este problema no puede ser la postura de Richard Dawkins “1941-vivo”. La controversia simplemente nunca desaparecerá al nivel de la sociedad, sin embargo, existen formas de congeniar las posturas. El modo en que cada estudiante valla a congeniar su propia postura es algo en lo que no debo influir, cada uno es libre de construir su propio sistema de creencias, mi problema recae en conocer y cómo puedo evitar que las creencias y preconcepciones tergiversen diversos conceptos fundamentales como la selección natural. En otras palabras, no he de pedir que me crea, pero tan solo le pido que conozca y no tuerza los conceptos, acéptelos tal como son, critíquelos tal como son, pero no los modifique a tal punto que se conviertan en una sátira.
De Santo Tomas de Aquino “1224-1274” he de tomar dos elementos conciliadores, la primera es la advertencia puesta en el primer párrafo del presente escrito, donde el aquitense advierte en no realizar afirmaciones sin fundamento académico, lo cual se hace muy común en los debates sociales sobre la evolución, el origen de la vida y el origen del universo. La segunda es la noción de causa primera y causas secundarias. En esta idea se nos dice que la causa primaria es Dios, semejante a la propuesta de Aristóteles como motor fundamental, causa incausada etc etc. El punto son las causas secundarias. Si realizamos una analogía diríamos que lo más cercano a las causas secundarias es lo que conocemos como las leyes de la naturaleza. Evidentemente desde las ciencias de la naturaleza es relativamente común la descripción de las leyes de la naturaleza, pero la existencia de una causa primera es objeto de muchos más problemas. Evadiendo todos estos dilemas filosóficos sobre la causa primera simplemente diremos que, la ciencia es un juego en el que solo se vale usar causas secundarias para explicar fenómenos naturales, es decir, donde acudir a la causa primaria es hacer trampa.
2.3 Independencia de la teoría de la evolución con respecto a otros campos de investigación

Figura 2.3. La vida es compleja. ¿Cómo resolver esta maraña de colores y formas? la explicación esencialista es reconfortante, pero bloquea las preguntas que pueden surgir a través de esta primera pregunta. Ya sabemos que podemos acudir al Dios lo hizo y san se acabó, el punto es ver que podemos aprender si asumimos que la causa primera no interviene y que el proceso de generación de especies es dinámico.
En este capítulo deberemos hacer una distinción entre evolución como paradigma y teoría de la evolución como campo de investigación en la biología. El paradigma evolutivo hace referencia a un modo general en que la mayor parte de la comunidad científica entiende el origen y desarrollo del universo. Bajo este paradigma se sostiene que el universo ha cambiado desde un pasado remoto a un presente. Tres campos de investigación se pueden distinguir bajo este paradigma.
👉 El origen y evolución del universo, campo de estudio que emplea las herramientas de la física y la química fundamentalmente y cuya explicación más confiable parece ser la teoría del BigBang en la actualidad.
👉 El origen de la vida, campo de estudio que emplea las herramientas de la química, la física y la biología, este campo del conocimiento aún carece de una teoría que genere un consenso al interior de las comunidades científicas, aunque lo más cercano a eso es sin duda la propuesta de Oparin “1894-1980” y Haldane “1892-1964”.
👉 Finalmente, la evolución de los seres vivos, campo de estudio que emplea principalmente las técnicas de la biología pero que también emplea a la química y la física. La teoría que genera más consenso en la actualidad es la teoría sintética de la evolución “TSE” aunque en los últimos 5-10 años varios autores han propuesto renombrarla como la síntesis expandida de la evolución.
Cada uno de los tres campos de investigación es independiente y posee un programa de investigación diferente, aun cuando todos obedecen al mismo paradigma. En este capítulo, la palabra evolución estará exclusivamente ligada a la evolución de los seres vivos.
2.4 El problema a resolver por las teorías evolutivas
Cualquier teoría evolutiva busca encontrar una explicación a la diversidad de los seres vivos mediante el uso del naturalismo metodológico. Antes de iniciarse la edad de la exploración en el siglo XV y XVI la diversidad de seres vivos conocidos por loes europeos era muy limitada. Con pocas especies por clasificar la noción tipológica de la naturaleza era fácil de evidenciar ya que existían muy pocas especies que sirvieran como nexos morfológicos entre grupos claramente diferenciables, por ejemplo, entre los reptiles y las aves. Sin embargo, con la edad de la exploración los huecos morfológicos comenzaron a llenarse, ya fuera por medio de especímenes raros capturados desde los confines del planeta o por medio de una nueva rama en la historia natural que sería denominada paleontología. Para la época de Darwin el registro fósil aún era fragmentario, pero las pocas especies con las que contaban estimulaban lo suficiente la imaginación como para aventurarse a proponer ideas radicales sobre el origen de las especies.
3. El pensamiento evolutivo desde Grecia hasta la edad media
3. El pensamiento evolutivo desde Grecia hasta la edad media
Solemos asociar todo pensamiento evolutivo a Charles Darwin, sin embargo, una de las principales ideas a las cuales nos tenemos que acostumbrar en este curso es que, hubo evolución antes que Darwin, y hay evolución posterior a Darwin. es semejante a la idea de que la teoría atómica se desplegó antes que Dalton y se desarrolló mucho después de Dalton. en esta lección analizaremos las ideas evolutivas anteriores, no a Darwin, sino a los primeros naturalistas de la edad de la exploración.
3.1 El pensamiento evolutivo en Grecia
Heredamos de nuestra cultura una serie de sistemas de pensamiento y modos de organizar la realidad natural y social que nos son invisibles. Una de las funciones de la historia es evidentemente hacer visible lo invisible. En el pensamiento evolutivo existen una serie de estructuras de pensamiento que facilitan el entendimiento de los conceptos modernos, pero del mismo modo existe otros que los dificultan. En la Grecia antigua debemos plantear al menos tres conceptos que nos acompañarán a todo lo largo de esta historia a cera de la evolución, la primera es el devenir, la segunda es el esencialismo y la última es la teleología. De los tres, los dos últimos son verdaderas taras debido a que son intuitivamente fáciles de entender y aplicar, y a que ciertas estructuras religiosas los acogieron como criterios para justificar aparte de su teología.

Figura 3.1. Heráclito según Rafael en la pintura “La Escuela de Atenas”.
👉 Presocráticos
Heráclito “535-484 antes de Cristo” nació en Éfeso una de las ciudades estado de cultura griega ubicada en la costa occidental de Asia menor en lo que actualmente es Turquía. El pensamiento de Heráclito tradicionalmente se asocia con el cambio y el devenir, de Heráclito se conoce la inmortal expresión: “Es imposible entrar en un río dos veces” esto es porque con el tiempo tanto el río como la persona han cambiado. De hecho, debe recordarse que la palabra evolución es un sinónimo de la palabra cambio, por lo que en últimas la estructura de pensamiento que facilita la comprensión de los procesos de cambio, específicamente los evolutivos en la biología se deberían a la aproximación de Heráclito. En Heráclito también podemos señalar otra cosa, para Heráclito el devenir causa un conflicto, pero un conflicto que en términos generales causa armonía (Michael & Ruse, 2009; Osborn, 1896; Wysong, 1976).
El pensamiento de Parménides “530-515” es de cierta forma lo opuesto al de Heráclito. Para Parménides lo importante es la identidad de las cosas, su esencia fundamental, la cual es estática. Mientras que el pensamiento de Heráclito es un pensamiento del cambio, en Parménides se desarrolló un pensamiento de lo estático. El pensamiento de lo estático y esencial influiría posteriormente en Platón y finalmente en todo el pensamiento occidental debido a la adopción por parte de la cristiandad del esencialismo. Con estas dos estructuras de pensamiento abriremos y cerraremos a la teoría de la evolución, pues, aunque la mayor parte de la comunidad científica ha adoptado el pensamiento heraclídeo del cambio, muchas personas influyentes al exterior de las ciencias de la naturaleza poseen un fuerte pensamiento esencialista basado en las creencias religiosas (Michael & Ruse, 2009; Osborn, 1896; Wysong, 1976).

Figura 3.2. Parménides según Rafael en la pintura “La Escuela de Atenas”.
Aunque de Heráclito debíamos rescatar la estructura de pensamiento que acepta al cambio como la constante de todas las cosas, el pensamiento evolutivo sobre los seres vivos en específico debe ser rastreado a otro filósofo presocrático, en este caso Anaximandro. Anaximandro de Mileto “610-546 antes de Cristo” propuso que los primeros animales vivieron en el agua durante un antiguo periodo de tiempo donde estaba húmeda, luego los primeros ancestros del hombre debieron haber nacido en el agua y solo pasaban parte de su vida en la tierra seca. Anaximandro también propuso que el primer humano como se lo conocía en su tiempo debía ser descendiente de otro tipo de animal.
👉 Los filósofos clásicos
Platón “428-348 AC” ha sido llamado por biólogos célebres como Ernst Mayr como el gran “Antihéroe del evolucionismo” debido a que su obra filosófica es el fundamento de dos de los más grandes obstáculos conceptuales para que las personas puedan entender los conceptos evolutivos, estos son el esencialismo y la escala natural.

Figura 3.3. Aristóteles y Platón. (Izquierda) Platón según Rafael en la pintura “La Escuela de Atenas”. (Derecha) Aristóteles según Rafael en “La pintura La Escuela de Atenas”.
Debemos recordar que para Platón lo único real son los ideales abstractos y que la realidad física es una distorsión, y eso aplicado a la biología se denomina la teoría esencialista de las formas fijas o simplemente esencialismo. De cierta forma podemos afirmar que es un desarrollo de las ideas de Parménides a cerca de las esencias. En el esencialismo los seres vivos se encuentran adscritos a tipos ideales que habitan en el mundo de los ideales. Existe por ejemplo un ideal de león, la variación que presenta cada individuo obedece a corrupciones que ese ideal experimenta al transferirse a la realidad física. Platón construyó un sistema cuasi-teológico ya que la transferencia de un ideal a la realidad, o la creación misma de cada forma ideal no era espontánea, dependía de una deidad universal que llamó el Demiurgo. Queda demás resalta que sus ideas influenciaron poderosamente a la cristiandad y a todas las teorías religiosas sobre la biología, y su proponente más moderno sería el Diseñador Inteligente (Michael & Ruse, 2009; Osborn, 1896; Wysong, 1976).
👉 La gran cadena del ser
La escala natural o la gran cadena del ser también es otra de las ideas que más han influido en la biología, sea para bien o para mal y del mismo modo que con e esencialismo la fuente principal es la obra llamada Timeo. En el Timeo Platón plantea que los demás seres vivos diferentes del ser humano son una corrupción paulatina de este, de este modo los menos corruptos serán más semejantes al ser humano y así sucesivamente desde el hombre hasta el reino de los minerales (Flannelly, 2017; Kutschera, 2011; Thein, 2006).
👉 La idea de la escala Natural también llamada la gran cadena del ser pone en la cúspide a los Dioses/Dios, luego las criaturas espirituales/ángeles.
👉 Posteriormente viene el hombre “masculino”, por debajo la mujer.
👉 Por debajo se encuentran los animales superiores o vertebrados, luego los inferiores o invertebrados.
👉 Después de los animales prosiguen las plantas superiores o terrestres y las plantas inferiores o acuáticas.
👉 Por debajo de las plantas están los minerales nobles “oro y plata” y luego los minerales bajos.
👉 La generación de escalas naturales será igual que con el esencialismo recolectada por la cristiandad y reencausada en los estudios filosóficos sobre filosofía natural.
Aristóteles “384-322” es el filósofo griego más influyente en la tradición occidental, después de todo sus opiniones son siempre tenidas en cuenta en las fases introductorias a muchas disciplinas como la genética, los modelos atómicos, la anatomía, la taxonomía y en este caso la evolución. Los escritos de Aristóteles resultaron de su investigación sobre el mundo natural de la isla de Lesbos y ha sobrevivido en cuatro libros que al ser traducidos al latín se conocen como: De Anima “acerca de la esencia de la vida”, Historia animalium “investigación sobre zoología” De generatione animalium “sobre la reproducción” y De partibus animalium “anatomía animal”. En este punto hay que remarcar que al ser alumno de Platón Aristóteles continuó la tradición de su maestro sobre la escala natural.

Figura 3.4. La cadena del ser y la escala natural. En Platón la escala natural sirve como un pensamiento evolutivo y lineal en la que lo más perfecto se degrada paulatinamente a lo menos perfecto. En Aristóteles la cadena del ser opera más como un sistema de clasificación. En cualquier caso, la escala natural será una idea fundamental que es muy difícil de desligar del pensamiento
En Historia animalium por ejemplo, Aristóteles clasificó a los organismos en relación a una escalera jerarquizada o cadena del ser, colocando en las partes superiores a los organismos con mayor complejidad estructural. De Aristóteles encontramos otra noción que dificulta el entendimiento moderno de la teoría de la evolución y es la noción de causa final u objetivo final de los seres vivos (Singer, 1931). Bajo esta idea se sostiene que debido a que los seres vivos exhiben exquisitas adaptaciones para funciones muy bien definidas, estos debieron haber sido creados para estas funciones finales, son creados para esos roles. A esta idea en la actualidad se la conoce como teleología. Aristóteles rechaza las ideas de Empédocles "490-430" de que los seres vivos se crearan al azar, debido precisamente a la exquisitez de sus adaptaciones (Ayala, 1970; Falcon, 2006; Gotthelf, 1999; Meyer, 1992; Singer, 1931).
👉 Epicuro
Epicuro “341-270 AC” es acreditado como el gran abuelo de la selección natural, pues, aunque no la explicita de forma completa, algunas ideas en su escuela de pensamiento parecen tocar el concepto. En el sistema de Epicuro muchas especies fueron generadas de forma espontánea por Gaia en el pasado, pero solo las más funcionales sobrevivieron para tener descendientes. Los epicúreos no anticiparon una teoría completa de la evolución tal como fue desarrollada posteriormente por Darwin y Wallace en el siglo XIX y aparentemente postularon que las especies se originaban por actos de creación independientes desde Gaia, en otras palabras, no existían la noción de ancestros comunes (Clark, Foster, & York, 2007; Stott, 2012a; Zirkle, 1941).
👉 Resumen
La filosofía griega nos sirve toda la discusión, sin embargo, dado que los filósofos más influyentes son sin duda Aristóteles y Platón resumiremos de ellos, tres conceptos que denominaremos obstáculos epistemológicos para el entendimiento de la evolución. El esencialismo, la noción de que las especies obedecen a tipos ideales e inmutables que han sido creados por el Demiurgo, Dios o el Diseñador Inteligente. La escala natural, la idea de que los seres vivos se organizan de forma lineal desde lo más perfecto a lo menos perfecto, con el ser humano en la cima. La teleología, la idea de que los seres vivos han sido creados de forma explícita e intencionada para cumplir un rol. Platón y Aristóteles serían retomados posteriormente por las escuelas de pensamiento de la cultura occidental y en especial de la cristiandad. Por este motivo durante la edad media y la ilustración los temas a tratar son básicamente estos tres.
3.2 El pensamiento evolutivo en la antigua China
Antiguos filósofos chinos como Zhuangzi “369-286 AC” de la escuela taoísta expresaron ideas sobre la naturaleza de los seres vivos. El taoísmo niega de forma explícita que las especies biológicas sean entidades fijas, y de hecho algunos filósofos taoístas especularon a cerca de como los atributos de los seres vivos diferían de acuerdo con los diferentes ambientes. En el tao todo lo que existe se encuentra en un estado de constante cambio, semejante a lo que pensó de forma independiente Heráclito, pero opuesto a la tradición que se impuso en el pensamiento occidental cristiano de las esencias de Platón (Jang, 2017; Shen, 2015; P. Zhang, 2016, 2018).
3.3 El pensamiento evolutivo en Roma y el mundo heleno
Tito Lucrecio Caro “99-55 AC” fue un filósofo griego que se oponía a los aristotélicos y estoicos en cuanto a la naturaleza de la materia, él era uno de los pocos atomistas. En su poema titulado De rerum Natura se preserva la mejor explicación sobreviviente de los filósofos de la escuela de Epicuro. En ella se describe el desarrollo del cosmos, la Tierra, los seres vivos y la sociedad humana a través de mecanismos naturales sin ninguna referencia a los dioses. Sin embargo, los epicúreos no representaban las líneas de pensamiento generales que se desarrollaban en el primer siglo antes de Cristo y que en los cuatro siglos siguientes se afianzarían en base a los estoicos. La revancha de la escuela epicúrea se desarrollaría con la ilustración después del siglo XVI (Ayala & Avise, 2014; Haldane, 1964; Hewes, 1993; Sweeney, 2002).

Figura 3.5. Marco Tulio Cicerón “106-43 AC”. Reconocido universalmente como uno de los más importantes autores de la historia romana, es responsable de la introducción de las más célebres escuelas filosóficas helenas en la intelectualidad republicana, así como de la creación de un vocabulario filosófico en latín. Gran orador y reputado abogado, Cicerón centró —mayoritariamente— su atención en su carrera política. Hoy en día es recordado por sus escritos de carácter humanista, filosófico y político.
A pesar de la visión de la cultura romana durante los primeros siglos de nuestro siglo como un grupo de disolutos y hedonistas la verdad era otra muy distinta. Aunque ciertas partes de la elite romana si se adecuaba este estereotipo la verdad es que muchos otros miembros de la elite romana eran muy diferentes. Un ejemplo de esto es Plinio el Viejo “23-79” quien veía en un simple beso entre marido y mujer en público como un acto contra la moralidad pública. Esta escuela extremadamente moralizante que pugnaba por el autocontrol fue denominada estoicismo. Los estoicos a parte de unos estándares morales muy altos también poseían una visión teológica y teleológica muy fuerte del mundo natural que posteriormente serían asumidos por el cristianismo. Algunos estoicos de importancia fueron Marco Tulio Cicerón “106-43 AC”, Lucio Anneo Séneca “4-65” y Plinio el viejo “23-79”. Los estoicos veían que la naturaleza intencionadamente producida seres vivos muy bien adaptados (Ayala & Avise, 2014; Bowler, 1992b; Fleischner, 2005).

Figura 3.6. Agustín de Hipona. onocido también como san Agustín (en latín, Aurelius Augustinus Hipponensis; Tagaste, 13 de noviembre de 354-Hipona, 28 de agosto de 430), es un santo, padre y doctor de la Iglesia católica. Después de su conversión, fue obispo de Hipona, al norte de África y lideró una serie luchas contra las herejías de los maniqueos, los donatistas y el pelagianismo.
En la línea del pensamiento griego en el siglo IV después de Cristo el obispo y teólogo cristiano posteriormente declarado Santo de la iglesia católica llamado San Agustín de Hipona "354-430" escribió que la historia del Génesis no debería ser leída de forma demasiado literal. En su libro De Genesi ad litteram “Sobre la interpretación literal del génesis” es que en algunos casos nuevas criaturas pudieron desarrollarse por descomposición de formas anteriores.
Para San Agustín los seres vivos han sido creados en un estado de imperfección y potencialidad para perfeccionarse en contraposición de entidades teológicamente perfectas como los ángeles, el firmamento y el alma humana. Según el profesor de teología de la Universidad Pontificia de Santa Cruz en Roma Giuseppe Tanzella-Nitti las ideas de San Agustín sobre el desarrollo lento de la vida puede sugerir o ser denominada una forma de pensamiento evolutivo (Greggs, 2009; Layton, 2004; Woods, 2009).
3.4 El pensamiento evolutivo en la edad media
A pesar de que las ideas evolutivas de los griegos y los romanos murieron con la caída del Imperio Romano, estas no se perdieron para la tradición islámica que preservó muchos de los textos originales griegos.
3.5 El islam
Los filósofos naturales del Islam vieron su apogeo entre los siglos VIII y XIII de nuestra era explorando varios tipos de ideas acerca de la filosofía natural, la alquimia y la historia natural. Estas ideas incluían la transmutación de lo vivo a partir desde lo no vivo, desde los minerales a las plantas, de las plantas a los animales, y desde los animales hacia el hombre. Resulta notable como al igual que sucede con el cristianismo, las ideas evolutivas del islam temprano no son vistas de forma peyorativa, o por lo menos de forma tan antagónica como se han venido a desarrollar en la actualidad. A continuación, se verán ejemplos de algunos filósofos islámicos de la edad media que reflexionaron a cerca de algunos problemas de la naturaleza relacionables con un tipo de razonamiento evolutivo. Bajo este contexto el pensamiento evolutivo hace referencia más al tipo de reflexión de Heráclito más que a la descendencia con modificación de Darwin, aunque como en todo hay que leer algunos casos realmente interesantes.
👉 La filosofía Mtazili
Al-Jāḥiẓ fue un escritor de prosa en árabe y autor de obras de literatura árabe, biología, zoología, historia, filosofía islámica, Mu'tazili Kalam (teología basada en la razón, la argumentación, el libre albedrío y el naturalismo. Además, fue el primero en sugerir la transformación de las especies animales, cuyo mecanismo se basaba en tres principios:
👉 La voluntad de Dios y Su Poder, El puede transformar una especie en otra cuando quiera.
👉 La lucha por la existencia, que para Al Jahiz es una ley decretada por Dios para preservar el equilibrio en la naturaleza.
👉 El efecto del clima, la alimentación y la adaptación de los animales a ambientes cambiantes debido a las migraciones, lo que provoca el uso y el desuso de los órganos.

Figura 3.7Abu Uthman Amr ibn Bahr al-Kinani al-Fuqaimi al-Basri “ al-Jāḥiẓ”. (Basora, hacia el 781-diciembre de 868 o enero de 869) fue un famoso intelectual árabe, del que se cree que tenía ascendencia afro-árabe de África Oriental.
👉 Primeras ideas sobre el efecto de las interacciones ecológicas
La obra más importante de Al-Jahiz es sin duda el Kitab al-Hayawan “el libro de los animales”. E texto aparentemente no va más allá de lo que ya sabían griegos como Aristóteles excepto por cierta reflexión notada por Conway Zirkle en 1941 mientras realizaba una reconstrucción histórica del concepto de selección natural. En la cita que se referencia mayoritariamente se describe lo que Darwin o Malthus describirían varios siglos después como la lucha por la existencia.
“La rata sale por su comida, y es hábil en obtenerla de aquellos animales más débiles que ella, del mismo modo, tiene que evadir a las serpientes y aves de presa que la buscan para devorarla debido a que son más fuertes de la rata.” “Los mosquitos saben instintivamente que la sangre es la cosa que los hace vivir, y cuando ven un animal saben que deben encontrar la piel para obtener su alimento. Del mismo modo las moscas cazan al mosquito que es la comida que ellas más desean, mientras que sus depredadores se alimentan de las moscas”. “Todos los animales, en resumen, no pueden vivir sin alimento, y al mismo tiempo no pueden evitar ser cazados para servir de alimento a otros. Todo animal débil devora al que es más débil que si mismo. Los animales fuertes no pueden escapar de ser devorados por animales que son más fuertes que ellos. Y en este respecto, los hombres no se diferencian de los animales, algunos con respecto a otros, aunque algunos no llegan a tantos extremos. En resumen, Alá ha dispuesto que algunos seres humanos sean la causa de la vida de otros, y del mismo modo, él ha dispuesto que los últimos sean la causa de la muerte de los primeros”.
Al-Jahiz también realizó descripciones de cadenas alimenticias (Ayoub, 2005; Bayrakdar, 1983; Shah, 2016; Stott, 2012b). Ibn Khaldun también conocido como Abū Zayd ‘Abdu r-Raḥmān bin Muḥammad bin Khaldūn Al-Ḥaḍrami “1332-1406” fue un filósofo e historiador bastante importante del norte de África. Algunos comentadores lo señalan como el padre fundador de la Sociología, y sus textos mencionan conceptos técnicos muy empleados por la sociología moderna. La relación de Khaldun con la evolución se relaciona con ciertos pasajes de su libro Mugaddimah donde afirma que los seres humanos se desarrollaron a partir del mundo de los monos mediante un proceso en el cual algunas especies se hacen más numerosas. En el capítulo I, Khaldun a diferencia de los filósofos naturales europeos señalaba la existencia de conexiones continuas entre diferentes tipos de seres mediante un patrón que es al mismo tiempo remarcable e interminable (Asghar, 2013; Asghar, Wiles, & Alters, 2010; Katsiaficas, 1999).
4. La escala natural en la edad media
4. La escala natural en la edad media
El pensamiento evolutivo no resultó particularmente evidente o convincente para la mayoría de los filósofos antiguos, y medievales por una razón particular, no se contaba con un registro relativamente completo de todas las especies vivientes que habitan en la tierra, y sus diversas y sobre todo graduales características físicas. fue sólo hasta la edad de los grandes imperios coloniales y de la exploración, que los naturalistas de los imperios coloniales tuvieron la posibilidad de empezar a registrar la enorme diversidad de los seres vivos del planeta, y sólo hasta entonces fue que la teoría evolutiva comenzó a emerger, aunque inicialmente de manera bastante tímida debido a las restricciones religiosas.
4.1 El conocimiento en la edad media
Durante la edad media la mayor parte de los trabajos de los griegos fueron perdidos para los académicos de occidente. Esto hace referencia a la división del imperio romano en dos, el imperio de Occidente que cayó entre los siglos IV-V y el imperio romano de oriente “también llamado imperio bizantino” que fue destruido en el siglo XV. Cuando los dos imperios se separaron, no solo rompieron en términos políticos, también lo hicieron en términos intelectuales y religiosos.
Adicionalmente Alejandría y otras ciudades de oriente que poseían una gran cantidad de centros intelectuales como la Gran Biblioteca cayeron en manos de los árabes islámicos. De esta forma tres bloques emergieron con diferentes políticas e ideas. El primero en caer fue el imperio romano de occidente y al hacerlo gran parte de sus logros académicos se perdieron, y eso sumado a la falta de “reabastecimiento” por parte de otras comunidades académicas en los otros imperios provocó que se perdiera el conocimiento. El conocimiento solo volvió a ser accesible a los europeos occidentales por dos rutas: la primera fue gracias a los contactos comerciales con los árabes que habían preservado y expandido lo que habían hecho los griegos. La segunda ruta fue la caída de Constantinopla y la destrucción del imperio romano de oriente, con lo cual muchos filósofos y académicos migraron a occidente en el siglo XV. De las dos rutas tal vez la más importante fue la primera ya que influyó mucho antes, permitiendo realizar copias europeas propias de los trabajos de filósofos clásicos como Aristóteles en el siglo XII. Eso sin mencionar que los filósofos naturales del Islam habían hecho aportes propios muy importantes en diversos campos del conocimiento como la historia natural, la filosofía natural, la alquimia y las matemáticas.

Figura 4.1. Durante la edad media sólo la ciencia y la filosofía musulmana podían compararse con la ciencia de China, pues Europa occidental se encontraba sumida en una edad oscura debido al colapso del imperio romano y la pérdida de gran parte del conocimiento clásico de los griegos.
Los pensadores cristianos que más pueden destacarse en la edad media son los escolásticos como Abelardo “1079-1142” y Santo Tomás de Aquino “1225-1274”, los cuales combinaros la clasificación aristotélica con las ideas de Platón, especialmente las que correspondían al esencialismo y la perfección de las entidades abstractas o ideales.
4.2 La gran cadena del ser en la edad media
De este modo el concepto de la Cadena del Ser o Escala Natural se convirtieron en parte del modo de pensamiento occidental, tanto así que aun hoy siguen permeando el discurso sobre la naturaleza. Como se mencionó en la sección de los griegos la cadena del ser postula que los seres vivos existen en una jerarquía. Esta jerarquía en la edad media se fusionó con las creencias cristianas, de este modo
👉 Dios quedaba en el trono superior y luego vienen los seres espirituales.
👉 Debajo de ellos los hombres y así sucesivamente la misma escala natural de los griegos.
En esta versión cristianizada del universo perfecto de Platón, las especies no podían cambiar, pues estaban fijas a un ideal creado por Dios mismo. Las criaturas de escalones cercanos se asemejarían las unas a las otras, lo cual se expresa en el vocablo latino: “natura non facit saltum”; o en español: la naturaleza no hace saltos. Como sistema de clasificación la cadena del ser se convertiría en uno de los fundamentos de la taxonomía Linneana (Artigas, Martnez, Glick, & Martínez, 2006; Carroll, 2000; Franklin, 1986; Witham, 2002).

Figura 4.2. La escala natural en la edad media. Representación de la gran cadena del ser mezclada con un esquema de "árbol", en la copa del árbol se encuentran Dios y los seres celestiales, luego están los santos, luego los hombres, y así sucesivamente hasta llegar a los seres inertes.
4.3 Las especies no cambian
Con el desarrollo de la Gran Cadena del Ser y el argumento de diseño por los teólogos cristianos la imagen de un mundo estático con categorías fijas, con límites claros se hizo bastante común en la cultura occidental. Algunos teólogos estuvieron más abiertos a la posibilidad de que el mundo fuera dinámico, y que este dinamismo dependiera de reglas definidas o también llamados procesos naturales.
Uno de esos teólogos fue Santo Tomas de Aquino quién es famoso por la distinción entre la causa primaria y las causas secundarias. Santo Tomás de Aquino insistió mucho más que San Agustín de Hipona en que los textos de las escrituras como Génesis no debían ser interpretados de una forma literal que fuera en conflicto o que impidiera a los filósofos naturales la comprensión del mundo natural. Santo Tomás sentía que la autonomía de la naturaleza era un signo de la bondad de Dios y que no había conflicto entre la concepción de una creación divina y de que el universo se desarrollara de forma dinámica, aunque en ese dinamismo la naturaleza en general tendría un fundamento teleológico, es decir una razón final y fundamental.
5. La escala natural durante la edad de la exploración
5. La escala natural durante la edad de la exploración
Durante la Edad del Renacimiento y la expansión de los grandes imperios coloniales, el concepto de la Gran Cadena del Ser o escala natural jugó un papel importante en la forma en que las personas entendían el mundo y su lugar en él.
5.1 Importancia cultural en el ánimo imperialista
La Gran Cadena del Ser era una idea que se remonta a la filosofía griega y se desarrolló más plenamente en la Edad Media y el Renacimiento. Sostenía que todos los seres vivos, desde los más simples hasta los más complejos, estaban organizados en una jerarquía ordenada, o "escala natural", con Dios en la cima y la materia inanimada en la parte inferior.
Esta idea tenía implicaciones importantes para la comprensión de la naturaleza y la relación entre los seres humanos y el mundo natural. Por ejemplo, algunos creían que los seres humanos eran superiores a los animales y otras formas de vida, y que estaban destinados a gobernar y dominar el mundo natural. Esta idea se utilizó a menudo para justificar la exploración y la colonización de nuevos territorios, así como para justificar la subyugación de los pueblos indígenas y la explotación de los recursos naturales.

Figura 5.1. "Redcoats" (casacas rojas en español) era el término que se utilizaba para referirse a los soldados británicos durante la época del Imperio británico. Los casacas rojas fueron utilizados como parte del esfuerzo del Imperio británico para expandir su dominio y explotar los recursos naturales en sus territorios colonizados. Su presencia y uso de la fuerza militar ayudó a establecer la autoridad británica en estas regiones y a proteger los intereses económicos y comerciales de Gran Bretaña.
Sin embargo, también había quienes cuestionaban la idea de la Gran Cadena del Ser y la jerarquía natural que implicaba. Algunos filósofos y científicos del Renacimiento comenzaron a cuestionar la autoridad de las enseñanzas tradicionales y buscaron una comprensión más científica y empírica del mundo natural. Esto sentó las bases para la revolución científica que se produciría más tarde en el siglo XVII.
En resumen, la idea de la Gran Cadena del Ser fue importante durante la Edad del Renacimiento y la expansión de los grandes imperios coloniales porque influyó en la forma en que las personas entendían la naturaleza y su relación con ella. Fue utilizado tanto para justificar la exploración y la colonización como para cuestionar la autoridad de las enseñanzas tradicionales y sentar las bases para la revolución científica (Deloria, 1997; Lovejoy, 2011; Pagden, 1986).
5.2 El renacimiento y la ilustración
No es objeto de este texto realizar un examen exhaustivo del cambio cultural que ocurrió paulatinamente en Europa posterior al siglo XV, sin embargo, vale la pena hablar de ciertas generalidades. Durante los siglos XV y XVI los contactos con las ciencias del islam y un paulatino cambio cultural se apoderaron de Europa. Varios eventos dieron lugar a este giro de la actitud, los que los historiadores citan con mayor frecuencia son: la caída de Constantinopla “1453”, el descubrimiento de América “1492”, la peste negra “1347-1353” y la invención de la imprenta “1450”.
👉 Los burgueses
El renacimiento y la ilustración también se enmarcan dentro del ascenso de una nueva clase social dominante, los comerciantes ricos de las ciudades “burgos” que en honor a estas fueron llamados burgueses, los cuales se habían originado en las últimas etapas de la edad media. Estos comerciantes influirían mucho en los modos de pensamiento y la práctica científica posterior. Los comerciantes estaban interesados en encontrar fuentes de capital prácticos, y en ese contexto los conocimientos de los griegos e islámicos se convirtieron en algo importante para la competencia capitalista sin reglas durante el renacimiento.
👉 La revolución científica
De este modo las ciencias abstractas de los griegos se amalgamaron con el espíritu empirista y aplicado desarrollado por los burgueses, lo cual generó el contexto propicio para que las ciencias modernas despegaran con la llegada de filósofos naturales como Leonardo da Vinci "1452-1519", Galileo Galilei “1564-1642”, Vesalius “1514-1564”, Paracelso “1493-1541” entre otros. De esta forma no es aventurado decir que la revolución científica fue fruto de la revolución social del capitalismo y la burguesía; y al mismo tiempo cuando la burguesía incluyó a la ciencia moderna en sus prácticas se generó un ciclo de crecimiento donde los burgueses invertían en ciencia, y esta generaba prácticas que generaban más riqueza.

Figura 5.2. René Descartes. también llamado Renatus Cartesius (en escritura latina) (La Haye en Touraine, 31 de marzo de 1596-Estocolmo, Suecia, 11 de febrero de 1650), fue un filósofo, matemático y físico francés, considerado como el padre de la geometría analítica y de la filosofía moderna, así como uno de los epígonos con luz propia en el umbral de la revolución científica.
5.3 El ser vivo como una máquina
Para la primera mitad del siglo XVII la historia natural se vio impregnada de una nueva idea, o mejor dicho una nueva analogía proveniente precisamente de los desarrollos tecnológicos de la época, la máquina.

Figura 5.3. El cuerpo-máquina. Leonardo da Vinci creó los primeros autómatas modernos empleando poleas, cables y palancas. La forma humanoide no es para nada azarosa en este contexto donde el cuerpo humano empezaba a ser visto bajo la perspectiva de una máquina muy compleja, pero al fin de cuentas una máquina que podía ser estudiada por los métodos de la filosofía natural y el naturalismo metodológico.
Ya Leonardo da Vinci “1452-1519” había creado estupendos artilugios mecánicos, los cuales incluían los primeros autómatas “robots”, por lo que la idea no era para nada nueva. Fue René Descartes “1596-1650” quien en base a su filosofía mecanicista propuso asumir que los cuerpos de los seres vivos incluyendo al ser humano son como máquinas, una metáfora que podía ser extendida al mismo universo. Este concepto caracterizaría a la revolución científica de la ilustración en los siglos XVI, XVII y XVIII. Dado que esta idea parte de una analogía física, Erns Mayr “1904-2005” asocia la analogía de la máquina a la postura filosófica que en la biología vendría a denominarse como fisicalismo, la cual también involucra la inclusión de las matemáticas en el pensamiento de los biólogos (Oldroyd, 1988).
5.4 Esencialismo
Entre los años de 1650 a 1800 varios autores se alinearon con el pensamiento de Heráclito de un universo dinámico que cambia y se desarrolla con el tiempo, de estos vale la pena destacar a Benoît de Maillet “1656-1738” quien apoyaba la idea de que el universo, la Tierra y la vida misma se habían desarrollado de forma mecánica sin una guía divina.

Figura 5.4. William Paley. (Peterborough, Northamptonshire, Inglaterra, 14 de julio de 1743 – 25 de mayo de 1805), fue un filósofo y teólogo utilitarista británico, recordado por su analogía del relojero y sus argumentos para demostrar la existencia de Dios en su obra Teología Natural, en especial el argumento teológico.
Sin embargo, eran excepciones, siglos de escolástica y de la fusión de la teología con las ideas de Platón y Aristóteles habían generado un impacto profundo en el pensamiento occidental, especialmente en lo que refiere al esencialismo y la teleología. De hecho, la mayoría de los filósofos que se especializaban realmente en la historia natural como los taxónomos estaban más a favor de que los seres vivos obedecieran a tipos perfectos creados al principio de los tiempos por Dios. Estos tipos podían ser clasificados en un orden estricto, lo cual permitía la generación de categorías bien diferenciadas. Este tipo de pensamiento tipológico es lo que permitió la creación de los sistemas taxonómicos de entre los cuales el que nos ha llegado es el sistema de Carlos Linneo “1707-1778” (Oldroyd, 1988).
5.5 Teleología y diseño inteligente
La teleología es la idea de que los seres vivos han sido creados con un propósito, esta idea se adjudica originalmente a Aristóteles, pero en el renacimiento encuentra sus defensores más fuertes dentro de los filósofos ingleses. De todos ellos el que ha saltado más a la celebridad histórica es William Paley “1743-1805” con su famosa analogía del reloj. La analogía del relojero se basa en la comparación de un fenómeno natural con un reloj. Normalmente se presenta como preludio para el argumento teleológico, a menudo de esta manera: Los complejos mecanismos del interior de un reloj requieren un diseñador inteligente. Al igual que el reloj, la complejidad de X (un órgano u organismo, la estructura del sistema solar, la vida, el universo, todo) requieren un diseñador. En esta presentación, la analogía del reloj no ejerce de premisa para un argumento, sino que sirve de recurso retórico y preámbulo. Su objetivo es establecer la viabilidad de la premisa general: mediante el simple hecho de observar algo, se puede saber si es o no un producto de diseño inteligente. El argumento se puede expresar del siguiente modo:
“Al observar un mecanismo tan sencillo como un reloj a nadie se le ocurre dudar que éste es el producto de una creación, que es el resultado de un trabajo intencional. A ninguna persona en su sano juicio se le puede ocurrir pensar que un mecanismo como el del reloj, con sus engranajes dentados, su solenoide y su bobina dispuestos de manera precisa entre sí para funcionar y medir el tiempo es consecuencia de una sucesión de casualidades que, progresivamente, han ido dando forma a sus partes y que, además, han dado con el acople entre sí de dichas partes para dar con la función deseada.
¡Nadie que no esté loco puede pensar que un reloj es consecuencia del azar! Así pues, ¿quién puede pensar que un organismo como el humano, mucho más complejo que el de un reloj, es producto del azar? A ninguna persona razonable se le puede ocurrir negar que todo ser vivo, con sus partes dispuestas entre sí idóneamente, cada una cumpliendo su función, su finalidad, interdependientes entre sí es el producto de un artesano sumamente hábil y poderoso que nos concibió. Nadie en su sano juicio puede dudar que somos criaturas de Dios.” (Denton & Scott, 1986)
6. Los primeros evolucionistas
6. Los primeros evolucionistas
Pensamiento evolutivo, evolución esto, evolucionismo aquello, manoseamos mucho una palabra en épocas remotas que no fue acuñado sino hasta 1762, antes de esto las concepciones de un universo dinámico no serían concebibles con la palabra evolución por sus protagonistas históricos. Es por esto que durante los artículos anteriores hemos preferido emplear expresiones como universo dinámico o pensamiento de Heráclito para describir la idea de cosas que cambian con el tiempo en oposición al pensamiento esencialista. La palabra evolución viene de la raíz latina Evolutio que significa desenvolver un pergamino, en otras palabras, casi como todo en bilogía es una analogía.

Figura 6.1. Charles Bonnet (Ginebra, 13 de marzo de 1720-20 de mayo de 1793) fue un biólogo y filósofo suizo. Fue uno de los principales exponentes de la idea de Scala naturae y fue autor de importantes descubrimientos biológicos, como la partenogénesis.
6.1 Charles Bonnet
La analogía en este caso hace referencia a expandir lo que está contenido como un potencial. Este desarrollo de un potencial “evolutio” fue aplicado originalmente en 1762 por Charles Bonnet “1720-1793” para ser aplicado a las ideas preformacionistas de la herencia. En palabras modernas lo más aproximado es la palabra desarrollo. En otras palabras, evolutio se aplicó para describir el desarrollo embrionario de los seres vivos, pero con el tiempo fue ganando más y más generalidad (Bowler, 1975; Opitz & Reynolds, 1985).
6.2 El conde de Buffon
Posteriormente en el siglo XVIII el filósofo francés Georges Louis Leclerc, conde de Buffon “1707-1788”, uno de los filósofos naturales más influyentes de su tiempo, sugirió que lo que la mayoría de las personas se referían como especies tipológicamente distinguibles no eran más que variedades bien definidas al interior de un grupo más general, las cuales se habían modificado debido a factores ambientales. En otras palabras, tomando la taxonomía de Linnaeus “1739-1778” sugería que los grupos en los que se clasificaban los seres vivos habrían tenido una existencia como si se tratara de un ancestro común tipológico. Posteriormente especuló que las aproximadamente 200 especies de mamíferos conocidos habrían descendido de unas 38 formas tipológicas independientes. Las ideas evolutivas de Buffon eran limitadas, el creía que cada forma original se había formado mediante generación espontánea, siendo estos moldes tipológicos a los que los seres vivos se asemejaban parcialmente. Los trabajos de Buffon buscaban explicar a la naturaleza mediante mecanismos naturalistas, cuestionando la idea de que todas las especies fueran entidades fijas e inmutables. Su trabajo influiría en otros filósofos e historiadores naturales en los tiempos por venir (Farber, 2003; Gayon, 1996).

Figura 6.2. Georges Louis Leclerc, conde de Buffon (Montbard, Borgoña, 7 de septiembre de 1707 – París, 16 de abril de 1788) fue un naturalista, botánico, matemático, biólogo, cosmólogo y escritor francés. Buffon pretendió compendiar todo el saber humano sobre el mundo natural en su obra en 44 volúmenes Histoire naturelle. Su enfoque influyó en la Enciclopedia de Diderot y sus ideas también lo hicieron sobre las siguientes generaciones de naturalistas y en particular sobre Lamarck, Cuvier y Darwin.
7. La teoría lamarckiana de la evolución
7. La teoría lamarckiana de la evolución
Jean-Baptiste Pierre Antoine de Monet, caballero de Lamarck “1744-1829” propuso en su obra Philosophie Zoologique “filosofía zoológica” de 1809 una nueva y radical teoría sobre la evolución de las especies. Lamarck no creía que todas las especies compartieran un solo ancestro común, aunque se alineó con Buffon en el sentido de que algunos grupos de especies altamente relacionados si debieron descender de un ancestro. Hasta aquí nada nuevo, el punto de Larmarck es su énfasis en el proceso que permitía esta evolución. Para Lamarck existían fuerzas vitales que influían en los seres vivos en la medida en que estos ejercían su lucha por la existencia. La teoría de Lamarck tiene cuatro componentes principales, la complejización, la adaptación, el uso, el esfuerzo y la herencia (Burkhardt, 2013; Gayon, 1996; Lamarck, 2013).

Figura 7.1. Jean-Baptiste Pierre Antoine de Monet, caballero de Lamarck (Bazentin, 1 de agosto de 1744-París, 18 de diciembre de 1829) fue un naturalista francés, uno de los grandes hombres de la época de la sistematización de la Historia Natural, cercano en su influencia a Linneo, Leclerc y Cuvier. Lamarck formuló la primera teoría de la evolución biológica, en 1802 acuñó el término «biología» para designar la ciencia de los seres vivos y fue el fundador de la paleontología de los invertebrados.
7.1 La fuerza de complejización
Lamarck así como otros naturalistas observaban que la naturaleza poseía una tendencia hacia a hacerse más complejos. Esta tendencia sin embargo aparece solo si se tiene en mente que la teoría sobre el orden de la naturaleza que prevalecía en su tiempo es la Gran Cadena del Ser. Bajo la Gran Cadena del Ser en donde se coloca al ser humano en la cúspide de los seres vivos se observaría una tendencia hacia la complejización. En esta idea se anticipa una especie de combinación entre las ideas evolutivas del pensamiento de Heráclito con la idea de la Gran Cadena del Ser como si los seres vivos de un escalón inferior tendieran a subir de escalón con el tiempo.
¿Qué hace que los seres vivos tienden a subir la escalera de la organización?, Lamarck lo intentó empleando una analogía de las ciencias físicas, especialmente al concepto de fuerzas intangibles. A pesar de que las fuerzas intangibles no parecen un concepto muy científico una se había vuelto vital en las explicaciones sobre el mundo natural y era la gravedad. La gravedad era una fuerza intangible, un fenómeno que ejercía fuerzas a distancia sin contacto entre los dos objetos y fue matematizada en ecuaciones por sir Isaac Newton “1642-1727”.
7.2 La fuerza vital en Lamarck
Lamarck creía que la fuerza que el trataba de determinar tenía una naturaleza semejante a la gravedad, un intangible que afectaba de manera específica a los seres vivos y no afectaba a los inertes más que para crear la generación espontánea. La idea de la fuerza vital no era nueva, pero se hizo muy popular entre los historiadores naturales ya que permitía explicar la adaptación de los seres vivos y el origen de la vida mediante una “ley natural” y al mismo tiempo explicaría la teleología. La fuerza vital haría evolucionar a los seres vivos hacia fines determinados “teleología”. Siendo una fuerza semejante a la gravedad y por lo tanto una explicación naturalista. De Lamarck también cabe destacar que pensaba que tanto la historia natural “lo que llamamos biología” y la alquimia “lo que llamarías química” se encontraban altamente relacionadas. En esta visión del ascenso de la escalera, los seres vivos simples que se hacían más complejos eran reemplazados por otros seres vivos simples que eran creados por generación espontánea. Finalmente se debe resaltar que la fuerza vital de Lamarck tenía más un principio mecanicista o físico que un sentido espiritual.
7.3 La fuerza de adaptación
La segunda parte de la teoría de la evolución de Lamarck concernía a la adaptación de los seres vivos a su medioambiente. Desde las épocas de Aristóteles y aun antes los conceptos de la lucha por la existencia, la competencia y la depredación fueron más que evidentes aun para cualquier cazador o campesino iletrado, son fenómenos más que evidentes aun cuando nos parezcan crueles. El punto es que los seres vivos parecen haber sido creados para estar adaptados a ciertos problemas o necesidades para su subsistencia de una forma altamente compleja e integrada, este fenómeno fue lo que llevó a Aristóteles a proponer el concepto de teleología. Para Lamarck la fuerza vital conectaba a los componentes de los seres vivos que más eran empleados en su interacción con el medioambiente. Aquellas partes que más se empleaban se fortalecían y esa fortaleza sería la clave de su teoría solo si podía ser heredable a las siguientes generaciones.
7.4 Las leyes de Lamarck
No estamos muy acostumbrados a aprendernos leyes naturales de teorías fallidas o anticuadas, sin embargo, en su contexto eran principios naturales aceptados por una gran cantidad de miembros de las comunidades científicas de la época. Entender cómo operan estas leyes servirá como base para poder contrastar como es que operan las leyes de la naturaleza de las teorías modernas. Lamarck desarrolló su teórica mediante dos leyes. Las leyes de Lamarck a pesar de lo que muchos creen no serían eliminadas por los trabajos de Darwin y Wallace, y perdurarían casi hasta el final del siglo XIX hasta que biólogos experimentales como August Weismann “1834-1914” las refutaran mediante experimentos controlados. Sin embargo, algunos rasgos de la segunda ley de Lamarck aún perduran en una nueva rama de la biología llamada epigenética aunque de un modo mucho más sutil.
👉 La primera ley de Lamarck, de uso y desuso
La primera le dé Lamarck dice que cada animal que no ha pasado el límite de su desarrollo puede fortalecer un órgano mediante su uso continuo. La cantidad de esfuerzo colocado en dicho órgano es directamente y es proporcional al poder y tamaño que este va adquiriendo.

Figura 7.2. El lamarckismo es lineal y perfeccionante. Lo bello de la teoría lamarckiana es que concordaba con la idea de la Gran Cadena del Ser y con la telología de Aristóteles, haciendo de la evolución un procedo lineal, unidireccional y orientado a un propósito adaptativo.
Por otro lado, los órganos que no se emplean durante el desarrollo del ser vivo se hacen débiles y más pequeños con el tiempo. Si haces ejercicio fortaleces determinados órganos, si no caminas los músculos de tus piernas se atrofian. Estos fenómenos eran bien conocidos desde la antigüedad, por lo que la primera ley de Lamarck no era más que una descripción de lo que todo el mundo sabía. El punto era lograr una explicación de órganos claramente hipertrofiados en algunos seres vivos en comparación con otros de tipos semejantes, por ejemplo, la jirafa con respecto a sus ancestros de cuello corto o en comparación a sus semejantes de cuello corto como el okapi. La primera ley de Lamarck en este contexto sería inútil sin una teoría de la herencia, la cual se enuncia en su segunda ley.
👉 La segunda ley de Lamarck, la herencia blanda
La segunda ley de Lamarck hace referencia a la herencia y es de hecho una teoría propia sobre la herencia. En esta se plantea que toda la ganancia del uso de los órganos, así como toda la perdida por órganos no usados puede transmitirse a la progenie, de esta forma en varias generaciones un órgano muy empleado puede crecer sin límites determinados. A esta teoría de la herencia se la denomina herencia blanda, debido a que los factores reproductivos son blandos y pueden ser influenciados por los cambios en el cuerpo general del ser vivo. Su teoría opuesta también fue desarrollada en el siglo XIX y se denominó herencia dura, en esta los factores reproductivos son duros y no pueden ser alterados por modificaciones en el cuerpo general del ser vivo. De las dos la herencia dura ganó predominancia con los trabajos de A. Weismann de los mendelianos en la última década del siglo XIX como Carl Correns.
8. Definiciones de Darwinismo
8. Definiciones de Darwinismo
Darwin y el darwinismo, son dos vocablos que tienden a emplearse de forma alegre sin desarrollar una definición clara, es por esto que con el tiempo se han transformado en expresiones polisémicas con multitud de significados. Por lo anterior se repasarán algunas definiciones comunes del vocablo darwinismo, poniendo especial énfasis en el que se asumirá de aquí en adelante. El lector debe recordar que sin una definición clara u homogénea sobre un concepto es imposible generar una discusión y una crítica clara de un concepto determinado.
8.1 El darwinismo como sinónimo de evolución
En un sentido general, y al mismo tiempo siendo una simplificación exagerada y por muchas razones errónea, se asume que el darwinismo es lo mismo que evolución en general. Como hemos visto en artículos precedentes Darwin no se inventó el concepto de evolución de la nada, y tampoco fue el primero en acuñar el termino evolutio “desenrollar o desenvolver” para aplicarlo en la historia natural. Darwin simplemente dio una explicación a la evolución que inclusive no es la misma que se maneja por las comunidades científicas en la actualidad, de Darwin lo que aún perdura y con grandes limitaciones es el concepto de selección natural.
8.2 El darwinismo como gradualismo filetico
La segunda definición de Darwinismo se corresponde a una línea de pensamiento al interior de las comunidades científicas sobre los ritmos o velocidades con que los seres vivos cambian a lo largo del registro geológico. En este contexto darwinismo hace referencia a un cambio gradual y lento, a lo cual se lo denomina de forma más formal como el gradualismo filetico. La teoría opuesta al gradualismo filetico es el equilibrio puntuado de Gould y Eldredge. Sin embargo, estas discusiones se enmarcan al interior de la teoría sintética de la evolución en la segunda mitad del siglo XX por lo que no serán abarcadas en el contexto de Darwin.
8.3 El darwinismo como sinónimo de selección natural
Una tercera acepción de darwinismo es a la corriente de pensamiento al interior de las comunidades científicas que la única o principal fuente de cambio morfológico es la selección natural. Si bien existen algunos científicos de esta opinión, con la introducción de la teoría sintética de la evolución en la primera mitad del siglo XX esta concepción comenzó a ser muy criticada. En ese contexto la selección natural paso de ser la reina en las explicaciones a ser solo uno mecanismo más entre otros. Adicionalmente se desarrollaron instrumentos estadísticos que sirven para distinguir entre la selección natural y otras fuerzas evolutivas como la deriva genética. Adicionalmente una de las subteorías que nacieron del desprendimiento de la selección natural como fuerza evolutiva reinante fue la Teoría Neutral de la Evolución Molecular.
8.4 El darwinismo como sinónimo de naturalismo metodológico
Como explicación naturalista se refiere a una explicación naturalista de la evolución, para diferenciarla de las que ocurren a elementos o causas teleológicas de orden no natural. En tal caso el darwinismo se vuelve un sinónimo del concepto de naturalismo metodológico que será discutido en próximos artículos. Tal vez lo más importante de este aspecto es la firme oposición del darwinismo a la postura filosófica de los vitalistas. En este sentido filosóficamente se opone a la presunción de fuerzas invisibles que actúan sobre los seres vivos con conciencia o premeditación. Es importante recalcar que muchas veces se habla de selección natural o de evolución en general como una fuerza vitalista, pero esto es solo una extensión del lenguaje que tergiversaría las conclusiones a las que han llegado en la actualidad las comunidades científicas.
8.5 El darwinismo como sinónimo de ciencias que estudian el cambio
Como sinónimo de las ciencias que estudian a la naturaleza desde la perspectiva de Heráclito, es decir como entidades en un devenir constante, en un constante cambio. Ya hemos visto como el pensamiento de Heráclito es fundamental, pues el asume que las cosas cambian con el tiempo siempre. De este modo su se asumen ambos como sinónimos ocurre que problemas científicos tan dispares como:
👉 El origen y desarrollo del universo, las galaxias y los planetas.
👉 El origen y desarrollo de la vida.
👉 El origen y desarrollo de las especies.
👉 El origen y desarrollo de la conciencia.
👉 El origen y desarrollo de las comunidades humanas.
Pasarían a ser entendidas todas bajo el sinónimo de darwinismo, como si fuera un paradigma. Este “darwinismo en sensu lato” es bastante problemático ya que extiende mucho las críticas o discusiones en un debate. Es por esta razón que en este blog la expresión de Darwinismo siempre hará referencia al gradualismo filetico o a la primera teoría de la evolución en su contexto de tiempo "darwinismo en sensu stricto".
9. Los tiempos de Darwin y Wallace
9. Los tiempos de Darwin y Wallace
Ernest Rutherford trabajó en física, con métodos y cálculos físicos… le dieron el premio Nobel de química (…), enojado proclamó unas palabras que ofenderían amor propio de muchos científicos y disciplinas a lo largo del planeta “la única ciencia real es la Física, lo demás es filatelia”. La filatelia es la afición a coleccionar sellos y sobres postales, y esta comparación es más que pertinente para los orígenes de la taxonomía, y de la biología misma.

Figura 9.1. Compañeros, rivales, mentor y aprendiz. Charles Darwin “izquierda” y Alfred Russell Wallace “derecha” protagonizaron uno de los conflictos de prioridades científicas más famosos, y tal vez, el mejor ejemplo de caballerosidad y camaradería más grandes de la historia de la ciencia, por el mérito del concepto de Selección Natural.

Figura 9.2. Una ciencia de colecciones. Sin una visión medianamente completa de la diversidad biológica, y especialmente, sin estar expuestos al problema de las especies complejas, es impensable entender el origen de las teorías evolutivas modernas de corte darwiniano.
La Historia Natural por milenios fue de hecho una filatelia, un arte de coleccionistas, de hecho, el propio Charles Darwin se inició por los caminos de la historia natural recolectando insectos del campo, catalogándolos y organizándolos en colecciones.

Figura 9.3. El Imperio Británico. El Imperio Británico hasta el final de la guerra de independencia norteamericana, era tan extenso que se decía que, en él, el Sol nunca se ponía. A parte de la sensación de orgullo y optimismo por su extensión, el imperio dependía de la exploración marítima.
Las clasificaciones de los seres vivos debían obedecer a los cánones establecidos por dos autores clásicos, que filosofías opuestas, Platón y Aristóteles. De Platón se tomaba la Gran Cadena del Ser, concepto que ya hemos discutido con anterioridad. En la gran cadena del ser todos los seres vivos están dispuestos en una organización teleológica y escalar, en el extremo superior se encuentra Dios/dioses, luego los seres celestiales, luego el hombre, por debajo la mujer y por debajo las demás criaturas vivientes del mundo. De Aristóteles se tomó la noción de dividir al mundo natural en tres grandes grupos o reinos, el Vegetal, el Animal y el Mineral. Nuevamente asumimos al mineral como un reino en toda regla ya que muchos autores de la época de Linnaeus aun abogaban por la Alquimia, donde se consideraba a los materiales como el oro, la plata o los cristales preciosos como entidades vivas.

Figura 9.4. Especie compleja o complejo de especies. En un complejo de especies, distinguir entre razas y especies es difícil, lo cual indica que el proceso de conversión entre razas a especies es gradual.
A pesar de ser Filatelia, las colecciones naturales de seres vivos poseían un enorme poder, el conocimiento, al saber las especies de animales y vegetales de sus territorios, los gobernantes podían estimular los cultivos de ciertas especies en ciertas zonas de sus territorios. Una planta encontrada en una colonia pequeña o una exploración aventurera podía ser llevada a una colonia más estable y grade, con mayor población y clima semejante para explotarla. Sin embargo, la organización era clave, reconocer que la planta que era llevada a la corte era la misma enviada a las colonias para el cultivo era la misma se hizo preponderante, de allí surgió la necesidad de crear un sistema de clasificación, reconocimiento y nombramiento de las especies naturales. El método de clasificación que más impacto ha tenido fue establecido en el siglo XVIII por Carlos Linnaeus, el cual rápidamente lo convirtió en la autoridad más importante de Europa en términos de Historia Natural.
9.1 Contexto histórico de Charles Darwin y Alfred R. Wallace
El contexto histórico de Charles Darwin “1809-1882” y Alfred Russel Wallace “1823-1913” corresponde la Inglaterra Victoriana, es decir al imperio británico del siglo XIX. La Inglaterra de esta época era muy diferente a la actual, pues a pesar de haber perdido su colonia en américa “actualmente los Estados Unidos”, aún seguía poseyendo una gran cantidad de colonias, como la India. Era un imperio marítimo y colonial, lo cual le aseguraba una gran cantidad de vienes alrededor del mundo. La Inglaterra de esta época era muy diferente a la actual, pues a pesar de haber perdido su colonia en américa “actualmente los Estados Unidos”, aún seguía poseyendo una gran cantidad de colonias, como la India. Era un imperio marítimo y colonial, lo cual le aseguraba una gran cantidad de vienes alrededor del mundo.
Estos bienes no solo eran económicos, también correspondían a especímenes raros de plantas y animales que eran recolectados por los historiadores naturales a bordo de los barcos del rey. Una buena descripción de la relación de los historiadores naturales puede verse en la película “Master and Commander: Al otro lado del mundo, 2003” en forma del doctor Stephen Maturin “Paul Bettany”. Darwin y Wallace fueron historiadores naturales comisionados en viajes alrededor del mundo, lo cual fue crítico para poder desarrollar sus teorías evolutivas. Los historiadores naturales seleccionaban especímenes raros que eran enviados a la metrópolis imperial, en este caso Londres para ser analizados en universidades y/o ser expuestos en museos para exponer ante todos, la magnificencia, poder y habilidad del Imperio Británico y sus hombres.

Figura 9.5. Una película para entender a Darwin y Wallace. Master and Commander nos sitúa en el contexto de los (B) historiadores naturales del siglo XIX comisionados en la marina británica, compañeros de los capitanes de clases nobles, los Lords británicos (B), siendo gente ilustrada al mando de marinos de clases bajas.
La Inglaterra del siglo XIX también es famosa por un aspecto, la revolución industrial, el empleo de las tecnologías desarrolladas por los físicos gracias a las ecuaciones de Newton y sus seguidores. Esto le otorgó al imperio un súbito incremento en su productividad y riqueza, aunque al mismo tiempo generando un desequilibrio social del cual se queja Malthus en su ensayo sobre el Principio de las Poblaciones. Básicamente los ricos se hicieron más ricos, los pobres más pobres y su número se incrementó, las poblaciones de las ciudades se incrementaron masivamente, lo cual generó problemáticas sociales. Para tener una idea de la vida en las calles de la Londres de esta época solo basta con leer o ver alguna película sobre Jack el destripador, una historia que se centra en los barrios bajos de Londres y de un asesino en serie que mataba prostitutas. Otra serie de relatos importantes sobre este periodo histórico son las novelas de Sherlock Holmes de Sir Arthur Conan Doyle “1859-1930”.

Figura 9.6. La ciencia y la sociedad británica. Con las novelas de sir Arthur Conan Doyle sobre el detective Holmes nos encontramos con una irrupción del método científico en actividades más cercanas a la sociedad, el método hipotético deductivo comenzó a emplearse en el siglo XIX en la criminalística y los análisis forenses.
Un último aspecto corresponde a la ciencia, la cultura británica a pesar de aferrarse a una serie de valores altamente moralistas tradicionales, abrazó con gran fervor las novedades en ingeniería e industria, prueba de ello fue que en el año de 1851 se realizará en Londres la Gran Feria Mundial, donde 34 naciones alrededor del mundo expusieron lo último en avances científicos así como las expectativas sobre las tecnologías del mañana. Gran parte de la ciencia escolar que es enseñada en el bachillerato fue propuesta durante esta época, no solo por científicos ingleses, también por sus competidores en Rusia, Francia, Estados Unidos, Austria y Prusia.
Solo para nombrar algunos nombres notables que cualquier escolar con buena memoria reconocería son: Amadeo Avogadro “1776-1856, Italia”, Ludwig Boltzmann “1844-1906, Austria”, Marie Curie “1867-1934, Polonia-Francia”, Pierre Curie “1859-1906, Francia”, Gregor Mendel “1822-1884, Austria”, Carl Correns “1864-1933, Alemania”, Christian Doppler “1803-1853”, Thomas Edison “1847-1931, Estados Unidos”, Nikola Tesla “1856- 1943, Croacia-Estados Unidos”, Michael Faraday “1791-1867, Inglaterra”, Carl Friedrich Gauss “1777-1855, Alemania”, Josiah Willard Gibbs “1839-1903, Estados Unidos”, Ernst Haeckel “1834-1919, Prusia-Alemania”, Heinrich Rudolf Hertz “1857-1894, Prusia-Alemania”, Friedrich Wilhelm Heinrich Alexander von Humboldt “1769-1859, Prusia”, James Clerk Maxwell “1831-1879, Escosia”, Dmitri Ivanovich Mendeleev “1834-1907, Rusia”, Alfred Bernhard Nobel “1833-1896, Suecia”, Luis Pasteur “1822-1895, Francia”, Santiago Ramón y Cajal “1852-1934, España”, William Thomson primer Barón de Kelvin “1824-1907”.

Figura 9.7. Una guerra a nivel científico. De la combinación de la mecánica y la química nacieron armas más poderosas con mayor alcance. Cañones y ametralladoras, bombarderos y tanques. De aquí en adelante las ciencias de la naturaleza y en especial la física serían vistas como parte de la defensa estratégica de las potencias, cuestión que se materializaría varias décadas después en la segunda guerra mundial con el proyecto Mahattan.
No por nada algunos autores llaman al siglo XIX como un maravilloso siglo científico, una época en que el idealismo por las ciencias se elevaría a sus más altos confines de la esperanza humana, solo para ser aplastado por la barbarie que se impondría con el siglo XX con la primera guerra mundial “1914-1918”, momento en el cual casi todas las tecnologías desarrolladas para esa época seria empleadas para la destrucción.
9.2 Charles Darwin
(1809-1882) Fue un naturalista, geólogo y biólogo inglés, mejor conocido por sus contribuciones a la ciencia de la evolución. Estableció que todas las especies de vida han descendido a lo largo del tiempo de antepasados comunes y, en una publicación conjunta con Alfred Russel Wallace, introdujo su teoría científica de que este patrón de ramificación de la evolución resultó de un proceso que él llamó selección natural, en el cual la lucha por la existencia tiene un efecto similar a la selección artificial involucrada en la reproducción selectiva (Darwin, 1993, 2018).
👉 Obra magna
Darwin publicó su teoría de la evolución con convincente evidencia en su libro de 1859 Sobre el origen de las especies, superando el rechazo científico de conceptos anteriores de transmutación de especies. En la década de 1870, la comunidad científica y la mayoría del público educado habían aceptado la evolución como un hecho. Sin embargo, muchas explicaciones competitivas favorecidas y no fue hasta la aparición de la síntesis evolutiva moderna desde la década de 1930 hasta la década de 1950 que se desarrolló un amplio consenso en el que la selección natural era el mecanismo básico de la evolución. El descubrimiento científico de Darwin es la teoría unificadora de las ciencias de la vida, que explica la diversidad de la vida (Darwin, 1993, 2018).
👉 Viajes
El interés temprano de Darwin en la naturaleza lo llevó a descuidar su educación médica en la Universidad de Edimburgo; en su lugar, ayudó a investigar invertebrados marinos. Los estudios en la Universidad de Cambridge (Christ's College) alentaron su pasión por las ciencias naturales. Su viaje de cinco años en el HMS Beagle lo estableció como un eminente geólogo cuyas observaciones y teorías respaldaban las ideas uniformadas de Charles Lyell, y la publicación de su diario del viaje lo hizo famoso como autor popular (Darwin, 1993, 2018).
Desconcertado por la distribución geográfica de la vida silvestre y los fósiles que recopiló en el viaje, Darwin comenzó investigaciones detalladas, y en 1838 concibió su teoría de la selección natural.
👉 Otras obras
Aunque discutió sus ideas con varios naturalistas, necesitó tiempo para una extensa investigación y su trabajo geológico tuvo prioridad. Estaba escribiendo su teoría en 1858 cuando Alfred Russel Wallace le envió un ensayo que describía la misma idea, lo que provocó la publicación conjunta inmediata de ambas de sus teorías. El trabajo de Darwin estableció a la descendencia con modificación como la explicación científica dominante de la diversificación en la naturaleza. En 1871 examinó la evolución humana y la selección sexual en El origen del hombre, y Selección en relación con el sexo, seguida de La expresión de las emociones en el hombre y los animales (1872). Darwin ha sido descrito como una de las figuras más influyentes en la historia de la humanidad, y fue honrado por el entierro en la Abadía de Westminster (Darwin, 1993, 2018).
9.3 Alfred Russell Wallace
(1823-1913) Fue un naturalista, explorador, geógrafo, antropólogo y biólogo inglés. Él es mejor conocido por concebir de forma independiente la teoría de la evolución a través de la selección natural; su artículo sobre el tema fue publicado conjuntamente con algunos de los escritos de Charles Darwin en 1858. Esto llevó a Darwin a publicar sus propias ideas en Sobre el origen de las especies.
👉 Viajes
Wallace hizo un extenso trabajo de campo, primero en la cuenca del río Amazonas y luego en el archipiélago malayo, donde identificó la división de fauna que ahora se denomina Línea de Wallace, que separa el archipiélago indonesio en dos partes distintas: una parte occidental en la que los animales son en gran parte Origen asiático, y una porción oriental donde la fauna refleja especies de Australasia (Brackman, 1980; Raby, 2002; Shermer, 2002; Smith & Beccaloni, 2010; Williams-Ellis, 1966).
Fue considerado el principal experto del siglo XIX en la distribución geográfica de las especies animales y, a veces, se le llama el "padre de la biogeografía". Wallace fue uno de los principales pensadores evolutivos del siglo XIX y realizó muchas otras contribuciones al desarrollo de la teoría evolutiva además de ser co-descubridor de la selección natural. Estos incluyeron el concepto de coloración de advertencia en animales, y el efecto Wallace, una hipótesis sobre cómo la selección natural podría contribuir a la especiación fomentando el desarrollo de barreras contra la hibridación. El libro de Wallace de 1904 El lugar del hombre en el universo fue el primer intento serio de un biólogo de evaluar la probabilidad de vida en otros planetas. También fue uno de los primeros científicos en escribir una exploración seria sobre el tema de si había vida en Marte
👉 Idealista filosófico
Wallace se sintió fuertemente atraído por ideas no convencionales (como la evolución). Su defensa del espiritismo y su creencia en un origen inmaterial para las facultades mentales superiores de los humanos tensaron su relación con algunos miembros la comunidad científica.
👉 Activismo social
Además del trabajo científico, era un activista social que criticaba lo que consideraba un sistema social y económico injusto (capitalismo) en la Gran Bretaña del siglo XIX. Su interés en la historia natural resultó en que él sea uno de los primeros científicos prominentes en expresar su preocupación por el impacto ambiental de la actividad humana.
👉 Obras
También fue un autor prolífico que escribió sobre cuestiones científicas y sociales; su relato de sus aventuras y observaciones durante sus exploraciones en Singapur, Indonesia y Malasia, The Malay Archipelago, fue popular y muy respetado. Desde su publicación en 1869, nunca se agotó.
👉 Financiamiento
Wallace tuvo dificultades financieras durante gran parte de su vida. Sus viajes por el Amazonas y el Lejano Oriente fueron apoyados por la venta de especímenes que recolectó y, después de perder la mayor parte del considerable dinero que obtuvo de esas ventas en inversiones fallidas, tuvo que apoyarse principalmente en las publicaciones que produjo. A diferencia de algunos de sus contemporáneos en la comunidad científica británica, como Darwin y Charles Lyell, no tenía ningún patrimonio familiar para recurrir, y no tuvo éxito en encontrar un puesto asalariado a largo plazo, no recibiendo ingresos regulares hasta que se le concedió una pequeña pensión del gobierno, a través de la influencia de Darwin, en 1881.
9.4 Importancia de pertenecer al imperio británico
La pertenencia al Imperio Británico tuvo un impacto significativo en el trabajo de Charles Darwin y Alfred Russel Wallace, los co-descubridores de la teoría de la evolución por selección natural.
En el caso de Darwin, su experiencia de viaje en el HMS Beagle a través de América del Sur y el Pacífico Sur, pues se trataba de un barco militar, que al viajar con bandera imperial, le aseguraba toda clase de contactos en los puertos a los que llegaba, así como comodidades para realizar incursiones en tierra como buenos caballos y los mejores guías para investigar las áreas de importancia científica. La pertenencia al impero también le aseguró contactos con otros científicos y naturalistas de todo el mundo, por lo que su obra seria insertada inmediatamente en la comunidad científica más importante. Además, la filosofía social y política del imperialismo británico influyó en el desarrollo de las ideas de Darwin, especialmente en su idea de la lucha por la existencia y la selección natural, que se basó en gran medida en la competencia y el conflicto que observó en las sociedades humanas y en la naturaleza.
Por su parte, Wallace también viajó extensamente en América del Sur y el sudeste asiático, recolectando muestras y observando la fauna y la flora. Su experiencia en las colonias británicas de Asia, en particular, lo llevó a considerar la importancia de la competencia y la selección natural en la evolución de las especies, y su trabajo fue financiado en gran parte por sociedades científicas y empresas británicas.
En resumen, la pertenencia al Imperio Británico brindó a Darwin y Wallace las oportunidades y los recursos necesarios para desarrollar sus ideas sobre la evolución y la selección natural, y la filosofía social y política del imperialismo británico influyó en sus concepciones sobre la competencia y el conflicto en la naturaleza (M. A Largent, 1995)
10. El viaje de Darwin
10. El viaje de Darwin
En el siglo XVIII y principios del XIX, los naturalistas europeos se centraban en la descripción y clasificación de las especies que se encontraban en Europa, que tenía una biodiversidad relativamente baja en comparación con otras partes del mundo, como América del Sur y el sudeste asiático. Por lo tanto, los naturalistas europeos no estaban expuestos a la amplia variedad de especies y la diversidad de formas de vida que se encuentran en otras partes del mundo, lo que podría haberlos llevado a pensar de manera diferente sobre cómo surgen y cambian las especies (Bowler, 1989).
10.1 Importancia de la biodiversidad para el pensamiento evolutivo
Por otro lado, la baja biodiversidad en Europa podría haber llevado a algunos naturalistas a creer que las especies eran estáticas e inmutables, y que no se podían producir cambios significativos en ellas. Sin embargo, tanto Darwin como Wallace tuvieron la oportunidad de viajar a regiones con mayor diversidad biológica, lo que les permitió observar la variabilidad de las especies y la competencia por los recursos naturales. Esto llevó a la formulación de la teoría de la evolución y la selección natural, que explicaba cómo las especies podían cambiar a lo largo del tiempo y adaptarse a su entorno a través de un proceso de selección natural (Bowler, 1989).
10.2 Darwin y el contacto con una región megadiversa
La biodiversidad de seres vivos en las islas británicas es baja, los historiadores naturales tendían a caminar horas y horas para ver algún insecto interesante, pero eso no es así en la selva húmeda tropical del Amazonas, cuando Darwin la visitó se quedó absorto ante tantos colores y formas extraños, siendo incapaz de prestar atención a un insecto exótico, pues este caminaba sobre una flor extraña. Al inicio de su viaje Charles Darwin modelaba la naturaleza en base a dos ideas:

Figura 10.1. El gliptodonte. Glyptodon (griego para «diente con surco o esculpido») es un género extinto de un gran mamífero acorazado perteneciente a la subfamilia Glyptodontinae, emparentado con los armadillos que vivió durante la época del Pleistoceno. Era aproximadamente del mismo tamaño y peso que un Volkswagen Escarabajo, aunque de forma más aplanada. Con su caparazón óseo redondeado y extremidades agazapadas, recuerda superficialmente a las tortugas, y a los dinosaurios anquilosaurios, como ejemplo de la evolución convergente de linajes distantes hacia formas similares.
👉 El fijismo que significa que las especies creadas por Dios no habían cambiado desde el inicio de la creación.
👉 La teleología que implicaba que cada adaptación había sido creada con un propósito por Dios para su supervivencia o para el disfrute de los seres humanos.
10.3 El problema de la extinción
Durante su viaje Darwin visitó las costas de Uruguay y Argentina, por dos razones, la primera era que tenía fuertes mareos en el mar y la segunda era para investigar la fauna local, pero lo que llamó su atención eran los fósiles. Muchos de ellos como el gliptodonte (género Glyptodon spp.) y el megaterio (género Megatherium spp.) parecían versiones gigantes de los armadillos y perezosos que habitaban esas tierras. El dato más relevante era el hecho de la extinción, evidentemente eran especies que fallaron para sobrevivir, ¿Cómo explicar la extinción desde el punto de vista de un diseño inteligente? (Charles, 2008; Vizcaíno, Fariña, & Fernicola, 2009).

Figura 10.2. Megaterio. El Megatherium fue un mamífero extinto del orden Pilosa. Es también conocido como el perezoso terrestre gigante y se halló desde Argentina hasta Texas, poseía unas impresionantes dimensiones, de hasta 6 metros en los adultos. Su sólido esqueleto y su fuerte pelvis, excepcionalmente ancha, indican que un animal de movimientos lentos, podía erguirse sobre las patas traseras, en esa posición era más alto que un T. rex y podía sentarse sobre los cuartos traseros mientras con las extremidades anteriores arrancaba hojas, pero su dieta no consistía sólo en eso, era omnívoro. De este animal, como del Nothrotheriops, también se han hallado esqueletos completos y restos momificados que aún conservan piel, carne y pelo adheridos a los huesos.
10.4 El problema de las especies hermanas
Darwin amaba el estilo de vida de los gauchos argentinos “vaqueros y pastores de las planicies” y cuando exploró con ellos se encontró con un fenómeno desconcertante, las especies hermanas. Las especies hermanas son dos especies muy similares entre sí y que viven muy cerca la una de la otra. En este caso eran dos especies de ñandú que actualmente se conocen como Rhea americana y Rhea pennata, la segunda especie fue la descrita por Darwin y también es conocido como ñandú petiso o ñandú de Darwin (Charles, 2008).

Figura 10.3. Especies complejas y Darwin. Los ñandúes y su obvia relación con los avestruces fueron uno de los primeros contactos que tuvo Darwin con el problema de las especies complejas-complejos de especies.
10.5 El problema de los órganos vestigiales o que han cambiado de función estándar
Darwin encontró un problema con la creación especial, un diseñador no tendría problemas para diseñar estructuras específicas para funciones específicas, sin embargo, las aves patagónicas podían emplear una estructura específica “el ala aviar que es un quiridio con tres dedos” como alas, remos y aletas de estabilización, en otras palabras, no había función específica. Darwin estaba contrariado con dos fuentes de información opuestas. El pato vapor Tachyeres pteneres es evidentemente un pato, pero es incapaz de volar, lo cual sugería que era una especie que había perdido dicha capacidad recientemente ya que aún tenía alas (Charles, 2008).
10.6 El problema fósil
Pero en los registros históricos humanos, aun desde las primeras civilizaciones, daban constancia de que los animales no habían cambiado. ¿Cómo conciliar estas dos fuentes de información? Los fósiles de ostras de aguas poco profundas encontrados en las cercanías y altitudes del Monte Aconcagua en Argentina en principio no eran problemáticas dado el mito del Diluvio Universal, el problema es que tantas ostras no podrían crecer en un mismo lugar en solo 40 días, al ser animales que no se mueven fácilmente, requieren tiempo para colonizar. Darwin tuvo que emplear las sugerentes hipótesis de Charles Lyell que sostenían que las montañas podían formarse desde el océano muy lentamente por fenómenos ordinarios como sismos, los volcanes, la erosión y la sedimentación. Darwin mismo recibió reporte de que la roca se había elevado 3 metros sobre el mar (Charles, 2008).

Figura 10.4. El pato vapor. Tachyeres pteneres, o pato vapor porque al nadar sobre el agua parecía un barco de vapor de la época fue uno de los más grandes problemas para un Darwin defensor del diseño inteligente, pues las alas y su forma de nado no eran eficientes.
Debido a lo que Charles Darwin se encontró en las montañas de los Andes y de las ideas de Charles Lyell tuvo que sondear la inquietante idea de que la edad del mundo era mucho mayor que unos 4000 años como había creído hasta ese momento.
10.7 ¿Los pinzones de Darwin?
La siguiente parada fue en las islas Galápagos en el Ecuador. Esta es quizá la parada más famosa y envuelta en leyendas, nadie se pone de acuerdo en que especie de aves fue la que capturó la mente de Darwin, algunos dicen que fueron los pinzones, los sinsontes, los ruiseñores o los picogordos, y la realidad es mucho más complicada ya que solo de pinzones hay 14 especies solo en las Galápagos. Todos son sin embargo aves cantoras pequeñas clasificadas en el suborden Passeri que significa “con forma de gorrión” (Charles, 2008).


Figura 10.5. Sinsontes o pinzones de Darwin. Hablando de especies complicadas, pues, si se parecen, pero a Darwin le interesaron los sinsontes (arriba), no los pinzones (abajo). De las cuatro especies del complejo de sinsontes Nesomimus spp., de las Galápagos, Darwin capturó tres. Posteriormente los pinzones ganaron más notoriedad ya que su complejo de 14 especies era más complicado y gradual.
La clave de la biodiversidad de las Galápagos era que cada islita tenía su propia colección de aves, sucedía lo mismo con pinzones, sinsontes, ruiseñores. De hecho, el mismo patrón se repetía con reptiles que no podían volar como las Pinzones.
10.8 Islas realmente aisladas
Durante su viaje por el pacífico, Darwin se encontró con un tipo de isla diferente del de las Galápagos. Las Galápagos estan cerca al continente y por ende, la biodiversidad puede fluir con cierto grado de aislamiento, vertebrados como reptiles y aves pueden viajar fácilmente, pero en el pacífico se encontró con islas verdaderamente aisladas, los atolones.
Las islas de atolones de coral se consideran aisladas biogeográficamente debido a la distancia geográfica y la falta de conexión con otras regiones más continentales. Estas islas se forman a partir de arrecifes de coral que crecen en aguas tropicales poco profundas, y a menudo se encuentran a cientos o miles de kilómetros de otras islas o continentes.
Aunque las islas de atolones de coral están rodeadas de agua, esto no significa necesariamente que estén conectadas biogeográficamente con otras regiones. Los organismos terrestres y marinos no pueden viajar a través del agua sin ayuda, y muchos no tienen la capacidad de hacerlo a tan grandes distancias. Además, las corrientes oceánicas y las condiciones climáticas pueden limitar la capacidad de los organismos para viajar de una isla a otra.
Estas limitaciones en la dispersión de los organismos son importantes porque la biogeografía se ocupa de cómo se distribuyen los organismos en el espacio y cómo evolucionan en diferentes lugares. Las islas de atolones de coral son un ejemplo de una región aislada biogeográficamente que ha dado lugar a la evolución de especies únicas y endémicas. Debido a la falta de intercambio genético con otras poblaciones, los organismos en las islas de atolones de coral a menudo evolucionan de manera diferente de los de otras regiones.
10.9 El problema de la biodiversidad en los atolones
A Darwin también le interesó la biogeografía de estas islas. En los corales alrededor la biodiversidad era inmensa, pero sobre las islas era pobre. La hipótesis más aceptada era que en lugares aislados Dios había generado centros de creación independientes, con su propia diversidad, la pregunta era ¿Por qué en islas cercanas al continente Dios había creado más diversidad como en las Galápagos, pero en islas aisladas del continente, la diversidad creada por dios era más pobre?
La respuesta de Darwin a la pregunta anterior fue ignorarla por completo y generar una explicación completamente naturalista.
Esto implicaba que las plantas y animales terrestres debieron haber llegado de otras regiones y solo los que podían cruzar las barreras geográficas podían hacerlo. El principal problema o la principal consecuencia de la idea anterior radicaba en que las especies de las islas eran un poco diferentes de las de los continentes, lo cual a su vez implicaba que las especies de las islas habían cambiado al aislarse, la palabra para describir el cambio a través del tiempo que empleó fue transmutación, pero actualmente la llamamos “Evolución” (Charles, 2008).
11. El viaje de Wallace
11. El viaje de Wallace
Wallace era una generación más joven de Darwin y no conocía la respuesta a la gran pregunta de la historia natural de la época ¿de dónde surgen las especies?
Aunque Darwin había encontrado la respuesta tuvo miedo de sus repercusiones y no publicó nada. Alfred Rusell Wallace se embarcaría en su propio viaje de exploración, esta vez a las islas de Malaysia que se encuentran entre el sudeste de Asia y Australia, allí se encontraría con una serie de paradojas sobre la biogeografía, es decir sobre el modo en que los seres vivos se distribuyen geográficamente (Rubin, 2013).

Figura 11.1. Wallace y las especies complejas. Al igual que con Darwin, Wallace solo pudo analizar una fracción de la diversidad, que sin embargo le hubiera dicho la misma historia, la de un complejo de especies que se desarrollan en aislamiento generando características únicas.
Otro complejo de especies
El complejo de especies que capturó la atención de Wallace fueron las mariposas alas de pájaro (género Ornithoptera spp.,). Un complejo de especies son un grupo de especies muy semejante que a primera vista parecen ser una sola, pero en realidad son especies hermanas estrechamente relacionadas evolutivamente.
Otra distribución biogeogrpafica
El patrón de distribución de las mariposas alas de pájaro es semejante al de los animales de las islas Galápagos, en aislamiento lo que parecería ser una misma especie se diversifica de manera independiente, por lo que se puede reconocer de que isla pertenece un determinado espécimen solo por su apariencia externa. Al ser un mismo patrón, Wallace concluyó lo mismo que Darwin sin saberlo, debió haber una sola especie de mariposas alas de pájaro que se aisló en diferentes especies cuando las islas se separaron generando especies hermanas, en resumen, las especies cambian (Rubin, 2013).

Figura 11.2. Sin especies complejas no hay teoría evolutiva Wallace es un ejemplo de por qué no pudo haber verdadero evolucionismo antes de la edad de la exploración, sin un conocimiento realista de la diversidad y distribución de los seres vivos, detalles como la ley de Wallace de sobre donde surgen las especies no serían evidentes.
La ley de Wallace
La ley de Wallace establece que las especies hermanas evolutivamente deben distribuirse biogeográficamente de manera muy cercana “no al azar”, aunque separadas por barreras reproductivas como ríos, océanos y montañas.
A demás establece que los cambios de cada población aislada si son al azar, de lo contrario animales como el jaguar y el tigre que son similares entre si y viven en ecosistemas semejantes serian exactamente iguales, lo mismo sucedería con cacatúas y las guacamayas, si los cambios no fueran al azar ambos linajes aislados deberían verse exactamente igual.
La consecuencia de la ley de Wallace era que todas las especies debían estar emparentadas y que esto podía representarse por medio de un árbol de ramificaciones, en la que cada rama representa un linaje, del cual pueden surgir otros o extinguirse.
Una consecuencia era la existencia de organismos transicionales, por ejemplo, entre ballenas completamente adaptadas al mar y animales terrestres, sin embargo, aún hoy viven animales con diversos grados de adaptación gradual al mar como las nutrias, las focas, los leones marinos y los manatís. Wallace también se encontró una frontera invisible en las islas malayas, en el oeste vivían primates euterios en los árboles y en el este, canguros arborícolas marsupiales, esta frontera fue llamada la línea de Wallace (Rubin, 2013).
La catástrofe Mathusiana
Wallace sabía que los individuos de una misma especie son todos diferentes, y esto plantea diferencias en sus interacciones ecológicas, algunos pueden defenderse mejor de los depredadores, otros son mejores competidores y otros resisten mejor a los parásitos. Aquellos que con su variación especial pueden enfrentar de mejor forma a la suma de las interacciones ecológicas sobrevivirá y pasará su especial combinación exitosa a la siguiente generación. La suma de las interacciones ecológicas se conoce como la Selección Natural.

Figura 11.3. La catástrofe de Malthus. La selección natural surge como un modelo que relaciona a las poblaciones reproductivas y variantes con el equilibrio natural y la crueldad de las interacciones ecológicas. Los seres vivos luchan por la existencia, muchos mueren para mantener el equilibrio, los que sobreviven lo hacen porque tienen cuerpos superiores a los de sus compañeros en cada generación.
Debido a que las especies son super-fecundas y a la “ley de Malthus”, una combinación de caracteres exitosa se multiplicará en la población rápidamente con algunas variaciones nuevas en cada generación lo que reinicia el ciclo de selección (Rubin, 2013).
12. El nacimiento de una teoría
12. El nacimiento de una teoría
Después del regreso de Darwin a Inglaterra en 1836, comenzó a trabajar en sus notas y diarios de viaje, recopilando información y datos para apoyar su teoría de la evolución. Sin embargo, Darwin no completó su obra maestra "El origen de las especies" hasta 1859, más de 20 años después de su regreso.
Durante este tiempo, Darwin se dedicó a una variedad de investigaciones y proyectos, incluyendo estudios detallados de especies de plantas y animales, correspondencia con otros científicos y la realización de experimentos para investigar cuestiones relacionadas con la evolución y la selección natural.
A pesar de que "El origen de las especies" no fue publicado hasta 1859, Darwin había estado trabajando en la teoría de la evolución durante muchos años antes de eso. De hecho, la idea de la selección natural como mecanismo de evolución se le ocurrió a Darwin en la década de 1830, durante su viaje en el HMS Beagle. Fue solo después de muchos años de observación, recopilación de datos y reflexión que Darwin pudo desarrollar su teoría de manera completa y convincente.
En resumen, pasaron más de 20 años desde el regreso de Darwin en 1836 hasta la publicación de su libro "El origen de las especies" en 1859, aunque Darwin había estado trabajando en la teoría de la evolución y la selección natural durante gran parte de ese tiempo (Browne, 1995).
12.1 De regreso en el imperio
En cuanto Darwin analizó la biodiversidad que vio en su viaje en base a la idea de linajes de seres vivos que, al aislarse en diferentes territorios por medio de océanos o ríos, pudo plantear una relación o explicación común, el mismo fenómeno podía aplicarse a los ñandús, a las aves de las Galápagos, a las especies que había aisladamente en las montañas, pero al considerar que lo mismo sucedía en los seres humanos se dio cuenta de que le traería más de un problema.
Las claves de la nueva hipótesis de Darwin recaían en
👉 una gran cantidad de tiempo,
👉 la capacidad interna de las especies para cambiar con el tiempo, y
👉 algún mecanismo naturalista que causara que algunas se extinguieran mientras que otras fueran exitosas por algún tiempo.
El único esquema o gráfica que Darwin planteó en sus escritos originales fue un diagrama de árbol ramificado “en la actualidad lo llamamos dendrograma”, en el que las especies se diversifican generando nuevas especies.
12.2 Importancia de los complejos de especies o especies complejas
Los complejos de especies, también conocidos como especies análogas o especies crípticas, fueron importantes para el nacimiento de la teoría de la evolución moderna propuesta por Darwin y Wallace. Estos complejos consisten en grupos de especies que son muy similares morfológicamente, pero que en realidad son diferentes especies que han evolucionado a partir de antepasados comunes similares.
Antes de la teoría de la evolución, los naturalistas creían que las especies eran fijas e inmutables, lo que significa que no cambiaban con el tiempo. Los complejos de especies presentaban un desafío a esta idea, ya que mostraban que las especies podían ser muy similares, pero aun así diferentes. Esto llevó a Darwin y Wallace a cuestionar la idea de que las especies eran inmutables y a proponer la idea de que las especies podían cambiar y evolucionar con el tiempo.
Además, los complejos de especies fueron importantes para la teoría de la selección natural de Darwin y Wallace. Los complejos de especies que habían evolucionado en diferentes ambientes pero que se parecían morfológicamente, podían ser explicados por la selección natural. La selección natural puede producir características similares en diferentes especies que viven en ambientes similares, lo que lleva a la evolución de especies análogas o crípticas.
En resumen, los complejos de especies fueron importantes para el nacimiento de la teoría de la evolución moderna propuesta por Darwin y Wallace porque cuestionaron la idea de que las especies eran fijas e inmutables y proporcionaron evidencia de que las especies podían evolucionar con el tiempo. Además, los complejos de especies ayudaron a explicar cómo la selección natural podría producir especies análogas o crípticas en diferentes ambientes (Mayr, 2013).
12.3 La selección natural
El mecanismo evolutivo propuesto por Darwin se basa en un fenómeno intrínseco a las especies y otros que son externos a ellas. El fenómeno interno es que toda especie tiene la capacidad de reproducirse más rápido que sus recursos como el alimento y el resguardo. Los fenómenos externos a las especies se conocen en la actualidad como interacciones ecológicas: relación depredador-presa, competencia, comensalismo, mutualismo, parasitismo entre otras. La suma de las interacciones como la depredación y el parasitismo controlan las poblaciones y las especies, seleccionando naturalmente aquellos individuos más adaptados a un ambiente cambiante. A esta idea se la conoce como la Selección Natural (Charles, 2008).
Figura 12.1. YouTube: El origen de las especies: construyendo una teoría | HHMI BioInteractive Video.
12.4 Dos honorables caballeros del imperio
En ciencia quien publica primero se lleva el prestigio, sin embargo, a defenecía de otros casos como el de Carl Correns y Hugo de Vries o el de Isaac Newton y Gottfried Leibniz que se enfrascaron en amargas disputas de prioridades; Darwin y Wallace decidieron publicar conjuntamente en 1858, aunque Wallace siempre reconoció que Darwin tenía más datos y llevaba más tiempo estudiando esta nueva teoría. Darwin publicaría los frutos de 20 años de esfuerzos en El origen de las especies (1859), mientras que Wallace continuaría robusteciéndola con los años, publicando un resumen de su obra en un libro dedicado al viejo maestro que llamó Darwinismo (1889), el mismo año en que la genética clásica nace realmente de la mano de Hugo de Vries y Carl Correns (Rubin, 2013).
12.5 El origen de las especies
A diferencia de Wallace, Darwin soportó sus afirmaciones con un gran cuerpo de hechos llevados a cabo más que todo mediante experimentos de crianza selectiva y registro fósil. También proveyó observaciones directas de organismos que existían en sus ambientes naturales. Este reporte fue hecho principalmente en la obra magna de Charles Darwin, publicada un año después del artículo original sobre la selección natural ante la comunidad científica. Junto con los Principia de Newton, esta obra marca el inicio de una revolución científica que no deja de causar polémicas aun hasta nuestros días, su nombre es: El Origen de la Especies, Mediante la Selección Natural. El contenido del texto pude dividirse en las siguientes secciones:(Kutschera & Niklas, 2004).

Figura 12.2. El Origen de las Especies. —título original en inglés: On the Origin of Species— es un libro de Charles Darwin publicado el 24 de noviembre de 1859, considerado uno de los trabajos precursores de la literatura científica, el fundamento de la teoría de la biología evolutiva, y la fundación de la biología como ciencia no teológica.
👉 La descendencia común: La idea de que todos los seres vivos están emparentados entre sí.
👉 El origen de las variaciones: el aspecto más débil del texto, ya que se emplean hipótesis como la lamarkiana para explicar la herencia de los caracteres y su cambio entre generaciones.
👉 La aparición de variedades mediante la variación y la selección natural.
👉 El concepto de selección natural.
👉 El cambio gradual de una variedad en otras en base a datos biogeográficos y de aclimatación de variedades a diversas condiciones.
Darwin critica en concepto morfológico de especie, debido a múltiples casos en los que individuos con morfologías “polimórficos” diferentes se pueden reproducir entre sí; los ciclos de vida; los dimorfismos sexuales entre otros. Sin embargo, de forma similar a la variación, no se da un concepto claro.

Figura 12.3. Controversia social. Las alas conservadoras de la sociedad victoriana apreciaban la competencia, pero no pudieron aceptar el hecho de que el ser humano estuviera atado al mismo orden que los demás seres vivos, por lo que las ideas de Darwin fueron mal recibidas en esta área de la población imperial.
12.6 Características de la primera teoría de la evolución
Treinta años después, el codescubridor de la selección natural publicó una serie de lecturas bajo el título de Darwinismo (Wallace 1889), que trataba de los mismos tópicos que Darwin pero a la luz de hechos y datos que eran desconocidos por Darwin que había muerto en 1882. Un análisis comparativo entre Darwin y Wallace revela que las contribuciones de Wallace fueron más significativas de lo que usualmente es reconocido. Por lo anterior se ha propuesto que la selección natural sea reconocida con el principio de Darwin-Wallace. Aunque Darwin es usualmente acreditado como el autor principal de la teoría evolutiva, Erns Mayr señala que no es correcto referirse a “la teoría Darwiniana de descendencia con modificación” (la palabra evolución no aparece en el artículo original de Darwin y Wallace, solo en posteriores ediciones de El Origen de las Especies y en el libro de Wallace, Darwinismo) Si igualamos la palabra Darwinismo con el contenido del libro El Origen de las Especies, podemos distinguir entre cinco conceptos separados (Kutschera & Niklas, 2004):
👉 La evolución en sí. Las especies no son entidades fijas, son un conjunto de poblaciones con una estructura interna diferenciada. Los individuos son diferentes entre sí, y esta variación es heredable. Esto hace que cada generación sea un poco diferente de la anterior.
👉 Todos los seres vivos descienden de uno o unos pocos ancestros comunes.
👉 Para que una especie se convierta en dos deben darse unas condiciones específicas como el aislamiento reproductivo y el tiempo necesario para que sutiles diferencias de tipo continuo se acumulen.
👉 El proceso de acumulación de cambios continuos más selección natural conlleva a la formación de nuevas especies.
👉 La selección natural emerge de la lucha por la existencia de una especie que se encuentra al borde de la catástrofe malthusiana, solo aquellos capaces de vencer en esa lucha pueden pasar sus rasgos a la siguiente generación.
12.7 El problema de la primera teoría de la evolución
El problema fundamental con la primera teoría de la evolución es que en realidad no explicaba cómo se origina una especie, además de que la base de este problema era la carencia de una teoría de la herencia que permitiera explicar cómo operaba la selección natural al interior del material hereditario. Y no es que la explicación estuviera cerca, aunque el artículo de Mendel de 1866 estuvo enmarcado en el mismo periodo de tiempo que los trabajos de Darwin y Wallace (1858-1859), estos nunca entraron en contacto. Adicionalmente Mendel tuvo muy poco aporte, ya que la teoría de la genética clásica tal como la entendemos fue formulada entre los años de 1890-1910. Sin una teoría de la herencia sólida, la teoría de la evolución de Darwin siempre estuvo coja, e incluso durante los años de 1900-1910 gran parte de la comunidad científica expresaba serias dudas sobre su utilidad, denominándose a ese periodo como el ocaso del darwinismo (Kutschera & Niklas, 2004).
13. La selección natural darwiniana
13. La selección natural darwiniana
Existen dos formas de entender a la selección natural “SN”, la primera es a través de la dinámica de poblaciones y la segunda es por medio de las interacciones ecológicas. Pero, sin la explicación de la SN se desconecta de estos dos fundamentos puede conllevar a un vacío conceptual. Por ejemplo, podemos tomar los cuestionamientos hacia la selección natural planteados por el profesor Emilio cervantes:
“Confuso por la avalancha de críticas e insultos recibidos por formular la pregunta ¿Qué es la selección natural? he buscado en las fuentes más recientes, concretamente en el diccionario de la Neolengua para informarme. Ahí encuentro lo siguiente: La selección natural es un mecanismo evolutivo que se define como la reproducción diferencial de los genotipos en el seno de una población biológica. Cuando uno explica selección natural desde sus múltiples definiciones se convierte en un concepto extremadamente abstracto que raya en el razonamiento circular, pues como bien dice Carl Popper, no hay mucha diferencia en decir “sobrevive el más apto” a decir “sobrevive el que sobrevive””.
13.1 La población
Evidentemente el problema aquí es que la definición de selección natural dada sin ton ni son, es una respuesta a una pregunta que no se ha hecho, y sin la pregunta nada tiene sentido.
👉 Superfecundidad
Como he mencionado al inicio, la selección natural se entiende en su sentido ecológico: dinámica de poblaciones e interacciones ecológicas; ambas presuponiendo el principio de superfecundidad que es ejemplificado crudamente por las ostras.
👉 Una pareja de ostras tiene la capacidad de producir más de 114´000.000 huevos en tan solo una puesta, lo que implicaría que, si todos sobrevivieran, en tan solo 5 generaciones, se generarían más individuos que electrones en el universo. Del mismo modo, de esos 114´000.000/ciclo solo se requieren entre 2 y 4 sobrevivientes “en todos los ciclos” para mantener la estabilidad de la población (Sadava, Berenbaum, & Hillis, 2014).
👉 En los sistemas ecológicos, las poblaciones mantienen una estabilidad dinámica, lo cual implica que entre 2 y 4 huevos logran sobrevivir y el resto (113 999 998) se mueren.
👉 Azar o determinismo
Este dato nos permite hacer la pregunta, ¿Cuál es la razón de que sobrevivan los que sobreviven? Puedes tener dos respuestas. Si la supervivencia es indiferencial, entonces estamos en un azar absoluto al que se le llama deriva. Pero si la supervivencia es diferencial o dependiente de rasgos anatómicos de los seres vivos, entonces el azar disminuye y nos encontramos ante una selección. La selección natural no es un argumento circular en cuanto no es una afirmación pueril de sobreviven los que sobreviven, pues tal afirmación no distingue efectivamente entre la deriva “azarosa” de un proceso selectivo “antiazaroso”. La selección natural busca responder la razón por la cual sobreviven los pocos que sobreviven.
👉¿Qué es un mecanismo?
La pregunta debe trasladarse a que es un mecanismo en sí. Dependiendo del campo de estudio un mecanismo significa cosas completamente diferentes: En la ingeniería un mecanismo es un cuerpo rígido de elementos organizados para transmitir un movimiento mecánico. En química puede referirse a una secuencia de reacciones, o a los pasos que modelan una sola reacción química mediante estados de transición de muy corta duración. En biología un mecanismo es un modelo que explica como un determinado rasgo es generado. Nuevamente, si nos acogemos solo a las definiciones textuales de selección natural nada es posible de explicar.
👉 Interacciones ecológicas
Retomando nuestros dos puntos de análisis, en este caso son las interacciones ecológicas más que la dinámica de poblaciones lo que nos será útil. Una especie se relaciona con su medioambiente por medio de su cuerpo. Estas interacciones son la depredación, la simbiosis, la competencia, la obtención de recursos abióticos y la tolerancia a los factores abióticos. Cada una de esas relaciones puede verse como un mecanismo pues operan físicamente sobre las unidades estructurales que son las poblaciones, eliminando a la mayoría de los individuos y dejando solo a los que puede mantenerlas el tiempo suficiente para reproducirse. Sin embargo, vale la pena aclarar que no es un mecanismo constante, dado que la selección natural emerge de relaciones con entidades bióticas y abióticas, y ambas son de naturaleza variable y diversa.
13.2¿Qué compone a la selección natural?
Si la selección natural es un mecanismo, entonces: ¿De qué mecanismo se trata? ¿Qué elementos intervienen, qué estructuras y cómo están ordenadas? Se trata de un mecanismo biológico que opera en los niveles de la población, la comunidad y el ecosistema. Los elementos que intervienen son:
👉 Las frecuencias de los alelos en los individuos de la población
👉 Los individuos de la misma población
👉 Las especies con las que se puede interactuar en un lugar geográfico
👉 Los factores abióticos a aprovechar o tolerar.
👉 Propiedades emergentes
La organización es de una jerarquía anidada y evidentemente de un sistema auto-organizado con propiedades emergentes.
Las propiedades emergentes son características o comportamientos que aparecen en un sistema complejo que no pueden ser explicadas únicamente por la comprensión de las partes individuales que lo componen. En otras palabras, cuando se combinan o interactúan las partes individuales en un sistema complejo, surgen nuevas propiedades o comportamientos que no existen en las partes individuales. Estas propiedades emergentes no pueden ser reducidas a sus componentes individuales, sino que surgen de las interacciones y relaciones entre ellos. Las propiedades emergentes son comunes en la biología, la física, la química y en otros campos de la ciencia, y son importantes para entender cómo funcionan los sistemas complejos en la naturaleza y en la sociedad.
👉 Ciertos niveles de organización no manifiestan algunas propiedades emergentes
👉 Un solo individuo aislado de todo no puede sufrir selección natural, tampoco puede evolucionar.
👉 La población tampoco experimenta selección a menos que se la analice a lo largo del tiempo reproductivo-evolutivo, es decir varias generaciones. En la población se da la relación de competencia por recursos, competencia por sexo y selección sexual.
👉 En la comunidad se da la depredación, la simbiosis y la competencia.
👉 En el ecosistema se trata de aprovechar y tolerar. Los individuos deben poder entablar relaciones individuales de cada uno de esos tipos, pero es la población en su conjunto quien cambia por la selección generada individualmente por cada relación.
Dado que cada especie puede generar más de una relación, el producto neto o selección natural generada por la suma de toda relación natural es algo muy complejo, matemáticamente caótico pero ordenado, solo que es difícil de describir.
13.3¿Puede tratarse acaso de un mecanismo sin ninguna estructura, elementos, ni orden?
No realmente, es un mecanismo que depende de la estructura de un sistema ecológico, con elementos que se auto-organizan en un sistema caótico cuyo orden no es predecible. Debe recordarse que cuando se tiene un sistema no lineal de elementos que interactúan se generan propiedades de auto-organización complejas no predecibles. Aun sistemas relativamente simples como tres planetas generan soluciones organizadas difíciles de predecir. Este fenómeno de orden no previsible se lo denomina teoría de Caos y fue propuesto por Poincaré en 1890. El hecho de que no sea predecible hace que generalmente se lo denomine aleatorio o azaroso, sin embargo, este azar no implica carencia de orden, es solo un orden impredecible.
13.4¿Podrá, en ese caso ser el mismo mecanismo responsable de la reproducción diferencial de todos los genotipos en el seno de todas y cada una de las poblaciones?
Sí y no. La selección natural puede entenderse como el producto neto de todas las presiones generadas por cada una de las interacciones en los niveles de organización de la población, la comunidad y el ecosistema. Dado que los componentes de una población, una comunidad y un ecosistema cambian de un lugar a otro, el producto neto cambiará de un lugar a otro. Como producto neto si es el mismo mecanismo, ya que independientemente del tipo de relaciones, siempre existirán relaciones que generan presiones de selección Sin embargo debido a que la naturaleza de cada sistema ecológico cambia, la selección natural de cada lugar será un poco diferente.
13.5¿El mismo mecanismo de selección natural regirá cambios en poblaciones y la formación de especies?
En lo referente a cambios morfológicos evidentemente si, ya que una especie se define como el conjunto de poblaciones que la forman, si la población cambia entonces la especie cambia. Sin embargo, vale la pena distinguir entre el cambio de la especie a través del tiempo llamado anagénesis, de los procesos que conducen a la formación de nuevas especies llamado cladogénesis. La cladogénesis es un proceso complejo y se tiene evidencia de que está regido más por la deriva que por la selección “lo más común es que ambas estén mezcladas”, por lo menos en los mecanismos que generan la incompatibilidad reproductiva a nivel celular.
13.6¿El mismo mecanismo en la naturaleza y en el laboratorio?
En el laboratorio la mayoría de variables del ecosistema se simplifican, lo que hace muy fácil ver la influencia de una determinada relación en la adaptación de la población, por ejemplo, en las relaciones depredador-presa. En la naturaleza opera el mismo mecanismo, pero mucho más complejo debido a una mayor cantidad de integrantes y a una mayor cantidad de relaciones ecológicas. Adicionalmente, los cambios evolutivos en el laboratorio solo se observan generalmente en modelos biológicos que (1) se reproducen mucho y (2) tienen ciclos de vida muy cortos como las bacterias.
13.7¿Es serio llamar a la selección natural mecanismo?
Si en el sentido de que la selección natural es una propiedad de un sistema ecológico con múltiples componentes que se relacionan entre sí de formas específicas. El problema es que es un sistema complejo, por lo que los órdenes que genera no son predecibles por nuestra tecnología moderna. Las relaciones ecológicas a su vez no tienen nada de abstracto, las relaciones depredador presa, simbiosis, competencia dependen de contactos físico-químico-mecánicos para operar, como aplastar, atravesar, desgarrar entre otros. Es por esto que se dice que: nada tiene sentido en la evolución sino se lo mira desde la perspectiva ecológica.
14. Una nueva teoría, el neodarwinismo
14. Una nueva teoría, el neodarwinismo
En base a lo propuesto por autores como Kutschera U, Niklas K (2004) he decidido separar a la Síntesis Evolutiva Moderna de lo que se ha denominado new-darwinismo. En base a esto, el neodarwinismo iría desde la refutación de Weismann de la herencia blanda en 1883 y por lo tanto dándole una estoca fatal a la teoría de la herencia tomada por Darwin en su libro “El Origen de las Especies”, hasta la publicación en 1916 por parte de T. H. Morgan de su libro “crítica al Darwinismo” obra seminal que daría inicio a una nueva era en la biología. En este marco de tiempo de unos 33 años las teorías de Darwin no se instituyeron rápida o fácilmente como la teoría dominante que describe la variación de los seres vivos como uno pensaría en base a los libros de texto. En esta época se pueden resaltar varios aspectos:
👉 El rechazo de la herencia blanda o lamarckismo como teoría válida de la herencia.
👉 El ocaso del darwinismo, con un creciente número de dudas sobre la efectividad de la selección natural por autores tan prominentes como William Bateson.
👉 El rechazo de la variación continua darwiniana debido a la naturaleza discreta de los genes.
👉 Propuesta de la teoría mutacionista por Hugo de Vries.
Este estadio del desarrollo de la teoría evolutiva puede ser rastreado hasta el zoólogo y citólogo A. Weismann "1834-1914" que proveyó evidencia experimental en contra el principio lamarckiano de herencia blanda y además propuso que la reproducción sexual (recombinación en la segunda fase de la meiosis) genera en cada generación una nueva y variable población de individuos. La selección natural actúa sobre esta variación y determina el curso del cambio evolutivo. Desde que la teoría expandida de Darwin (Neo-darwinismo), enriqueció el concepto original de Darwin llevando la atención en como la variación biológica es generada y excluyendo el principio de Lamarck de herencia blanda como un mecanismo plausible para la evolución. Wallace en 1889 fue quien popularizó el termino (Darwinismo) e incorporó completamente los aportes de Weismann y fue desde entonces uno de los primeros proponentes del Neo-darwinismo.
14.1 August Weismann
August Friedrich Leopold Weismann (17 de enero de 1834 - 5 de noviembre de 1914) fue un biólogo evolutivo alemán. Ernst Mayr lo clasificó como el segundo teórico evolutivo más notable del siglo XIX, después de Charles Darwin. Weismann se convirtió en el Director del Instituto Zoológico y el primer Profesor de Zoología en Freiburg. Su principal contribución fue la teoría del plasma germinal, también conocida como Weismannismo, según la cual la herencia (en un organismo multicelular) solo tiene lugar por medio de las células germinales, los gametos, como los óvulos y los espermatozoides. Otras células del cuerpo -las células somáticas- no funcionan como agentes de la herencia. El efecto es unidireccional: las células germinales producen células somáticas y no se ven afectadas por nada que las células somáticas aprendan o, por lo tanto, ninguna habilidad que un individuo adquiera durante su vida. La información genética no puede pasar de soma a germoplasma y a la próxima generación. Los biólogos se refieren a este concepto como la barrera de Weismann. Esta idea, si es cierta, descarta la herencia de las características adquiridas según lo propuesto por Jean-Baptiste Lamarck (Churchill, 2015; Harman, 2009; Laubichler & Rheinberger, 2006).

Figura 14.1. August Weismann. (1834, Fráncfort del Meno - 1914, Friburgo de Brisgovia) fue un biólogo alemán. Se formó en la Universidad de Gotinga y enseñó zoología en Friburgo (1866-1912). Ernst Mayr lo situó como el segundo más notable teórico evolucionista del s. XIX, detrás de Charles Darwin.
La hipótesis de los caracteres adquiridos avanzó desde Jean-Baptiste de Lamarck (1744–1829) y fue aceptada por Darwin (1859, 1872) fue vigorosamente refutada por August Weismann (1834–1914). Durante la época en que Weisman y otros realizaron los experimentos que desacreditaron la noción de herencia blanda, los genetistas mendelianos habían desarrollado una base firme para explicar la variación entre los individuos en una población tal y como lo requería la teoría de Darwin. Desde 1960 los genetistas moleculares demostraron que los cambios en la composición de pares de bases en el ADN se traducían en cambios en la estructura de las proteínas. Lo anterior demostró que la noción de Weismann de herencia dura era un hecho irrefutable.
La idea de la barrera Weismann es central para la síntesis moderna de mediados del siglo XX, aunque los académicos no la expresan hoy en los mismos términos. En opinión de Weismann, el proceso de mutación en gran medida aleatorio, que debe ocurrir en los gametos (o células madre que los producen) es la única fuente de cambio para que la selección natural funcione. Weismann se convirtió en uno de los primeros biólogos en negar totalmente el lamarckismo y en consecuencia al primer darwinismo de manera parcial en su explicación sobre la herencia. Las ideas de Weismann precedieron al redescubrimiento del trabajo de Gregor Mendel, y aunque Weismann fue cauteloso al aceptar el mendelismo, sus sucesores más jóvenes pronto hicieron la conexión. Weismann es muy admirado hoy. Ernst Mayr lo consideró el pensador evolutivo más importante entre Darwin y la síntesis evolutiva alrededor de 1930-1940, y "uno de los grandes biólogos de todos los tiempos".
14.2 El gran fallo de Darwin
Weismann comenzó al igual que muchos de los historiadores naturales/biólogos del siglo XIX como Charles Darwin sus investigaciones asumiendo una herencia continua que es afecta por las condiciones del ambiente, una herencia blanda afectada por factores externos. Hacia 1876 aceptaba la noción de que las especies se transmutaban en otras especies por la acción directa del ambiente, en otras palabras, en adición a la selección natural existían otros factores que generaban la diversidad sobre la cual la selección natural eliminaba variantes. De hecho, Weismann escribió lo siguiente “si cada variación es el resultado de una reacción de los organismos a las condiciones externas, como una desviación de la línea de desarrollo, se sigue que ninguna evolución podrá darse sin un cambio en el ambiente”. Aunque la teoría neutral y el equilibrio puntuado actualmente son rutas de pensamiento que permiten llegar a la misma conclusión actualmente es importante resaltar como la primera teoría de la evolución y los conceptos de Lamarck no son antagónicos en su contexto, más bien son una amalgama en desarrollo durante esta época. Weismann también emplea la metáfora de Lamarck sobre los órganos de usados y no usados.
El primer rechazo de Weismann al concepto de herencia blanda, o mejor dicho herencia de los caracteres adquiridos se dio en una ponencia de 1883 titulada “sobre la herencia (Über die Vererbung). El estilo de Weismann es eminentemente comparativo con respecto a ejemplos concretos, es decir busca una serie de ejemplos concretos sobre las cuales aplicar los modelos teóricos y poder determinar cual explica sin mayores hipótesis auxiliares los fenómenos observados. Un ejemplo que le llamó la atención fue la existencia de castas no reproductivas al interior de una serie de artrópodos como las hormigas o las termitas, las cuales no pueden explicarse con la teoría de los caracteres adquiridos. En lugar de ello plantea una teoría radical para su tiempo a la cual denominará la teoría del plasma germinal, la cual logra explicar la existencia de las castas de hormigas no reproductivas como una analogía de las células somáticas que mueren mientras que las células germinativas pasan a la siguiente generación (Kutschera & Niklas, 2004).
14.3 La teoría del plasma germinal
La teoría del plasma Germinal, propuesta por A. Weismann es la base sobre la cual actualmente distinguimos dos tipos diferentes de líneas células al interior de los seres vivos multicelulares. El cuerpo de los animales y las plantas que interactúa con el medio ambiente está compuesto por la línea somática, la cual consiste en células especializadas para funciones específicas. Protegido al interior de la línea somática se encuentra la línea germinal o plasma germinal, la cual son las células que se reproducen, las que pasan la información genética a la siguiente generación. Las mutaciones que afectan a la línea somática solo afectan al cuerpo del poseedor, pero no necesariamente son transmitidas al plasma germinal que se encuentra aislado del ambiente. Es por esto que se logra el rechazo de la herencia blanda, aquellos factores que alteran a la línea somática como el ejercicio o las lesiones no transfieren su efecto a la línea germinal, a lo cual se le denomina la barrera de Weismann. Otro aspecto importante en la teoría de Weismann era que las células somáticas a medida que experimentaban su especialización perdían su información genética, quedándose únicamente con lo que necesitaban parta su labor especializada, mientras que el plasma germinal mantenía la totalidad de la información genética (Kirkwood & Cremer, 1982; Winther, 2001).
Actualmente se sabe que la herencia no está completamente aislada del medio ambiente debido a la existencia de proteínas que transfieren información desde el medio a los genes, pero su efecto no es tan grande como para invalidar la teoría en general. A los factores que pueden pasar por la barrera de Weismann los llamamos factores epigenéticos. Por otra parte, la noción de Weismann de que las células del soma pierden información durante su especialización si fue completamente refutada con los experimentos de clonación por transferencia de núcleo somático como los hechos con la oveja Dolly. Estos experimentos demostraron que las células del soma no pierden su información genética. La teoría del plasma germinal fue desarrollada entre los años de 1865-1889, y el fundamento de esta propuesta fue hecho originalmente por Francis Galton pero en una versión más débil (Kutschera & Niklas, 2004).
14.4 La mitosis y la meiosis
El significado de la mitosis y la meiosis fue descrito por A. Weismann en 1890 quien trabajó con huevos de erizos de mar, en las cuales observó diferentes tipos de división celular,
👉 a una la denominó división ecuatorial (Äquatorialteilung = mitosis) y
👉 a la otra división reductora (Reduktionsteilungres = meiosis).
Weismann notó como en la meiosis la cantidad de cromosomas disminuía a la mitad, y por lo tanto indujo la consecuencia de que durante la fecundación dos células con la mitad de los cromosomas debían unirse para que el número de cromosomas de la especie se mantuviera constante. Esto obviamente ya lo sabían otros biólogos previos a Weismann. Lo que realmente importa es que Weismann le dio un papel evolutivo a los procesos que fueron el fundamento de su teoría del plasma germinal, en donde la meiosis es quien se encarga de producir las células especializadas de dicho plasma (Churchill, 1970).
Durante esta época los trabajos de Mendel habían caído en el olvido, por lo que la conexión entre la genética mendeliana y los mecanismos de reproducción celular solo se haría evidente varias décadas después. Debido a la teoría del plasma germinal, la noción de herencia blanda de la primera teoría de la evolución que Darwin había tenido que introducir debido a la falta de explicación sobre el origen de la variabilidad fue refutada, de este modo Weismann es uno de los proponentes de la segunda teoría de la evolución o Neo-Darwinismo en la cual la herencia es independiente a los factores externos, concepto al cual se hará referencia como herencia dura.
14.5 El experimento de la cola de los ratones
La teoría del plasma germinal pone final absoluta a la teoría de Lamarck de los caracteres adquiridos. Aunque los experimentos de Weismann pueden verse un poco tontos en nuestro contexto, ay que recordar que en su época la situación era diferente.

Figura 14.2. El experimento de la cola de los ratones. El célebre experimento de la cola de los ratones demostró que estados de caracteres grandes como apéndices no eran afectados por fuerzas externas, lo que refutó la noción de herencia blanda del lamarckismo y el primer darwinismo.
Con frecuencia había reportes a cerca de como al cortarle la cola a un gato sus descendientes nacían sin cola, por ejemplo. Aunque estos reportes carecían de una fundamentación o corroboración seria, si hacían que la cuestión se discutiera en base a premisas imprecisas. Para corroborar o refutar los reportes Weismann condujo una serie de experimentos en los que tomó a 68 ratones y durante 5 generaciones repetidamente les cortó la cola a todos. Y a pesar de los 901 descendientes producidos durante 5 generaciones siempre presentaron colas al momento de nacer. Los resultados eran consistentes con su teoría del plasma germinal, refutando de forma casi definitiva la propuesta de Lamarck (Niklas & Kutschera, 2014).
14.6 La hipótesis de Weismann sobre el sexo
El término “Hipótesis de Weismann” ahora se concentra en la proposición explicita de que la reproducción sexual funciona como un proveedor de variación para la selección natural (Little, Jones, Burt, & Perrett, 2007; Mayr, 1982). A pesar de la importancia adaptativa del sexo (y el como el sexo evolucionó) ha permanecido como un área de considerable debate debido al costo del 50% en la aptitud, lo que en teoría debería favorecer a la reproducción asexual (Niklas, 1997). Nótese que, durante la reproducción sexual, cada progenitor solo le hereda el 50% de su genoma a su descendencia. El resultado de la vacación genomita introducido de esta forma puede conllevar a la aparición de individuos mal adaptados. En contraste, la reproducción asexual, asegura que los nuevos individuos están adaptados como sus padres al ambiente, lo que conllevaría a que cada miembro de la población generase clones de sí mismo. Por lo cual la pregunta del millón es ¿Por qué es tan común entre los eucariontes, la reproducción sexual?
De acuerdo con Mayr (1982) el dilema de la perdida de la aptitud en un 50% debido a la reproducción sexual se puede resolver si consideramos el hecho de que el ambiente al que estaban adaptados los padres cambia a través del tiempo y puede que no sea el mismo que enfrentara la descendencia. Niklas (1997) señaló que la reproducción sexual y asexual no son mutuamente excluyentes, una combinación de ambas se puede observar en plantas, hongos y un sin fin de organismos eucariontes microscópicos. La descendencia variable producida por la reproducción sexual puede contener (y debido al número, contiene) algunos individuos mejor adaptados a nuevos ambientes, mientras que la reproducción asexual produce organismos que retienen los caracteres adaptativos para un ambiente estable.
14.7 Como usan los hongos al sexo
Muchos hongos poseen ambos tipos de reproducción Figura 11.3, cuando llegan a un ambiente rico en recursos ellos empiezan a generar una población clonal generada por reproducción asexual, cuando las condiciones del ambiente se hacen menos favorables, los hongos entran en un estado que induce la generación de gametos y estructuras secundarias que sirven para dispersar a largas distancias los cigotos producidos por la reproducción sexual.

Figura 14.3. Ciclo de vida de los zigomicetes. Los hongos tienen dos ciclos de vida, uno asexual para colonizar ambientes estables y ricos en nutrientes y uno sexual para colonizar nuevos ambientes con condiciones impredecibles y diversas.
Esta estrategia no es única de los hongos, y la reproducción sexual siempre emerge como un fenómeno de emergencia, cuando las condiciones del ambiente se ponen poco favorables. Hamilton et al (1990) propusieron que la variabilidad genética que la reproducción sexual provee ayuda a las plantas, animales, hongos entre otros a lidiar contra parásitos y enfermedades. En la carrera armamentista evolutiva, los organismos patogénicos pueden crecer y adaptarse rápidamente debido a su gran número y corto ciclo de vida, lo que les permite evadir las defensas del organismo huésped. Rice y Chippindale (2001) han mostrado evidencia de que la reproducción sexual es ventajosa debido a que acelera la evolución genotípica sin el riesgo de la mutación aleatoria que puede generar tanto alelos ventajosos como perjudiciales. Todas estas ideas y experimentos soportan el concepto original de Weismann (1892) de que la reproducción sexual produce una progenie variable lo que promueve la evolución adaptativa.
15. Otros conceptos neodarwinianos
15. Otros conceptos neodarwinianos
15.1 Huxley y el eslabón perdido

Figura 15.1. El eslabón perdido. En la presente Escala Natural platónica puede distinguirse el concepto de eslabón perdido.
Thomas Henry Huxley “1825-1895” fue uno de los más grandes defensores del Darwinismo, aunque al principio aceptó con lentitud las ideas de Darwin. También es el primero en emplear un famoso término en forma de a una predicción, el eslabón perdido en 1863 (Beer, 1992; Weiss, 2004). El concepto de eslabón hace referencia a una cadena, que no es otra cosa que la Cadena del Ser también conocida como la Escala Natural. En ese contexto un eslabón perdido haría referencia a un nivel de complejidad que debería estar lleno con un ser. Con el advenimiento de Darwin, el concepto de eslabón perdido hace referencia a seres que estuvieran cercanamente emparentados o que inclusive fuesen los ancestros comunes de las especies vivas muy diferentes “humanos-primates”, “aves-reptiles”. El eslabón perdido debía ser un fósil que hubiera vivido en un periodo de tiempo específico con características mezcladas entre primate y ser humano.
15.2 Los primeros fósiles de transiciones
Después de que esta idea de eslabón perdido fuera lanzada, los arqueólogos y paleontólogos se lanzaron a la búsqueda de tal criatura. La búsqueda se centró en Asia y África tal como lo habían hipotetizado Darwin en su libro “la descendencia del hombre” y Wallace en su libro “Darwinismo”. La búsqueda terminó cuando Eugene Dubois “1858-1940” descubrió en 1894 el primer espécimen de Homo herectus, aunque originalmente se lo llamó Pithecanthropus erectus (Dubois, 1894). Otros fósiles que cumplían el criterio de eslabón perdido fueron encontrados durante la misma época, como, por ejemplo, los primeros especímenes de Archaeopteryx spp., entre los años de 1860-1880 (Charig, 1986; Wellnhofer, 2010). Esta época puede describirse como el inicio fuerte de la paleontología, y una fiebre de fósiles se apoderó de los dueños de museos. En cualquier caso, una vez descubierto un eslabón perdido ya no está perdido, por lo que desde hace mucho tiempo se los denomina como fósiles transicionales, supongo que el nombre de eslabón encontrado no es tan cool.
15.3 Haeckel y los embriones
Ernst Haeckel “1834-1919” fue uno de los más grandes biólogos del siglo XIX y también uno de los más controversiales.

Figura 15.2. Ernst Haeckel Uno de los primeros evolucionistas alemanes, si figura se ha vuelto controversial con los años, aunque sigue siendo uno de los biólogos más importantes del siglo XIX.
De nacionalidad pruso-alemana fue quien llevó el pensamiento de Darwin a los germanoparlantes. A él se le deben términos como el de Filum y Ecología. En términos teóricos podemos definir que, Haeckel estaba más cercano a la primera teoría de la evolución, ya que aceptaba muchos de los mecanismos lamarkianos o de herencia blanda que en el neodarwinismo quedan proscritos experimentalmente (Amundson, 2005; Hall, 2003; Hoßfeld & Olsson, 2003; Manfred Dietrich Laubichler & Maienschein, 2007).

Figura 15.3. El feto no es lo que parece. Esto no es un ser humano, ni se le deben dar derechos civiles propios de un ciudadano, si no le gusta, pues de malas, porque es un elefante en estado embrionario. Aun cuando los dibujos de Haeckel tienen varias licencias artísticas las fotos de los embriones nos revelan la misma idea. En el nivel embrionario los animales son muy semejantes.

Figura 15.4. Embrión de delfín. Puede notarse que embrionariamente los delfines forman extremidades traseras, las cuales se retraen con el tiempo. Sin embargo, algunos individuos fallan en eliminarlas.
15.4 El darwinismo imperfecto
En cualquier caso, con Hackel podemos ver que las ideas de la escala natural aún seguían persistentemente incrustadas en el pensamiento de los biólogos posteriores a Darwin, y esto se puede ver claramente en su árbol filogenético-taxonómico en el cual en la copa está el hombre y en la base están los seres vivos unicelulares. Es decir, la tendencia de la Escala Natural de poner en la base lo que se considera menos perfecto y en la copa lo más perfecto, aunque en el camino se observen las ramificaciones que proponía Darwin.
El proceso aleatorio no fue tomado en cuenta por Haeckel quien pesaba que la evolución procedía por un camino de complejización inexorable. También vale la pena mencionar que el impulsó la integración de la embriología con la teoría evolutiva con su propia teoría de la recapitulación, en la cual se proponía que la ontogenia “desarrollo embrionario” resume la filogenia “desarrollo evolutivo”. Aunque tal ley, junto con varios detalles de sus dibujos están desacreditados hoy en día, algunos detalles de esta sin se han confirmado, por ejemplo, la presencia de rasgos arcaicos en el desarrollo embrionario.

Figura 15.5. Darwinismo imperfecto. El árbol filogenético de Ernst Haeckel aún se encuentra influido por las ideas de la escala natural, dado que el tronco sigue siendo lineal. Los primeros Darwinistas no entendieron completamente la falta de dirección en el proceso evolutivo, es decir, la falta de perfección de algún ser vivo sobre otro.
15.5 La crisis del neodarwinismo
Julian Huxley usó la frase "el eclipse del darwinismo" para describir el estado de las cosas antes de lo que llamó la síntesis moderna, cuando la evolución en sentido no-darwiniano fue ampliamente aceptada en círculos científicos, pero relativamente pocos biólogos creían que la selección natural fuera su mecanismo principal o que esta afectara a una herencia continua. Los historiadores de la ciencia como Peter J. Bowler han usado la misma frase como etiqueta para el período dentro de la historia del pensamiento evolutivo desde la década de 1880 hasta alrededor de 1920, cuando se desarrollaron y exploraron alternativas a la selección natural -como muchos biólogos consideraban la selección natural ha sido una suposición errónea por parte de Charles Darwin, o al menos por una importancia relativamente menor. Se ha propuesto un término alternativo, la interfase del darwinismo o crisis del neodarwinismo. Si bien hubo varias explicaciones de la evolución que incluyeron el vitalismo, el catastrofismo y el estructuralismo durante el siglo XIX, cuatro alternativas principales a la selección natural estaban en juego a comienzos del siglo XX (Bender, 2004; Bowler, 1992a, 2005; Mark A Largent, 2009):
👉 La evolución teísta fue la creencia de que Dios guió directamente la evolución.
👉 El neolamarckismo fue la idea de que la evolución fue impulsada por la herencia de características adquiridas durante la vida del organismo.
👉 La ortogénesis era la creencia de que los organismos se veían afectados por fuerzas internas o leyes de desarrollo que impulsaban la evolución en direcciones particulares.
👉 El mutacionismo fue la idea de que la evolución fue en gran medida el producto de mutaciones que crearon nuevas formas o especies en un solo paso.
La evolución teísta desapareció en gran parte de la literatura científica a fines del siglo XIX, cuando los llamamientos directos a las causas sobrenaturales llegaron a considerarse anticientíficas. Las otras alternativas tuvieron seguidores significativos hasta bien entrado el siglo XX, además de tener ciertos grados de amalgamamiento que las hace difíciles de distinguir en ciertos autores; la biología dominante los abandonó en gran medida solo cuando los avances en genética los hicieron parecer cada vez más insostenibles, y cuando el desarrollo de la genética de poblaciones y la síntesis moderna demostraron el poder explicativo de la selección natural. Ernst Mayr escribió que aún en 1930 la mayoría de los libros de texto aún enfatizaban tales mecanismos no-darwinianos.
El desarrollo de la teoría de herencia discreta o mendelismo fue la clave de este eclipse, pues llevó a muchos biólogos mendelianos a negar de plano la variación continua de los caracteres, por ejemplo, intentando encajonar la variación continua del color de la piel humana solo en tres o cuatro categorías para poder intentar modelarlas en un sistema mendeliano simple o saltasionista, sin intermedios. Estas ideas hicieron también su irrupción en las teorías evolutivas, ya que, si se negaba la variación continua, la evolución neodarwinista no tendría sustento.
15.6 El saltacionismo
Hay que tener en cuenta que este saltacionismo también es una teoría evolutiva en derecho propio e igual de naturalista, solo que no era gradual. Este saltacionismo podía tener tres vertientes:
👉 uno neo-larmarckiano en el que los saltos siempre perfeccionaban a su antecesor por una fuerza externa
👉 otro llamado ortogénesis en el que los saltos siempre perfeccionaban a su antecesor por una fuerza interna. Junto con el anterior generaba un patrón saltacionista o mendeliano hacia el perfeccionamiento de manera lineal, lo cual se modelaba con esquemas como “la marcha de los primates”;
👉 y otro de corte más mutacionista en el que el salto no era ni bueno ni malo, sino que dependía del ambiente.
El descubrimiento de Hugo de Vries 1848-1935 de una mutación de salto de especie en la planta de Onagra, junto con el éxito de los mendelianos hizo que durante los años de 1900-1915 el neodarwinismo estuviera casi al borde de ser eliminado de los escritos serios de los biólogos, y a este periodo de tiempo en general se lo conoce como la crisis del neodarwinismo, y el autor más prominente en esta época fue William Bateson 1861-1926 (Bowler, 2000, 2005; Nordmann, 1992).
Aunque el mendelismo es útil para casos concretos, los mendelianos se enceguecieron al tratar de encajonar caracteres continuos en cajones discretos, es por esto que en libros de texto es común encontrar falsos ejemplos de herencia mendeliana como en el caso de: el arco del pulgar, el pico de viuda “entradas de Vegeta”, control de la lengua, el color de los ojos, el color de la piel, los hoyuelos en las mejillas, el cabello rojo, o la sensibilidad a ciertos químicos. Estos caracteres realmente obedecen a una herencia continua, que es la base de una evolución neodarwiniana. Otro detalle que conllevó a ignorar a las malas a la herencia continua fue el hecho de querer encajonar a la población humana con herencia continua en cajones discretos mendelianos llamados razas. Solo a través del racismo se puede ver una variación continua como el color de la piel en cajones discretos.
15.7 La ortogénesis
La ortogénesis era la teoría de que la vida tiene una tendencia innata “internalista” al cambio, de una manera unilineal en una dirección particular. El término fue popularizado por Theodor Eimer, un zoólogo alemán, en su libro de 1898 sobre Orthogenesis: Y la impotencia de la selección natural en la formación de especies. Había estudiado la coloración de las mariposas y creía haber descubierto características no adaptativas que no podían explicarse por selección natural. Eimer también creía en la herencia lamarckiana de las características adquiridas, pero sentía que las leyes internas de crecimiento determinan qué características se adquirirían y guiaron la dirección de la evolución a largo plazo por ciertos caminos, en lugar de fuerzas lamarckianas de naturaleza externalista (Bowler, 1979).
La ortogénesis tuvo un seguimiento significativo en el siglo XIX, sus defensores incluyeron al biólogo ruso Leo S. Berg y al paleontólogo estadounidense Henry Fairfield Osborn (Levit & Olsson, 2006; Osborn, 1922). La ortogénesis fue particularmente popular entre algunos paleontólogos, que creían que el registro fósil mostraba patrones de cambio unidireccional gradual y constante. Los que aceptaron esta idea, sin embargo, no aceptaron necesariamente que el mecanismo que impulsaba la ortogénesis fuera teleológico (dirigido a un objetivo). Creían que las tendencias ortogénicas no eran adaptativas; de hecho, sintieron que en algunos casos condujeron a desarrollos que eran perjudiciales para el organismo, como las grandes cornamentas de los alces irlandeses que, según ellos, condujeron a la extinción del animal, por lo que esta teoría se amalgamaba parcialmente con un mutacionismo azaroso saltacionista.

Figura 15.6. La ortogénesis fue muy usada por los paleontólogos, de allí que se filtrara a la cultura popular como el modo en que se entiende la evolución de manera errónea.
16. El inicio de la síntesis moderna
16. El inicio de la síntesis moderna
La síntesis evolutiva extendida consiste en un conjunto de conceptos teóricos más completos que la síntesis moderna anterior de la biología evolutiva que tuvo lugar entre 1918 y 1942. La síntesis evolutiva extendida fue solicitada en la década de 1950 por CH Waddington, argumentada sobre la base de puntuado Equilibrio de Stephen Jay Gould y Niles Eldredge en la década de 1980, y fue reconceptualizado en 2007 por Massimo Pigliucci y Gerd B. Müller. La síntesis evolutiva extendida revisita la importancia relativa de los diferentes factores en juego, examinando varios supuestos de la síntesis anterior y ampliándola con factores causales adicionales. Incluye selección multinivel, herencia epigenética transgeneracional, construcción de nicho, potencial evolutivo y varios conceptos de evo-devo. No todos los biólogos están de acuerdo con la necesidad o el alcance de una síntesis extendida. Muchos han colaborado en otra síntesis en biología evolutiva del desarrollo, que se enfoca en la evolución de la genética molecular y la evolución para comprender cómo la selección natural opera en procesos de desarrollo y homologías profundas entre organismos a nivel de genes altamente conservados. En general no hay un cambio de paradigma radical en el sentido de que en general sigue siendo gradual en el fenotipo, y la selección natural junto con la deriva genética conservan su importancia, por lo que en este texto no nos referimos a la síntesis expandida como una teoría aparte sino más bien como la tercera etapa de la TSE que sigue y seguirá expandiéndose con el tiempo (D. J. Futuyma, 2015; Stoltzfus, 2017; Tanghe, De Tiège, Pauwels, Blancke, & Braeckman, 2018).
16.1 Repaso general de la historia de la teorìa
Al igual que con la teoría atómica, la teoría evolutiva posee modelos que se suceden históricamente, siendo los más relevantes:
👉 el darwinismo de 1838,
👉 el neodarwinismo de 1895 y
👉 la Teoría Sintética de la Evolución “TSE” que a su vez se divide en momentos:
👉 primera generación 1915-1937,
👉 segunda generación 1937-1947 y expansión continua hasta la actualidad.
16.2 Primera generación
La TSE posee su fundamento en el trabajo de T. H. Morgan en 1916 quien indicó la labor que debían hacer los biólogos futuros, una fusión entre matemáticas, genética mendeliana y neodarwinismo en un solo cuerpo teórico, de allí el nombre de “sintética”. Este trabajo fue realizado por sus sucesores, la primera generación de los sintéticos-evolutivos, R. Fisher, J. Haldane y S. Wright quienes crearon matemáticas nuevas para la biología. Hacia mediados de los años 20 los filósofos de la ciencia señalaron que para que una Ciencia de la Naturaleza sea declarada como tal, esta debe estar matematizada o axiomatizada, algunos decían que la biología no era ciencia por su falta de rigor matemático, los primeros intentos de resolver esto vinieron de la mano de los mendelianos, pero con limitaciones que fueron resueltas por Fisher, Haldane y Wright. Pero aún faltaba algo, una conexión entre la matemática evolutiva teórica con ramas empíricas de la biología como la ecología, la anatomía y especialmente la paleontología (Kutschera & Niklas, 2004).
16.3 Thomas Hunt Morgan
Aunque inicialmente escéptico ante conceptos tanto del mendelismo como del neodarwinismo, Thomas Hunt Morgan 1866-1945 logra vincular la herencia continua a través de fenómenos mendelianos a través de los caracteres poligenéticos.
👉 Caracteres poligénicos
Si un solo carácter como el color de la piel estuviera vinculado por las de un gen, y los heterocigotos manifestaran rasgos de dominancia intermedia no es difícil racionalizar una expansión simple.

Figura 16.1. Thomas Hunt Morgan (25 de septiembre 1866 - 4 de diciembre 1945) fue un genetista estadounidense. Estudió la historia natural, zoología, y macromutación en la mosca de la fruta Drosophila melanogaster. Sus contribuciones científicas más importantes fueron en el campo de la Genética. Fue galardonado con el Premio Nobel de Fisiología y Medicina en 1933 por la demostración de que los cromosomas son portadores de los genes, lo que se conoce como la teoría cromosómica de Sutton y Boveri. Gracias a su trabajo, Drosophila melanogaster se convirtió en uno de los principales organismos modelo en Genética.
👉 con un solo gen y dos alelos los fenotipos son 3
👉 con dos genes los fenotipos son 9
👉 con tres genes los fenotipos serían 27
👉 con cuatro genes los fenotipos serían 81.
Tenga en cuenta que en esta expansión solo hemos tenido en cuenta dos alelos en cada uno de los 4 genes, pero en la realidad cada gen puede tener entre 20 y 200 lelos, por lo que la cantidad de fenotipos se eleva a una cantidad astronómica que da la apariencia de una continuidad, de hecho, aun asumiendo solo dos alelos por cada gen tenemos entre 27 y 81 tonos de piel diferentes, como cuando uno tiene una caja de colores finos, aunque cada lápiz de colores es discreto, como hay muchos y cada uno varia sutilmente, se genera la apariencia de continuidad de la que hablaban los neodarwinistas.
👉 El blanco de la selección natural
De esta forma la selección natural opera eliminando combinaciones de alelos específicas, mientras que la mutación y recombinación genera nuevas variantes al azar.

Figura 16.2. La paradoja de la gradualidad. Aunque filosóficamente hay quien encuentra extraño que desde lo discreto emerja lo continuo, una caja de colores finos nos demuestra que una gran cantidad de caracteres discontinuos que solo cambian un poco unos de otros generan la ilusión de continuidad de la que hablaban los darwinianos, y neodarwinianos.
De esta manera el mendelismo no era una teoría opuesta al neodarwinismo, sino más bien un caso especial, el caso especial más fácil y que podía modelarse matemáticamente más fácilmente. Morgan y sus alumnos dejaron sentadas las bases para fusionar a la evolución neodarwiniana y a la genética mendeliana en un solo cuerpo que sería llamado años más tarde la Síntesis Moderna o Teoría Sintética de la Evolución.
17. La primera generación de sintéticos evolutivos
17. La primera generación de sintéticos evolutivos
La historia de la síntesis evolutiva moderna se divide en dos generaciones porque hubo dos períodos importantes en los que se desarrollaron y consolidaron las ideas y teorías que conformaron esta síntesis.
👉 La primera generación de la síntesis evolutiva moderna se produjo en las décadas de 1920 y 1930, y estuvo liderada por biólogos como Ronald Fisher, J.B.S. Haldane y Sewall Wright. Durante este período, se establecieron los fundamentos teóricos de la evolución mediante la combinación de la genética mendeliana con la teoría de la evolución de Darwin. Estos biólogos desarrollaron la teoría de la selección natural y establecieron los principios de la genética de poblaciones.
👉 La segunda generación de la síntesis evolutiva moderna tuvo lugar en las décadas de 1940 y 1950, y se caracterizó por la inclusión de nuevas disciplinas como la paleontología, la ecología y la sistemática en la síntesis. Además, se comenzó a tener en cuenta la importancia de los procesos de especiación y la evolución molecular. Algunos de los líderes de esta segunda generación incluyen a Theodosius Dobzhansky, Ernst Mayr y George Gaylord Simpson.
En resumen, la división de la síntesis evolutiva moderna en dos generaciones se debe a que se produjeron importantes avances teóricos y metodológicos en dos períodos diferentes, liderados por diferentes científicos y con diferentes enfoques y disciplinas involucradas. Ambas generaciones fueron cruciales para la consolidación de la teoría de la evolución moderna.
17.1 Fisher
El primer autor es Sir Ronald Aylmer Fisher “1890-1962” quién trabajó específicamente en poblaciones de vegetales. El punto de su investigación puede distinguirse en dos líneas,
👉 Maestro estadístico
la primera hace referencia a la medida de la variación entre los diferentes rasgos de un carácter. Para esta fecha ya era obvio que los caracteres reales NO eran mendelianos, es decir, poseían una herencia continua de muchos rasgos “no dos, como en la simplificación mendeliana”. De hecho, la variación era enorme y difícil de distinguir porque los cambios son graduales, un factor muy común de los caracteres de herencia continua. Los métodos estadísticos de Fisher se desarrollaron para poder estudiar este tipo de variación compleja.

Figura 17.1. Sir Ronald Aylmer Fisher (Londres, Reino Unido, 17 de febrero de 1890 – Adelaida, Australia, 29 de julio de 1962) fue un estadístico y biólogo que usó la matemática para combinar las leyes de Mendel con la selección natural, de manera que ayudó así a crear una nueva síntesis del Darwinismo conocida como la síntesis evolutiva moderna, y también un prominente eugenista en la parte temprana de su vida.
A continuación, se presentan algunas de las técnicas estadísticas más importantes desarrolladas por Fisher:
👉 Análisis de varianza (ANOVA): Fisher desarrolló esta técnica para analizar la varianza en conjuntos de datos, lo que permitió identificar las fuentes de variabilidad en los experimentos. Esta técnica ha sido ampliamente utilizada para analizar datos en biología y otros campos.
👉 Distribución de Fisher: Fisher desarrolló la distribución de Fisher, que es una distribución de probabilidad utilizada para analizar datos en la estadística inferencial. La distribución de Fisher se utiliza comúnmente en pruebas de hipótesis y en análisis de varianza.
👉 Prueba exacta de Fisher: Fisher desarrolló una prueba exacta para evaluar la significancia de los datos de conteo. Esta prueba se utiliza comúnmente en la genética para analizar datos de segregación.
👉 Estimación de máxima verosimilitud: Fisher desarrolló el método de estimación de máxima verosimilitud, que es un método utilizado para estimar los parámetros de un modelo estadístico a partir de datos observados. Este método se utiliza ampliamente en la genética para estimar la frecuencia de alelos y la tasa de mutación.
En resumen, las técnicas estadísticas desarrolladas por Fisher han sido fundamentales para el estudio de la evolución y la variabilidad en la biología y otros campos. Su trabajo ha permitido a los científicos analizar y comprender mejor los datos, lo que ha llevado a importantes avances en la comprensión de la genética, la evolución y la biología en general.
👉 Diseño experimental
La segunda línea tenía que ver con el diseño experimental, donde aplicó lo aprendido en el estudio de la variabilidad “varianza” al diseño de experimentos. Este trabajo le convirtió en la máxima autoridad en el diseño experimental, y de hecho, su trabajo sobre métodos estadísticos para distinguir la varianza y la similitud, se emplean hoy en áreas como la economía, la física o la ingeniera, esto gracias al desarrollo de pruebas estadísticas como los tests exactos, la distribución z y la F de Fisher, para las tablas de análisis de varianza o ANOVAs (Charlesworth & Edwards, 2018; Gordon, 2015; Vyas & Desai, 2015).
El trabajo fundacional de Fisher sobre la Teoría de la Evolución fue un libro llamado “La teoría genética de la Selección Natural” de 1930, donde el autor expone varios principios que son ahora lineamientos básicos de la teoría moderna de la evolución. Entre otros aportes caben señalar la demostración de que las macromutaciones tienden a disminuir la aptitud de supervivencia de los individuos, mientras que las micromutaciones no tienen un impacto tan perjudicial en caso de ser deletéreas.
Evidentemente son tendencias estadísticas, por lo que pueden encontrarse excepciones. Los métodos estadísticos de conteo de alelos en una población para diferentes rasgos y su conexión con las teorías de la Ecología, esto gracias a la colaboración de Fisher con ecólogos influyentes como Edmund Brisco Ford “1901-1988”. Fisher también se le acredita por el concepto de Vigor Híbrido, en el cual los heterocigotos poseen una mayor tendencia a sobrevivir que las variedades puras.
17.2 Haldane

Figura 17.2. John Burdon Haldane ( Oxford, 5 de noviembre de 1892 - Bhubaneswar, India, 1 de diciembre de 1964) fue un genetista y biólogo evolutivo británico. Junto con Ronald Fisher y Sewall Wright, fue uno de los fundadores de la genética de poblaciones.
👉 Una persona muy diferente Lord Fisher
Para los estándares modernos John Burdon Haldane “1892-1964” sería considerado un científico realmente peculiar, un ateo con tendencias comunistas y a experimentar consigo mismo, lo que contrastaba con Fisher que era cristiano anglicano devoto, y aun así con creencias personales tan contrastantes ambos concordaron en la realidad evolutiva de estilo darwinista y la importancia de su estudio. Haldane comenzó a trabajar ya desde 1908 junto con su hermana en la genética mendeliana, su modelo biológico inicial fue el conejillo de indias (Ellison, 2018; Mahanti, 2015; Nicoglou, 2018).
👉 Experimentos de Haldane
Sin embargo, su primer trabajo técnico a cerca de genética postmendeliana fue publicado en 1915 empleando ratones como modelos biológicos, aunque se conoce que a lo largo de sus investigaciones empleó otros mamíferos como conejos o el mismo, así como aves de corral como los pollos. Su trabajo lo llevó a demostrar que el ligamiento de genes en un solo cromosoma también era posible en los mamíferos y las aves, esto ya lo había demostrado Morgan y su grupo de investigación entre 1910 y 1915 pero solo para la mosca de la fruta.
👉 Conceptos ampliados por Haldane
Sin embargo, lo que en realidad lo pone en esta sección es una serie de 10 artículos publicados entre 1924 y 1934 en los cuales desarrolla la Teoría Matemática de la Selección Natural y Artificial (Haldane, 1930, 1931, 1932). En estos, Haldane analizó:
👉 los efectos de la endogamia “1924”
👉 la mutación y su selección “1927”
👉 el aislamiento reproductivo “1930”
👉 la variación y la potencia de la selección “1931”
👉 la estabilidad de las poblaciones en términos de genética y selección natural “1932”
👉 los efectos de fuertes presiones de selección “1932” entre otros.
👉 Haciendo predicciones audaces y concretas
Junto con los trabajos de Fisher esta aproximación matemática de la biología constituye el núcleo fuerte de la Teoría Sintética de la Evolución. Aunque filósofos de la ciencia dudaron del fundamento epistemológico de la teoría de la evolución, como Imre Lakatos 1922-1974, es posible usar su propia filosofía para demostrar su error. Para Imre Lakatos un programa de investigación válido debe realizar predicciones audaces que luego son validadas por la experimentación. Haldane realizó una predicción muy concreta, el propuso que en poblaciones con fuertes índices de malaria “una enfermedad mortal”, las poblaciones tenderían a desarrollar anormalidades que, si bien son limitantes, las protegerían de la enfermedad. Esta propuesta se conoce como la hipótesis de la malaria y fue propuesta en 1949. Estas anormalidades fueron confirmadas por Anthony Clifford Allison en 1954 en forma de ciertas enfermedades como la malaria falciforme y algunas talasemias, que si bien causan limitaciones a los individuos que las padecen, también los hacen inmunes a una enfermedad mortal como es la malaria (Allison, 2002; Livingstone, 1983; Siniscalco et al., 1966; Weatherall, 1987; Yuthavong & Wilairat, 1993).
17.3 Wright

Figura 17.3. Sewall Green Wright (21 de diciembre de 1889 – 3 de marzo de 1988) fue un genetista estadounidense conocido por su influyente trabajo en teoría evolutiva. Sus artículos sobre endogamia, sistemas de apareamiento y deriva genética lo convirtieron en uno de los principales fundadores de la genética poblacional, junto con Ronald Fisher y J.B.S. Haldane.
Sewall Green Wright “1889-1988” fue el tercer fundador de la teoría sintética, y al igual que los otros dos, su campo fue la matematización de la biología por medio de la estadística moderna, de hecho, él trabajó con Fisher. Tal vez el aporte singular más importante de Wright a la teoría de la evolución es el concepto de la deriva genética “1929” (Dietrich & Skipper Jr, 2012; Orr, 2005, 2009; Provine, 1989).
👉 La deriva genética
La deriva genética es una fuerza evolutiva que emerge de fenómenos azarosos que son indiferentes de las capacidades de los individuos, es decir, a diferencia de la selección natural, donde algunos individuos de la población pueden afrontarla con mayor éxito, en la deriva genética cualquier miembro de la población, aun el más poderoso, muere. Como pareja de la selección natural, la deriva genética es su antítesis, pues opera de forma azarosa, mientras que la selección natural es anti-azarosa. En la naturaleza los fenómenos que generan deriva genética son catástrofes no predecibles como volcanes, inundaciones, terremotos, o meteoros y cometas entre otros. A pesar de ser fenómenos raros, los componentes de la deriva genética son importantes al borrar poblaciones adaptadas a un ambiente, abriendo los nichos a nuevos linajes. Un ejemplo de eso fue el meteorito que impactó Chicxulub hace 65 millones de años (Frohlich, 2014; Manning, 2001), el cual junto a una serie de explosiones volcánicas aniquilaron al linaje dominante en la época, los dinosaurios no aviares. Cuando el ecosistema recuperó su equilibrio, serían los mamíferos y los dinosaurios aviares los que retomarían el control de los ecosistemas. Evidentemente Wright realiza sus aportes desde una perspectiva de matemáticas duras.
👉 El paisaje adaptativo
Su segundo aporte es la idea del paisaje adaptativo en 1932. Los individuos vivos no pueden abarcar todas las posibilidades de combinaciones de alelos teóricamente posibles. En tal caso, la población posee una variabilidad genética real que es inferior a la teórica. Del mismo modo algunas combinaciones teóricas son más aptas para la población, lo que implica que los individuos más cercanos a dichas combinaciones tienen más éxito reproductivo, y sus descendientes tienden a ubicarse más cerca de estas zonas. A las zonas que representan combinación genéticas teóricas exitosas se las llama picos adaptativos. Pero a las combinaciones que llevan a la muerte prematura de los individuos se las llama valles adaptativos. Las poblaciones naturales tienden a acercarse a los picos mediante su diversificación natural, mediada por la recombinación genética y las mutaciones.

Figura 17.4. La idea del paisaje adaptativo es que cada alelo genera un cambio en el potencial de supervivencia, la selección natural opera eliminando los alelos de baja aptitud, y preservando los de alta aptitud.
👉 Impacto de las nuevas tecnologías moleculares
Las nuevas tecnologías químicas como la electroforesis permitieron medir en el campo, los cambios de frecuencias genéticas y poner a prueba las ideas de las matemáticas evolutivas. Debido a que las caminatas en paisajes adaptativos son aleatorias las especies pueden alcanzar picos adaptativos inferiores de otros que son potencialmente más altos, y lo que es más, dos especies que evolucionan independientemente para la misma función pueden terminar en picos adaptativos de eficiencia diferente, permitiendo que una sea evidentemente superior a la otra. Las especies invasoras son aquellas que han alcanzado picos adaptativos superiores y pueden eventualmente desplazar por competencia a las especies nativas que han evolucionado en aislamiento si logran invadir sus ecosistemas aislados.
18. La segunda generación de sintéticos evolutivos
18. La segunda generación de sintéticos evolutivos
La aparición de la segunda generación de sintéticos evolutivos se debió a varios eventos importantes en el desarrollo de la biología y la genética durante las décadas de 1940 y 1950.
Uno de los eventos más significativos fue la Segunda Guerra Mundial, que interrumpió la investigación científica en Europa y llevó a muchos científicos europeos a emigrar a Estados Unidos, donde se establecieron en instituciones académicas y centros de investigación. Esta diáspora científica contribuyó a la mezcla de diferentes disciplinas y enfoques en la síntesis evolutiva moderna.
Otro evento importante fue el descubrimiento de la estructura del ADN en 1953 por James Watson y Francis Crick. Este descubrimiento revolucionó la genética y sentó las bases para la investigación en biología molecular y evolución molecular, lo que permitió a la segunda generación de sintéticos evolutivos integrar la genética molecular en la teoría evolutiva.
Además, durante las décadas de 1940 y 1950 se produjeron importantes avances en la paleontología, la ecología y la sistemática, lo que permitió a los científicos comprender mejor los procesos de especiación y la evolución de los organismos en un contexto más amplio.
En resumen, la aparición de la segunda generación de sintéticos evolutivos fue el resultado de una mezcla de factores, incluyendo la Segunda Guerra Mundial, el descubrimiento de la estructura del ADN y los avances en diferentes disciplinas biológicas, que permitieron una comprensión más amplia y profunda de la evolución de los seres vivos.
18.1 Dobzhansky
Theodosius Dobzhansky 1900-1975 y sus maestros rusos ya estaban persiguiendo la síntesis genética-evolución-matemáticas, el problema fue el incidente de Lysenko cuando la Unión Soviética decidió que la genética matemática era una ciencia burguesa que iba en contra de los ideales socialistas-comunistas, así que todo el que trabajaba en esa línea de investigación debería, bien trabajar en otra cosa; afrontar un juicio y posterior fusilamiento; o escapar a occidente.
En 1937 Dobzhansky realiza un resumen de los trabajos anteriores con su publicación “Genética y el Origen de las especies” (Dobzhansky & Dobzhansky, 1982). El objetivo de Dobzhansky era transferir las matemáticas de los sintéticos de primera generación a los ecólogos de campo. Esto fue importante ya que las matemáticas de la TSE son de alto grado de dificultad, “y es por eso que no las vemos en este curso introductorio”, de hecho, muchos biólogos aún se inscribían en la carrera de biología al pensar que era la menos matemática de las tres Ciencias de la Naturaleza. Con Dobzhansky la biología alcanza su estatus de ciencia madura, con una estructura matemática que puede contrastarse experimentalmente (Dobzhansky, 1949; Hossfeld & Olsson, 2002).
El trabajo de Theodosius Dobzhansky reveló que la variabilidad genética de las poblaciones naturales era mucho mayor de lo que los teóricos matemáticos habían estimado inicialmente, y que las sub-poblaciones genéticamente distintas “razas biológicas” eran importantes en la evolución. En 1937, Dobzhansky populariza entre los ecólogos la idea de que las mutaciones son la principal fuente de cambios evolutivos junto con la recombinación genética. Posteriormente la deriva genética, la selecciona natural, la inmigración y la emigración, la hibridación y la no hibridación determinarían la aparición de nuevas especies.
18.2 Ford
Uno de los ecólogos influidos por Dobzhansky y Fisher fue Edmund Brisco Ford 1901-1988 quien en 1940 puso a prueba las ideas de Dobzhansky y antecesores con su trabajo de campo sobre especies silvestres de mariposas y polillas, especialmente al cuantificar la selección natural.

Figura 18.1. Edmund Brisco Ford (23 de abril de 1901 - 2 de enero de 1988) fue un genetista ecológico británico. Fue un líder entre los biólogos británicos que investigaron el papel de la selección natural en la naturaleza. Cuando era un escolar, Ford se interesó en los lepidópteros, el grupo de insectos que incluye mariposas y polillas. Luego estudió la genética de las poblaciones naturales e inventó el campo de la genética ecológica. Ford recibió la Medalla Darwin de la Royal Society en 1954.
Ford logra identificar los polimorfismos genéticos en la naturaleza que habían sido predichos por Fisher y Haldane casi dos décadas antes. Una vez que Ford y Dobzhansky demuestran la coherencia de la TSE, otros biólogos comienzan a sumarse a la síntesis, ya no desde la genética, la ecología o las matemáticas, sino de ramas de la biología más clásicas como la botánica y la paleontología (Clarke, 1995; Ford, 1945; Huxley, 1929; Huxley, Hardy, & Ford, 1954).
18.3 Schmalhauzen

Figura 18.2. Iván Ivánovich Shmalgauzen ( Kiev, 23 de abril de 1884–Leningrado, 7 de octubre de 1963). Morfólogo y embriólogo evolutivo ruso. Junto con C. H. Waddington fue un gran abogado de la integración de la biología del desarrollo en la teoría evolutiva, siendo considerado uno de los precedentes principales de la moderna evo-devo “biología evolutiva del desarrollo”.
Iván Ivánovich Shmalgauzen 1884-1963 en 1941 señala una de las deducciones del paisaje adaptativo, cuando una especie alcanza un pico adaptativo en un ambiente estable, la selección natural mantendrá el fenotipo general estable por un largo periodo, a esto se lo conoce como la hipótesis de la selección estabilizante, y su opuesto es la hipótesis de la reina roja que veremos en capítulos posteriores (Dobretsov, Kolchanov, & Suslov, 2008; Hoßfeld, Olsson, & UH, 2001; Kureichik, Kureichik, & Sorokoletov, 2007; Ozernyuk, 2010).
18.4 Huxley el joven
No confundir con el primer Huxley que fue contemporáneo de Darwin.

Figura 18.3. Julian Sorell Huxley fue un biólogo evolutivo, escritor, humanista, eugenista internacionalista británico, conocido por sus contribuciones a la popularización de la ciencia a través de libros y conferencias. Fue el primer director de la Unesco y fue nombrado caballero británico en 1958. Apoyó las teorías de la selección natural, fue una de las figuras destacadas de la síntesis evolutiva moderna. Fue el secretario de la Sociedad Zoológica de Londres y uno de los fundadores del Fondo Mundial para la Naturaleza.
Julian Sorell Huxley 1887-1975 en 1942 se encarga populariza la evolución matematizada despojándola de sus matemáticas y ofrece a cambio ejemplos ecológicos de los principios evolutivos para poder divulgar la nueva teoría de la evolución al público general que no entiende mucho de matemáticas. Su trabajo convenció a una nueva generación de biólogos de la realidad de la evolución y la importancia de su estudio. En el mismo libro de 1942 Huxley bautiza a esta evolución matematizada con el nombre con el que se la conoce actualmente, el libro se llamó “Evolución la Síntesis Moderna” (Huxley, 1942).
18.5 Mayr
Este es el autor que mas ha influenciado mi visión sobre los seres vivos.

Figura 18.4. Ernst Walter Mayr, 5 de julio de 1904, Kempten, Alemania – 3 de febrero de 2005, Bedford, Massachusetts Estados Unidos), fue uno de los más notables biólogos evolutivos del siglo XX. Fue, además un renombrado taxónomo, explorador de los trópicos, ornitólogo, historiador de la ciencia y naturalista. Su obra contribuyó a la revolución conceptual que llevó a la síntesis moderna de la teoría de la evolución y al desarrollo del concepto biológico de especie.
Ernst Walter Mayr 1904-2005 en 1942 trabajó sobre el origen de nuevas especies por aislamiento geográfico y sobre la clasificación de los seres vivos desde un punto de vista evolutivo. Ambos temas serán trabajados con mayor énfasis en capítulos posteriores, pero hay que dejar claro que Mayr es uno de los biólogos más influyentes en la segunda mitad del siglo XX, siendo tal vez el autor que más ha influido en el escritor del presente texto (Bock, 1994, 2005; Mayr, 1997).
18.6 Simpson
George Gaylord Simpson 1902-1984 en 1944 introduce la TSE a la paleontología que hasta ese momento había estado fuertemente influida por las ideas neo-lamarkianas de evolución lineal perfeccionante y a saltos conocida simplemente como ortogénesis. Los paleontólogos ortogenesistas organizaban los fósiles linealmente como cadenas progresivas de perfeccionamiento, pero esto generaba paradojas temporales, dado que muchos organismos extintos en estas líneas en realidad habían convivido por periodos largos de tiempo geológico o se encontraban vivos en la actualidad junto con sus supuestos descendientes.

Figura 18.5. George Gaylord Simpson (16 de junio de 1902 - 6 de octubre de 1984) fue un paleontólogo, geólogo y biólogo estadounidense. Especialista en los mamíferos del Mesozoico y del Cenozoico, Simpson es, junto con Theodosius Dobzhansky y Ernst Mayr, uno de los principales teóricos de la Teoría evolutiva sintética. Tempo and Mode in Evolution y The Meaning of Evolution son sus obras más representativas.
Desde Simpson, toda representación evolutiva de los fósiles deberá organizase como árboles de ramificación “dendrogramas” que idealmente tengan en cuenta el tiempo geológico en que aparecen los fósiles “dendrogramas cronocalibrados”. El problema de muchos libros de texto es que estas dos imágenes diferentes de la evolución se mezclan, por lo que es común encontrar imágenes de sucesiones lineales ortogenesistas con etiquetas que la señalan como procedentes de la TSE o simplemente como darwinismo, como, por ejemplo, en la marcha de los caballos o en la marcha de los primates. Esta idea errónea es tan potente que, al colocar evolución o darwinismo en google, lo que aparecerá será esta marcha ortogenesista, en lugar de las verdaderas ideas darwinistas de árbol de ramificación (Laporte, 2000; Stewart & Barber, 1959).
18.7 Stebbins
George Ledyard Stebbins 1906-2000 en 1950 introduce el punto de vista de las plantas, señalando mecanismos evolutivos propios de las plantas como las poliploidias, que permite generar nuevas especies rápidamente (Smocovitis, 1997, 2006; Solbrig, 1979). La manipulación consciente de la evolución de las plantas permitió un rápido mejoramiento de las especies cultivables para que sobrevivieran en terrenos áridos con nutrientes adecuados proporcionados en fertilizantes. La omisión de los principios evolutivos por el contrario conlleva a cosechas con eficacia muy limitada y a la hambruna como sucedió en la Rusia comunista de la mano de Trofim Lysenko (Buiatti, 2005; Singh, 2015). Debido que todo fenómeno biológico puede ser entendido y conectado con otros por medio de la TSE, Dobzhansky afirmó en 1973 que “Nada en biología tiene sentido excepto bajo la luz de la evolución. La TSE cohesiona la genética, la biología de sistemas y órganos con la ecología (Dobzhansky, 2013).

Figura 18.6. George Ledyard Stebbins jr ( 6 de enero de 1906-19 de enero de 2000) fue un botánico y genetista estadounidense, considerado uno de los biólogos más destacados del siglo XX uno de los miembros fundadores de la síntesis evolutiva moderna. Su libro de 1950 Variation and Evolution in Plants (Variación y Evolución en Plantas) llevó la botánica dentro de la síntesis.
18.8 Consolidación de la síntesis y enfrentamiento final con la ortogénesis
Una idea problemática que mal interpreta la TSE y la confunde con otras líneas de pensamiento se resume en la siguiente acotación ¿por qué si descendemos de los monos, aún hay monos el día de hoy? Tal idea se desprende de la famosa imagen de la marcha de los primates y que resume algo llamado neo-lamarckismo u ortogénesis, dos de las teorías evolutivas que compitieron con el neodarwinismo en la primera década del siglo XX e incluso con la TSE a mediados de siglo. Ambas fueron hipótesis biológicas según la cuales la vida tiene una tendencia innata a evolucionar de un modo unilineal debido a alguna "fuerza directriz", ya sea interna “ortogenesista” o externa “neolamarckista”.
La hipótesis tiene bases filosóficas de esencialismo, finalismo, la teleología y la teología, y propone una fuerza intrínseca que lentamente transforma las especies desde lo menos perfecto a lo más perfecto. Ronald Fisher puso fin a la ortogénesis cuando arguyó en su libro de 1930 “La teoría genética de la Selección Natural” en favor de la herencia particulada. La ortogénesis fue particularmente aceptada por los paleontólogos que vieron en sus fósiles un cambio direccional. La hipótesis ortogenética / neolamarckista comenzó a colapsarse cuando quedó claro que no podría explicar los patrones encontrados por nuevos paleontólogos en el registro de fósil, el cual era no lineal.
La hipótesis fue generalmente abandonada cuando no se encontró ningún mecanismo que explicaba el proceso, a diferencia de la teoría de la evolución por selección natural, la cual prevaleció. La TSE, en la cual los mecanismos genéticos de la evolución fueron encontrados, refutaron la hipótesis de la ortogénesis totalmente. Cuanto mejor se conocían estos mecanismos recién descubiertos, más patente se hacía que era imposible que la herencia pudiera tener dirección lineal prefijada a la perfección, aunque algunos teólogos como Teilhard de Chardin 1881-1955, un paleontólogo jesuita, la seguían defendiendo para 1959 (Pi, 2017).
19. Selección sexual
19. Selección sexual
La selección sexual es un proceso mediante el cual ciertos rasgos que mejoran la capacidad de un individuo para competir por, atraer o elegir una pareja son favorecidos y, por lo tanto, se vuelven más comunes en una población con el tiempo. Es una forma de selección natural que actúa específicamente en los rasgos relacionados con el éxito reproductivo, y puede operar a través de dos mecanismos principales:
👉 Selección intrasexual, que implica la competencia entre miembros del mismo sexo por el acceso a parejas, y
👉 Selección intersexual, que implica la elección de pareja por parte de un sexo (generalmente las hembras) basada en rasgos exhibidos por los miembros del sexo opuesto.
En ambos casos, el resultado final es un aumento en la frecuencia de ciertos rasgos en una población, lo que puede implicar un costo en términos de otros rasgos que son menos importantes para el éxito reproductivo. Ejemplos de rasgos seleccionados sexualmente incluyen ornamentación elaborada, colores brillantes, tamaño grande y exhibiciones de cortejo complejas, entre otros.
19.1 El handicap o desventaja evolutiva
La selección sexual fue propuesta por primera vez por Charles Darwin en El origen de las especies (Darwin, 1866)y se desarrolló en El origen del hombre y la selección en relación con el sexo (Darwin, 1872), ya que consideraba que la selección natural por sí sola no podía explicar ciertos tipos de adaptaciones de no supervivencia. Una vez le escribió a un colega que "la vista de una pluma en la cola de un pavo real, cada vez que la miro, me enferma" pues la selección natural, entendida como interacciones ecológicas de competencia, depredación o simbiosis no explicaba la existencia de caracteres suicidas, que hacían más difícil la supervivencia. Los biólogos han llegado a dar un nombre a estas características desventajosas como Handicap “Desventaja”.

Figura 19.1. A diferencia de lo que se podría esperar, Darwin no llegó inmediatamente al concepto de selección sexual, y antes de eso decía que la visión de un pavo real (Pavo cristatus) le daban ganas de vomitar ya que la selección natural por interacciones ecológicas exteriores a la especie, como la relación depredador-presa no lograban explicar el plumaje de los machos.
Las colas de los pavos reales son una desventaja evolutiva o “handicap” para su propia supervivencia, son muy grandes y no se camuflan, por lo que solo los machos más poderosos pueden sobrevivir con ellas ante los depredadores, además su sistema inmune debe ser superior, ya que los parásitos afectarían su brillo y suavidad. Sin embargo, sin colas ostentosas no hay sexo, las hembras no se aparearán con machos con colas mediocres. La reproducción es el objeto fundamental sobre el que se define a la selección natural, aunque seas el mejor sobreviviente, si no te reproduces no existes evolutivamente (Lumley et al., 2015; Pomiankowski, 1987; Zahavi, 1975).

Figura 19.2. Espécimen de lagarto cola de lático (Cnemidophorus) que se reproduce por partenogénesis.
Para las mantis “orden Mantodea” la situación se lleva al extremo, pues los machos que se han reproducido pueden servir mejor a la causa de sus crías sirviendo como alimento, al ser devorados por las hembras (Fisher, Cornell, Holwell, & Price, 2018).
19.2 La hipótesis de la reina roja y la importancia del macho
Algunas especies de lagartos cola de látigo “género Cnemidophorus” se reproducen por medio de la partenogénesis en la cual la población solo posee hembras autoreproductoras (Larose, Parker, & Schwander, 2018). Por lo general los machos no comparten las responsabilidades de la crianza en los animales, y al existir especies de vertebrados que se reproducen sin machos, se debe plantear una pregunta fundamental ¿Cuál es la utilidad de la reproducción sexual? y ¿para qué sirven los machos aparte de competir por recursos con las hembras de sus respectivas especies?
En el desierto de Sonora en Méjico se pueden encontrar especies de peces del género “Poeciliopsis spp.,” con subpoblaciones sexuales y asexuales que viven en un mismo estanque, allí los reproductores asexuales son más atacados por parásitos tremátodos “gusanos planos” del género “Neascus sp.,” que los reproductores sexuales (Leberg & Vrijenhoek, 1994). Una de las consecuencias del proceso evolutivo se conoce como la hipótesis de la Reina Roja propuesta por Leigh Van Valen en 1973 (Liow, Van Valen, & Stenseth, 2011; Van Valen, 1973), la cual sostiene que la mayoría de la evolución ocurre para que la especie pueda sobrevivir a duras penas, manteniéndose en su nicho ecológico sin cambios aparentes, es decir, aun cuando en apariencia la evolución se detiene cuando las especies se adaptan a su ambiente, en el interior la evolución sigue en marcha para enfrentar a depredadores y parásitos que también evolucionan en cada generación, esta condición también se conoce como carrera armamentista evolutiva (Liow et al., 2011).
Cuando una especie deja de evolucionar para responder a los desafíos de las interacciones ecológicas: depredador-presa, competencia, parasitismo, comensalismo y relaciones con factores abióticos; dicha especie se extingue. Las poblaciones con reproducción sexual poseen sistemas inmunes únicos y más fuertes que pueden enfrentar a los parásitos de manera efectiva, disminuyendo su transmisión en la población. La reproducción sexual solo funciona para acelerar la evolución si los reproductores no están emparentados. Si los reproductores sexuales proceden de una misma familia no habrá recombinaciones genéticas nuevas, lo que hace que evolucionen más lento y que acumulen malformaciones congénitas, fenómeno conocido como endogamia o incesto. Cuando una población endogámica recupera su diversidad o variabilidad con la introducción de individuos que no son parientes, recuperan su capacidad de recombinación genética y los beneficios de la reproducción sexual (Kutschera & Niklas, 2004).
19.3 La profase de la meiosis 1
Ya hablamos de la reproducción celular denominada meiosis durante el capítulo de genética clásica, donde la aplicamos justo como corresponde a su etimología siendo esta la reproducción reductora es decir disminuyendo la cantidad de cromosomas de una célula a la mitad. Sin embargo, la meiosis también posee un truco especial, durante su primera división denominada la meiosis 1 los cromosomas individuo que tienen la misma forma y función “cromosomas homólogos” van a interactuar de manera íntima entre sí intercambiando brazos cromosómicos, de forma tal que van a generar cromátidas normales y cromátidas recombinantes. A este proceso se lo denomina la recombinación genética.

Figura 19.3. La existencia de la recombinación genética fue detectada accidentalmente por el equipo de Morgan, al percatarse de que los cálculos esperados para la segregación independiente vs para la segregación ligada no concordaba exactamente en ninguna de las dos hipótesis anteriores con los datos obtenidos, pues algunos grupos de genes que parecían cumplir casi perfectamente la segregación independiente, a veces se comportaban como si estuvieran ligados, y viceversa (Lobo & Shaw, 2008).
Recuerde que una cromátida es lo que se va a convertir en un cromosoma simple cuando el cromosoma duplicado con forma de x se rompe durante la finalización de la meiosis 1.

Figura 19.4. Partes de un cromosoma. Los cromosomas con forma de x son entidades duplicadas, es decir la cromátide izquierda posee en esencia la misma información genética que la cromátide derecha. Ambas estructuras han evolucionado para romperse cuando proteínas especiales del citoesqueleto se anclen al cinetocoro y comiencen a jalar en lados opuestos para llevarse una cromátida a los polos opuestos de las nuevas células.
La importancia de esa recombinación genética y la formación de los cromosomas recombinantes radica en que esto incrementa la diversidad posible de la descendencia sin someterse al riesgo de la mutación aleatoria, incrementando la biodiversidad de la población, especialmente en especies con reproducción lenta en comparación con los parásitos que los atacan que pueden reproducirse mucho más rápido.
A su vez un incremento en la biodiversidad genética de un individuo también le asegura un sistema inmune mucho más poderoso. Así pues, la importancia del macho radica en que permite recombinar los genes.
19.4 Tipos
Charles Darwin clasificó la selección sexual en dos modos de acción:
👉 Lucha directa o indirecta de los machos por acaparar a las hembras.
La lucha directa puede ocasionarse por combates en los cual es el perdedor puede resultar herido o muerto pero cuyo resultado más importante es que no pueden generar descendencia, si el macho perdedor no consigue en algún momento de su existencia ganar algún combate para acceder a alguna hembra, entonces su material genético se perderá para siempre (Clutton-Brock, 2007; Ercit & Gwynne, 2015; D. J. Futuyma, 2005; Heywood, 1989).
La lucha indirecta de los machos puede clasificarse a su vez en dos tipos, la lucha ritual y el acaparamiento de recursos. En un combate ritual los machos se enfrascan en competiciones en los cuales exhiben sus caracteres sexuales secundarios, así como su tamaño y coordinación de forma tal que uno de los dos machos decida que entrar en combate físico directo no es recomendable (Clutton-Brock, 2007; Ercit & Gwynne, 2015; D. J. Futuyma, 2005; Heywood, 1989).
El acaparamiento de recursos implica que los machos van a competir por los recursos clave que necesite una hembra o grupo de hembras para poder realizar sus labores de crianza. En este caso el combate no se da por la hembra en sí sino por un territorio de crianza (Clutton-Brock, 2007; Ercit & Gwynne, 2015; D. J. Futuyma, 2005; Heywood, 1989).
👉 Elección femenina
Por lo general debido a que la hembra invierte más energía en producir los gametos, es ella quien realiza un proceso denominado selección sexual. Las hembras seleccionan caracteres que en el ambiente disminuyen la supervivencia de los machos, así que solo los machos más poderosos pueden sobrevivir y reproducirse. La selección sexual por lo tanto explica la aparición del dimorfismo sexual, es decir caracteres diferentes entre machos y hembras, y también explica por qué los caracteres masculinos son deletéreos o desventajosos como:
👉 la melena del león aumenta el acaloramiento, y disminuye la capacidad de camuflaje,
👉 la cola del pavo real aumenta el peso, disminuye la capacidad de camuflaje y la agilidad,

Figura 19.5. El dimorfismo sexual debe ser explicado a través de una forma especial de selección natural llamada selección sexual.
👉 la cornamenta de muchos animales especialmente aquellas que solo crecen en la etapa reproductiva, son molestas pues los machos gastan energía en generarlas, y su crecimiento genera pucha picazón, lo que los distrae de los depredadores,
👉 la espalda plateada en los gorilas disminuye la capacidad de camuflaje
👉 o la barba en el ser humano que aumenta la propensión a adquirir parásitos como los piojos y pulgas.
(Clutton-Brock, 2007; Ercit & Gwynne, 2015; D. J. Futuyma, 2005; Heywood, 1989).
Sin embargo, en ocasiones no solo basta con heredar un buen sistema inmune, si el macho se queda a cuidar a la hembra y a las crías, estas aumentarán su probabilidad de supervivencia más allá de sus sistemas inmunes, por lo que en la selección sexual por elección femenina también puede estar involucrado la selección de un comportamiento protector protector a largo plazo (Clutton-Brock, 2007; Ercit & Gwynne, 2015; D. J. Futuyma, 2005; Heywood, 1989)
19.5 Historia del concepto
Alfred Russel Wallace se opuso en cierta medida a estos puntos de vista, principalmente después de la muerte de Darwin. Aceptó que la selección sexual podía ocurrir, pero argumentó que era una forma de selección relativamente débil. Argumentó que las competencias macho-macho eran formas de selección natural, pero que la coloración "monótona" de la pava real es en sí misma adaptativa como camuflaje. En su opinión, atribuir la elección de pareja a las hembras era atribuir la capacidad de juzgar los estándares de belleza a los animales (como los escarabajos) demasiado subdesarrollados cognitivamente para ser capaces de tener sentimientos estéticos (Wallace, 1892).
Las ideas de Darwin sobre la selección sexual fueron recibidas con escepticismo por sus contemporáneos y no se consideraron de gran importancia, hasta que en la década de 1930 los biólogos decidieron incluir la selección sexual como un modo de selección natural en la síntesis evolutiva moderna. Este caso es interesante dentro de la filosofía de la ciencia, siendo un concepto que propone uno de los autores originales de una teoría, pero que no se acepta por la comunidad científica contemporánea, por ende, la selección sexual no es considerada como un concepto clave dentro del darwinismo original o en el neodarwinismo, pero que se vuelve importante en la síntesis moderna (G. Miller, 2011).
Solo en el siglo XXI se han vuelto más importantes en biología; el concepto ahora se considera una forma especial de selección natural generada por interacciones intraespecíficas. Recuerde que las interacciones intraespecíficas son modos estándar de comportamiento entre individuos de una misma especie, los cuales generan patrones de selección, sin la necesidad de una mente inteligente que seleccione cuales formas físicas ofrecen ventajas a dichas interacciones estandarizadas.
Un estudio de diez años, variando experimentalmente la selección sexual en los escarabajos de la harina manteniendo constantes otros factores, mostró que la selección sexual protegía incluso a una población endogámica contra la extinción (Lumley et al., 2015).
20. Carrera armamentista y coevaluación
20. Carrera armamentista y coevaluación
En biología evolutiva, una carrera armamentista evolutiva es una lucha continua entre conjuntos competitivos de genes coevolutivos, rasgos fenotípicos y de comportamiento que desarrollan adaptaciones crecientes y contraadaptaciones entre sí, asemejándose a una carrera armamentista.
20.1 Tipos
Existen varios tipos de carrera armamentista en la cual los individuos pugnan por adaptaciones y contra-adaptaciones para mejorar su éxito de supervivencia y reproducción.
👉 Interespecificas
Son las relaciones de conflicto que se entablan entre individuos de diferentes especies, como en una carrera armamentista evolutiva entre una especie depredadora/parásita y su presa/anfitrión/hospedante (Vermeij, 1993) o entre competidores por un mismo recurso (Thompson, 1994).
👉 Intraespecíficas
Alternativamente, la carrera armamentista puede ser entre miembros de la misma especie, como en el modelo de comunicación de resistencia a la manipulación/ventas (Dawkins & Krebs, 1979) o como en la evolución desbocada o los efectos de la Reina Roja. Un ejemplo de una carrera armamentista evolutiva es el conflicto sexual entre los sexos, a menudo descrito con el término fugitivo Fisheriano, en el cual, la selección sexual genera caracteres con una desventaja muy marcada (Fieher, 1922; Pomiankowski & Iwasa, 1998).
20.2 Carrera armamentista depredador-presa o parásito-presa
La carrera armamentista depredador/parásito-presa/anfitrión es un proceso evolutivo en el que los depredadores o parásitos y las presas desarrollan, a lo largo del tiempo, adaptaciones cada vez más complejas y sofisticadas para cazar o escapar/inmunizarse, respectivamente. Este proceso se debe a la presión selectiva que ejercen unos sobre otros, en la que las presas que tienen características que les permiten escapar de los depredadores tienen más posibilidades de sobrevivir y reproducirse, mientras que los depredadores que tienen características que les permiten cazar con mayor éxito tienen más posibilidades de sobrevivir y reproducirse también (Cortez & Weitz, 2014; Holding, Biardi, & Gibbs, 2016; Scheuerl, Cairns, Becks, & Hiltunen, 2019; Watve, 1998).
Como resultado de esta competencia evolutiva, las presas desarrollan estrategias para evitar ser capturadas, como camuflaje, mimetismo, velocidad, agilidad y otras defensas, mientras que los depredadores desarrollan estrategias para superar estas defensas, como mejoras en la velocidad, la fuerza, la agilidad, la visión, el olfato y otras habilidades para detectar y capturar a sus presas (Cortez & Weitz, 2014; Holding et al., 2016; Scheuerl et al., 2019; Watve, 1998).
Este proceso se llama "carrera armamentista" porque los depredadores y las presas están en una constante competición para superar las defensas y estrategias del otro, como si se tratara de una carrera en la que cada uno trata de superar al otro. Este proceso también es conocido como "coevolución depredador-presa" (Cortez & Weitz, 2014; Holding et al., 2016; Scheuerl et al., 2019; Watve, 1998).
20.3 Ejemplos de la carrera depredador-presa
Hay dos presiones de selección o interacciones ecológicas que son particularmente notables a la hora de entender la selección natural, la relación depredador-presa y la relación de simbiosis parasítica, debido a que los dos miembros de la interacción evolucionan mecanismos para combatirse o para convivir de manera más prolongada.
Las adaptaciones generadas por interacciones de dos o más especies se conocen como coevolución. En la coevolución de la relación depredador-presa tenemos varios ejemplos de una alta especialización, en la cual un depredador se especializa en consumir una presa que nadie más puede, como en el caso del tritón granuloso “Taricha granulosa” que solo puede ser consumido por la serpiente de jarretera “Thamnophis sirtalis” y otras serpientes del género “Thamnophis spp.”

Figura 20.1. Ejemplar del tritón granuloso (Taricha granulosa).
Aunque estas serpientes no son venenosas, han evolucionado independientemente genes que les confiere resistencia al veneno de los tritones, los cuales han ido evolucionando en cada generación, de forma tal que las serpientes modernas son más resistentes al veneno y los tritones modernos son más venenosos (Brodie III & Brodie Jr, 1991; Hanifin, Yotsu-Yamashita, Yasumoto, & Brodie, 1999; Williams, Hanifin, Brodie, & Brodie III, 2010).

Figura 20.2. Ejemplar de la serpiente de jarretera (Thamnophis sirtalis).
La clave del proceso evolutivo es que los diferentes miembros de una especie respondan de manera diferente al mismo estímulo, por lo que los biólogos evolutivos prestan mucha atención a detectar y cuantificar las diferentes respuestas a una presión de selección como la respuesta al veneno de una presa. Las carreras armamentistas cobran un precio a los depredadores, pues muchas veces especializarse en una presa les impide cazar otras o los hace más vulnerables a sus propios depredadores, dado que la evolución no es un proceso de perfeccionamiento, por lo general tenemos organismos imperfectos con adaptación óptimas, pero con varios puntos débiles.

Figura 20.3. Ejemplar de guepardo (Acinonyx jubatus).
Por ejemplo los guepardos “Acinonyx jubatus” solo pueden cazar además de presas pequeñas a la ágil gacela de Thompson “Eudorcas thomsonii”, pero a cambio no pueden cazar otras grandes presas, tienen poca grasa corporal por lo que no pueden aguantar hambre por mucho tiempo (Bro‐Jørgensen, 2016). Aunque los humanos ya no tenemos miedo generalmente de los grades e impresionantes depredadores, aún sigue existiendo una relación ecológica a la que tememos, la relación de parasitismo. Los seres humanos podemos ser parasitados por un sinnúmero de tipos de seres vivos, desde artrópodos como las moscas carnívoras, pasando por gusanos redondos y gusanos planos, todo tipo de protistas como los que causan la malaria, bacterias que están en todas partes y por los virus que se encuentran entre lo vivo y o no vivo.

Figura 20.4. Ejemplar de la gacela de Thomson (Eudorcas thomsonii).
20.4 Ejemplos de la carrera parásito-presa
Las bacterias fueron por mucho tiempo los parásitos más temibles, aunque no fueran conocidos, causantes de enfermedades como la peste negra y la tuberculosis hasta que a mediados del siglo XX se culminó el desarrollo de los antibióticos descubiertos casi por accidente por Alexander Flemming. La Penicilina y los antibióticos que le siguieron fueron muy exitosos en controlar enfermedades infeccionas de tipo bacteriano, a tal punto que muchos pensaban que la humanidad había vencido a las enfermedades infecciosas de una vez por todas (Commission & Commission, 1958; Walsh, 2003; Zaffiri, Gardner, & Toledo-Pereyra, 2012). La tuberculosis es una infección bacteriana contagiosa que compromete principalmente a los pulmones, pero puede propagarse a otros órganos. La especie de bacteria más importante y representativa causante de tuberculosis es Mycobacterium tuberculosis o bacilo de Koch, el cual se pensaba había sido controlado por el uso extendido de antibióticos (O’Neill et al., 2018). La tuberculosis puede mantenerse como un comensal latente hasta que el sistema inmune del anfitrión se debilita, momento en el cual pasa a ser un parásito que destruye los pulmones y se disemina a otros órganos. El problema radica en que el mal uso de los antibióticos selecciona aquellas variantes con resistencia, y con los años estas resistencias se han acumulado haciendo que muchas enfermedades infecciones vuelvan a ser problemas de salud pública.
Hace tan sólo unas décadas, los antibióticos se consideraban medicamentos milagrosos por lo bien que curaban las enfermedades mortales. Irónicamente, muchos antibióticos se han vuelto menos eficaces precisamente porque funcionaban tan bien, que se han utilizado con demasiada frecuencia. La era de los antibióticos comenzó en 1929 con la observación de Alexander Fleming de que las bacterias no crecían cerca de colonias del hongo Penicillium sp. En los decenios posteriores a este descubrimiento trascendental, las moléculas producidas por ciertos hongos y bacterias se han utilizado con éxito para combatir enfermedades bacterianas como la tuberculosis y la neumonía, y los antibióticos han hecho que disminuya drásticamente la mortalidad asociada a muchas enfermedades infecciosas. La edad de oro de los antibióticos resultó ser corta (Commission & Commission, 1958; Walsh, 2003; Zaffiri et al., 2012).
Durante los últimos decenios, muchas cepas de bacterias han adquirido resistencia a los antibióticos debido a la presión de selección impuesta por los propios antibióticos sobre seres vivos que evolucionan muy rápido. Un ejemplo es Neisseria gonorrhoeae, la bacteria causante de la gonorrea, que se muestra a la derecha. En la década de 1960, la mayoría de los casos de gonorrea podían tratarse con penicilina o ampicilina. En la actualidad, más del 24% de las bacterias de la gonorrea de los EE.UU. son resistentes a al menos uno de los dos antibióticos y el 98% de las bacterias de la gonorrea del Sudeste de Asia son resistentes a la penicilina. Las bacterias infecciosas son mucho más difíciles de tratar que sus predecesoras de hace diez o veinte años. Los médicos echan de menos «los viejos tiempos» en que los antibióticos que recetaban curaban sistemáticamente a sus pacientes (Berkeley, n.d.).
Dadas las condiciones de tiempo, herencia y variabilidad, cualquier organismo vivo (incluidas las bacterias) evoluciona cuando se introduce una presión selectiva (en este caso, los antibióticos). No obstante, la teoría evolutiva también proporciona a los médicos y pacientes algunas estrategias específicas para retrasar que se extienda, más aún, la evolución de la resistencia a los antibióticos. Entre estas estrategias se encuentran:
👉 No utilizar los antibióticos para tratar las infecciones virales
👉 Evitar las dosis bajas de antibióticos durante periodos prolongados
👉 Cuando se trate una infección bacteriana con antibióticos, se deben tomar todas las pastillas
👉 Utilizar una combinación de medicamentos para tratar una infección bacteriana
👉 Disminuir y eliminar el uso «preventivo» de antibióticos en el ganado y los cultivos
En definitiva, reconocer que las bacterias son seres que evolucionan y comprender su evolución debería ayudarnos a controlar esa evolución, permitiéndonos prolongar el tiempo de vida útil de los antibióticos (Berkeley, n.d.).
20.5 Ejemplos de la carrera competidor-competidor
Los machos de morsa Odobenus rosmarus compiten por el acceso a las hembras y utilizan sus colmillos como arma en las luchas territoriales. A lo largo del tiempo, los colmillos de los machos se han vuelto cada vez más grandes y gruesos a medida que los competidores han evolucionado para resistir las defensas de sus rivales. Esto ha llevado a una carrera armamentista en la que los colmillos se han vuelto más grandes y más resistentes con el tiempo (Boersma & Pyenson, 2015; Emlen, 2014).

Figura 20.5. Ejemplares de morsa Odobenus rosmarus, de arriba hacia abajo, cría, hembra y macho.
La competencia entre plantas por la luz: En los bosques y otros hábitats con una gran cantidad de plantas, la competencia por la luz puede ser feroz. Las plantas que tienen éxito en la competencia por la luz tienen una mayor capacidad para fotosintetizar y, por lo tanto, pueden crecer más y reproducirse mejor. Esto ha llevado a una carrera armamentista en la que las plantas evolucionan para tener hojas más grandes y un mayor crecimiento vertical para superar a sus competidores por la luz (Keddy, 2023; Walker, Blackshaw, & Dekker, 1988).
21. Extinciones y radiaciones evolutivas
21. Extinciones y radiaciones evolutivas
La extinción en biología se refiere a la desaparición de una especie o grupo taxonómico completo de organismos vivos. Esto puede ocurrir de manera natural o como resultado de la actividad humana, y puede ser causado por diversos factores, como cambios ambientales, competencia con otras especies, enfermedades, predación, caza, entre otros (Pimm & Raven, 2000).
La extinción es un proceso importante en la evolución porque libera recursos y nichos ecológicos para que otras especies puedan evolucionar y ocupar esos espacios vacíos rápidamente. La extinción también puede desencadenar procesos evolutivos, como la radiación adaptativa, en la que las especies supervivientes pueden experimentar una diversificación rápida y una evolución adaptativa en ausencia de competidores. Además, la extinción puede tener un impacto significativo en la biodiversidad y la estabilidad de los ecosistemas, lo que a su vez puede afectar el bienestar humano y la sostenibilidad del planeta (Pimm & Raven, 2000).
Por lo tanto, la comprensión de los patrones y procesos de extinción es esencial para la biología evolutiva y la conservación de la biodiversidad (Pimm & Raven, 2000).
21.1 La extinción y el diseño inteligente
La paradoja entre el proceso de extinción y el diseño inteligente en biología se debe a la idea de que, según la afirmación del diseño inteligente, la vida en la Tierra es el resultado de la intervención divina en la creación del mundo y sus seres vivos. Esto implica que todas las especies han sido creadas por un ser supremo, y que su presencia en el planeta es el resultado de un diseño inteligente, por lo que estas están perfectamente adaptadas y en perfecto equilibrio unas con otras a menos que el ser humano las afecte. Así pues, la extinción sería culpa única del ser humano. De lo anterior se deduce que la extinción no debería ocurrir en ausencia del ser humano.
Sin embargo, la evidencia científica muestra que la extinción es un proceso natural que ha ocurrido durante la historia de la vida en la Tierra, antes de la aparición del ser humano como especie o linaje. Muchas especies han desaparecido debido a factores naturales como cambios ambientales, competencia con otras especies invasoras, enfermedades y otros eventos que no pueden ser explicados por un diseño inteligente.
Por lo tanto, la paradoja entre el proceso de extinción y el diseño inteligente en biología radica en la idea de que, si la vida en la Tierra ha sido diseñada de manera inteligente por un ser supremo, entonces ¿por qué las especies están sujetas a la extinción? Esto llevó a contemporáneos y antecesores de Darwin a cuestionar la validez del diseño inteligente.
En resumen, la paradoja entre el proceso de extinción y el diseño inteligente en biología radica en la discrepancia entre la idea de que todas las especies fueron creadas por un ser supremo de manera perfecta y la evidencia científica que muestra que la extinción es un proceso natural que ha ocurrido durante la historia de la vida en la Tierra, lo cual implica imperfecciones tan incompatibles con la vida, que causan la desaparición de especies completas (K. R. Miller, 2004; Pennock, 2003).
21.2 Generalidades del proceso de extinción
La extinción, definida como la desaparición de linajes de seres vivos es un tema común en la historia, cerca del 95-99% de las especies que han existido en el planeta se han extinto. En promedio una morfo-especie se mantiene por unos 4 millones de años. Las razones de la extinción son variadas y pueden ser causadas por cualquier interacción biológica, pero en general se da cuando los cambios en los factores abióticos son devastadores. En general los eventos de extinción son de deriva genética, eventos fortuitos como la caída de un asteroide.
Cuando un ecosistema alcanza su pico de estabilidad existen muy pocos cambios físicos entre las especies que sean evidentes, pero después de una extinción en masa las reglas cambian, al no haber competencia especies no-aptas para una función, pueden realizarla sin competencia, y adaptarse con el curso del tiempo y las generaciones, hasta llegar al pico de adaptación, cuando esto sucede en ecosistemas completos se lo denomina radiación o explosión adaptativa (Kutschera & Niklas, 2004).
21.3 Las grandes extinciones
Las cinco grandes extinciones en masa son (Hallam & Hallam, 2005):
👉Extinción del Ordovícico-Silúrico: ocurrió hace aproximadamente 443 millones de años y se estima que aproximadamente el 85% de las especies de vida marina fueron eliminadas.
👉Extinción del Devónico: ocurrió hace aproximadamente 359 millones de años y se estima que entre el 70% y el 80% de las especies desaparecieron.
👉Extinción del Pérmico-Triásico: ocurrió hace aproximadamente 252 millones de años y se estima que el 96% de las especies marinas y el 70% de las especies terrestres fueron extintas.
👉Extinción del Triásico-Jurásico: ocurrió hace aproximadamente 201 millones de años y se estima que alrededor del 20% de las especies de la Tierra fueron eliminadas.
👉Extinción del Cretácico-Paleógeno: ocurrió hace aproximadamente 66 millones de años y se estima que alrededor del 75% de todas las especies del planeta, incluidos los dinosaurios, se extinguieron.
21.4¿Y la extinción causada por el ser humano?
La "extinción del Antropoceno" se refiere a la actual tasa de extinción de especies, que es causada por la actividad humana. Aunque es cierto que la extinción en curso es una grave preocupación y una amenaza para la biodiversidad y el equilibrio ecológico del planeta, actualmente no se considera como una de las cinco grandes extinciones en masa porque la tasa de extinción actual es mucho menor que las tasas de las extinciones pasadas.
Las cinco grandes extinciones en masa ocurrieron durante periodos de tiempo geológicos específicos y fueron eventos dramáticos que involucraron una gran cantidad de especies extinguidas en un período de tiempo geológico, donde corto ya involucra millones de años. En comparación, la extinción actual se produce en un período de tiempo más corto “por lo que aún no tiene una clasificación geológica clara” y afecta principalmente a una cantidad relativamente pequeña de especies en comparación con las extinciones masivas anteriores.
Es importante señalar que, aunque la extinción del Antropoceno no se considera actualmente una de las grandes extinciones en masa, sigue siendo una preocupación importante y requiere acciones urgentes para prevenir la pérdida de especies y la disminución de la biodiversidad (Wake & Vredenburg, 2008).
Para no alargar mas este artículo, solo discutiremos algunas características de dos de las cinco grandes.
21.5 La gran muerte o extinción del Pérmico-Triásico
En la historia geológica del planeta se registran 5 grandes extinciones masivas, de las cuales la más devastadora ocurrió hace unos 250 millones de años al final del período Pérmico. Durante el Pérmico hubo una gran diversidad de arcosaurios como los listrosauros “Lystrosaurus spp.,” y los gorgonoxidos “Gorgonopsia spp.,”, pero la gran mayoría desaparece al final del Pérmico (King, 1990). Conocer las razones de una extinción en masa no siempre es fácil porque deben ser factores planetarios diversos y simultáneos. Debido a que los ecosistemas se encuentran organizados en red, una red dependiente de los factores abióticos como la temperatura, la luz y la disponibilidad de agua, cambios abruptos en estos factores afectan a muchas especies al mismo tiempo, que a la vez afectan a otras especies en la red, colapsando el sistema ecológico. Cuando un ecosistema colapsa aniquilando a muchas especies tenemos un evento de extinción masiva local.
Una extinción en masa es un evento en el que muchos ecosistemas a nivel planetario afrontan un colapso simultáneo.

Figura 21.1. Esqueleto fósil y reconstrucción artística de Lystrosaurus sp.
Después de la extinción pérmica ocurrió una radiación adaptativa en dos linajes de arcosaurios, uno de estos se especializa en un modo de vida semi-bípedo dando como resultado a los dinosaurios, mientras que el segundo es relegado a la vida nocturna y es el que dará lugar a los mamíferos (Benton, 1983). Para finales del periodo triásico los dinosaurios parecen haber ganado la carrera evolutiva llenando los nichos más importantes, lo que les permite diversificarse en animales grandes, y para el siguiente periodo, el jurásico un grupo de dinosaurios da lugar a las primeras aves primitivas. Los mamíferos sin embargo continuarán ocupando el nicho de las ratas, viviendo como ratas, teniendo forma de ratas y teniendo el tamaño de ratas (Huttenlocker, Grossnickle, Kirkland, Schultz, & Luo, 2018; Lillegraven, 1979; Lovegrove, Lobban, & Levesque, 2014).
21.6 Extinción del cretáceo-eoceno
Hace unos 65 millones de años, al final del periodo cretáceo un asteroide impacta con lo que hoy es Méjico, desencadenando una serie de eventos que destruyen los ecosistemas, destruyendo a los dinosaurios no aviares, a los reptiles voladores, a los reptiles marinos y a otras especies menos impresionantes, pero las aves y los mamíferos, así como algunos anfibios sobreviven (Kutschera & Niklas, 2004).
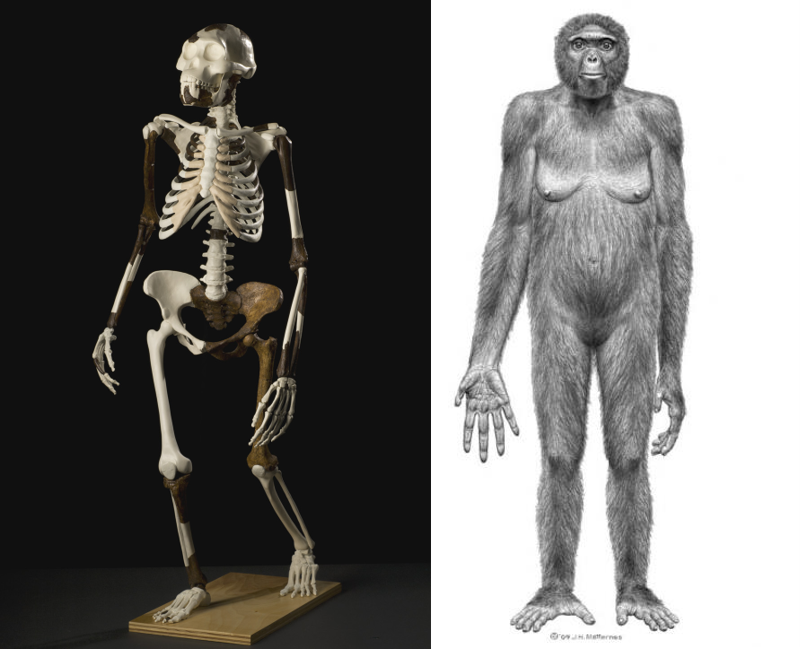
Figura 21.2. Esqueleto fósil parcialmente reconstruído y reconstrucción artística de Australopithecus afarensis, espécimen apodado Lucy.
Después del evento cretáceo-terciario hay que esperar a que la vida vegetal retorne para que los mamíferos pasen de un modo de vida de ratas a diversificarse en su propia radiación adaptativa, al igual de las aves. Los mamíferos han demostrado ser muy adaptables, ya que existen linajes que se especializaron en modos de vida acuáticos parciales o permanentes, pero también está el hecho que dentro de uno de los linajes de mamíferos se encuentra el grupo al cual pertenece el ser humano, los primates. De los grandes primates dos géneros destacan los austrolopitecinos “Australopithecus spp.,” y los miembros del género humano “Homo spp.,” todos caracterizados por caderas en forma de cuencos y mandíbulas más pequeñas, así como por cerebros más grandes y una mayor adaptación a caminar (Grabowski, 2013). Los seres humanos hemos cambiado las reglas de juego, y nuestro efecto en los factores abióticos como la temperatura y la disponibilidad de agua están provocando una sexta extinción en masa (Braje & Erlandson, 2013).
21.7 Radiaciones evolutivas
La radiación adaptativa es un proceso que describe la rápida evolución de una o varias especies para llenar muchos nichos ecológicos. Este es un proceso de la evolución (evolución divergente en el caso de dos o más especies con origen común), cuyas herramientas son la mutación y la recombinación genética. La radiación adaptativa ocurre con frecuencia cuando se introduce una especie en un nuevo ecosistema, o cuando hay especies que logran sobrevivir en un ambiente que les era hasta entonces inalcanzable o cuando ocurre una extinción masiva. Por ejemplo, los pinzones de Darwin de las islas Galápagos se desarrollaron a partir de una única especie de pinzón que llegó a las islas donde no había competencia con especies nativas adaptadas que ya habían alcanzado su pico adaptativo. Otros ejemplos incluyen la introducción por el hombre de mamíferos predadores en Australia, el desarrollo de las primeras aves que repentinamente tuvieron la capacidad de expandir su territorio por el aire, o el desarrollo de los peces pulmonados “Dipnoi” durante el Devónico, hace cerca de 300 millones de años (Lianhai Hou, Martin, Zhou, & Feduccia, 1996; Schluter, 2000; THOMSON, 1969)

Figura 21.3. Ejemplar de un pez pulmonado moderno (Neoceratodus forsteri).
La dinámica de la radiación adaptativa es tal que, dentro de un corto período de tiempo, muchas especies se derivan de una o varias especies de ancestros comunes. De este gran número de combinaciones genéticas, sólo unas pocas pueden sobrevivir con el pasar del tiempo cuando la competencia entre las especies que intentan cumplir el mismo nicho se hace más intensa. Por tanto, una radiación adaptativa posee dos momentos:
👉 una diversificación rápida inicial que provoca que el registro fósil aparente generar nuevos fósiles casi de la nada
👉 y luego una estabilización del fenotipo a medida que las especies alcanzan su pico morfológico de adaptación. La estabilización puede involucrar la eliminación de especies por desplazamiento competitivo o el particionamiento de nichos y especialización para que especies con nichos semejantes puedan convivir.
Una radiación adaptativa indica generalmente un episodio en el que un linaje se diversifica rápidamente y los linajes recién formados desarrollan diferentes adaptaciones. Hay distintos factores que pueden desencadenar las radiaciones adaptativas, pero cada uno de ellos es una respuesta a una oportunidad; entre ellos figuran:
👉 Adaptaciones clave
aparición de una adaptación clave, que es una adaptación que permite al organismo evolucionar para explotar recurso o nicho nuevo. Una adaptación clave puede hacer accesibles a un organismo muchos nichos nuevos y proporcionar la oportunidad para una radiación adaptativa. Por ejemplo, las adaptaciones de los escarabajos pudieron haber sido desencadenadas por alimentarse de las plantas con flor.
👉 Extinciones
Al librarse de la competencia, los linajes que invaden islas pueden dar lugar a radiaciones adaptativas, debido a que los invasores no tienen competencia de otras especies. En el continente otras especies llenan todos los nichos ecológicos posibles, haciendo imposible que un linaje se separe en formas nuevas y se diversifique. Sin embargo, es posible que en una isla estos nichos estén vacíos. Las extinciones también pueden generar el efecto de nicho vacío y hacer posible una radiación adaptativa. Por ejemplo, los nichos vacantes dejados por las extinciones de dinosaurios permitieron la radiación de los mamíferos en esas posiciones en la red trófica terrestre.
👉 Particionamiento de nicho
La especialización y el particionamiento de nicho, mediante la especialización un nicho único puede subdividirse en muchos nichos nuevos. Por ejemplo, los peces cíclidos de los lagos del este de África se han diversificado en más de 600 especies y esa diversificación ha sido posible porque los diferentes linajes de peces evolucionaron para explotar alimentos diferentes (incluidos los insectos, algas, moluscos, peces pequeños, peces grandes, las escamas de otros peces e, incluso, los ojos de otros peces).
22. Grandes saltos evolutivos
22. Grandes saltos evolutivos
El concepto de "gran salto evolutivo" se refiere a un cambio significativo y abrupto en la evolución de una especie, que resulta en una transformación importante en sus características físicas, biológicas o comportamentales. Estos saltos evolutivos a menudo se producen debido a una mutación genética importante que permite la generación de una adaptación clave que se propaga rápidamente a través de la población, y puede dar lugar a una nueva especie o a una subespecie.
Aunque la mayoría de las transformaciones evolutivas son graduales y ocurren a lo largo de largos períodos de tiempo, los grandes saltos evolutivos son relativamente raros, pero pueden tener un impacto significativo en el curso de la evolución de una especie, y grupos de especies relacionadas. Algunos ejemplos de grandes saltos evolutivos en la historia de la vida en la Tierra incluyen la aparición de las primeras células eucariotas, la evolución de los mamíferos a partir de los reptiles, y la aparición de los seres humanos (BioInteractive, 2013; Szathmáry & Smith, 1995).
22.1 La importancia del registro fósil
El registro fósil es una herramienta invaluable para estudiar los grandes saltos evolutivos, ya que proporciona una visión detallada de la historia de la vida en la Tierra. Los fósiles son restos de organismos del pasado que se han conservado en las rocas y pueden proporcionar información sobre la anatomía, el comportamiento y la distribución geográfica de las especies antiguas.
En particular, el registro fósil es útil para estudiar los grandes saltos evolutivos porque permite a los científicos rastrear los cambios graduales que llevaron a la aparición de nuevas especies y la evolución de nuevas características biológicas. Por ejemplo, el registro fósil muestra cómo los primeros vertebrados evolucionaron de los peces acuáticos y se adaptaron a la vida en tierra firme, lo que dio lugar a la aparición de los primeros reptiles.
Además, el registro fósil también puede proporcionar pistas sobre las causas de los grandes saltos evolutivos, como cambios ambientales, selección natural o mutaciones genéticas. Al estudiar las capas de roca y los fósiles que se encuentran en ellas, los científicos pueden reconstruir cómo era el clima, la vegetación y otros aspectos del medio ambiente en el pasado, lo que puede ayudar a explicar por qué ciertas especies evolucionaron de ciertas maneras.
En resumen, el registro fósil es una herramienta esencial para el estudio de los grandes saltos evolutivos, ya que proporciona información detallada sobre la historia de la vida en la Tierra y permite a los científicos rastrear los cambios graduales y las causas subyacentes que llevaron a la evolución de nuevas especies y características biológicas (Kidwell & Flessa, 1995; Prothero, 2009; Stanley, 1982).
22.2 El ejemplo de las ballenas
Una de las ramas más fascinantes de la teoría evolutiva moderna es la conclusión de que la vida tiene una historia cambiante, de que los seres vivos del pasado son diferentes a los del presente, pero aun así están vinculados por relaciones de parentesco. Una de estas historias es la evolución de los cetáceos, ya que por muchos años no se pudieron encontrar ni especímenes vivos o fósiles que explicara su evolución de manera directa desde el suelo seco al mar donde viven actualmente en forma de ballenas y delfines (Nummela, Thewissen, Bajpai, Hussain, & Kumar, 2004).

Figura 22.1. Esqueleto fósil y reconstrucción paeloartística de Pakicetus spp. Para ser francos el involucrum no es la única cosa que salta a la vista cuando te acostumbras a los cráneos de las ballenas primitivas, la forma de sus dientes y hocicos son muy características.
Uno de los primeros fósiles en la transición al mar de los cetáceos descubiertos fue Pakicetus spp., en 1981 y confirmado por otros especímenes desde 1983 hasta 2010. Pakicetus spp., fue vinculado a los cetáceos por poseer una estructura del sentido del oído única llamada involucrum de la bulla timpánica ósea, que en la actualidad solo poseen las ballenas y delfines, aunque se trataba de un depredador terrestre parecido a un perro (Nummela et al., 2004). Encontrar un esqueleto completo y articulado es una rareza comparable con sacarse la lotería, lo más común es encontrarse piezas sueltas parcialmente erosionadas, probablemente restos de la merienda de algún carroñero o depredador, por lo que los paleontólogos deben ser muy pacientes.

Figura 22.2. Esqueleto fósil y reconstrucción paeloartística de Basilosauru spp.
Un ejemplo de esto fue basilosaurio (Basilosaurus spp.) una ballena primitiva con forma alargada, conocida casi totalmente para la ciencia, excepto por su pelvis que aun guardaba un secreto, una cintura pelviana completa con extremidades posteriores, es decir era una ballena con patas traseras que no servían para mucho más que para tener sexo (Gingerich, Smith, & Simons, 1990). Desde entonces se han ido descubriendo otros miembros de la familia de los cetáceos como el Indohyus, Ambulocetus, Kutchicetus, Remingtonocetus, Rodhocetus, Protocetus, Dorudon, Squalodon, Kentriodon, Aulophyseter, Brygmophyseter, Aetiocetus, Janjucetus, Cetotherium, y otros Basilosauridae entre otros. El proceso evolutivo se explica mucho por un proceso denominado reclutamiento de caracteres “cooptar” que básicamente funciona retomando características antiguas usadas para otra cosa y reajustándolas para una función aparentemente nueva, aun cuando se sigan comportando a la manera antigua, como el desplazamiento por flexión vertical de la columna en los cetáceos.
22.3 El ejemplo de los primeros tetrápodos
Otra transición importante pero esta vez del agua a la tierra fue la que realizaron los ancestros de los tetrápodos, es decir los ancestros de todos los animales con cuatro extremidades como los reptiles, aves, mamíferos, anfibios y cierta clase de peces. Las rocas que contienen la transición entre los peces primitivos y los anfibios primitivos son muy antiguas, el periodo de denomina Devónico o informalmente como “la edad de los peces”, por lo que o se encuentran muy meteorizadas por el movimiento del planeta o se encuentran en regiones muy apartadas como en Groenlandia. Antes de 1987 se pensaba que los peces habían desarrollado primero los pulmones y que las patas se habían desarrollado posteriormente como sucede como el pez saltarín del fango Periophthalmus spp., pero en 1987 Jennifer A. Clarck rastreó una pista de 1933 en Groenlandia desenterrando a Acanthostega gunnari (Coates, 1996).

Figura 22.3. Esqueleto fósil y reconstrucción paeloartística de Acanthostega gunnari.
Acanthostega sp., tenía patas bien definidas, pero la cintura pectoral era débil, y la cintura pelviana estaba desconectada de la columna “no tenían hueso sacro” por lo que sería imposible que este animal pudiera desplazarse en suelo seco, por lo que es un pez con patas y pulmones además de arcos branquiales bien definidos. Otros fósiles de esta transición se han ido descubriéndose con el tiempo como Ichthyostega sp., Tiktaalik sp., y Panderichthys sp., (Lu et al., 2012)pero aún siguen existiendo muchas preguntas, pues los ecosistemas marinos aún pueden contener fósiles relevantes.

Figura 22.4. Estructura básica del quiridio o extremidad base de los vertebrados terrestres.
Las aletas tienen formas diversas, pero existe una que destaca y se llama quiridio, se caracteriza por tener un hueso de eje con la cintura “estilopodio”, dos huesos que siguen la conexión “zeugopodio” y un conjunto de huesecillos que componen una muñeca y unas falanges “autopodio”, esta estructura fue heredada por todos los descendientes, los tetrápodos, generando extremidades estandarizadas en comparación con las aletas que son más diversas (Kardong, 2014). La transición de los quiridios primitivos como los de Panderichthys sp., a las extremidades de un anfibio se basa principalmente en la modificación de la forma y en la pérdida de algunas partes (Boisvert, 2005).
22.4 El ejemplo de la explosión cámbrica
Una de las etapas geológicas más fascinantes para los paleontólogos es el periodo cámbrico. Antes se pensaba que todos los animales habían aparecido en el cámbrico. Casi todo lo que sabemos del cámbrico proviene de un único yacimiento en Canadá conocido como Burgess Shale (Erwin & Tweedt, 2012), cuyas rocas han preservado a animales en muchos de sus detalles más finos. Algunos de los animales más representativos del cámbrico son artrópodos conocidos como los trilobites emparentados con los crustáceos, los insectos y los arácnidos. Los artrópodos del cámbrico son extraños experimentos de una radiación evolutiva que no superaron las pruebas de la competencia, como el Anomalocaris spp (Morris, 2000). Sin embargo decir que los animales aparecieron en el cámbrico es un poco problemático, pues aunque en efecto allí aparecen algunas de las primeras formas o filos, estas formas primitivas son diferentes de sus descendientes y de sus ancestros registrados en rocas más antiguas, por ejemplo los vertebrados están representados con animales que están en la transición con los invertebrados, por lo que se trata de criaturas parecidas a gusanos, con notocordios de cartílago o fibra, cráneos imperfectos, sin mandíbulas, espinas, vértebras, o extremidades como Pikaia spp (Chen, Huang, & Li, 1999).
22.5 El ejemplo de la evolución de las aves
Los fósiles eran conocidos en la época de Darwin y representaban una historia natural del pasado, pero no dan información acerca de cómo ocurren los cambios evolutivos, de allí que fósiles tan famosos como Archaeopteryx spp., (Owen, 1863)no fueran tan importantes a la hora del desarrollo de la Selección Natural. En la actualidad la genética ha avanzado mucho, y estudia complejas interacciones entre los genes, sus productos activos llamados proteínas y como es que estos interactúan para alterar un embrión para formar un organismo completo. Wiliam Bateson reportó algunas anomalías en los patrones de desarrollo embrionario de los seres vivos, y esto le convenció de que el mecanismo evolutivo debía ser genético y categórico, es decir interno y a saltos, por lo que para él la selección natural y el gradualismo darwiniano debían estar en un error.

Figura 22.5. Esqueleto fósil y reconstrucción paeloartística de Archaeopteryx spp.
22.6 Ejemplos moleculares, los genes de los ojos
Cuando los genetistas mejoraron sus métodos, comenzaron a percatarse de que la segmentación estaba controlada por genes reguladores que controlaban la formación de secciones completas o segmentos y su identidad, en la actualidad los conocemos como los genes Hox. Posteriormente los genetistas comenzaron a preguntarse si existían homologías en los genes del mismo modo que las hay en los cuerpos, por ejemplo, en los genes encargados de comandar el inicio del desarrollo de los ojos. Para eso, se suprimió el gen generador de ojos en las moscas “llamado pax-6” y se les transfirió su homólogo de los ratones, la respuesta fue que las moscas generaron ojos, pero ojos de mosca, por lo que el gen solo controla el inicio del desarrollo, no sus detalles (Harzsch & Hafner, 2006).
En la actualidad entendemos que el proceso evolutivo es poco creativo, lo que hace es reciclar procesos e información y los modifica levemente para dar resultados cualitativa y aparentemente distintos, pero en el fondo los mecanismos son los mismos. Dado que la única manera de tener los mismos genes es por relaciones de parentesco, se deduce que los genes Hox presentes en todos los animales y que controlan la arquitectura del cuerpo debieron haber sido heredados por un ancestro común (Montavon, Le Garrec, Kerszberg, & Duboule, 2008).
22.7 La aparición del ser humano
En cuanto a la evolución humana temprana que ocurrió entre 8-4 millones de años la información es fragmentaria, se sabe de algunos primates grandes diferentes de chimpancés (Pan troglodytes) que vagaron por África y por el sur de Europa “en aquella época había puentes de tierra”, pero los fósiles decididamente humanoides comienzan a aparecer hacer unos 3,9 millones de años con los austrolopitecinos, y el género Homo spp., solo hace su aparición hace unos 1,5 millones de años. El estudio de los primates africanos primitivos es difícil ya que sus fósiles son escasos, por lo que se deben emplear analogías como los lémures que presentan diversos modos de locomoción.
Otro aspecto que complica el estudio de la evolución humana es que de todo nuestro género y nuestra familia de seres vivos somos los únicos supervivientes, los parientes más cercanos que sobrevivieron al día de hoy son el chimpancé común (Pan troglodytes) y el bonobo (Pan paniscus) con una arquitectura corporal casi idéntica, incluso con los mismos tipos de sangre, por no mencionar la masiva similitud genética (Bourlière & Howell, 2013). Los chimpancés no son nuestros ancestros, y son tan diferentes del ancestro común como nosotros, por ejemplo, al desplazarse en tierra lo hacen con los nudillos, una especialización de la que carecería el ancestro común con los humanos.

Figura 22.6. De izquierda a derecha: bonobos hembra y macho; chimpancé común macho y hembra.
22.8 Importancia de los genes Hox
Los genes homeóticos o Hox participan en el desarrollo embrionario de los organismos y que determinan la identidad de los segmentos o partes individuales del embrión en sus etapas iniciales. La función normal de los genes homeóticos consiste en conferir a la célula especialización espacial o posicional inequívoca en diferentes regiones a lo largo del eje anteroposterior del cuerpo. Estos genes indican a la célula si forma parte de la cabeza, del tórax o del abdomen del individuo. En el caso de la mosca de Drosophila melanogaster, en primer lugar, se expresan los genes de polaridad del huevo, que determinan los ejes anteroposterior y dorsoventral del huevo; después lo hacen los genes de segmentación, que determinan el número y la organización de los segmentos del cuerpo.
El término homeótico viene del griego homeo, que significa semejante. Los genes homeóticos reciben este nombre porque tras sufrir una mutación adquieren la capacidad de transformar un segmento de su cuerpo en otro. Ejemplos típicos en D. melanogaster son las mutaciones Antennapedia “pata por antena” y Bithorax (dos tórax). La primera tiene como resultado la aparición de patas en lugar de las antenas; la segunda provoca la repetición del segmento del tórax, obteniendo dos tórax con sus respectivas alas y respectivos halterios. Aunque inicialmente se descubrieron en D. melanogaster, los genes homeóticos se han identificado en la mayoría de los seres vivos, incluidos los seres humanos. En éstos, al igual que en el resto de los organismos, las mutaciones que afectan a estos genes son responsables de la aparición de alteraciones en el desarrollo corporal.
23. Deriva genética
23. Deriva genética
La deriva genética (deriva alélica o efecto Sewall Wright) es el cambio en la frecuencia de una variante genética existente (alelo) en una población debido al muestreo aleatorio de organismos. Los alelos de la descendencia son una muestra de los de los padres, y el azar influye en la determinación de si un individuo determinado sobrevive y se reproduce. La frecuencia alélica de una población es la fracción de las copias de un gen que comparten una forma particular.
La deriva genética puede hacer que las variantes genéticas desaparezcan por completo y, por lo tanto, reducir la variación genética sin tener en cuenta las consecuencias en la aptitud que puedan tener estas variantes, pues la deriva genética funciona al azar. También puede hacer que los alelos inicialmente raros se vuelvan mucho más frecuentes e incluso fijos, y nuevamente sin tener en cuenta que estos alelos, consecuentemente, generen problemas al individuo.
Casi todos los fenómenos se ven afectados simultáneamente por factores aleatorios (impredecibles) y no aleatorios, o determinísticos (predecibles). Cualquiera de nosotros puede sufrir un accidente automovilístico debido al comportamiento impredecible de otros conductores, pero es previsible que tengamos más probabilidades de hacerlo si lo hacemos después de beber. Lo mismo ocurre con la evolución. La selección natural es un proceso determinista, no aleatorio. Pero, al mismo tiempo, hay importantes procesos aleatorios en la evolución, incluida la mutación y las fluctuaciones aleatorias en las frecuencias de los alelos o haplotipos llamada deriva genética aleatoria (D. J. Futuyma, 2005).
La deriva genética y la selección natural son las dos causas más importantes de sustitución de alelos, es decir, del cambio evolutivo en las poblaciones. La deriva genética ocurre en todas las poblaciones naturales porque, al igual que las poblaciones ideales en el Equilibrio de Hardy-Weinberg, las poblaciones naturales son de tamaño finito. Las fluctuaciones aleatorias en las frecuencias de los alelos pueden resultar en el reemplazo de alelos viejos por otros nuevos, dando como resultado una evolución no adaptativa. Es decir, "si bien la selección natural da como resultado la adaptación, la deriva genética no lo hace, por lo que este proceso no es responsable de esas características anatómicas, fisiológicas y de comportamiento de los organismos que los equipan para la reproducción y la supervivencia. Sin embargo, la deriva genética tiene muchos aspectos importantes". consecuencias, especialmente a nivel genético molecular: parece explicar gran parte de la diferencia en las secuencias de ADN entre especies.
Debido a que todas las poblaciones son finitas, los alelos de todos los loci están potencialmente sujetos a una deriva genética aleatoria, pero no todos están necesariamente sujetos a la selección natural. Por esta razón, y debido a que los efectos esperados de la deriva genética pueden describirse matemáticamente con cierta precisión, algunos genetistas evolucionistas sostienen la opinión de que la deriva genética debería ser la "hipótesis nula" utilizada para explicar una observación evolutiva a menos que haya evidencia positiva de selección natural o algún otro factor. Esta perspectiva es análoga a la "hipótesis nula" en estadística: la hipótesis de que los datos no se apartan de los esperados basándose únicamente en el azar ". Según este punto de vista, no deberíamos asumir que una característica o una diferencia entre poblaciones o especie, es adaptativa o ha evolucionado por selección natural a menos que exista evidencia para esta conclusión.

Figura 23.1. Motoo Kimura (13 de noviembre de 1924 - 13 de noviembre de 1994) fue un biólogo japonés mejor conocido por introducir la teoría neutral de la evolución molecular en 1968 (Kimura, 1968). Se convirtió en uno de los genetistas teóricos de poblaciones más influyentes. Es recordado en genética por su uso innovador de ecuaciones de difusión para calcular la probabilidad de fijación de alelos benéficos, perjudiciales o neutrales (Watterson, 1996). Combinando la genética de poblaciones teóricas con datos de evolución molecular, también desarrolló la teoría neutral de la evolución molecular en la que la deriva genética es la fuerza principal que cambia las frecuencias alélicas (Ohta & Gillespie, 1996).
La teoría de la deriva genética, gran parte de la cual fue desarrollada por el genetista estadounidense Sewall Wright a partir de la década de 1930, y por el genetista japonés Motoo Kimura a partir de la década de 1950, incluye algunos de los modelos matemáticos más refinados en biología.
23.1 Equilibrios con la selección natural
En las poblaciones naturales, la deriva genética y la selección natural no actúan de forma aislada; ambos fenómenos están siempre en juego, junto con la mutación y la migración. La evolución neutral es el producto tanto de la mutación como de la deriva. De manera similar, incluso cuando la selección supera la deriva genética, solo puede actuar sobre la variación que proporciona la mutación.
Mientras que la selección natural tiene una dirección, guiando la evolución hacia adaptaciones heredables que responden a presiones de supervivencia, la deriva genética no tiene dirección y está guiada solo por las matemáticas del azar (Futuyma, 2004). Como resultado, la deriva actúa sobre las frecuencias genotípicas dentro de una población sin tener en cuenta sus efectos fenotípicos. Por el contrario, la selección favorece la propagación de alelos cuyos efectos fenotípicos aumentan la supervivencia y / o reproducción de sus portadores, disminuye las frecuencias de los alelos que causan rasgos desfavorables e ignora los que son neutrales (Cavalli-Sforza, Cavalli-Sforza, Menozzi, & Piazza, 1994).
23.2 Ley de los números grandes
La ley de los grandes números predice que cuando el número absoluto de copias del alelo es pequeño (por ejemplo, en poblaciones pequeñas en peligro de extinción), la magnitud de la deriva en las frecuencias alélicas por generación es mayor.

Figura 23.2. Recuerde que una población natural vieja y bien adaptada es por naturaleza, altamente diversa.
La magnitud de la deriva es lo suficientemente grande como para abrumar la selección en cualquier frecuencia alélica cuando el coeficiente de selección es menor que 1 dividido por el tamaño efectivo de la población. Por lo tanto, la evolución no adaptativa resultante del producto de la mutación y la deriva genética se considera un mecanismo consecuente de cambio evolutivo principalmente dentro de poblaciones pequeñas y aisladas (Zimmer, 2011).
Aunque estos razonamientos pueden parecer bastante abstractos en la realidad, traen consecuencias muy fuertes en cuanto a la hipótesis de diseño. Normalmente los filósofos antiguos se concentraron mucho en los ejemplos de organismos que se encuentran finamente adaptados a su ambiente, ignorando repetidamente el hecho de que la mayoría de los individuos muere y además no conocían el otro hecho importante de que la mayoría de las especies se extinguen. La mayoría de las especies, por lo tanto, son un fracaso a largo plazo y por ende, la hipótesis de diseño no se sostiene, pues un diseñador inteligente va a diseñar a sus organismos de forma tal que no se extingan. Por el contrario, la falta de previsión de los modelos naturalistas como el de deriva genética, otorgan un poder explicativo de por qué poblaciones, específicamente poblaciones aisladas en islas, se extinguen o son ineptas para competir con especies introducidas del continente.
Cuando la frecuencia alélica es muy pequeña, la deriva también puede dominar la selección incluso en poblaciones grandes. Por ejemplo, mientras que las mutaciones desventajosas en general se eliminan rápidamente en grandes poblaciones, las nuevas mutaciones ventajosas son casi tan vulnerables a la pérdida por deriva genética como las mutaciones neutrales. Hasta que la frecuencia alélica de la mutación ventajosa no alcance un cierto umbral, la deriva genética no tendrá ningún efecto.
23.3 Cuellos de botella
Un cuello de botella poblacional ocurre cuando una población se contrae a un tamaño significativamente menor durante un corto período de tiempo debido a algún evento ambiental aleatorio. En un verdadero cuello de botella poblacional, las probabilidades de supervivencia de cualquier miembro de la población son puramente aleatorias y no mejoran con ninguna ventaja genética inherente en particular. El cuello de botella puede resultar en cambios radicales en las frecuencias alélicas, completamente independientes de la selección (Ørsted, Hoffmann, Sverrisdóttir, Nielsen, & Kristensen, 2019; Wein & Dagan, 2019).
El impacto de un cuello de botella en la población puede mantenerse, incluso cuando el cuello de botella sea causado por un evento único, como una catástrofe natural. Un ejemplo interesante de un cuello de botella que causa una distribución genética inusual es la proporción relativamente alta de individuos con ceguera total al color de las células bastón (acromatopsia) en el atolón Pingelap en Micronesia (Spaethe, Schmidt, Hickelsberger, & Chittka, 2001). Después de un cuello de botella, aumenta la consanguinidad. Esto aumenta el daño causado por mutaciones deletéreas recesivas, en un proceso conocido como depresión endogámica (López-Wilchis et al., 2021; Rojas-Bracho & Jaramillo-Legoretta, 2009). Se seleccionan mutaciones al azar, y con azar es más probable que se dispersen mutaciones deletéreas. Esto conduce a una mayor pérdida de diversidad genética. Además, una reducción sostenida del tamaño de la población aumenta la probabilidad de que se produzcan más fluctuaciones alélicas derivadas de la deriva en las generaciones venideras, por lo que la población tiende a escaparse de la capacidad adaptativa de la selección natural, disminuyendo su potencial competitivo contra otras especies semejantes.

Figura 23.3. El modelo anterior representa de izquierda a derecha: una población vieja y diversa, que al experimentar un evento de deriva reduce su diversidad genética con la que restituye una población grande, pero menos diversa.
La variación genética de una población puede reducirse en gran medida por un cuello de botella, e incluso las adaptaciones beneficiosas pueden eliminarse permanentemente (Futuyma, 2005). La pérdida de variación deja a la población superviviente vulnerable a nuevas presiones de selección, como enfermedades, cambios climáticos o cambios en la fuente de alimento disponible, porque adaptarse en respuesta a los cambios ambientales requiere una variación genética suficiente en la población para que tenga lugar la selección natural (Cornuet & Luikart, 1996; O’Corry-Crowe, 2008).
Ha habido muchos casos conocidos de cuellos de botella demográficos en el pasado reciente. Antes de la llegada de los europeos, las praderas de América del Norte eran el hábitat de millones de pollos de las praderas más grandes. Solo en Illinois, su número se desplomó de aproximadamente 100 millones de aves en 1900 a aproximadamente 50 aves en la década de 1990. La disminución de la población se debió a la caza y la destrucción del hábitat, pero una consecuencia ha sido la pérdida de la mayor parte de la diversidad genética de la especie. El análisis de ADN que compara las aves de mediados de siglo con las aves de la década de 1990 documenta una fuerte disminución en la variación genética solo en las últimas décadas. Actualmente, el gran pollo de las praderas está experimentando un escaso éxito reproductivo (Kaiser, 1997).
Sin embargo, la pérdida genética causada por el cuello de botella y la deriva genética puede aumentar la aptitud, como en las bacterias Ehrlichia (Dale & Moran, 2006).
La caza excesiva también provocó un grave cuello de botella en la población del elefante marino del norte en el siglo XIX. La disminución resultante en la variación genética se puede deducir comparándola con la del elefante marino del sur, que no fue cazado de manera tan agresiva (Kalkvik, Stout, & Parkinson, 2012).
23.4 Efecto fundador
El efecto fundador es un caso especial de cuello de botella poblacional, que ocurre cuando un pequeño grupo de una población se separa de la población original y forma una nueva. Se espera que la muestra aleatoria de alelos en la nueva colonia recién formada represente erróneamente a la población original en al menos algunos aspectos. Incluso es posible que el número de alelos para algunos genes en la población original sea mayor que el número de copias de genes en los fundadores, haciendo imposible la representación completa. Cuando una colonia recién formada es pequeña, sus fundadores pueden afectar fuertemente la composición genética de la población en el futuro.
Un ejemplo bien documentado se encuentra en la migración de los Amish a Pensilvania en 1744. Dos miembros de la nueva colonia compartían el alelo recesivo del síndrome de Ellis-Van Creveld. Los miembros de la colonia y sus descendientes tienden a ser aislados religiosos y permanecen relativamente aislados reproductivamente. Como resultado de muchas generaciones de endogamia, el síndrome de Ellis-Van Creveld es ahora mucho más frecuente entre los Amish que entre la población general (Liping Hou et al., 2013; Shook, Nelson, Aguilera, & Braff, 2019).
La diferencia en las frecuencias de genes entre la población original y la colonia también puede provocar que los dos grupos diverjan significativamente en el transcurso de muchas generaciones. A medida que aumenta la diferencia, o la distancia genética, las dos poblaciones separadas pueden volverse distintas, tanto genética como fenéticamente, aunque no solo la deriva genética, sino también la selección natural, el flujo de genes y la mutación contribuyen a esta divergencia. Este potencial de cambios relativamente rápidos en la frecuencia genética de la colonia llevó a la mayoría de los científicos a considerar el efecto fundador (y por extensión, la deriva genética) como una fuerza impulsora significativa en la evolución de nuevas especies. Sewall Wright fue el primero en atribuir este significado a la deriva aleatoria y a las poblaciones pequeñas recién aisladas con su teoría del equilibrio cambiante de la especiación (Wolf, Brodie, & Wade, 2000).
24. Aplicaciones de la teoría evolutiva
24. Aplicaciones de la teoría evolutiva
La biología evolutiva, en particular la comprensión de cómo los organismos evolucionan a través de la selección natural, es un área de la ciencia con muchas aplicaciones prácticas (Bull & Wichman, 2001; Lindberg, 2007), las cuales podemos dividir en tres categorías, selección artificial e ingeniería genética, medicina y biología computacional.
24.1 Selección artificial
La cría selectiva (también llamada selección artificial) es el proceso mediante el cual los seres humanos utilizan la cría de animales y la cría de plantas para desarrollar selectivamente rasgos (características) fenotípicos particulares al elegir qué machos y hembras típicamente animales o vegetales se reproducirán sexualmente y tendrán descendencia juntos. Los animales domesticados se conocen como razas, normalmente criados por un criador profesional, mientras que las plantas domesticadas se conocen como variedades, cultígenos, cultivares o razas. Dos animales de raza pura de diferentes razas producen un cruce, y las plantas cruzadas se denominan híbridos. Las flores, hortalizas y árboles frutales pueden ser cultivados por aficionados y profesionales comerciales o no comerciales: los cultivos principales suelen ser de procedencia de los profesionales.
En la cría de animales, se utilizan técnicas como la consanguinidad, la cría en línea y el cruzamiento. En el fitomejoramiento se utilizan métodos similares. Charles Darwin discutió cómo la cría selectiva había tenido éxito en producir cambios a lo largo del tiempo en su libro de 1859, Sobre el origen de las especies. Su primer capítulo analiza la cría selectiva y la domesticación de animales como palomas, gatos, ganado y perros. Darwin usó la selección artificial como trampolín para introducir y apoyar la teoría de la selección natural (Darwin, 1866).
La explotación deliberada de la cría selectiva para producir los resultados deseados se ha vuelto muy común en la agricultura y la biología experimental.
La cría selectiva puede ser involuntaria, por ejemplo, como resultado del proceso de cultivo humano; y también puede producir resultados no deseados, deseables o indeseables. Por ejemplo, en algunos granos, un aumento en el tamaño de la semilla puede deberse a ciertas prácticas de arado más que a la selección intencional de semillas más grandes. Lo más probable es que haya habido una interdependencia entre factores naturales y artificiales que han dado lugar a la domesticación de las plantas (Purugganan & Fuller, 2009).
Más recientemente, dicha selección se ha convertido en una parte vital de la ingeniería genética, con marcadores seleccionables como genes de resistencia a antibióticos que se utilizan para manipular el ADN en biología molecular. También es posible utilizar rondas repetidas de mutación y selección para desarrollar proteínas con propiedades particulares, como enzimas modificadas o nuevos anticuerpos, en un proceso llamado evolución dirigida (Bunzel, Anderson, & Mulholland, 2021; Jäckel, Kast, & Hilvert, 2008; Zeymer & Hilvert, 2018).
24.2 Medicina
La resistencia a los antibióticos puede ser el resultado de mutaciones puntuales en el genoma del patógeno a una tasa de aproximadamente 1 en 108 por replicación cromosómica. La acción antibiótica contra el patógeno puede verse como una presión ambiental; las bacterias que tienen una mutación que les permite sobrevivir vivirán para reproducirse. Luego pasarán este rasgo a su descendencia, lo que dará como resultado una colonia completamente resistente.
La comprensión de los cambios que han ocurrido durante la evolución del organismo puede revelar los genes necesarios para construir partes del cuerpo, genes que pueden estar involucrados en los trastornos genéticos humanos (Maher, 2009).

Figura 24.1. Astyanax mexicanus es una especie de peces de la familia Characidae en el orden de los Characiformes, que han perdido sus ojos durante el curso de su historia evolutiva.
Por ejemplo, el tetra mexicano “Astyanax mexicanus” es un pez de las cavernas albino que perdió la vista durante la evolución. La cría de diferentes poblaciones de este pez ciego produjo algunas crías con ojos funcionales, ya que se habían producido diferentes mutaciones en las poblaciones aisladas que habían evolucionado en diferentes cuevas (Borowsky, 2008). Esto ayudó a identificar los genes necesarios para la visión y la pigmentación, como las cristalinas y el receptor de melanocortina 1 (Gross, Borowsky, & Tabin, 2009). De manera similar, al comparar el genoma del draco antártico (Channichthyidae spp), que carece de glóbulos rojos, con parientes cercanos como el cocodrilo antártico (Trematomus bernacchii), se revelaron los genes necesarios para producir estas células sanguíneas (Yergeau, Cornell, Parker, Zhou, & Detrich III, 2005).
24.3 Computación evolutiva
En informática, la computación evolutiva es una familia de algoritmos para la optimización global inspirados en la evolución biológica, y el subcampo de la inteligencia artificial y la computación blanda que estudian estos algoritmos. En términos técnicos, son una familia de solucionadores de problemas de prueba y error basados en la población con un carácter de optimización metaheurística o estocástica.
En la computación evolutiva, se genera y actualiza iterativamente un conjunto inicial de soluciones candidatas. Cada nueva generación se produce eliminando estocásticamente las soluciones menos deseadas e introduciendo pequeños cambios aleatorios. En terminología biológica, una población de soluciones está sujeta a selección natural (o selección artificial) y mutación aleatoria. Como resultado, la población evolucionará gradualmente para aumentar su aptitud, en este caso la función de aptitud elegida del algoritmo, sin que el ingeniero deba preocuparse por diseñar variantes manualmente.
Las técnicas de computación evolutiva pueden producir soluciones altamente optimizadas en una amplia gama de entornos de problemas, lo que las hace populares en la informática. Existen muchas variantes y extensiones, adecuadas para familias más específicas de problemas y estructuras de datos. La computación evolutiva también se usa a veces en biología evolutiva como un procedimiento experimental in silico para estudiar aspectos comunes de los procesos evolutivos generales.
👉 Historia
El uso de principios evolutivos para la resolución automatizada de problemas se originó en la década de 1950. No fue hasta la década de 1960 que se empezaron a desarrollar tres interpretaciones distintas de esta idea en tres lugares diferentes.
La programación evolutiva fue introducida por Lawrence J. Fogel en los Estados Unidos, mientras que John Henry Holland llamó a su método un algoritmo genético. En Alemania, Ingo Rechenberg y Hans-Paul Schwefel introdujeron estrategias de evolución. Estas áreas se desarrollaron por separado durante unos 15 años. Desde principios de los noventa en adelante se unifican como diferentes representantes ("dialectos") de una tecnología, denominada computación evolutiva. También a principios de los noventa, había surgido una cuarta corriente que seguía las ideas generales: la programación genética. Desde la década de 1990, los algoritmos inspirados en la naturaleza se están convirtiendo en una parte cada vez más importante de la computación evolutiva.
Estas terminologías denotan el campo de la computación evolutiva y consideran la programación evolutiva, las estrategias de evolución, los algoritmos y la programación genéticos como subáreas.
Las primeras simulaciones computacionales de la evolución utilizando algoritmos evolutivos y técnicas de vida artificial fueron realizadas por Nils Aall Barricelli en 1953 (Dorin & Taylor, 2020), con los primeros resultados publicados en 1954 (Barricelli, 1954). Otro pionero en la década de 1950 fue Alex Fraser, quien publicó una serie de artículos sobre simulación de selección artificial (Fraser, 1958). La evolución artificial se convirtió en un método de optimización ampliamente reconocido como resultado del trabajo de Ingo Rechenberg en la década de 1960 y principios de la de 1970, quien utilizó estrategias de evolución para resolver problemas complejos de ingeniería (unter Rudolph & Schwefel, 1994). Los algoritmos genéticos en particular se hicieron populares a través de la escritura de John Holland (Holland, 1992). A medida que crecía el interés académico, los aumentos dramáticos en el poder de las computadoras permitieron aplicaciones prácticas, incluida la evolución automática de los programas de computadora (Koza, 1994). Los algoritmos evolutivos se utilizan ahora para resolver problemas multidimensionales de manera más eficiente que el software producido por diseñadores humanos, y también para optimizar el diseño de sistemas (Jamshidi, 2003; Onwubolu & Babu, 2013).
👉 Algoritmos evolutivos
Los algoritmos evolutivos forman un subconjunto de la computación evolutiva en el sentido de que generalmente solo involucran técnicas que implementan mecanismos inspirados en la evolución biológica como la reproducción, mutación, recombinación, selección natural y supervivencia del más apto. Las soluciones candidatas al problema de optimización desempeñan el papel de los individuos en una población, y la función de costes determina el entorno en el que "viven" las soluciones. La evolución de la población tiene lugar luego de la aplicación repetida de los operadores anteriores.
En este proceso, hay dos fuerzas principales que forman la base de los sistemas evolutivos: la mutación de recombinación y el cruce crean la diversidad necesaria y, por lo tanto, facilitan la novedad, mientras que la selección actúa como una fuerza que aumenta la calidad.
Muchos aspectos de este proceso evolutivo son estocásticos. Las piezas de información modificadas debido a la recombinación y la mutación se eligen al azar. Por otro lado, los operadores de selección pueden ser deterministas o estocásticos. En el último caso, los individuos con una mayor aptitud tienen una mayor probabilidad de ser seleccionados que los individuos con una menor aptitud, pero normalmente incluso los individuos débiles tienen la posibilidad de convertirse en padres o de sobrevivir.
👉 Algoritmos evolutivos y biología
Los algoritmos genéticos ofrecen métodos para modelar sistemas biológicos y biología de sistemas que están vinculados a la teoría de sistemas dinámicos, ya que se utilizan para predecir los estados futuros del sistema. Esta es solo una forma vívida (pero quizás engañosa) de llamar la atención sobre el carácter ordenado, bien controlado y altamente estructurado del desarrollo en biología.
Sin embargo, el uso de algoritmos e informática, en particular de la teoría computacional, más allá de la analogía con los sistemas dinámicos, también es relevante para entender la evolución misma.
Esta visión tiene el mérito de reconocer que no existe un control central del desarrollo; los organismos se desarrollan como resultado de interacciones locales dentro y entre las células. Nos parece que las ideas más prometedoras sobre los paralelos de desarrollo de programas son las que apuntan a una analogía aparentemente cercana entre los procesos dentro de las células y el funcionamiento de bajo nivel de las computadoras modernas. Por lo tanto, los sistemas biológicos son como máquinas computacionales que procesan información de entrada para calcular los siguientes estados, de modo que los sistemas biológicos están más cerca de un cálculo que el sistema dinámico clásico (Ochoa, 2018).
Además, siguiendo conceptos de la teoría computacional, los microprocesos en los organismos biológicos son fundamentalmente incompletos e indecidibles (completitud (lógica)), lo que implica que “hay más que una burda metáfora detrás de la analogía entre células y computadoras (Danchin, 2008).
La analogía con la computación se extiende también a la relación entre los sistemas de herencia y la estructura biológica, que a menudo se piensa que revela uno de los problemas más urgentes para explicar los orígenes de la vida.
Los autómatas evolutivos (Burgin & Eberbach, 2010, 2012, 2013), una generalización de las máquinas evolutivas de Turing (Eberbach, 2002, 2005), se han introducido con el fin de investigar con mayor precisión las propiedades de la computación biológica y evolutiva. En particular, permiten obtener nuevos resultados sobre la expresividad de la computación evolutiva (Eberbach & Burgin, 2009).
25. Debates sociales sobre la evolución
25. Debates sociales sobre la evolución
En el siglo XIX, particularmente después de la publicación de El origen de las especies en 1859, la idea de que la vida había evolucionado fue una fuente activa de debate académico centrado en las implicaciones filosóficas, sociales y religiosas de la evolución. Hoy en día, la síntesis evolutiva moderna es aceptada por una gran mayoría de científicos (Müller, 2017). Sin embargo, la evolución sigue siendo un concepto polémico para algunos teístas (Zuckerkandl, 2006).
25.1 Debates religiosos
Si bien varias religiones y denominaciones han reconciliado sus creencias con la evolución a través de conceptos como la evolución teísta, hay creacionistas que creen que la evolución se contradice con los mitos de la creación que se encuentran en sus religiones y que plantean diversas objeciones a la evolución (Hameed, 2008; Ross, 2005; Scott & Matzke, 2007). Como lo demostraron las respuestas a la publicación de Vestigios de la historia natural de la creación en 1844, el aspecto más controvertido de la biología evolutiva es la implicación de la evolución humana de que los humanos comparten un ancestro común con los simios y que las facultades mentales y morales de la humanidad tienen los mismos tipos de causas naturales que otros rasgos hereditarios en los animales (Bowler, 1989).
En algunos países, especialmente en los Estados Unidos, estas tensiones entre ciencia y religión han alimentado la actual controversia creación-evolución, un conflicto religioso centrado en la política y la educación pública (J. D. Miller, Scott, & Okamoto, 2006). Mientras que otros campos científicos como la cosmología (Spergel et al., 2003) y las ciencias de la Tierra (Wilde, Valley, Peck, & Graham, 2001) también entran en conflicto con las interpretaciones literales de muchos textos religiosos, la biología evolutiva experimenta una oposición significativamente mayor por parte de los literalistas religiosos.
25.2 La enseñanza de la evolución en la escuela
La enseñanza de la evolución en las clases de biología de las escuelas secundarias estadounidenses fue poco común en la mayor parte de la primera mitad del siglo XX. La decisión de del juicio de Scopes de 1925 hizo que el tema se volviera muy raro en los libros de texto de biología secundaria estadounidenses durante una generación, pero se volvió a introducir gradualmente más tarde y se protegió legalmente con la decisión de Epperson v. Arkansas de 1968. Desde entonces, la creencia religiosa competitiva del creacionismo fue legalmente prohibida en los planes de estudio de la escuela secundaria en varias decisiones en las décadas de 1970 y 1980, pero regresó en forma pseudocientífica como diseño inteligente (DI), para ser excluida una vez más en el 2005 Kitzmiller v. Dover (Branch, 2007). El debate sobre las ideas de Darwin no generó una controversia significativa en China (Jin, 2019).
25.3 La situación en Colombia
No tengo conocimiento de algún conflicto específico reportado en Colombia en relación a la enseñanza de la síntesis evolutiva moderna o del darwinismo en escuelas o universidades. Sin embargo, es posible que haya habido debates y discusiones sobre estos temas en algunos círculos académicos y científicos, ya que en ocasiones hay diferentes interpretaciones o enfoques en la teoría evolutiva.
Es importante tener en cuenta que la enseñanza de la evolución es un tema sensible en algunos lugares del mundo, incluyendo América Latina. En algunos casos, puede haber resistencia o controversia por parte de grupos religiosos o conservadores que rechazan la teoría evolutiva por motivos ideológicos o culturales. Sin embargo, en general, la síntesis evolutiva moderna es ampliamente aceptada como la explicación científica más sólida y ampliamente respaldada sobre el origen y diversidad de las especies en la actualidad (Restrepo Forero, 2009).
25.4 La situación en América latina
Sí, existen reportes de conflictos y controversias relacionados con la enseñanza de la evolución en algunos países de América Latina, por ejemplo, en un artículo del periódico La Jornada de México, publicado en 2013, se titula "Enseñanza de la evolución, un desafío en México", y trata sobre los desafíos y obstáculos que enfrenta la enseñanza de la evolución en las escuelas mexicanas. El artículo señala que la evolución es un tema polémico en México debido a la influencia de grupos religiosos que rechazan la teoría de la evolución y proponen alternativas como el creacionismo o el diseño inteligente.
El artículo cita a diversos expertos y académicos que señalan la importancia de enseñar la evolución de manera rigurosa y científica en las escuelas para fomentar el pensamiento crítico y la comprensión del mundo natural. También se mencionan las dificultades que enfrentan los maestros para enseñar la evolución debido a la falta de capacitación y recursos.
El artículo concluye con una reflexión sobre la necesidad de promover el diálogo y la colaboración entre los diferentes actores involucrados en la enseñanza de la evolución en México, incluyendo a los científicos, maestros, estudiantes y grupos religiosos, para avanzar en la comprensión y aceptación de esta importante teoría científica.
25.5 La situación en la unión europea
La enseñanza de la evolución en la Unión Europea se rige por las políticas y regulaciones de cada país miembro, aunque existe un consenso generalizado en la comunidad científica europea sobre la importancia de enseñar la evolución de manera rigurosa y científica.
En 2007, la Comisión Europea publicó un informe titulado "La enseñanza de las ciencias en la Unión Europea" (EURYDICE., Educación, Cultura, & Deporte, 2012), que destaca la importancia de la enseñanza de la evolución en el contexto de la educación científica. En este informe se señala que la evolución es una teoría científica bien establecida y que su enseñanza es esencial para fomentar el pensamiento crítico y la comprensión del mundo natural.
Sin embargo, en algunos países de la Unión Europea, como España, Italia y Polonia, se han registrado casos de conflictos y controversias en torno a la enseñanza de la evolución. En estos países, existen grupos religiosos y políticos que rechazan la evolución y promueven alternativas como el creacionismo o el diseño inteligente.
En general, la Unión Europea promueve la enseñanza de la evolución como parte de la educación científica y apoya la investigación y difusión de la ciencia evolutiva (Peker, Comert, & Kence, 2010; Savvides, 2003; Xanthoudaki, 2010).
25.6 La situación en rusia
En general, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo en Rusia sigue las mismas líneas generales que en otros países. La teoría evolutiva se enseña en las escuelas y universidades como una parte integral de la biología y otros campos científicos.
Sin embargo, es importante tener en cuenta que Rusia es un país grande y diverso, con una gran cantidad de opiniones y actitudes diferentes hacia la ciencia y la educación. En algunos casos, puede haber resistencia a la enseñanza de la evolución y la biología en general, particularmente en áreas rurales y más conservadoras.
En cuanto a la síntesis evolutiva moderna, hay una comunidad activa de biólogos y genetistas en Rusia que trabajan en esta área, y las investigaciones y publicaciones sobre el tema son abundantes. Además, hay varias universidades y centros de investigación en Rusia que se enfocan en la evolución y la genética, y que ofrecen cursos y programas en estas áreas.
En resumen, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo en Rusia sigue las mismas líneas generales que en otros países, aunque puede haber diferencias regionales en la aceptación y comprensión de estas teorías (Menter, 2021).
25.7 La situación en Japón y Corea del sur
En general, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo en Japón y Corea del Sur sigue las mismas líneas generales que en otros países desarrollados. La teoría evolutiva se enseña en las escuelas y universidades como una parte integral de la biología y otros campos científicos.
En Japón, la evolución y la síntesis evolutiva moderna se enseñan en la educación secundaria y superior, y los libros de texto de biología utilizados en las escuelas japonesas presentan la evolución de manera detallada. También existen instituciones de investigación y universidades que tienen un fuerte enfoque en la investigación y la enseñanza de la evolución y la biología en general (Unoura, 1999).
En Corea del Sur, la evolución también se enseña en las escuelas secundarias y en las universidades como una parte importante de la biología. Sin embargo, a veces hay controversia sobre la enseñanza de la evolución en algunas comunidades conservadoras o religiosas. En 2012, hubo protestas contra el Ministerio de Educación en Corea del Sur por incluir la evolución en el plan de estudios nacional, pero la evolución sigue siendo una parte importante de la educación científica en el país (Kim & Nehm, 2011; Park, 2022).
En resumen, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo en Japón y Corea del Sur es similar a la de otros países desarrollados. La evolución se enseña ampliamente en las escuelas y universidades, aunque puede haber controversias ocasionales en algunas comunidades conservadoras o religiosas.
25.8 La situación en China
En China, la enseñanza de la evolución y la síntesis evolutiva moderna está presente en los planes de estudio de biología en la educación secundaria y superior. La teoría evolutiva se enseña como parte de la biología, y se considera un tema importante en la educación científica en China. Los libros de texto de biología utilizados en las escuelas chinas presentan la evolución como una teoría científica aceptada y fundamental.
En la esfera de influencia del sur de China, que incluye a países como Vietnam, Laos, Camboya, Tailandia y Myanmar, la situación es similar. En estos países, la evolución se enseña en las escuelas y universidades como parte de la biología y otros campos científicos. Sin embargo, en algunos casos, las creencias religiosas y culturales pueden influir en la aceptación de la teoría evolutiva en estas regiones.
Aunque la enseñanza de la evolución está presente en la educación en China y su esfera de influencia del sur, hay cierta controversia sobre la enseñanza de la teoría evolutiva en China. Algunos críticos afirman que la enseñanza de la evolución puede ser insuficiente o presentada de manera insuficientemente crítica en las escuelas chinas. Sin embargo, la evolución sigue siendo una parte importante de la educación científica en China y su esfera de influencia del sur.
En resumen, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo en China y su esfera de influencia del sur sigue las mismas líneas generales que en otros países desarrollados. La teoría evolutiva se enseña en las escuelas y universidades como parte de la biología y otros campos científicos, aunque puede haber controversias ocasionales (M. Zhang, Weisberg, Zhu, & Weisberg, 2022).
Cabe destacar que la situación puede variar en diferentes regiones de China y de su esfera de influencia del sur, y que puede ser influenciada por factores culturales, religiosos y políticos.
25.9 La situación es los Estados Unidos de América
En los Estados Unidos, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo es parte del plan de estudios de biología en la mayoría de las escuelas secundarias y universidades. La evolución es considerada una teoría científica establecida y fundamental en la educación científica de los Estados Unidos. Sin embargo, la enseñanza de la evolución a menudo se enfrenta a oposición de grupos religiosos y políticos que abogan por la enseñanza del creacionismo o el diseño inteligente en las escuelas.
Los estados del sur de los Estados Unidos, que incluye estados como Texas, Alabama, Louisiana, Mississippi y otros, la enseñanza de la evolución a menudo enfrenta una mayor oposición y crítica por parte de grupos religiosos y políticos conservadores. En algunos estados, ha habido intentos de introducir leyes o políticas que permitan o incluso requieran la enseñanza del creacionismo o el diseño inteligente en las escuelas públicas.
A pesar de estos desafíos, la enseñanza de la evolución sigue siendo una parte importante de la educación científica en los Estados Unidos y su esfera de influencia del sur. Los defensores de la educación en evolución trabajan activamente para promover la enseñanza de la evolución y contrarrestar los intentos de socavarla (Laats & Siegel, 2016; Lynn, Glaze, Evans, & Reed, 2017; Plutzer, Branch, & Reid, 2020).
En resumen, la enseñanza de la síntesis evolutiva moderna y otras formas de darwinismo en los Estados Unidos sigue siendo un tema polémico, pero la evolución se enseña ampliamente como parte de la educación científica en la mayoría de las escuelas y universidades. La situación puede variar de un estado a otro y puede verse influida por factores políticos, culturales y religiosos.
25.10 La situación en el mundo musulmán
En el mundo musulmán, la enseñanza de la evolución y la aceptación de la síntesis evolutiva moderna varían en diferentes países y comunidades. Algunos países musulmanes han adoptado la evolución como una teoría científica establecida y la enseñan ampliamente en sus sistemas educativos, mientras que otros países musulmanes han sido más reticentes a adoptarla.
En algunos países musulmanes, la evolución se enseña en las escuelas y universidades sin controversia significativa. Por ejemplo, en Turquía, la enseñanza de la evolución es obligatoria en la educación secundaria y se enseña en la mayoría de las universidades. En Marruecos, la enseñanza de la evolución también es parte del plan de estudios de biología en la educación secundaria y universitaria.
Sin embargo, en algunos otros países musulmanes, la enseñanza de la evolución enfrenta desafíos significativos. En algunos países, las creencias religiosas se ven como incompatibles con la teoría de la evolución, y algunos grupos religiosos han expresado su oposición a la enseñanza de la evolución. En algunos casos, se ha intentado prohibir o restringir la enseñanza de la evolución en las escuelas (Asghar, 2013; Asghar, Hameed, & Farahani, 2014; Danner, 1991).
En general, la actitud hacia la enseñanza de la evolución y la aceptación de la síntesis evolutiva moderna en el mundo musulmán varía ampliamente. La actitud hacia la evolución a menudo está influenciada por factores religiosos y culturales, así como por la educación científica y la comprensión de la teoría de la evolución en sí.
26. Una síntesis evolutiva más moderna
26. Una síntesis evolutiva más moderna
La síntesis evolutiva extendida consiste en un conjunto de conceptos teóricos que se argumenta que son más completos que la síntesis moderna anterior de la biología evolutiva que tuvo lugar entre 1918 y 1942. La síntesis evolutiva extendida fue solicitada en la década de 1950 por CH Waddington, argumentado en el base del equilibrio puntuado por Stephen Jay Gould y Niles Eldredge en la década de 1980, y fue reconceptualizado en 2007 por Massimo Pigliucci y Gerd B. Müller.
La síntesis evolutiva ampliada revisa la importancia relativa de los diferentes factores en juego, examinando varios supuestos de la síntesis anterior y ampliándolos con factores causales adicionales pero igual de naturalistas (Laland et al., 2015; Wade, 2011). Incluye selección multinivel, herencia epigenética transgeneracional, construcción de nichos, capacidad de evolución y varios conceptos de la biología evolutiva del desarrollo (Manfred D Laubichler & Renn, 2015; Müller, 2007; Pigliucci & Finkelman, 2014).
No todos los biólogos han coincidido en la necesidad o el alcance de una síntesis ampliada. Muchos han colaborado en otra síntesis en biología del desarrollo evolutivo, que se concentra en la genética molecular del desarrollo y la evolución para comprender cómo la selección natural opera en los procesos de desarrollo y las homologías profundas entre organismos al nivel de genes altamente conservados.
26.1 La síntesis moderna previa
La síntesis moderna fue la síntesis ampliamente aceptada de principios del siglo XX que reconcilia la teoría de la evolución por selección natural de Charles Darwin y la teoría de la genética de Gregor Mendel en un marco matemático conjunto. Estableció la evolución como paradigma central de la biología. Las ideas del siglo XIX sobre la selección natural de Darwin y la genética mendeliana fueron unidas por investigadores que incluyeron a Thomas H. Morgan fundador de la genética moderna y Ronald Fisher, uno de los tres fundadores de la genética de poblaciones, y JBS Haldane y Sewall Wright, entre 1918 y 1932. Julian Huxley introdujo la frase "síntesis moderna" en su libro de 1942, Evolution: The Modern Synthesis (Huxley, 1942).
Sin embargo, este nombre tiene una trampa, moderno significa al modo de hoy, por ende, la síntesis moderna siempre deberá ser la forma mas actualizada del conocimiento genético-evolutivo del momento.
26.2 Primeros llamados a la modernización de la síntesis moderna
Durante la década de 1950, el biólogo inglés C. H. Waddington pidió una síntesis ampliada basada en su investigación sobre epigenética y asimilación genética (Rutherford & Henikoff, 2003; Van Speybroeck, 2002; Waddington, 1953). El zoólogo austríaco Rupert Riedl también propuso una síntesis ampliada, con el estudio de la capacidad de evolución (Marchesini & Celentano, 2021; Wagner & Laubichler, 2004).
26.3 Equilibrio puntuado
En la década de 1980, los paleontólogos estadounidenses Stephen Jay Gould y Niles Eldredge abogaron por una síntesis ampliada basada en su idea de equilibrio puntuado, el papel de la selección de especies dando forma a patrones evolutivos a gran escala y la selección natural trabajando en múltiples niveles que se extienden de genes a especies (Eldredge, 1985; Stephen J Gould, 1987; Stephen Jay Gould, 1982). El etólogo John Endler escribió un artículo en 1988 en el que analizaba los procesos de evolución que, en su opinión, habían sido descuidados (Endler & McLellan, 1988).
26.4 Biología evolutiva del desarrollo
Algunos investigadores en el campo de la biología del desarrollo evolutivo propusieron otra síntesis. Argumentan que las síntesis modernas y extendidas deberían centrarse principalmente en los genes y sugerir una integración de la embriología con la genética molecular y la evolución, con el objetivo de comprender cómo opera la selección natural en la regulación genética y las homologías profundas entre organismos a nivel de genes altamente conservados, factores de transcripción. y vías de señalización (Davidson, 2010). Por el contrario, una rama diferente de evo-devo sugiere que un enfoque de organismo (Bateson, 2005; Gilbert, Opitz, & Raff, 1996; Huneman, 2010) contribuye a la síntesis ampliada al enfatizar (entre otros) el sesgo de desarrollo (Brakefield, 2006), la capacidad de evolución (Brakefield, 2006) y la inherencia de la forma (Hall, 2012; Müller, 2021; Newman & Müller, 2006) como factores primarios en la evolución de estructuras complejas y novedades fenotípicas.
26.5 Historia reciente
La idea de una síntesis ampliada fue relanzada en 2007 por Massimo Pigliucci, y Gerd B. Müller, con un libro en 2010 titulado Evolution: The Extended Synthesis (Pigliucci & Müller, 2010), que ha servido como punto de partida para trabajar en la síntesis ampliada. Esto incluye:
👉 El papel de las configuraciones previas, las estructuras genómicas y otros rasgos del organismo en la generación de variaciones evolutivas.
👉 Cómo la creciente dimensionalidad de los paisajes de fitness afecta nuestra visión de la especiación.
👉 El papel de la selección multinivel en las principales transiciones evolutivas.
👉 Nuevos tipos de herencia, incluida la herencia cultural y epigenética.
👉 La forma en que el desarrollo del organismo y la plasticidad del desarrollo canalizan las vías evolutivas y generan una novedad fenotípica
👉 Cómo los organismos modifican los entornos a los que pertenecen a través de la construcción de nichos.
Los proponentes dicen que otros procesos como la capacidad de evolución, la plasticidad fenotípica, la evolución reticulada, la evolución sexual y la simbiogénesis han sido excluidos o no incluidos en la síntesis moderna. El objetivo de la síntesis ampliada de Piglucci y Müller es llevar la evolución más allá del enfoque genético de la genética de poblaciones para considerar enfoques más centrados en el organismo y la ecología. Muchas de estas causas se consideran actualmente secundarias en la causalidad evolutiva, y los defensores de la síntesis ampliada quieren que se consideren causas evolutivas de primera clase. El biólogo Eugene Koonin escribió en 2009 que "los nuevos desarrollos en biología evolutiva de ninguna manera deben verse como una refutación de Darwin. Al contrario, están ampliando los caminos que Darwin abrió hace 150 años y revelan la extraordinaria fertilidad de su pensamiento" (Koonin, 2009).
Referencias bibliográficas de las teorías de la evolución
Referencias bibliográficas de las teorías de la evolución
Allison, A. C. (2002). The discovery of resistance to malaria of sickle‐cell heterozygotes. Biochemistry and Molecular Biology Education, 30(5), 279–287.
Amundson, R. (2005). The changing role of the embryo in evolutionary thought: roots of evo-devo. Cambridge University Press.
Artigas, M., Martnez, R. A., Glick, T. F., & Martínez, R. A. (2006). Negotiating Darwin: the Vatican confronts evolution, 1877–1902. JHU Press.
Asghar, A. (2013). Canadian and Pakistani Muslim teachers’ perceptions of evolutionary science and evolution education. Evolution: Education and Outreach, 6(1), 10.
Asghar, A., Hameed, S., & Farahani, N. K. (2014). Evolution in biology textbooks: A comparative analysis of 5 Muslim countries. Religion & Education, 41(1), 1–15.
Asghar, A., Wiles, J. R., & Alters, B. (2010). The origin and evolution of life in Pakistani high school biology. Journal of Biological Education, 44(2), 65–71.
Ayala, F. J. (1970). Teleological explanations in evolutionary biology. Philosophy of Science, 37(1), 1–15.
Ayala, F. J., & Avise, J. C. (2014). Essential readings in evolutionary biology. JHU Press.
Ayoub, M. M. (2005). Creation or evolution? The reception of Darwinism in modern Arab thought. Science and Religion in a Post-Colonial World: Interfaith Perspectives, 173–190.
Barricelli, N. A. (1954). Esempi numerici di processi di evoluzione. Methodos, 6(21–22), 45–68.
Bateson, P. (2005). The return of the whole organism. Journal of Biosciences, 30(1), 31–39.
Bayrakdar, M. (1983). Al-Jahiz and the rise of biological evolution. The Islamic Quarterly, 27, 307–315.
Beer, G. (1992). Forging the missing link: Interdisciplinary stories. CUP Archive.
Bender, B. (2004). Evolution and" the sex problem": American narratives during the eclipse of Darwinism. Kent State University Press.
Benton, M. J. (1983). Dinosaur success in the Triassic: a noncompetitive ecological model. The Quarterly Review of Biology, 58(1), 29–55.
Berkeley, U. (n.d.). Antibiotic resistance: delaying the inevitable - Understanding Evolution. Retrieved August 10, 2017, from https://evolution.berkeley.edu/evolibrary/article/medicine_03
BioInteractive, H. (2013). The Origin of Species: Lizards in an Evolutionary Tree. Video, 17, 45.
Blinn, J. F., & Goodstein, D. (1985). El Universo Mecánico: el universo cuántico mecánico. USA: PBS.
Bock, W. J. (1994). Ernst Mayr, naturalist: His contributions to systematics and evolution. Biology and Philosophy, 9(3), 267–327.
Bock, W. J. (2005). Ernst Mayr at 100: A life inside and outside ornithology. Ornithological Monographs, 2–16.
Boersma, A. T., & Pyenson, N. D. (2015). Albicetus oxymycterus, a new generic name and redescription of a basal physeteroid (Mammalia, Cetacea) from the Miocene of California, and the evolution of body size in sperm whales. PloS One, 10(12), e0135551.
Boisvert, C. A. (2005). The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion. Nature, 438(7071), 1145–1147.
Borowsky, R. (2008). Restoring sight in blind cavefish. Current Biology, 18(1), R23–R24.
Bourlière, F., & Howell, C. F. (2013). African ecology and human evolution. Routledge.
Bowler, P. J. (1975). The changing meaning of" evolution". Journal of the History of Ideas, 95–114.
Bowler, P. J. (1979). Theodor Eimer and orthogenesis: evolution by ‘definitely directed variation.’ Journal of the History of Medicine and Allied Sciences, 34(1), 40–73.
Bowler, P. J. (1989). Evolution: the history of an idea. Univ of California Press.
Bowler, P. J. (1992a). The eclipse of Darwinism: Anti-Darwinian evolution theories in the decades around 1900. JHU Press.
Bowler, P. J. (1992b). The Fontana history of the environmental sciences. Fontana press.
Bowler, P. J. (2000). Philosophy, instinct, intuition: what motivates the scientist in Search of a theory? Biology and Philosophy, 15(1), 93–101.
Bowler, P. J. (2005). Revisiting the eclipse of Darwinism. Journal of the History of Biology, 38(1), 19–32.
Brackman, A. C. (1980). A delicate arrangement: the strange case of Charles Darwin and Alfred Russel Wallace. Crown.
Braje, T. J., & Erlandson, J. M. (2013). Human acceleration of animal and plant extinctions: A Late Pleistocene, Holocene, and Anthropocene continuum. Anthropocene, 4, 14–23.
Brakefield, P. M. (2006). Evo-devo and constraints on selection. Trends in Ecology & Evolution, 21(7), 362–368.
Branch, G. (2007). Understanding creationism after Kitzmiller. BioScience, 57(3), 278–284.
Bro‐Jørgensen, J. (2016). Our Antelope Heritage–Why the Fuss? Antelope Conservation: From Diagnosis to Action, (16), 1.
Brodie III, E. D., & Brodie Jr, E. D. (1991). Evolutionary response of predators to dangerous prey‐reduction of toxicity of newts and resistance of garter snakes in island populations. Evolution, 45(1), 221–224.
Browne, E. J. (1995). Charles Darwin: a biography (Vol. 2). Knopf.
Buiatti, M. (2005). Biologies, agricultures, biotechnologies. Tailoring Biotechnologies, 1(2), 9–30.
Bull, J. J., & Wichman, H. A. (2001). Applied evolution. Annual Review of Ecology and Systematics, 32(1), 183–217.
Bunzel, H. A., Anderson, J. L. R., & Mulholland, A. J. (2021). Designing better enzymes: insights from directed evolution. Current Opinion in Structural Biology, 67, 212–218.
Burgin, M., & Eberbach, E. (2010). Bounded and periodic evolutionary machines. In IEEE Congress on Evolutionary Computation (pp. 1–8). IEEE.
Burgin, M., & Eberbach, E. (2012). Evolutionary automata: Expressiveness and convergence of evolutionary computation. The Computer Journal, 55(9), 1023–1029.
Burgin, M., & Eberbach, E. (2013). Recursively Generated Evolutionary Turing Machines and Evolutionary Automata. In Artificial Intelligence, Evolutionary Computing and Metaheuristics (pp. 201–230). Springer.
Burkhardt, R. W. (2013). Lamarck, evolution, and the inheritance of acquired characters. Genetics, 194(4), 793–805.
Carroll, W. E. (2000). Creation. Evolution and Thomas Aquinas. REVUE DES QUESTIONS SCIENTIFIQUES-LOUVAIN AND PARIS-, 171(4), 319–348.
Cavalli-Sforza, L. L., Cavalli-Sforza, L., Menozzi, P., & Piazza, A. (1994). The history and geography of human genes. Princeton university press.
Charig, A. J. (1986). Archaeopteryx is not a forgery. Science, 232, 622–627.
Charles, J. (2008). Darwin’s Lost Voyage. USA: National Geographic. Retrieved from https://www.youtube.com/watch?v=x7kIPlgAx2I
Charlesworth, B., & Edwards, A. W. F. (2018). A century of variance. Significance, 15(4), 20–25.
Chen, J.-Y., Huang, D.-Y., & Li, C.-W. (1999). An early Cambrian craniate-like chordate. Nature, 402(6761), 518–522.
Churchill, F. B. (1970). Hertwig, Weismann, and the meaning of reduction division circa 1890. Isis, 61(4), 429–457.
Churchill, F. B. (2015). August Weismann. Harvard University Press.
Clark, B., Foster, J. B., & York, R. (2007). The critique of intelligent design: Epicurus, Marx, Darwin, and Freud and the materialist defense of science. Theory and Society, 36(6), 515–546.
Clarke, B. C. (1995). Edmund Brisco Ford, 23 April 1901-21 January 1988. The Royal Society.
Clutton-Brock, T. (2007). Sexual selection in males and females. Science, 318(5858), 1882–1885.
Coates, M. I. (1996). The Devonian tetrapod Acanthostega gunnari Jarvik: postcranial anatomy, basal tetrapod interrelationships and patterns of skeletal evolution. Transactions of the Royal Society of Edinburgh: Earth Sciences, 87(03), 363–421.
Commission, F. T., & Commission, F. T. (1958). Economic report on antibiotics manufacture. United States Government Printing Office: Washington, DC.
Cornuet, J. M., & Luikart, G. (1996). Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics, 144(4), 2001–2014.
Cortez, M. H., & Weitz, J. S. (2014). Coevolution can reverse predator–prey cycles. Proceedings of the National Academy of Sciences, 111(20), 7486–7491.
Dale, C., & Moran, N. A. (2006). Molecular interactions between bacterial symbionts and their hosts. Cell, 126(3), 453–465.
Danchin, A. (2008). Bacteria as computers making computers. FEMS Microbiology Reviews, 33(1), 3–26.
Danner, V. (1991). Western evolutionism in the Muslim world. American Journal of Islam and Society, 8(1), 67–82.
Darwin, C. (1866). On the Origins of Species by Means of Natural Selection or the Preservation of Favored Races in the Struggle for Life (4th ed.). London: John Murray.
Darwin, C. (1872). The descent of man, and selection in relation to sex (Vol. 2). D. Appleton.
Darwin, C. (1993). The autobiography of Charles Darwin, 1809-1882: with original omissions restored. WW Norton & Company.
Darwin, C. (2018). Life and letters of Charles Darwin. Charles River Editors via PublishDrive.
Davidson, E. H. (2010). The regulatory genome: gene regulatory networks in development and evolution. Elsevier.
Dawkins, R., & Krebs, J. R. (1979). Arms races between and within species. Proceedings of the Royal Society of London. Series B. Biological Sciences, 205(1161), 489–511.
Deloria, V. (1997). Red earth, white lies: Native Americans and the myth of scientific fact. Fulcrum Publishing.
Denton, M., & Scott, R. (1986). Evolution: A theory in crisis.
Dietrich, M. R., & Skipper Jr, R. A. (2012). A shifting terrain: a brief history of the adaptive landscape. The Adaptive Landscape in Evolutionary Biology, 3–15.
Dobretsov, N. L., Kolchanov, N. A., & Suslov, V. V. (2008). On important stages of geosphere and biosphere evolution. In Biosphere Origin and Evolution (pp. 3–23). Springer.
Dobzhansky, T. (1949). Towards a modern synthesis. Evolution, 3(4), 376–377.
Dobzhansky, T. (2013). Nothing in biology makes sense except in the light of evolution. The American Biology Teacher, 75(2), 87–91.
Dobzhansky, T., & Dobzhansky, T. G. (1982). Genetics and the Origin of Species (Vol. 11). Columbia university press.
Dorin, A., & Taylor, T. (2020). Rise of the Self-Replicators: Early Visions of Machines, AI and Robots That Can Reproduce and Evolve. In AI and Robots That Can Reproduce and Evolve. (1st ed.). Springer.
Dubois, M. E. F. T. (1894). Pithecanthropus erectus: eine menschenähnliche Übergangsform aus Java. Landesdruckerei.
Eberbach, E. (2002). On expressiveness of evolutionary computation: is EC algorithmic? In Proceedings of the 2002 Congress on Evolutionary Computation. CEC’02 (Cat. No. 02TH8600) (Vol. 1, pp. 564–569). IEEE.
Eberbach, E. (2005). Toward a theory of evolutionary computation. BioSystems, 82(1), 1–19.
Eberbach, E., & Burgin, M. (2009). Evolutionary automata as foundation of evolutionary computation: Larry fogel was right. In 2009 IEEE Congress on Evolutionary Computation (pp. 2149–2156). IEEE.
Eldredge, N. (1985). Unfinished synthesis: biological hierarchies and modern evolutionary thought. Oxford University Press, USA.
Ellison, P. T. (2018). The evolution of physical anthropology. American Journal of Physical Anthropology, 165(4), 615–625.
Emlen, D. J. (2014). Animal weapons: the evolution of battle. Henry Holt and Company.
Endler, J. A., & McLellan, T. (1988). The processes of evolution: toward a newer synthesis. Annual Review of Ecology and Systematics, 19(1), 395–421.
Ercit, K., & Gwynne, D. T. (2015). Darwinian balancing selection: predation counters sexual selection in a wild insect. Evolution, 69(2), 419–430.
Erwin, D. H., & Tweedt, S. (2012). Ecological drivers of the Ediacaran-Cambrian diversification of Metazoa. Evolutionary Ecology, 26(2), 417–433.
EURYDICE., Educación, E. M. de, Cultura, & Deporte. (2012). La enseñanza de las ciencias en Europa: políticas nacionales, prácticas e investigación. Ministerio de Educación.
Falcon, A. (2006). Aristotle on causality.
Farber, P. (2003). Teaching evolution & the nature of science. The American Biology Teacher, 65(5), 347–354.
Fieher, R. A. (1922). The genetical theory of natural selection, 1930. Proceedings of the Royal Society of Edinburgh, 42, 321–341.
Fisher, A. M., Cornell, S. J., Holwell, G. I., & Price, T. A. R. (2018). Sexual cannibalism and population viability. Ecology and Evolution.
Flannelly, K. J. (2017). Greek Philosophy, Early Christian Theology, Purpose, and Change. In Religious Beliefs, Evolutionary Psychiatry, and Mental Health in America (pp. 11–18). Springer.
Fleischner, T. L. (2005). Natural history and the deep roots of resource management. Nat. Resources J., 45, 1.
Ford, E. B. (1945). Polymorphism. Biological Reviews, 20(2), 73–88.
Franklin, J. (1986). Aristotle on species variation. Philosophy, 61(236), 245–252.
Fraser, A. S. (1958). Monte Carlo analyses of genetic models. Nature, 181(4603), 208–209.
Frohlich, D. R. (2014). Biology, Peirce, and Biosemiotics: Commentaires’ Cénoscopic’d’un Biologiste. The American Journal of Semiotics, 30(1/2), 173–188.
Futuyma, D. (2004). Natural selection: How evolution works. Retrieved September, 26, 2015.
Futuyma, D. (2005). Evolution (1st ed.). Sunderland: Sinauer.
Futuyma, D. J. (2005). Evolution (1st ed.). Sunderland: Sinauer.
Futuyma, D. J. (2015). Can modern evolutionary theory explain macroevolution? In Macroevolution (pp. 29–85). Springer.
Gayon, J. (1996). The individuality of the species: a Darwinian theory?—from Buffon to Ghiselin, and back to Darwin. Biology and Philosophy, 11(2), 215–244.
Gilbert, S. F., Opitz, J. M., & Raff, R. A. (1996). Resynthesizing evolutionary and developmental biology. Developmental Biology, 173(2), 357–372.
Gingerich, P. D., Smith, B. H., & Simons, E. L. (1990). Hind limbs of Eocene Basilosaurus: evidence of feet in whales. Science, 249(4965), 154–157.
Gordon, S. (2015). Design of Experiments. In Modeling and Simulation in the Systems Engineering Life Cycle (pp. 187–200). Springer.
Gotthelf, A. (1999). Darwin on Aristotle. Journal of the History of Biology, 32(1), 3–30.
Gould, Stephen J. (1987). Is a new and general theory of evolution emerging? In Self-Organizing Systems (pp. 113–130). Springer.
Gould, Stephen Jay. (1982). Darwinism and the expansion of evolutionary theory. Science, 216(4544), 380–387.
Grabowski, M. W. (2013). Hominin obstetrics and the evolution of constraints. Evolutionary Biology, 40(1), 57–75.
Greggs, T. (2009). Barth, Origen, and universal salvation: Restoring particularity. Oxford University Press.
Gross, J. B., Borowsky, R., & Tabin, C. J. (2009). A novel role for Mc1r in the parallel evolution of depigmentation in independent populations of the cavefish Astyanax mexicanus. PLoS Genetics, 5(1), e1000326.
Haldane, J. B. S. (1930). A mathematical theory of natural and artificial selection.(Part VI, Isolation.). In Mathematical Proceedings of the Cambridge Philosophical Society (Vol. 26, pp. 220–230). Cambridge University Press.
Haldane, J. B. S. (1931). A mathematical theory of natural and artificial selection. Part VII. Selection intensity as a function of mortality rate. In Mathematical Proceedings of the Cambridge Philosophical Society (Vol. 27, pp. 131–136). Cambridge University Press.
Haldane, J. B. S. (1932). A mathematical theory of natural and artificial selection. Part IX. Rapid selection. In Mathematical Proceedings of the Cambridge Philosophical Society (Vol. 28, pp. 244–248). Cambridge University Press.
Haldane, J. B. S. (1964). A defense of beanbag genetics. Perspectives in Biology and Medicine, 7(3), 343–360.
Hall, B. K. (2003). Evo-Devo: evolutionary developmental mechanisms. International Journal of Developmental Biology, 47(7–8), 491–495.
Hall, B. K. (2012). Evolutionary developmental biology. Springer Science & Business Media.
Hallam, A., & Hallam, T. (2005). Catastrophes and lesser calamities: The causes of mass extinctions. Oxford University Press on Demand.
Hameed, S. (2008). Bracing for Islamic creationism. Science, 322(5908), 1637–1638.
Hanifin, C. T., Yotsu-Yamashita, M., Yasumoto, T., & Brodie, E. D. (1999). Toxicity of dangerous prey: variation of tetrodotoxin levels within and among populations of the newt Taricha granulosa. Journal of Chemical Ecology, 25(9), 2161–2175.
Harman, O. S. (2009). Neo-Darwinism and Random Variation, or Why It’s Important to Read Darwin Carefully. In Divine Action And Natural Selection: Science, Faith and Evolution (pp. 138–152). World Scientific.
Harzsch, S., & Hafner, G. (2006). Evolution of eye development in arthropods: phylogenetic aspects. Arthropod Structure & Development, 35(4), 319–340.
Hewes, G. W. (1993). A history of speculation on the relation between tools and language. Tools, Language and Cognition in Human Evolution, 20–31.
Heywood, J. S. (1989). Sexual selection by the handicap mechanism. Evolution, 1387–1397.
Holding, M. L., Biardi, J. E., & Gibbs, H. L. (2016). Coevolution of venom function and venom resistance in a rattlesnake predator and its squirrel prey. Proceedings of the Royal Society B: Biological Sciences, 283(1829), 20152841.
Holland, J. H. (1992). Adaptation in natural and artificial systems: an introductory analysis with applications to biology, control, and artificial intelligence. MIT press.
Hossfeld, U., & Olsson, L. (2002). From the modern synthesis to Lysenkoism, and back? Science, 297(5578), 55–56.
Hoßfeld, U., & Olsson, L. (2003). The road from Haeckel: The Jena tradition in evolutionary morphology and the origins of “Evo-Devo.” Biology and Philosophy, 18(2), 285–307.
Hoßfeld, U., Olsson, L., & UH, E. H. H. (2001). Between science and politics: Axolotl research at Jena University, Germany, during the Lysenko era (1950s–1960s). Axolotl News, 29, 12–15.
Hou, Lianhai, Martin, L. D., Zhou, Z., & Feduccia, A. (1996). Early adaptive radiation of birds: evidence from fossils from northeastern China. Science, 274(5290), 1164–1167.
Hou, Liping, Faraci, G., Chen, D. T. W., Kassem, L., Schulze, T. G., Shugart, Y. Y., & McMahon, F. J. (2013). Amish revisited: next-generation sequencing studies of psychiatric disorders among the Plain people. Trends in Genetics, 29(7), 412–418.
Huneman, P. (2010). Assessing the prospects for a return of organisms in evolutionary biology. History and Philosophy of the Life Sciences, 341–371.
Huttenlocker, A. K., Grossnickle, D. M., Kirkland, J. I., Schultz, J. A., & Luo, Z.-X. (2018). Late-surviving stem mammal links the lowermost Cretaceous of North America and Gondwana. Nature, 558(7708), 108.
Huxley, J. (1929). Julian Huxley. Biologist and Statesman Ofsci.
Huxley, J. (1942). Evolution the modern synthesis. George Allen and Unwin.
Huxley, J., Hardy, A. C., & Ford, E. B. (1954). Evolution as a Process. Geroge Allen And Unwin: London.
Jäckel, C., Kast, P., & Hilvert, D. (2008). Protein design by directed evolution. Annu. Rev. Biophys., 37, 153–173.
Jamshidi, M. (2003). Tools for intelligent control: fuzzy controllers, neural networks and genetic algorithms. Philosophical Transactions of the Royal Society of London. Series A: Mathematical, Physical and Engineering Sciences, 361(1809), 1781–1808.
Jang, J. (2017). Doctrine of theodicy in a scientific age: examining the evolutionary theology of John Haught and the Daoist philosophy of Zhuangzi.
Jin, X. (2019). Translation and transmutation: the Origin of Species in China. The British Journal for the History of Science, 52(1), 117–141.
Kaiser, J. (1997). Fingering a Genetic Bottleneck. Science, 277(5330), 1205.
Kalkvik, H. M., Stout, I. J., & Parkinson, C. L. (2012). Unraveling natural versus anthropogenic effects on genetic diversity within the southeastern beach mouse (Peromyscus polionotus niveiventris). Conservation Genetics, 13(6), 1653–1664.
Kardong, K. V. (2014). Vertebrates: Comparative Anatomy, Function, Evolution (7th ed.). McGraw-Hill Education.
Katsiaficas, G. (1999). Ibn Khaldun: a dialectical philosopher for the 21st century. New Political Science, 21(1), 45–57.
Keddy, P. A. (2023). Competition. In Causal Factors for Wetland Management and Restoration: A Concise Guide (pp. 73–80). Springer.
Kidwell, S. M., & Flessa, K. W. (1995). The quality of the fossil record: populations, species, and communities. Annual Review of Ecology and Systematics, 26(1), 269–299.
Kim, S. Y., & Nehm, R. H. (2011). A cross‐cultural comparison of Korean and American science teachers’ views of evolution and the nature of science. International Journal of Science Education, 33(2), 197–227.
Kimura, M. (1968). Evolutionary rate at the molecular level. Nature, 217(5129), 624–626.
King, G. M. (1990). Dicynodonts and the end Permian event.
Kirkwood, T. B. L., & Cremer, T. (1982). Cytogerontology since 1881: a reappraisal of August Weismann and a review of modern progress. Human Genetics, 60(2), 101–121.
Koonin, E. V. (2009). Towards a postmodern synthesis of evolutionary biology. Taylor & Francis.
Koza, J. R. (1994). Genetic programming as a means for programming computers by natural selection. Statistics and Computing, 4(2), 87–112.
Kureichik, V. V, Kureichik, V. M., & Sorokoletov, P. V. (2007). Analysis and a survey of evolutionary models. Journal of Computer and Systems Sciences International, 46(5), 779–791.
Kutschera, U. (2011). From the scala naturae to the symbiogenetic and dynamic tree of life. Biology Direct, 6(1), 33.
Kutschera, U., & Niklas, K. J. (2004). The modern theory of biological evolution: an expanded synthesis. Naturwissenschaften, 91(6), 255–276.
Laats, A., & Siegel, H. (2016). Teaching evolution in a creation nation. In Teaching Evolution in a Creation Nation. University of Chicago Press.
Laland, K. N., Uller, T., Feldman, M. W., Sterelny, K., Müller, G. B., Moczek, A., … Odling-Smee, J. (2015). The extended evolutionary synthesis: its structure, assumptions and predictions. Proceedings of the Royal Society B: Biological Sciences, 282(1813), 20151019.
Lamarck, J. B. P. (2013). Zoological philosophy. Theclassics Us.
Laporte, L. F. (2000). George Gaylord Simpson: paleontologist and evolutionist. Columbia University Press.
Largent, M. A. (1995). Darwin and imperialism: The reflection in his voyage journals. Ournal of the History of Biology, 28(1), 1-32.
Largent, Mark A. (2009). The so-called eclipse of Darwinism. Transactions of the American Philosophical Society, 99(1), 3–21.
Larose, C., Parker, D. J., & Schwander, T. (2018). Fundamental and realized feeding niche breadths of sexual and asexual stick-insects. BioRxiv, 377176.
Laubichler, Manfred D, & Renn, J. (2015). Extended evolution: A conceptual framework for integrating regulatory networks and niche construction. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 324(7), 565–577.
Laubichler, Manfred D, & Rheinberger, H.-J. (2006). August Weismann and theoretical biology. Biological Theory, 1(2), 195–198.
Laubichler, Manfred Dietrich, & Maienschein, J. (2007). From embryology to evo-devo: a history of developmental evolution. Mit Press Cambridge, MA.
Layton, R. A. (2004). Didymus the Blind and his circle in late-antique Alexandria: virtue and narrative in biblical scholarship. University of Illinois Press.
Leberg, P. L., & Vrijenhoek, R. C. (1994). Variation among desert topminnows in their susceptibility to attack by exotic parasites. Conservation Biology, 8(2), 419–424.
Levit, G. S., & Olsson, L. (2006). Evolution on rails”: mechanisms and levels of orthogenesis. Ann Hist Philos Biol, 11, 99–138.
Lillegraven, J. A. (1979). Reproduction in Mesozoic mammals. Mesozoic Mammals: The First Two–Thirds of the Mammalian History, 259–276.
Lindberg, D. R. (2007). The Evolving World: Evolution in Everyday Life. American Institute of Biological Sciences.
Liow, L. H., Van Valen, L., & Stenseth, N. C. (2011). Red Queen: from populations to taxa and communities. Trends in Ecology & Evolution, 26(7), 349–358.
Little, A. C., Jones, B. C., Burt, D. M., & Perrett, D. I. (2007). Preferences for symmetry in faces change across the menstrual cycle. Biological Psychology, 76(3), 209–216.
Livingstone, F. B. (1983). The malaria hypothesis. Distribution and Evolution of Hemoglobin and Globin Loci, 1.
Lobo, I., & Shaw, K. (2008). Thomas Hunt Morgan, genetic recombination, and gene mapping. Nature Education, 1(1), 205.
López-Wilchis, R., Méndez-Rodríguez, A., Juste, J., Serrato-Díaz, A., Rodríguez-Gómez, F., & Guevara-Chumacero, L. M. (2021). Genetic Consequences of Forest Fragmentation in a Widespread Forest Bat (Natalus mexicanus, Chiroptera: Natalidae). Diversity, 13(4), 140.
Lovegrove, B. G., Lobban, K. D., & Levesque, D. L. (2014). Mammal survival at the Cretaceous–Palaeogene boundary: metabolic homeostasis in prolonged tropical hibernation in tenrecs. Proc. R. Soc. B, 281(1796), 20141304.
Lovejoy, A. O. (2011). The great chain of being: A study of the history of an idea. Transaction Publishers.
Lu, J., Zhu, M., Long, J. A., Zhao, W., Senden, T. J., Jia, L., & Qiao, T. (2012). The earliest known stem-tetrapod from the Lower Devonian of China. Nature Communications, 3, 1160.
Lumley, A. J., Michalczyk, Ł., Kitson, J. J. N., Spurgin, L. G., Morrison, C. A., Godwin, J. L., … Chapman, T. (2015). Sexual selection protects against extinction. Nature, 522(7557), 470.
Lynn, C., Glaze, A., Evans, W., & Reed, K. (2017). Evolution Education in the American South. Springer.
Mahanti, S. (2015). John Burdon Sanderson Haldane.
Maher, B. (2009). Evolution: Biology’s next top model? Nature News, 458(7239), 695–698.
Manning, A. (2001). Time, life and the Earth. Geological Society, London, Special Publications, 190(1), 253–264.
Marchesini, R., & Celentano, M. (2021). From Evolutionary Epistemology to an Extended Evolutionary Synthesis. In Critical Ethology and Post-Anthropocentric Ethics (pp. 31–60). Springer.
Mayr, E. (1982). The growth of biological thought: Diversity, evolution, and inheritance. Harvard University Press.
Mayr, E. (1997). This is biology: the science of the living world. Universities Press.
Mayr, E. (2013). Animal species and evolution. In Animal species and evolution. Harvard University Press.
Menter, I. (2021). Teacher education in Russia: Past, present, and future. Routledge.
Meyer, S. S. (1992). Aristotle, teleology, and reduction. The Philosophical Review, 101(4), 791–825.
Michael, R., & Ruse, M. (2009). Darwin and design: does evolution have a purpose? Harvard University Press.
Miller, G. (2011). The mating mind: How sexual choice shaped the evolution of human nature. Anchor.
Miller, J. D., Scott, E. C., & Okamoto, S. (2006). Public acceptance of evolution. Science-New York Then Washington-, 313(5788), 765.
Miller, K. R. (2004). The flagellum unspun—the collapse of “Irreducible Complexity. Philosophy of Biology: An Anthology, 439–449.
Montavon, T., Le Garrec, J.-F., Kerszberg, M., & Duboule, D. (2008). Modeling Hox gene regulation in digits: reverse collinearity and the molecular origin of thumbness. Genes & Development, 22(3), 346–359.
Morris, S. C. (2000). The Cambrian “explosion”: slow-fuse or megatonnage? Proceedings of the National Academy of Sciences, 97(9), 4426–4429.
Müller, G. B. (2007). Evo–devo: extending the evolutionary synthesis. Nature Reviews Genetics, 8(12), 943–949.
Müller, G. B. (2017). Why an extended evolutionary synthesis is necessary. Interface Focus, 7(5), 20170015.
Müller, G. B. (2021). Evo-devo’s contributions to the extended evolutionary synthesis. Evolutionary Developmental Biology: A Reference Guide, 1127–1138.
Newman, S. A., & Müller, G. B. (2006). 2 Genes and Form: Inherency in the Evolution of Developmental Mechanisms. In Genes in Development (pp. 38–74). Duke University Press.
Nicoglou, A. (2018). The Concept of Plasticity in the History of the Nature-Nurture Debate in the Early Twentieth Century. In The Palgrave Handbook of Biology and Society (pp. 97–122). Springer.
Niklas, K. J. (1997). The evolutionary biology of plants. University of Chicago Press.
Niklas, K. J., & Kutschera, U. (2014). Amphimixis and the individual in evolving populations: does Weismann’s Doctrine apply to all, most or a few organisms? Naturwissenschaften, 101(5), 357–372.
Nordmann, A. (1992). Darwinians at war Bateson’s place in histories of Darwinism. Synthese, 91(1–2), 53–72.
Nummela, S., Thewissen, J. G. M., Bajpai, S., Hussain, S. T., & Kumar, K. (2004). Eocene evolution of whale hearing. Nature, 430(7001), 776.
O’Corry-Crowe, G. (2008). Climate change and the molecular ecology of Arctic marine mammals. Ecological Applications, 18(sp2), S56–S76.
O’Neill, M. B., Kitchen, A., Zarley, A., Aylward, W., Eldholm, V., & Pepperell, C. S. (2018). Global Phylogeography of Mycobacterium Tuberculosis Reveals Role of Recent Human History in Pathogen Dispersal. In AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY (Vol. 165, p. 193). WILEY 111 RIVER ST, HOBOKEN 07030-5774, NJ USA.
Ochoa, J. G. D. (2018). Elastic multi-scale mechanisms: Computation and biological evolution. Journal of Molecular Evolution, 86(1), 47–57.
Ohta, T., & Gillespie, J. H. (1996). Development of neutral and nearly neutral theories. Theoretical Population Biology, 49(2), 128–142.
Oldroyd, D. (1988). Evolution: The History of an Idea. JSTOR.
Onwubolu, G. C., & Babu, B. V. (2013). New optimization techniques in engineering (Vol. 141). Springer.
Opitz, J. M., & Reynolds, J. F. (1985). The developmental field concept. American Journal of Medical Genetics, 21(1), 1–11.
Orr, H. A. (2005). The genetic theory of adaptation: a brief history. Nature Reviews Genetics, 6(2), 119.
Orr, H. A. (2009). Fitness and its role in evolutionary genetics. Nature Reviews Genetics, 10(8), 531.
Ørsted, M., Hoffmann, A. A., Sverrisdóttir, E., Nielsen, K. L., & Kristensen, T. N. (2019). Genomic variation predicts adaptive evolutionary responses better than population bottleneck history. PLoS Genetics, 15(6), e1008205.
Osborn, H. F. (1896). From the Greeks to Darwin: an outline of the development of the evolution idea (Vol. 1). Macmillan.
Osborn, H. F. (1922). Orthogenesis as observed from paleontological evidence beginning in the year 1889. The American Naturalist, 56(643), 134–143.
Owen, P. (1863). On the Archeopteryx of von Meyer, with a description of the fossil remains of a long-tailed species, from the lithographic stone of Solenhofen. Philosophical Transactions of the Royal Society of London, 153, 33–47.
Ozernyuk, N. D. (2010). Correlation of ontogenetic and evolutionary processes in view of achievements of modern genetics: Role of gene duplication. Biology Bulletin, 37(2), 100–105.
Pagden, A. (1986). The fall of natural man: the American Indian and the origins of comparative ethnology. Cambridge University Press.
Park, H. W. (2022). South Koreans’ responses to evolution and creationism: A survey and its implications. Public Understanding of Science, 31(8), 1029–1045.
Peker, D., Comert, G. G., & Kence, A. (2010). Three decades of anti-evolution campaign and its results: Turkish undergraduates’ acceptance and understanding of the biological evolution theory. Science & Education, 19, 739–755.
Pennock, R. T. (2003). Creationism and intelligent design. Annual Review of Genomics and Human Genetics, 4(1), 143–163.
Pi, M. (2017). Darwinism after the modern synthesis. In The Darwinian Tradition in Context (pp. 89–103). Springer.
Pigliucci, M., & Finkelman, L. (2014). The extended (evolutionary) synthesis debate: where science meets philosophy. BioScience, 64(6), 511–516.
Pigliucci, M., & Müller, G. B. (2010). Evolution–the extended synthesis.
Pimm, S. L., & Raven, P. (2000). Biodiversity: Extinction by numbers. Nature403 (6772): 843-845.
Plutzer, E., Branch, G., & Reid, A. (2020). Teaching evolution in US public schools: a continuing challenge. Evolution: Education and Outreach, 13(1), 1–15.
Pomiankowski, A. (1987). Sexual selection: the handicap principle does work–sometimes. Proc. R. Soc. Lond. B, 231(1262), 123–145.
Pomiankowski, A., & Iwasa, Y. (1998). Runaway ornament diversity caused by Fisherian sexual selection. Proceedings of the National Academy of Sciences, 95(9), 5106–5111.
Prothero, D. R. (2009). Evolutionary transitions in the fossil record of terrestrial hoofed mammals. Evolution: Education and Outreach, 2, 289–302.
Provine, W. B. (1989). Sewall Wright and evolutionary biology. University of Chicago Press.
Purugganan, M. D., & Fuller, D. Q. (2009). The nature of selection during plant domestication. Nature, 457(7231), 843–848.
Raby, P. (2002). Alfred Russel Wallace: A Life. Princeton University Press.
Restrepo Forero, O. (2009). El darwinismo en Colombia: visiones de la naturaleza y la sociedad. Acta Biológica Colombiana, 14, 23–40.
Rojas-Bracho, L., & Jaramillo-Legoretta, A. M. (2009). Vaquita: Phocoena sinus. In Encyclopedia of marine mammals (pp. 1196–1200). Elsevier.
Ross, M. R. (2005). Who believes what? Clearing up confusion over intelligent design and young-Earth creationism. Journal of Geoscience Education, 53(3), 319–323.
Rubin, J. (2013). The Origin of Species: The Making of a Theory. USA: HHMI Biointeractive. Retrieved from https://www.hhmi.org/biointeractive/the-origin-of-species-the-making-of-a-theory
Rutherford, S. L., & Henikoff, S. (2003). Quantitative epigenetics. Nature Genetics, 33(1), 6–8.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of Biology (10th ed.). Sinauer & MacMillian.
Savvides, N. (2003). The European dimension in the national curriculum for England. Implementing European Union Education and Training Policy: A Comparative Study of Issues in Four Member States, 143–160.
Scheuerl, T., Cairns, J., Becks, L., & Hiltunen, T. (2019). Predator coevolution and prey trait variability determine species coexistence. Proceedings of the Royal Society B, 286(1902), 20190245.
Schluter, D. (2000). The ecology of adaptive radiation. OUP Oxford.
Scott, E. C., & Matzke, N. J. (2007). Biological design in science classrooms. Proceedings of the National Academy of Sciences, 104(suppl 1), 8669–8676.
Shah, M. S. (2016). Pre-Darwinian Muslim scholars’ views on evolution.
Shen, T. V. (2015). Evolutionism through Chinese eyes: Yan Fu, Ma Junwu and their translations of Darwinian evolutionism. ASIANetwork Exchange: A Journal for Asian Studies in the Liberal Arts, 22(1), 49–60.
Shermer, M. (2002). In Darwin’s shadow: The life and science of Alfred Russel Wallace: A biographical study on the psychology of history. Oxford University Press on Demand.
Shook, B., Nelson, K., Aguilera, K., & Braff, L. (2019). Forces of Evolution. Explorations.
Singer, C. J. (1931). A short history of biology: a general introduction to the study of living things. Clarendon Press.
Singh, R. S. (2015). Darwin’s legacy II: why biology is not physics, or why it has taken a century to see the dependence of genes on the environment. Genome, 58(1), 55–62.
Siniscalco, M., Bernini, L., Filippi, G., Latte, B., Khan, P. M., Piomelli, S., & Rattazzi, M. (1966). Population genetics of haemoglobin variants, thalassaemia and glucose-6-phosphate dehydrogenase deficiency, with particular reference to the malaria hypothesis. Bulletin of the World Health Organization, 34(3), 379.
Smith, C. H., & Beccaloni, G. (2010). Natural selection and beyond: the intellectual legacy of Alfred Russel Wallace. Oxford University Press.
Smocovitis, V. B. (1997). G. Ledyard Stebbins, Jr. and the evolutionary synthesis (1924–1950). American Journal of Botany, 84(12), 1625–1637.
Smocovitis, V. B. (2006). Keeping up with Dobzhansky: G. Ledyard Stebbins, Jr., plant evolution, and the evolutionary synthesis. History and Philosophy of the Life Sciences, 9–47.
Solbrig, O. T. (1979). George Ledyard Stebbins. In Topics in Plant Population Biology (pp. 1–17). Springer.
Spaethe, J., Schmidt, A., Hickelsberger, A., & Chittka, L. (2001). Adaptation, constraint, and chance in the evolution of flower color and pollinator color vision. In Cognitive ecology of pollination: animal behavior and floral evolution. Cambridge University Press.
Spergel, D. N., Verde, L., Peiris, H. V, Komatsu, E., Nolta, M. R., Bennett, C. L., … Kogut, A. (2003). First-year Wilkinson Microwave Anisotropy Probe (WMAP)* observations: determination of cosmological parameters. The Astrophysical Journal Supplement Series, 148(1), 175.
Stanley, S. M. (1982). Macroevolution and the fossil record. Evolution, 460–473.
Stewart, E. C., & Barber, S. B. (1959). Behavior and Evolution. JSTOR.
Stoltzfus, A. (2017). Why we don’t want another “Synthesis.” Biology Direct, 12(1), 23.
Stott, R. (2012a). Darwin’s ghosts: in search of the first evolutionists. Bloomsbury Publishing.
Stott, R. (2012b). Darwin’s ghosts: the secret history of evolution. Spiegel & Grau.
Sweeney, K. (2002). History of complexity. Complexity and Health Care. An Introduction, 19–35.
Szathmáry, E., & Smith, J. M. (1995). The major evolutionary transitions. Nature, 374(6519), 227–232.
Tanghe, K. B., De Tiège, A., Pauwels, L., Blancke, S., & Braeckman, J. (2018). What’s wrong with the modern evolutionary synthesis? A critical reply to Welch (2017). Biology & Philosophy, 33(3–4), 23.
Templeton, A. R. (2013). Biological races in humans. Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, 44(3), 262–271.
Thein, K. (2006). The Life Forms and Their Model in Plato’s Timaeus. Rhizai111, 241–273.
Thompson, J. N. (1994). The coevolutionary process. University of Chicago press.
THOMSON, K. S. (1969). The biology of the lobe‐finned fishes. Biological Reviews, 44(1), 91–154.
Unoura, H. (1999). Samurai Darwinism: Hiroyuki Kato and the reception of Darwin’s theory in modern Japan from the 1880s to the 1900s. History and Anthropology, 11(2–3), 235–255.
unter Rudolph, G., & Schwefel, H.-P. (1994). Evolution are Algorithmen: Ein robustes Optimierkonzept. Physikalische Bl Atter.
Van Speybroeck, L. (2002). From epigenesis to epigenetics: the case of CH Waddington. Annals of the New York Academy of Sciences, 981(1), 61–81.
Van Valen, L. (1973). A new evolutionary law. Evol Theory, 1, 1–30.
Vermeij, G. J. (1993). Evolution and escalation: an ecological history of life. Princeton University Press.
Vizcaíno, S. F., Fariña, R. A., & Fernicola, J. C. (2009). Young Darwin and the ecology and extinction of Pleistocene South American fossil mammals. Revista de La Asociación Geológica Argentina, 64(1), 160–169.
Vyas, S. A., & Desai, S. P. (2015). The Professor and the Student, Sir Ronald Aylmer Fisher (1890–1962) and William Sealy Gosset (1876–1937): Careers of two giants in mathematical statistics. Journal of Medical Biography, 23(2), 98–107.
Waddington, C. H. (1953). Genetic assimilation of an acquired character. Evolution, 118–126.
Wade, M. J. (2011). The neo-modern synthesis: The confluence of new data and explanatory concepts. American Institute of Biological Sciences Circulation, AIBS, 1313 Dolley ….
Wagner, G. P., & Laubichler, M. D. (2004). Rupert Riedl and the re‐synthesis of evolutionary and developmental biology: body plans and evolvability. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 302(1), 92–102.
Wake, D. B., & Vredenburg, V. T. (2008). Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proceedings of the National Academy of Sciences, 105(supplement_1), 11466–11473.
Walker, G. K., Blackshaw, R. E., & Dekker, J. (1988). Leaf area and competition for light between plant species using direct sunlight transmission. Weed Technology, 2(2), 159–165.
Wallace, A. R. (1892). Note on sexual selection. Natural Science, 1, 749–750.
Walsh, C. (2003). Antibiotics: actions, origins, resistance. American Society for Microbiology (ASM).
Watterson, G. A. (1996). Motoo Kimura’s use of diffusion theory in population genetics. Theoretical Population Biology, 49(2), 154–188.
Watve, M. (1998). The Red Queen: Sex and the evolution of human nature. Resonance, 3(1), 79–80.
Weatherall, D. J. (1987). Common genetic disorders of the red cell and the ‘malaria hypothesis.’ Annals of Tropical Medicine & Parasitology, 81(5), 539–548.
Wein, T., & Dagan, T. (2019). The effect of population bottleneck size and selective regime on genetic diversity and evolvability in bacteria. Genome Biology and Evolution, 11(11), 3283–3290.
Weiss, K. M. (2004). Thomas Henry Huxley (1825–1895) puts us in our place. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 302(3), 196–206.
Wellnhofer, P. (2010). A short history of research on Archaeopteryx and its relationship with dinosaurs. Geological Society, London, Special Publications, 343(1), 237–250.
Wilde, S. A., Valley, J. W., Peck, W. H., & Graham, C. M. (2001). Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago. Nature, 409(6817), 175–178.
Williams-Ellis, A. (1966). Darwin’s moon: A biography of Alfred Russel Wallace. Blackie.
Williams, B. L., Hanifin, C. T., Brodie, E. D., & Brodie III, E. D. (2010). Tetrodotoxin affects survival probability of rough-skinned newts (Taricha granulosa) faced with TTX-resistant garter snake predators (Thamnophis sirtalis). Chemoecology, 20(4), 285–290.
Winther, R. G. (2001). August Weismann on germ-plasm variation. Journal of the History of Biology, 34(3), 517–555.
Witham, L. A. (2002). Where Darwin meets the Bible: Creationists and evolutionists in America. Oxford University Press.
Wolf, J. B., Brodie, E. D., & Wade, M. J. (2000). Epistasis and the evolutionary process. Oxford University Press.
Woods, H. (2009). Augustine and evolution: A study in the Saint’s De Genesi ad litteram and De Trinitate. Wipf and Stock Publishers.
Wysong, R. L. (1976). The Creation-evolution Controversy:(implications, Methodology and Survey of Evidence): Toward a Rational Solution. Wysong Institute.
Xanthoudaki, M. (2010). Quality science education: Where do we stand? Guidelines for practice from a European experience. Museo Nazionale Della Scienza e Della Tecnologia Leonardo Da Vinci, Lifelong Learning Programme of the European Union. Retrieved from Www. Museoscienza. Org/Setac/Resources. Asp.
Yergeau, D. A., Cornell, C. N., Parker, S. K., Zhou, Y., & Detrich III, H. W. (2005). bloodthirsty, an RBCC/TRIM gene required for erythropoiesis in zebrafish. Developmental Biology, 283(1), 97–112.
Yuthavong, Y., & Wilairat, P. (1993). Protection against malaria by thalassaemia and haemoglobin variants. Parasitology Today, 9(7), 241–245.
Zabell, S. L. (2001). Ronald Aylmer Fisher. In Statisticians of the centuries (pp. 389–397). Springer.
Zaffiri, L., Gardner, J., & Toledo-Pereyra, L. H. (2012). History of antibiotics. From salvarsan to cephalosporins. Journal of Investigative Surgery, 25(2), 67–77.
Zahavi, A. (1975). Mate selection—a selection for a handicap. Journal of Theoretical Biology, 53(1), 205–214.
Zeymer, C., & Hilvert, D. (2018). Directed evolution of protein catalysts. Annual Review of Biochemistry, 87, 131–157.
Zhang, M., Weisberg, D. S., Zhu, J., & Weisberg, M. (2022). A comparative study of the acceptance and understanding of evolution between China and the US. Public Understanding of Science, 31(1), 88–102.
Zhang, P. (2016). The four ecologies, post-evolution and singularity. Explorations in Media Ecology, 15(3–4), 343–354.
Zhang, P. (2018). Mother Pelican. A Journal of Solidarity and Sustainability, 14(2).
Zimmer, C. (2011). Evolution: the triumph of an idea. Random House.
Zirkle, C. (1941). Natural Selection before the" Origin of Species". Proceedings of the American Philosophical Society, 71–123.
Zuckerkandl, E. (2006). Intelligent design and biological complexity. Gene, 385, 2–18.
No hay comentarios:
Publicar un comentario