
Índice
Portada

1. Introducción
|| Regresar al Índice ||
El sistema tegumentario es el sistema de
órganos que protege al cuerpo de diversos tipos de daños, como la pérdida de
agua o la abrasión desde el exterior. El sistema comprende la piel y sus
apéndices (incluyendo cabello, escamas, plumas, pezuñas y cuernos). El sistema
tegumentario tiene una variedad de funciones; Puede servir para
impermeabilizar, amortiguar y proteger los tejidos más profundos, excretar
desechos y regular la temperatura, y es el sitio de unión de los receptores
sensoriales para detectar dolor, sensación, presión y temperatura. En la
mayoría de los vertebrados terrestres con la exposición significativa a la luz
del Sol, el sistema tegumentario también proporciona la síntesis de la vitamina
D. El tegumento debe servir como barrera semipermeable, la cual permita de
algún modo el flujo de algunas sustancias, especialmente en organismos
unicelulares que carecen de órganos especializados para las demás funciones
biológicas. En este orden de ideas, el primer tegumento que tuvieron los seres
vivos y del cual aún depende la gran mayoría es la membrana celular. Sin
embargo, bacterias y hongos pueden poseer una capa externa extra denominada
pared celular.
En general, el sistema tegumentario juega
un papel importante en el mantenimiento de la homeostasis. El sistema
tegumentario es el sistema de órganos más externo del cuerpo y muchas de sus
funciones están relacionadas con esta ubicación. La piel protege al cuerpo
contra agentes patógenos y químicos, minimiza la pérdida o entrada de agua y
bloquea los efectos nocivos de la luz solar. Los receptores sensoriales en la
piel proporcionan información sobre el entorno externo, ayudando a la piel a
regular la temperatura corporal en respuesta a los cambios ambientales y
ayudando al cuerpo a reaccionar ante el dolor y otros estímulos táctiles. La
gran superficie de la piel lo hace ideal para la regulación de la temperatura.
La tasa de pérdida de calor puede ser regulada por la cantidad de sangre que
fluye a través de los vasos sanguíneos en la dermis cerca de la superficie de
la piel. Cuando la temperatura corporal aumenta, como por ejemplo durante el
ejercicio, el tono simpático se reduce y esto provoca la dilatación de los
vasos sanguíneos que irrigan la piel. El aumento en el flujo sanguíneo de la
piel permite que el calor se pierda más rápidamente para que la temperatura
corporal no se eleve por encima del rango homeostático normal. La tasa de pérdida
de calor también puede aumentarse mediante la producción de sudor, que absorbe
calor adicional a medida que se evapora. Por el contrario, si la producción de
calor es menor que la requerida, los vasos dérmicos se contraen, el sudor se
detiene y el cuerpo conserva el calor.
1.1 Hieronymus
Mercurialis
(1530-1606) Fue un médico italiano y
erudito en griego y latín. Fue profesor de medicina en Padua, Bolonia y Pisa, y
escribió libros de medicina, incluido uno sobre las dolencias de los niños. En
su libro sobre las enfermedades de los niños, dedicó tres capítulos a varios
aspectos de los problemas del habla y la voz. Uno fue sobre habla y voz, uno
sobre mutismo y el tercero sobre articulación y fluidez. Él es más famoso por
su trabajo fue De Arte Gymnastica (1659).
Mercurialis nació en la ciudad de Forli,
Italia y se educó en Bolonia y Padua. Recibió su doctorado en medicina en 1555.
Luego fue a Roma y comenzó a estudiar los clásicos y la filosofía antigua y las
prácticas médicas de Aristóteles, Hipócrates y Galeno. También estudió las
actitudes antiguas hacia la dieta, el ejercicio y la higiene y el uso de
métodos naturales para curar enfermedades. Estos estudios le llevaron a
escribir en 1569 De Arte Gymnastica.
En este libro ofreció explicaciones y principios para la fisioterapia y el
ejercicio.

Su trabajo académico lo llevó a que se le
pidiera que asumiera una cátedra académica en Padua en 1569. Fue en Padua donde
completó muchos de sus estudios posteriores, incluida una traducción de las
obras de Hipócrates, un libro sobre enfermedades
de la piel (1572) que es conocido como el primer trabajo científico
dedicado a la dermatología, uno sobre las enfermedades de la mujer (1583) y
otro sobre las enfermedades de los niños (1583).
En 1573, Mercurialis fue a Viena para
tratar al emperador del Sacro Imperio Romano Germánico, Maximiliano II. El
emperador, complacido con el tratamiento de Mercurialis, lo nombró Conde
Palatino.
Mercurialis permaneció en Padua hasta
1587, cuando comenzó a enseñar y ocupar el cargo de catedrático de medicina en
la Universidad de Bolonia. En 1589, se le ofreció un puesto de profesor en la
Universidad de Pisa. Permaneció en Pisa hasta los setenta y cinco años, cuando
se retiró a Forli en 1605. Murió allí un año después.
2. Tegumento viral
2.1 Membrana
viral
Algunos virus (por ejemplo, el VIH y muchos virus animales) tienen envolturas virales que cubren sus cápsides proteicas protectoras. Las envolturas se derivan típicamente de porciones de las membranas de la célula anfitrión (fosfolípidos y proteínas), pero incluyen algunas glucoproteínas virales. Pueden ayudar a los virus a evitar el sistema inmunitario del anfitrión. Las glicoproteínas en la superficie de la envoltura sirven para identificar y unirse a los sitios receptores en la membrana del anfitrión. La envoltura viral luego se fusiona con la membrana del anfitrión, permitiendo que la cápside y el genoma viral entren e infecten al anfitrión (Harrison, 2008).

Figura 2.1. Los virus
envueltos poseen además de la propia envoltura que los hace pasar como una
célula o vesícula, un espacio entre la superficie de la cápside y la membrana
llamada matriz, donde pueden ubicarse proteínas extra llamadas tegumento viral.
La célula de la cual brota el virus mismo
a menudo morirá o se debilitará y arrojará más partículas virales durante un
período prolongado. La envoltura de la bicapa lipídica de estos virus es
relativamente sensible a la desecación, el calor y los detergentes, por lo
tanto, estos virus son más fáciles de esterilizar que los virus sin envoltura,
tienen una supervivencia limitada fuera del ambiente del anfitrión y, por lo
general, deben transferirse directamente de un anfitrión a otro. Los virus
envueltos poseen una gran adaptabilidad y pueden cambiar en poco tiempo para
evadir el sistema inmune; y causar infecciones persistentes (Harrison, 2008).
2.2 Tegumento
real
Un tegumento viral, más comúnmente
conocido como matriz viral, es un grupo de proteínas que recubre el espacio
entre la envoltura y la nucleocápside de todos los herpesvirus (Yu et al.,
2011). El tegumento generalmente contiene proteínas que ayudan en la
replicación del ADN viral y la evasión de la respuesta inmune, típicamente con
la inhibición de la señalización en el sistema inmune y la activación de
interferones. El tegumento generalmente se libera poco después de la infección
en el citoplasma. Estas proteínas generalmente se forman dentro de la fase
tardía del ciclo infeccioso viral, después de que los genes virales se han
replicado. Se ha recopilado mucha información sobre los tegumentos virales del
estudio del virus del herpes simple (Grzesik et al., 2019).
Los tegumentos virales pueden estar
dispuestos simétricamente a través de proteínas estructurales y de andamiaje o
también pueden estar dispuestos asimétricamente, dependiendo del virus. Los
tegumentos rara vez se colocan al azar y generalmente involucran proteínas de
andamiaje en su formación alrededor de la nucleocápside. Las proteínas no esenciales
incluidas en el tegumento pueden ayudar en la supresión de la respuesta inmune,
la supresión de la transcripción del ARNm del anfitrión o la supresión de las
defensas intrínsecas o celulares.
Las proteínas esenciales incluirán
factores que ayudan en el tráfico de la cápside viral al núcleo (para
herpesvirus), reclutando factores de transcripción o traducción del anfitrión,
o directamente transcribiendo o traduciendo genes virales. El contenido
tegumental se libera en el citoplasma al ingresar a la célula sobre el cual se
activan muchas proteínas tegumentales. El tegumento también puede ayudar en la
inserción del genoma viral en el citoplasma o núcleo de la célula anfitrión.
2.3 Cápside
Una cápside es la cubierta proteica de un
virus. Se compone de varias subunidades estructurales oligoméricas hechas de
proteínas llamadas protómeros. Las subunidades morfológicas tridimensionales
observables, que pueden corresponder o no a proteínas individuales, se
denominan capsómeros. La cápside encierra el material genético del virus (York, 2017).
Las cápsides se clasifican ampliamente
según su estructura. La mayoría de los virus tienen cápsides con estructura
helicoidal o icosaédrica. Algunos virus, como los bacteriófagos, han
desarrollado estructuras más complicadas debido a restricciones de elasticidad
y electrostática. La forma icosaédrica, que tiene 20 caras triangulares
equiláteras, se aproxima a una esfera, mientras que la forma helicoidal se
asemeja a la forma de un resorte, ocupando el espacio de un cilindro pero no
siendo un cilindro en sí. Las caras de la cápside pueden consistir en una o más
proteínas. Por ejemplo, la cápside del virus de la fiebre aftosa tiene caras
que consisten en tres proteínas llamadas VP1-3 (York, 2017). Las funciones de la cápside son:
👉 proteger
el genoma
👉 entregar el genoma y
👉 interactuar con el anfitrión.
El virus debe ensamblar una cubierta
proteica estable y protectora para proteger el genoma de agentes químicos y
físicos letales. Estos incluyen formas de radiación natural, extremos de pH o
temperatura y enzimas proteolíticas y nucleolíticas. Para los virus sin
envoltura, la cápside en sí misma puede estar involucrada en la interacción con
los receptores en la célula anfitrión, lo que conduce a la penetración de la
membrana de la célula anfitrión y a la internalización de la cápside. El
suministro del genoma se produce mediante el posterior recubrimiento o
desensamblaje de la cápside y la liberación del genoma en el citoplasma, o
mediante la expulsión del genoma a través de una estructura portal
especializada directamente en el núcleo de la célula anfitrión (York, 2017).
Se ha sugerido que muchas proteínas de la
cápside viral han evolucionado en múltiples ocasiones a partir de proteínas
celulares funcionalmente diversas. El reclutamiento de proteínas celulares
parece haber ocurrido en diferentes etapas de la evolución, por lo que algunas
proteínas celulares fueron capturadas y refuncionalizadas antes de la
divergencia de los organismos celulares en los tres dominios contemporáneos de
la vida, mientras que otras fueron secuestradas más recientemente. Como
resultado, algunas proteínas de la cápside están muy extendidas en los virus
que infectan organismos relacionados de forma distante, mientras que otras
están restringidas a un grupo particular de virus (por ejemplo, proteínas de la
cápside de los alfavirus) (York, 2017).
3. La piel de los procariotas
|| Regresar al Índice ||
3.1 Membrana
celular
Los muros de una casa, los muros de una caja fuerte o las láminas de un carro separan al frágil interior de ambientes impredecibles y hostiles. Estas barreras son duras y rígidas, para poder realizar esta protección. Podríamos esperar que la barrera que separa lo vivo de lo no vivo fuera una estructura igual de poderosa, dura y rígida; y, aun así, la membrana celular es de hecho una barrera delgada, frágil y dinámica. Es una bicapa lipídica que delimita toda la célula. Es una estructura formada por dos láminas de fosfolípidos, glucolípidos y proteínas que rodean, limitan la forma y contribuyen a mantener el equilibrio entre el interior (medio intracelular) y el exterior (medio extracelular) de las células. Regula la entrada y salida de muchas sustancias entre el citoplasma y el medio extracelular. Es similar a las membranas que delimitan los orgánulos de células eucariotas.

Figura 3.1. Tal como se
entiende actualmente la membrana biológica es una capa doble de grasas
especiales, con un polo soluble en agua que se denomina cabeza que mira hacia
la parte externa, y dos colas grasosas que quedan empacadas dentro. En medio de
estas moléculas vibrantes hay proteínas insertadas que le otorgan propiedades
especiales a cada membrana.
Está compuesta por dos láminas que sirven
de "contenedor" para el citosol y los distintos compartimentos
internos de la célula, así como también otorga protección mecánica. Está
formada principalmente por fosfolípidos (fosfatidiletanolamina y
fosfatidilcolina), colesterol, glúcidos y proteínas (integrales y periféricas).
La principal característica de esta barrera es su permeabilidad selectiva, lo
que le permite seleccionar las moléculas que deben entrar y salir de la célula.
De esta forma se mantiene estable el medio intracelular, regulando el paso de
agua, iones y metabolitos, a la vez que mantiene el potencial electroquímico
(haciendo que el medio interno esté cargado negativamente). La membrana
plasmática es capaz de recibir señales que permiten el ingreso de partículas a
su interior. Cuando una molécula de gran tamaño atraviesa o es expulsada de la
célula y se invagina parte de la membrana plasmática para recubrirlas cuando
están en el interior ocurren respectivamente los procesos de endocitosis y
exocitosis.
La membrana celular es mucho más que una
barrera, y, de hecho, de estas estructuras que define a la vida que conocemos
actualmente. Bajo nuestro conocimiento actual, todo lo vivo posee una membrana
biológica que aísla el ambiente interno del ambiente externo y lleva a cabo una
serie de funciones. Todas las membranas poseen una serie de funciones base, y
luego dependiendo de qué célula u orgánulo, también posee capacidades extra,
que dependen la mayoría de las veces de las proteínas de membrana. Las
funciones básicas de una membrana biológica son:
👉 separación de ambientes interno y
externo,
👉 andamiaje enzimático,
👉 barrera semipermeable,
👉 barrera selectiva,
👉 barrera sensible a los estímulos y
👉 barrera que transforma la energía.
3.2 Pared
celular bacteriana
Alrededor del exterior de la membrana
celular está la pared celular bacteriana. Las paredes celulares bacterianas están
hechas de peptidoglicano (también llamado mureína), que está hecho de cadenas
de polisacáridos reticuladas por péptidos inusuales que contienen D-aminoácidos
(Heijenoort, 2001). Las
paredes celulares bacterianas son diferentes de las paredes celulares de las
plantas y hongos que están hechos de celulosa y quitina, respectivamente (Koch, 2003). La
pared celular de las bacterias es también distinta a la que posee las arcaicas,
que no contienen peptidoglicano. La pared celular es esencial para la
supervivencia de muchas bacterias, aunque las bacterias en forma de L pueden
ser producidas en el laboratorio que carecen de una pared celular (Joseleau-Petit, Liébart, Ayala, & D’Ari, 2007).
El antibiótico penicilina es capaz de
matar las bacterias mediante la prevención de la reticulación de peptidoglicano
y esto hace que la pared celular sea débil (Koch, 2003). La enzima lisozima también puede dañar las
paredes celulares bacterianas. Hay ampliamente hablando, dos tipos diferentes
de pared celular en bacterias, llamadas Gram-positivas y Gram-negativas. Los
nombres se originan a partir de la reacción de las células a la tinción de
Gram, una prueba muy empleada para la clasificación de las especies bacterianas
(Gram, 1884).
Las bacterias Gram-positivas poseen una pared celular gruesa que contiene muchas capas de peptidoglicano y ácidos teicoicos. Por el contrario, las bacterias Gram-negativas tienen una pared celular relativamente delgada que consiste en unas pocas capas de peptidoglicano rodeadas por una segunda membrana lipídica que contiene lipopolisacáridos y lipoproteínas. Estas diferencias en la estructura pueden producir diferencias en la susceptibilidad a los antibióticos, por ejemplo, la vancomicina puede matar sólo bacterias Gram-positivas y es ineficaz contra patógenos Gram-negativos, tales como Haemophilus influenzae o Pseudomonas aeruginosa.

Figura
3.2. La pared de los Gram-positivos es
gruesa y mira hacia el exterior de la célula, mientras que la pared de los
Gram-negativos es más delgada y esta empacada en medio de dos membranas
biológicas.
3.3 Pared celular
arcaica
Aunque no son verdaderamente únicas, las
paredes celulares de las arcaicas son inusuales. Considerando que el peptidoglicano
es un componente estándar de todas las paredes celulares bacterianas, todas las
paredes celulares arcaicas carecen de dicha sustancia (White, Drummond, & Fuqua, 1995), con la excepción de un
grupo de metanógenos (Howland, 2000). En
ese grupo, el peptidoglicano es una forma modificada muy diferente de la que se
encuentra en las bacterias (White et al., 1995). Hay cuatro tipos de pared celular actualmente conocidos entre las
arcaicas.
👉 Un tipo de pared es
el compuesto de pseudopeptidoglicano (también llamado pseudomureina).
Este tipo de pared se encuentra en algunos metanógenos, como Methanobacterium
y Methanothermus (Bender, Madigan, Michael, & Stahl, 2015). Mientras que la estructura general del pseudopeptidoglucano
arcaico se asemeja superficialmente a la del peptidoglicano bacteriano, hay una
serie de diferencias químicas significativas. Al igual que el peptidoglicano
encontrado en las paredes celulares bacterianas, el pseudopeptidoglicano
consiste en cadenas poliméricas de glicanos reticuladas por conexiones
peptídicas cortas. Sin embargo, a diferencia del peptidoglicano, el ácido
N-acetilmurámico del azúcar se reemplaza por el ácido N-acetiltalosaminurónico (White et al., 1995), y los dos azúcares están unidos con un enlace β, 1-3 glicosídico
en lugar de β, 1-4. Además, los péptidos de reticulación son L-aminoácidos en
lugar de D-aminoácidos como están en bacterias (Bender et al., 2015).
👉 Un segundo tipo de pared
celular arcaica se encuentra en Methanosarcina y Halococcus. Este
tipo de pared celular se compone enteramente de una gruesa capa de polisacáridos,
que puede ser sulfatada en el caso de Halococcus (Bender et al., 2015). La estructura en este tipo de muros es compleja y no se investiga
completamente.
👉 Un tercer tipo de pared
entre las arcaicas consiste en glicoproteinas, y ocurre en el hyperthermophiles,
Halobacterium, y algunos metanogenos. En Halobacterium, las
proteínas en la pared tienen un alto contenido de aminoácidos ácidos, dando a
la pared una carga negativa total. El resultado es una estructura inestable que
se estabiliza por la presencia de grandes cantidades de iones de sodio
positivos que neutralizan la carga (Bender et al., 2015). En consecuencia, Halobacterium prospera sólo en
condiciones con alta salinidad.
👉 En otras arcaicas, como Methanomicrobium
y Desulfurococcus, la pared puede estar compuesta solamente de proteínas
de la capa superficial (White et al., 1995), conocidas como capa S. Las capas S son comunes en las bacterias, donde
sirven como el único componente de la pared celular o una capa externa
conjuntamente con los polisacáridos. La mayoría de las arcaicas son
Gram-negativas, aunque al menos un miembro Gram-positivo es conocido(White et al., 1995).
4. La piel de los hongos y otros unicelulares
|| Regresar al Índice ||
Siguiendo la idea de la unicelularidad continuamos discutiendo los tegumentos o capas externas de diferentes seres vivos, lo cual es un sinónimo de discutir la conformación de diferentes tipos de paredes celulares.

Figura 4.1. La pared de los hongos es una
estructura compleja compuesta por glucanos, manoproteínas y quitina, un
material que también emplean los animales en sus propios tegumentos.
4.1 Paredes
celulares en los hongos
Hay varios grupos de organismos que han
sido llamados "hongos". Algunos de estos grupos (Oomycete y Myxogastria)
han sido transferidos fuera de los hongos, en parte debido a las diferencias
bioquímicas fundamentales en la composición de la pared celular. La mayoría de
los hongos verdaderos tienen una pared celular que consiste en gran parte de
quitina y otros polisacáridos (Figura 4.1) (George, 1998). Los verdaderos hongos no tienen celulosa en sus paredes celulares
(Webster & Weber, 2007). En los hongos, la pared celular es la capa más externa,
recubriendo a la membrana plasmática. La pared celular de hongos es una matriz
de tres componentes principales (Webster & Weber, 2007):
👉 quitina: polímeros que
consisten principalmente en cadenas no ramificadas de N-acetilglucosamina unida
a β- (1,4) en la Ascomycota y Basidiomycota, o N-acetilglucosamina
(quitosano) ligada con poli-β- (1,4) en la Zygomycota.
Tanto la quitina como el quitosano son sintetizados y extruidos en la membrana
plasmática.
👉 glucanos: polímeros de
glucosa que funcionan para unir polímeros de quitina o quitosano. Los
β-glucanos son moléculas de glucosa que y proporcionan rigidez a la pared
celular mientras que α-glucanos funcionan como parte de la matriz.
👉 proteínas: las enzimas
necesarias para la síntesis y la lisis de la pared celular además de las
proteínas estructurales están presentes en la pared celular. La mayoría de las
proteínas estructurales que se encuentran en la pared celular están
glicosiladas y contienen manosa, por lo que estas proteínas se denominan
manoproteínas o mananas.
Debido a la falta de una especialización
real, la “piel” de varias estructuras complejas de los hongos como los
ascocarpos o los basidiocarpos no son otra cosa que una colección de hifas
densamente empacadas.
4.2 Pseudo-hongos
Por otro lado, el grupo Oomycetes, también conocido como mohos
de agua, son patógenos de plantas saprotrópicas. Hasta hace poco se creía que
eran hongos, pero evidencia estructural y molecular (Sengbusch, 2003) ha
llevado a su reclasificación como heterocontos, relacionados con algas marrones
autotróficas y diatomeas. A diferencia de los hongos, los oomicetos normalmente
poseen paredes celulares de celulosa y glucanos en lugar de quitina, aunque
algunos géneros (como Achlya y Saprolegnia) tienen quitina en sus
paredes. La fracción de celulosa en las paredes no es más del 4 al 20%, mucho
menos que la fracción de glucanos (Alexopoulos, Mims, & Blackwell, 1996). Las paredes celulares de oomicetos también contienen el
aminoácido hidroxiprolina, que no se encuentra en las paredes celulares de
hongos.
Los dictióstelidos son otro grupo
anteriormente clasificado entre los hongos. Son llamados vulgarmente mohos de
limo que se alimentan como amebas unicelulares, pero se agregan en un tallo reproductivo y
esporangio bajo ciertas condiciones. Las células del tallo reproductor, así
como las esporas formadas en el ápice, poseen una pared de celulosa (Raper, 2014). Se ha
demostrado que la pared de esporas posee tres capas, la mitad de la cual se
compone principalmente de celulosa, y la más interna es sensible a la celulasa
y la pronasa.
4.3 Paredes
celulares en las algas
Al igual que las plantas, las algas tienen
paredes celulares (Webster & Weber, 2007). Las paredes celulares de algas contienen polisacáridos (tales
como celulosa (un glucano)) o una variedad de glicoproteínas (Volvocales) o ambas. La inclusión de
polisacáridos adicionales en las paredes de las células de algas se utiliza
como una característica sinapomórfica clasificatoria.
👉 Mannanos:
Forman microfibrillas en las paredes celulares de una serie de algas marinas
verdes, incluyendo las de los géneros, Codium,
Dasycladus y Acetabularia, así como en las paredes de algunas algas rojas, como Porphyra y Bangia.
👉 Xilanos: Es un
grupo de hemicelulosas que se encuentran en las paredes celulares de las
plantas y algunas algas. Los xilanos son polisacáridos hechos de unidades de
xilosa (un azúcar pentosa). Los xilanos son casi tan ubicuos como la celulosa
en las paredes celulares de las plantas y contienen predominantemente unidades
β-D-xilosa unidas como en la celulosa.
👉 Ácido algínico:
Es un polisacárido común en las paredes celulares de algas pardas.
👉 Polisacáridos sulfonados: Se producen en las paredes celulares de la mayoría de las algas;
Los comunes en las algas rojas incluyen agarosa, carragenano, porfirano,
furcelerano y funorano.
Otros compuestos que pueden acumularse en
las paredes celulares de algas incluyen esporofenol e iones de calcio.
El grupo de algas conocidas como diatomeas
sintetizan sus paredes celulares (también conocidas como frustulas o válvulas)
de ácido silícico (específicamente ácido ortosilícico, H4SiO4). El
ácido se polimeriza intracelularmente, después la pared se extruye para proteger
la célula. Significativamente, en comparación con las paredes celulares
orgánicas producidas por otros grupos, las frustes de sílice requieren menos
energía para sintetizarse (aproximadamente 8%), un gran ahorro en el
presupuesto de energía celular total (RAVEN, 1983) y
posiblemente una explicación de mayores tasas de crecimiento en diatomeas (Furnas, 1990) que
en otras algas. En las algas marrones, los florotanninos pueden ser un
componente de las paredes celulares (Koivikko, Loponen, Honkanen, & Jormalainen, 2005).
4.4 Otros
tegumentos protistas
Muchos protistas y bacterias producen
otras estructuras superficiales celulares aparte de las paredes celulares,
externas (matriz extracelular) o internas (Barsanti & Gualtieri, 2014;
Becker, 2000; Preisig et al., 1994). Muchas algas tienen una envoltura de mucílago fuera de la célula
hecha de exopolisacaridos. Las diatomeas construyen un frustula de la sílice
extraída del agua circundante; los radiolarios, foraminíferos, y otros también
producen un esqueleto a partir de minerales, llamado testa en algunos grupos.
Muchas algas verdes, como Halimeda y Dasycladales, y algunas algas rojas, los
Corallinales, envuelven sus células
en un esqueleto secretado de carbonato de calcio. En cada caso, la pared es
rígida y esencialmente inorgánica. Es el componente no vivo de la célula.
Algunas algas doradas, ciliados y coanoflagelados producen una cubierta
exterior protectora de tipo cáscara llamada lorica. Algunos dinoflagelados
tienen una teca de placas de celulosa, y los coccolitóforos tienen coccolitos.
5. La piel de las plantas
|| Regresar al Índice ||
Las plantas evolucionaron de las algas y
como tal poseen una pared celular, pero aparte de eso son, seres vivos
multicelulares con especialización de tejidos, por lo que algunos grupos de
células se especializan en la protección del interior, y en el transporte de
gases metabólicos como el dióxido de carbono, el dioxígeno y el vapor de agua.
5.1 Pared
celular de las plantas
Una canción popular de hace más de 50 años enumeró alimentos que el escritor dijo que no le gustaban. Cada versículo terminaba con la frase: "¡Me gustan los plátanos porque no tienen huesos!" De hecho, los plátanos y todas las plantas difieren de los animales más grandes por no tener huesos o estructuras esqueléticas internas similares. Sin embargo, los árboles grandes soportan ramas y hojas que pesan muchas toneladas. Pueden hacerlo porque la mayoría de las células vegetales tienen paredes rígidas que proporcionan el soporte que se les da a los animales por medio de huesos o paredes semirrígidas que proporcionan flexibilidad. Al mismo tiempo, las paredes protegen el delicado contenido de la célula. Cuando millones de estas células funcionan juntas como un tejido, su fuerza colectiva es enorme. Las secuoyas y los árboles de Eucalipto de Tasmania, que son los árboles más grandes hoy en día, superan la masa y el volumen de los animales terrestres más grandes, los elefantes, por más de cien veces. La madera de un árbol gigante de secoya podría soportar el peso combinado de mil elefantes.

Figura 5.1. La celulosa es
un azúcar complejo muy estable, empleado por las plantas y algas para construir
sus paredes celulares.
La primera
estructura celular descubierta por Robert Hooke en 1665 fue la pared celular, y
entre las estructuras de células vegetales observadas con un microscopio, la
pared celular es la más obvia, porque define la forma de la célula. Muchos de
los especímenes preparados observados con un microscopio en biología vegetal
son simplemente restos manchados de paredes sin contenido. Pero la gran
diversidad de paredes celulares dentro y entre especies cuenta una historia
sobre la estructura y función de cada célula. Por ejemplo, las células
epidérmicas, que forman una capa fina en las superficies de todos los órganos
de la planta, suelen tener formas y tamaños inusuales. Algunas de estas células
forman cabellos que pueden secretar sustancias que disuaden a los animales de
pastar en las plantas que los producen como alcaloides. Las células de paredes
delgadas que se encuentran debajo de la epidermis de las hojas están
especializadas para su función de fotosíntesis; Y las células de paredes
gruesas de madera ayudan a transportar el agua sin colapsar.
El principal componente estructural de las paredes celulares
es la celulosa, que está compuesta de 100 a 15.000 monómeros de glucosa en
cadenas largas, y es el polímero más abundante en la Tierra. Como fuente
primaria de alimento para los animales de pastoreo y al menos indirectamente
para casi todos los otros organismos vivos, podría decirse que la mayor parte
de la vida en la Tierra depende directa o indirectamente de la pared celular
vegetal. Los seres humanos también dependen de las paredes celulares porque
proporcionan ropa, refugio, muebles, papel y combustible. Además de la
celulosa, las paredes celulares contienen típicamente una matriz de
hemicelulosa (una sustancia semejante a un pegante que contiene fibrillas de
celulosa juntas), la pectina (el material orgánico que da rigidez a las jaleas
de frutas y puede fermentarse en metanol) y glicoproteínas (proteínas que
tienen azúcares asociadas con sus moléculas).
Una lámina central, que consiste en una capa de pectina, se
produce primero cuando se forman nuevas paredes celulares. Esta lámina central
normalmente es compartida por dos células adyacentes y es tan delgada que puede
no ser visible con un microscopio de luz ordinario a menos que esté
especialmente coloreada. Una pared primaria flexible, que consta de una fina
red de celulosa, hemicelulosa, pectina y glicoproteínas, se coloca a ambos
lados de la lámina central. La reorganización, la síntesis de nuevas moléculas
y la inserción de nuevos polímeros de pared conducen al reordenamiento de la
pared celular durante el crecimiento. Las paredes secundarias, que se producen
dentro de las paredes primarias, se derivan de paredes primarias por
espesamiento e inclusión de lignina, un polímero complejo. Las paredes celulares
secundarias de las plantas generalmente contienen más celulosa (40% a 80%) que
las paredes primarias. A medida que la célula envejece, el espesor de la pared
puede variar, ocupando tan poco como 5% a más del 95% del volumen de las
células. Durante la formación de la pared secundaria, las microfibrillas de
celulosa se incrustan en la lignina, al igual que las barras de acero se
incrustan en el hormigón para formar hormigón pretensado.
5.2 La
epidermis vegetal
La capa más externa de las células de todos los órganos jóvenes de las plantas se llama epidermis. Dado que está en contacto directo con el medio ambiente, está sujeto a modificaciones por el medio ambiente y a menudo incluye varios tipos diferentes de células. La epidermis suele tener una célula de espesor, pero unas pocas plantas producen raíces aéreas llamadas raíces de velamen (por ejemplo, orquídeas) en las que la epidermis puede tener varias células de espesor, con las células externas funcionando como una esponja. Esa epidermis de múltiples capas también se produce en las hojas de algunos higos tropicales y miembros de la familia de la pimienta (Piperaceae), donde protege a la planta de la desecación.

Figura 5.2. El hormigón y
la lignina son materiales muy estables, tanto que ningún animal por si solo
cuenta con las enzimas para digerirlos, por lo que deben entablar simbiosis con
fermentadores que descomponen los azúcares, de ese modo los fermentadores y sus
anfitriones se alimentan juntos. Las vacas por ejemplo almacenan a sus
bacterias fermentadoras en una cámara pre-estomacal llamada rumen, los primates
normalmente almacenan a sus fermentadores en el ciego o apéndice intestinal.
👉 La cutícula: La mayoría de las células epidérmicas secretan una sustancia
grasa llamada cutina dentro
y sobre la superficie de las paredes exteriores. Cutina forma una capa protectora
llamada cutícula. El espesor de la cutícula (o, lo que es más
importante, la cera secretada sobre la cutícula por la epidermis) determina en
gran medida la cantidad de agua que se pierde a través de las paredes celulares
por evaporación. La cutícula también es excepcionalmente resistente a bacterias
y otros organismos patógenos y ha sido recuperada de plantas fósiles de
millones de años.
👉 La cera cuticular: Las ceras depositadas en la cutícula en una serie de plantas
pueden llegar a la superficie por difusión, migrar entre las células o viajar a
través de canales microscópicos en las paredes celulares. La susceptibilidad de
una planta a los herbicidas puede depender del espesor de estas capas de cera.
Algunos depósitos de cera son lo suficientemente extensos como para tener valor comercial. La cera de carnauba, por ejemplo, se deposita sobre las hojas. Éste y otras ceras se cosechan para su uso en pulimentos y, en el pasado, para registros fonográficos. En épocas coloniales, una cera obtenida de las hojas y de los frutos hervidos del mirto de cera se utilizó para hacer velas.

Figura 5.3. Modelo del
corte lateral de una hoja mostrando tejidos y estructuras tegumentarias como la
cutícula externa, la epidermis los estomas y los tejidos internos como los
parénquimas y los vasos conductores de agua y azúcares.
👉 Pelos: En las hojas, las paredes
epidérmicas de las células perpendiculares a la superficie a menudo asumen
formas extrañas que, bajo el microscopio, les dan la apariencia de piezas de un
rompecabezas. Las células epidérmicas de las raíces producen extensiones
tubulares llamadas pelos radiculares a una corta distancia detrás de las puntas
de crecimiento. Los pelos radiculares aumentan enormemente el área de
absorción. Los cabellos de una naturaleza diferente se producen en la epidermis
de las partes superiores de las plantas. Estos pelos forman brotes que
consisten de una a varias células. Las hojas también tienen numerosos poros
pequeños, los estomas, bordeados por pares de células epidérmicas
especializadas llamadas células guardianas que forman el aparato del estoma.
👉 La
epidermis: La
epidermis superior cubre la superficie del haz y la epidermis inferior cubre la
superficie del envés. Muchas células en estas capas carecen de cloroplastos (no
son fotosintéticas) y, por lo tanto, son relativamente transparentes. Una
característica interesante de las células de la epidermis es que la pared celular
que mira hacia el exterior de la hoja es más gruesa que la pared que mira hacia
el interior. Este grosor extra le puede proveer a la planta protección
adicional contra las heridas mecánicas o a la evaporación de agua.
👉 Los
tricomas: La epidermis de muchas hojas está
cubierta por varias estructuras semejantes a pelos llamados tricomas, los
cuales poseen funciones diversas. Los tricomas de algunas plantas ayudan a
reducir la perdida de agua reteniendo una capa de aire fresco o reflejando
parte de la radiación solar que no es necesaria para los requerimientos
fotosintéticos de las plantas.

Figura 5.4. Los tricomas
permiten defenderse contra depredadores o atrapar presas en el caso de algunas
plantas carnívoras.
Otros tricomas segregan sustancias químicas altamente
irritantes para desalentar el pastoreo por parte de los herbívoros.
Adicionalmente, cuando los tricomas son los suficientemente largos, dificultan
la caminata de los insectos sobre la superficie de las hojas o que, de hecho,
se las coman.
5.3 Peridermo
En las plantas leñosas, la epidermis es
desprendida y reemplazada por un peridermo después de que el cambium del corcho
comience a producir nuevos tejidos que aumentan la circunferencia del tallo o
la raíz. El peridermo constituye la corteza externa y se compone principalmente
de células de corcho algo rectangulares y en forma de caja, que están muertas
en la madurez.

Figura 5.5. Estructuras
tegumentarias de las plantas leñosas.
Mientras que el citoplasma de las células
del corcho todavía está funcionando, secreta una sustancia grasa llamada
suberina en las paredes. Esto hace que las células de corcho impermeables y les
ayuda a proteger el floema y otros tejidos debajo de la corteza de la desecación,
lesiones mecánicas y temperaturas de congelación. Algunos tejidos de corcho,
como los producidos por el alcornoque, se cosechan comercialmente y se utilizan
para tapones de botellas y en la fabricación de linóleo y juntas. Algunas
partes de un cambium de corcho forman bolsillos de células de parénquima
sueltas que no están impregnadas de suberina. Estos bolsillos de tejido
sobresalen a través de la superficie del peridermo; Se llaman lenticelas y
funcionan en el intercambio de gases del tallo. Las fisuras en la corteza de
los árboles tienen lenticelas en sus bases.
5.4 Tejido
glandular
Todas las células secretan ciertas
sustancias que pueden dañar el citoplasma, si se les permite acumular
internamente. Tales materiales deben ser aislados del citoplasma de las células
en las que se originan en la vacuola o ser excretados fuera del cuerpo de la
planta. A menudo, las sustancias consisten en productos de desecho que no son
de uso adicional a la planta, pero algunas sustancias, tales como néctar,
perfumes y hormonas son vitales para las funciones normales de la planta. Las
células secretoras pueden funcionar individualmente o como parte de un tejido
secretor. Las células o tejidos secretores, que a menudo se derivan del
parénquima, pueden ocurrir en una amplia variedad de lugares en una planta.
Entre los tejidos secretores más comunes están los que secretan el néctar en
las flores; Aceites en cítricos, menta; Mucílago en los pelos glandulares de
las ramas de sol y otras plantas de captura de insectos; Látex en miembros de
varias familias de plantas, como la familia Euphorbia;
Y resinas en plantas coníferas, tales como pinos. El látex y las resinas suelen
ser secretadas por células que recubren conductos de tipo tubular que forman
redes a través de ciertas especies vegetales. Algunas secreciones vegetales,
tales como resina de pino, caucho, aceite de menta y opio, tienen un valor
comercial considerable.
5.5 Estomas
La epidermis de las hojas contiene
diminutas aberturas o estomas, utilizados para el intercambio de gases entre
las células de la hoja y el medio ambiente (respiración organísmica en
plantas). Entre las células de las estomas se cuentan las células de guardia,
estas se abren y cierran por turgencia al absorber o emitir agua y iones
salinos a las células que las rodean. Los estomas son las únicas zonas sin
cutícula por lo que son propensos a perder agua, y también son el blanco de los
parásitos vegetales. Las células de guardia están asociadas a unas células de
la epidermis especializadas llamadas células subsidiarias. Las células
subsidiarias proveen una reserva de agua e iones salinos que se mueven hacia o
desde las células de guardia. Mediante el llenado y vaciado de agua, las
células de guardia cambian de forma lo que provoca la abertura o el cierre del
estoma.
Los estomas son especialmente comunes en
la epidermis del envés de las hojas orientadas de manera horizontal (con un
promedio de 100 estomas por milímetro cuadrado) y en muchas especies se ubican
únicamente en la superficie del envés. Esto se debe a que de esta forma se
reduce la pérdida de agua, ya que así las aberturas de la planta quedan a la
sombra de la hoja, y por lo tanto no quedan expuestas a la luz. Algunos tejidos
vegetales pueden llegar a poseer hasta 400 estomas por milímetro cuadrado. De
manera opuesta, las hojas de las plantas flotantes como los lirios de agua
poseen sus estomas solo en el haz de la hoja, ya que el envés se encuentra
sumergido y por lo tanto el intercambio de gases sería mucho más lento, debido
a que la solubilidad de los gases en el agua es mucho menor a la que poseen en
el aire.
6. La piel de los poríferos
|| Regresar al Índice ||
La esponja es un animal único en que sus sistemas de órganos se pueden limitar al integumento solamente. Su "piel", o pared del cuerpo, consiste en dos capas de células. La capa externa se conoce como el Pinacodermo o la epidermis. Esta capa está hecha de células planas, delgadas y apretadas, llamadas pinacocitos y otras células que tienen aberturas para que el agua pase por los llamados poroquitos. La segunda capa, debajo de la primera, se llama el choanodermo o el gastrodermo, y se compone de los coanocitos. Éstas son células encontradas sólo en esponjas. Constantemente bombea el agua del interior de las esponjas. También captan nutrientes y partículas de alimentos y ayudan en la reproducción. Estas capas celulares hacen parte de la arquitectura principal de las esponjas conocido como el sistema acuífero, así que describiremos dicho sistema como parte de su aparato tegumentario.

Figura 6.1. Modelos de los
tres sistemas acuíferos de las esponjas.
Ambas capas son
de una sola célula de espesor. Entre estas dos láminas celulares finas está el
mesohilo que es la estructura que debe proteger el tegumento, este puede ser
muy fino en algunas esponjas simples, o masivo y grueso en especies más
grandes. El pinacodermo está perforado por pequeños orificios llamados poros
dérmicos u ostia (singular, ostium), dependiendo de si la abertura está rodeada
por varias células o una sola, respectivamente. El agua es atraída a través de
estas aberturas y es conducida a través del coanodermo por el movimiento de los
flagelos de los coanocitos. Los coanocitos bombean grandes volúmenes de agua a
través del cuerpo de la esponja a presiones muy bajas, estableciendo el sistema
de corriente de agua (acuífero). Una cutícula o capa de colágeno puede cubrir
(o incluso reemplazar) el pinacodermo en algunas especies. El pinacodermo en sí
mismo puede ser una lámina externa simple, pero típicamente también cubre
algunas de las cavidades internas del sistema acuífero donde no hay coanocitos.
Las células pinacodérmicas que recubren los canales internos se llaman
endopinacocitos. El coanodermo también puede ser simple y continuo, o plegado y
subdividido de varias maneras. El mesohilo varía en grosor y juega un papel
vital en la digestión, la producción de gametos, la secreción del esqueleto y
el transporte de nutrientes y productos de desecho por células ameboides
especiales. El mesohilo incluye una mesoglea coloidal no celular en la que
están incrustadas fibras de colágeno, espículas y varias células; Como tal, es
realmente un tipo de mesénquima. Se puede encontrar un gran número de tipos de
células en el mesohilo. La mayoría de estas células pueden cambiar de un tipo a
otro según sea necesario; Pero algunos se diferencian irreversiblemente, como
los que se comprometen con la reproducción o con la formación del esqueleto.
La movilidad de todas las células, incluidos los pinacocitos
y los coanocitos, ha sido demostrada por cinematografía. Las células del
pinacodermo y del coanodermo son más estables que las del mesohilo, pero en
general, toda la estructura puede considerarse como un sistema continuamente
móvil. De hecho, algunas esponjas se mueven de un lugar a otro. Las células
amebóides a lo largo de la base de la esponja "gatean" mientras que
otras dan soporte para el borde delantero de la esponja. Bond reportó que
algunos ameobocitos se pueden liberar de la esponja y pueden moverse por su
cuenta por un tiempo, regresando finalmente al cuerpo de la esponja de nodriza (Bond, 1998).
Esta locomoción en esponjas no es suficiente para proporcionarles un mecanismo
de escape rápido de depredadores, sin embargo; En las palabras de Bond,
"El campeón velocista entre las esponjas puede moverse regularmente a más
de cuatro milímetros por día." (Bond, 1998) Durante
el crecimiento, el pinacodermo y el coanodermo son cada uno de una sola célula
de espesor. Al aumentar su plegado a medida que aumenta el volumen de mesohilo,
estas capas mantienen una relación área / volumen suficiente para mantener un
intercambio adecuado de nutrientes y residuos en todo el individuo. El
coanodermo de una célula de espesor puede permanecer simple y continuo (la
condición de asconoide), o puede plegarse (la condición de sinconoide), o puede
subdividirse en cámaras flageladas separadas (la condición de leuconoide).
6.1 Estructura
asconoide del tegumento
La condición asconoide se encuentra en algunas esponjas
calcáreas adultas, radialmente simétricas (por ejemplo, Clathrina, Leucosolenia)
y en la fase inicial (Olynthus) de
esponjas calcáreas recién colocadas. Las esponjas asconoidas rara vez superan
los 10 cm de altura y permanecen como unidades tubulares simples en vaso. Las
finas paredes encierran una cavidad central llamada aurícula (= espongocelo),
que se abre hacia el exterior a través de un solo osculo. El pinacodermo de
asconoides y esponjas muy sencillas de siconia tiene células especializadas
llamadas porocitos. Durante la embriogénesis, cada porocito se alarga y rueda
para formar un tubo cilíndrico. El porocito se extiende a través del
pinacodermo, el mesohilo delgado y el coanodermo hacia el atrio, emergiendo
entre los coanocitos adyacentes. La abertura externa del canal de los poros es
llamada ostio o poro incurrente. El coanodermo es una capa simple, desplegada
de los coanocitos que recubre todo el atrio. El agua que se mueve a través de
una esponja asconoidica fluye a través de las siguientes estructuras: ostium →
spongocoel (sobre el choanodermo) → osculum.
6.2 Estructura
siconoide del tegumento
El plegado simple del pinacodermo y del coanodermo produce la
condición de siconoide, dentro de la cual son posibles varios niveles de
complejidad. A medida que aumenta la complejidad, el mesohilo puede espesar y
parece tener dos capas. La "región cortical" externa, o corteza, a
menudo contiene elementos esqueléticos que son diferentes de los encontrados en
la porción interior del mesohilo. En aquellas esponjas con una corteza, las
aberturas incurrentes están revestidas por varias células (no formadas por un
solo porocito) y se denominan poros dérmicos. En la condición de los
sianocitos, los coanocitos están restringidos a cámaras o divertículos
específicos de la aurícula llamados cámaras de coanocitos (o cámaras flageladas
o canales radiales). Cada cámara de los coanocitos se abre a la aurícula
mediante una amplia abertura llamada apopilo. Las esponjas siconoidales con una
corteza gruesa poseen un sistema de canales o canales incurrentes que conducen
desde los poros dérmicos a través de las cámaras mesohilo a las de los
coanocitos. Las aberturas de estos canales a las cámaras de los coanocitos se
llaman prosopilas. En una esponja tan compleja, el agua que se desplaza desde
la superficie hacia el cuerpo fluye a lo largo de la siguiente ruta: poro
incurrente (dermal) → conducto incurrente → prosopilo → cámara de los
coanocitos → apóptilo → atrio → osculum. La construcción de la sinconoide se
encuentra en muchas esponjas calcáreas (por ejemplo, Scypha, también
conocida como Sycon).
Algunas esponjas siconoidas aparecen radialmente simétricas, pero su compleja
organización interna es en gran medida asimétrica.
6.3 Estructura
del tegumento leuconoide
La condición de
leuconoide se produce por el plegamiento adicional del coanodermo y un
espesamiento adicional del mesohilo por crecimiento cortical. Estas
modificaciones se acompañan de la subdivisión de las superficies flageladas en
cámaras discretas de ovocitos de coanocitos. En la condición leuconoide, se
encuentra un aumento en el número y una disminución en el tamaño de las cámaras
de los coanocitos, que por lo general se agrupan en grupos en el mesohilo
espesado. La aurícula se reduce a una serie de canales excurrentes (o canales
exhalados) que llevan el agua de las cámaras de los coanocitos al ósculo. El
flujo de agua a través de una esponja leuconoide es: poro dérmico → conducto
incurrente → prosopilo → cámara de los coanocitos → apopilo → conducto
excurrente → osculum. La organización de los leuconoides es típica de la
mayoría de las esponjas calcáreas y de todos los miembros del Demospongiae. Es
importante darse cuenta de que el caudal no es uniforme a través de las
diversas partes del sistema acuífero. Funcionalmente, es crítico que el agua se
mueva muy lentamente sobre el coanodermo, permitiendo el tiempo para
intercambios de nutrientes, gases y desechos entre el agua y los coanocitos.
Los cambios en la
velocidad del flujo de agua a través de esta tubería son una función de los
diámetros acumulados de los canales a través de los cuales se mueve el agua. La
velocidad del flujo de agua disminuye a medida que aumenta el diámetro de la
sección transversal; Por lo tanto, en una esponja, las velocidades son más
bajas sobre el coanodermo. Además, el agua que sale de la oscula debe ser
llevada lo suficientemente lejos para evitar que sea reciclada por la esponja.
En ambientes de relativamente alta turbulencia, corrientes o acción de las
olas, este potencial reciclaje de desechos no es un problema ya que las
corrientes diluyen rápidamente los desehcos. Sin embargo, las esponjas que
residen en aguas relativamente tranquilas dependen del mantenimiento de altas
velocidades de flujo de agua a través de la oscula (o sobre formas modificadas
del cuerpo) para empujar el agua excurrente lo suficientemente lejos de la
esponja para evitar las corrientes entrantes. En una esponja leuconoide de
forma irregular que vive en agua tranquila, el diámetro transversal combinado
de todos los poros incurrentes es mucho menor que el de todas las cámaras de
los coanocitos. Pero el diámetro oscular total es incluso menor que el de los poros
incurrentes. En pocas palabras, el agua entra a cierta velocidad “x”, se
ralentiza a una pequeña fracción de “x” cuando pasa sobre el coanodermo, luego
sale de la esponja a una velocidad mucho mayor que “x”. En las esponjas
complejas, las diferencias de velocidad son dramáticas. La regulación de la
tasa de flujo también se facilita en algunas esponjas, en parte, por la
actividad de las células ameboides (llamadas células centrales) que residen
cerca de las apópilos de las cámaras de los coanocitos. Estas células pueden
retardar o acelerar la salida de agua de las cámaras cambiando la forma y la
posición a través del apóptilo.
El reconocimiento de los diversos niveles de organización y
complejidad entre los poríferos permite describir rápida y sencillamente el
plan anatómico básico de una esponja. Sin embargo, hay muy poca evidencia de
que el plan asconoide sea necesariamente el más primitivo, o que todos los
linajes de poriferos se hayan movido a través de estos tres niveles de
complejidad durante su evolución. Tampoco todas las esponjas pasan por tres de
estas etapas de desarrollo. Además, las gradaciones e intermedios entre los
tres planes básicos son comunes. Sin embargo, entre las esponjas adultas, las
organizaciones más simples (asconoide y siconoide) ocurren solamente en la
clase Calcarea, que
podría ser la clase más primitiva de poriferos vivos. Además, las esponjas
calcáreas de la condición leuconoide pasan a través de las etapas asconoide y
siconoide mientras crecen, y es solamente en esta clase que ocurren los tres
planes corporales organizacionales.
6.4 Las
esponjas de hexactinelidas
Las hexactinelidas difieren considerablemente de las esponjas
calcáreas y las esponjas. Los cuerpos de las esponjas de hexactinelida exhiben
un mayor grado de simetría radial, o superficial radial, que cualquier otro
grupo. No hay pinacodermo o su equivalente en hexactinelidos. Una membrana
dérmica está presente, pero es extremadamente delgada; Ninguna estructura
celular discreta o continua lo soporta. Poros incurrentes son agujeros simples
en esta membrana dérmica. El material celular está escasamente distribuido y
forma una red trabecular que se extiende a través de las cavidades internas de
interconexión llamadas las lagunas subdérmicas. Las cámaras flageladas en forma
de dedal están dispuestas en una sola capa y están soportadas dentro de la red
trabecular. Tanto la red trabecular como las paredes de las cámaras flageladas
parecen ser sincíticas (es decir, no existen coanocitos discretos, siendo estos
conectados por puentes citoplasmáticos y una membrana celular continua). El
agua entra en los poros incurrentes, pasa a las lagunas subdérmicas, y de allí
penetra en las cámaras de los coanocitos a través de las prosopilas. La
estructura única de los hexactinelidos es tan sorprendente que algunos autores
(por ejemplo, (Bergquist,
1985)
incluso han sugerido que los hexactinelidos podrían considerarse como un phylum
separado (el Symplasma). Sin embargo,
las relaciones filogenéticas se buscan mejor en las similitudes entre los
grupos, no en las diferencias, y por este razonamiento tratamos los
hexactinelidos como poriferos.
7. La piel de los cnidarios
|| Regresar al Índice ||
El tegumento cnidario -la epidermis externa y la gastrodermis interna- incluye células mioepiteliales, vistas por muchos trabajadores como las células musculares más primitivas de los Metazoa. Estas células columnares soportan extensiones basales aplanadas, contráctiles, llamadas mionemias. En la epidermis, estas células se denominan células epiteliomusculares, y en la dermis gástrica se les denomina células musculares nutritivas. Las mionemias reposan contra el mesoglea medio o mesenquima, y los extremos opuestos de las células forman el cuerpo externo y las superficies intestinales. Las mionemias son paralelos a las superficies frías y contienen miofibrillas contráctiles. Las mionemias de células vecinas se interconectan, formando a menudo hojas longitudinales y circulares capaces de contraerse como verdaderas capas musculares.

Figura 7.1. Los cnidarios
son básicamente un bolsillo con tentáculos venenosos, si nada es una medusa, si
está en el lecho es un pólipo.
Otros tipos de células mioepiteliales
ocurren en algunos otros animales; como los miocitos contráctiles de las
esponjas. Células similares son conocidas incluso entre mamíferos, donde se
encuentran asociadas con ciertos tejidos secretores. Algunos cnidarios también
poseen músculos subepidérmicos mesenquimales, aparentemente derivados de los
elementos contráctiles de las células mioepiteliales. En las anémonas, por
ejemplo, los esfínteres de tipo cordón se hunden por debajo del epitelio y
residen como distintos músculos completamente dentro del mesénquima. Además de
las células epiteliomusculares, la epidermis contiene células sensoriales,
células portadoras de arpones venenosos llamadas cnidocitos, células glandulares
y células intersticiales. Los últimos son indiferenciados y capaces de
desarrollarse en otros tipos de células. La gatrodermis es histológicamente
algo similar a la epidermis. Junto con las células nutritivas musculares
también contiene cnidocitos (excepto en el Hydrozoa)
y las células glandulares. En los hidrozoos la capa media es un mesoglea
bastante simple parecida a la gelatina, acelular en gran parte. Scyphomedusae y cubomedusae tienen capas
mesogleales muy gruesas con células dispersas. En los antozoos, la capa media
suele ser un mesénquima grueso y ricamente celular.
7.1 Los
cnidios
A parte de estas generalidades hay que
destacar que la estructura más relevante del grupo son los cnidocitos y los
cnidios “arpones venenosos”, de la cual el grupo obtiene su nombre. Cnidae, a menudo referido colectivamente
como nematocistos en trabajos fuentes antiguas, son únicos a los cnidarios.
Tienen una variedad de funciones, incluyendo captura de presas, defensa,
locomoción y unión a sustrato. Se producen dentro de las células llamadas
cnidoblastos, que se desarrollan a partir de células intersticiales en la
epidermis y, en muchos grupos, en la gastrodermis. Una vez que la cnidia está
completamente formada, la célula se llama apropiadamente un cnidocito. Durante
la formación de una cnidia, el cnidoblasto produce una gran vacuola interna en
la que tiene lugar una reorganización intracelular compleja pero poco
comprendida. Los cnidios pueden ser productos complejos de la secreción del
aparato de Golgi del cnidoblasto. También hay cierta evidencia de que los
cnidios podrían haberse originado simbióticamente de algunos protistas
antiguos, y se han descrito estructuras semejantes a cnidios de grupos tan
diversos como dinoflagelados, "esporozoos" y microsporanos.
Los cnidios están entre las estructuras
intracelulares más grandes y más complejas conocidas. Cuando están
completamente formadas, son cápsulas en forma de cigarros o frascos, de 5-100
μm o más de longitud, con paredes delgadas compuestas de una proteína similar
al colágeno. Un extremo de la cápsula se gira hacia adentro como un túbulo
largo, hueco, en espiral, eversible. La pared externa de la cápsula consiste en
proteínas globulares de función desconocida. La pared interna está compuesta
por haces de fibrillas similares a colágeno que tienen una separación de 50-100
nm, con estriaciones cruzadas cada 32 nm (en los nematocistos de Hydra). El patrón distintivo de fibras
de mini-colágeno proporciona la fuerza de tracción necesaria para soportar la
alta presión en la cápsula. Toda la estructura está anclada a células
epiteliales adyacentes (células de soporte) o al mesénquima subyacente. Cuando
se estimula suficientemente, el tubo se separa de la célula. En los miembros de
las clases Hydrozoa, Scyphozoa, y tal vez Cubozoa, la cápsula está cubierta por
una tapa con bisagras, u opérculo, que se abre cuando la cnidio descarga. En
los miembros de estas tres clases, cada cnidio lleva una larga cerda parecida a
un cilicio llamada cnidocilio, un mecanorreceptor que provoca la descarga
cuando se estimula. El cnidocilio responde a frecuencias específicas de
vibración transmitidas por el agua. Quimiorreceptores en las células de apoyo
adyacentes en realidad puede "afinar" el cnidocilio a la frecuencia
de recepción adecuada para las presas disponibles. Los cnidios de los
anthnozoos carecen de un cnidocilio, en su lugar poseen tienen una aleta
tripartita apical. Los cnidocitos son más abundantes en la epidermis de la
región oral y los tentáculos, donde a menudo se producen en grupos de
estructuras parecidas a las verrugas llamadas "baterías de
nematocistos".
7.2 Tipos de
cnidocitos
Alrededor de 30 tipos de cnidios se han
descrito, pero se pueden asignar a tres tipos básicos.
7.3 Cnidocitos
verdaderos
Los verdaderos nematocistos tienen cápsulas de doble pared que contienen una mezcla tóxica de fenoles y proteínas. El túbulo de la mayoría de los tipos está armado con espinas o púas que ayudan a la penetración y el anclaje en la carne de la víctima. La toxina se inyecta en la víctima a través de un poro terminal en el hilo o se lleva a la herida en la superficie del túbulo.

Figura 7.2. A la izquierda (a) tenemos un nematocisto cargado y en (b) uno accionado (YouTube).

Figura 7.3. Los cnidocilios
pueden tener estructuras secundarias que ayudan a la identificación taxonómica.
👉 Espirocistos: Los espirocistos tienen cápsulas de una
sola pared que contienen mucoproteína o glicoproteína. Sus túbulos adhesivos se
envuelven y se adhieren a la víctima en lugar de penetrarla. Los túbulos
cápsulas de espirocistos nunca tienen un poro apical.
👉 Pticocistos: El tercer tipo de cnidio, el pticocisto,
difiere morfológicamente y funcionalmente de los nematocistos y de
espirocistos. El tubo de la cápsula de un pticocisto carece de espinas y un
poro apical, y es estrictamente adhesivo. Además, el túbulo se pliega en lugar
de enrollarse dentro de la cápsula. Los pticocistos ocurren solamente en los
ceriantharios y sirven para formar el tubo único en el cual estos animales
residen.
7.4 El
pinchazo
Los cnidios han
sido generalmente vistos como efectores independientes, y, de hecho, a menudo
se descargan con la estimulación directa mecánica. Sin embargo, la evidencia
experimental sugiere que el animal tiene al menos un cierto control de la
acción de sus cnidios. Por ejemplo, las anémonas hambrientas parecen tener un
umbral de disparo más bajo que los animales saciados. También se ha demostrado
que la estimulación de la descarga de cnidios en un área del cuerpo da lugar a
la descarga en las áreas circundantes. Sin embargo, los estímulos químicos y
mecánicos, inicialmente percibidos por el cnidocilio o una estructura similar,
hacen que la mayoría de los cnidios se abstengan de pinchar. Se sabe que los
cnidarios descargan sus cnidios en presencia de varios azúcares, compuestos
aminicos de bajo peso molecular y ocasionalmente glutatión, siendo este último
un producto químico que se libera cuando los animales se lesionan o cuando se
rompe el tejido. El glutatión reducido también hace que los tentáculos de
alimentación y los ozooides gastrinos se vuelvan activos, se retuerzan y se
preparen para la alimentación.
La expulsión del
túbulo de un cnidio se llama exocitosis, y una cnidio individual puede ser
despedida sólo una vez. Se han propuesto tres hipótesis para explicar el
mecanismo de disparo:
👉 la
descarga es el resultado del aumento de la presión hidrostática causada por un
flujo rápido de agua (la hipótesis osmótica);
👉 las
fuerzas intrínsecas de tensión generadas durante la cnidogénesis se liberan en
la descarga (la hipótesis de la tensión); Y
👉 las
unidades contráctiles que envuelven la cnidio causan la descarga
"apretando" la cápsula (hipótesis contráctil).
Debido al pequeño tamaño de los cnidios y a la
extrema velocidad del proceso de exocitosis, estas hipótesis han sido difíciles
de contrastar experimentalmente. Un trabajo reciente que utiliza la
microcinematografía de alta velocidad sugiere que tanto el modelo osmótico como
el de tensión pueden estar funcionando simultáneamente, y que las cápsulas
tienen muy altas prestaciones internas. El túbulo capsular en espiral es
expulsado a la fuerza y arrojado fuera de la célula de ruptura para penetrar
o envolver alrededor de una porción de la víctima. Toma sólo unos pocos
milisegundos para que el cnidio dispare, y el túbulo expulsado puede alcanzar
una velocidad de 2 m/s lo que lo convierte en uno de los procesos celulares más
rápidos de la naturaleza.
7.5 El veneno
La mayoría de los
cnidocitos contienen varias toxinas diferentes que varían en actividad y
fuerza, pero son todos venenos biológicos, bastante potentes y capaces de
someter a la presas grandes y activas, incluyendo peces. La mayoría parecen ser
neurotoxinas. Las toxinas de algunos cnidarios son lo suficientemente poderosas
como para afectar a los seres humanos (por ejemplo, las de algunas medusas,
ciertas hidroides coloniales, tales como Lytocarpus,
muchos hidrocorales, como Millepora,
muchos sifonóforos, como Physalia).
La toxina de la mayoría de los cubomedusas es más potente que el veneno de una
cobra. En Australia tropical, el doble de personas muere anualmente de
cubomedusas que por ataques de tiburones. Las picaduras de Chironex (la avispa marina) y Chiropsalmus
suelen dar como resultado un dolor severo en el mejor de los casos, y una
insuficiencia respiratoria o cardiaca mortal en el peor de los casos. En el
norte de Australia, dos veces más personas han sido asesinadas por avispas de
mar que por tiburones. Ambos ambientes ácidos y alcalinos suprimen la acción de
los cnidocitos. Por lo tanto, si usted sale de las olas con un tentáculo de
medusas rocíela orina (ácido) o bicarbonato de sodio (alcalino) para reducir el
impacto. El ablandador de carne también funciona, presumiblemente por la
desnaturalización de las toxinas, pero la cantidad necesaria podría dañar su
piel.
8. La piel de los platelmintos
|| Regresar al Índice ||
8.1 La piel
en los tubelarios
La pared del cuerpo de los turbelarios es una capa de tipos de células funcionales. La epidermis está compuesta por un epitelio total o parcialmente ciliado, sincitial o celular, con células de las glándulas y terminaciones nerviosas sensoriales distribuidas en diversos patrones. Debajo de la epidermis se encuentra una membrana basal, que a menudo es lo suficientemente gruesa para prestar cierto apoyo estructural al cuerpo. En las órdenes Acoela, Catenulida y Macrostomida, la membrana basal está aparentemente ausente, pero esta condición se interpreta como secundariamente derivada. En el interior de la membrana basal se encuentran las células del músculo liso, frecuentemente dispuestas en capas longitudinales circulares, diagonales intermedias e interiores. El área entre la pared del cuerpo y los órganos internos suele estar llena de un mesénquima (a menudo llamado parénquima) que incluye una variedad de células sueltas y fijas, fibras musculares y tejido conectivo.
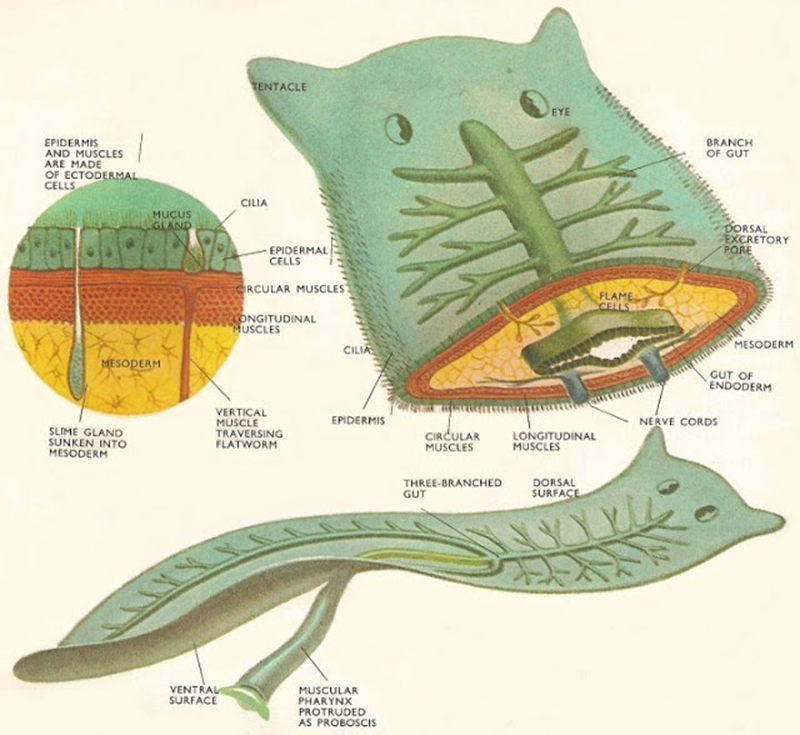
Figura 8.1. Anatomía
general de los tubelarios.
La mayoría de los acoelos, y quizás los macrostomidios, carecen de un mesénquima celular. Las células de la glándula de la pared del cuerpo generalmente se derivan del ectodermo. Cuando maduras, muchas de estas células se encuentran en el mesénquima con un "cuello" que se extiende entre las células epidérmicas a la superficie del cuerpo. Estas células producen secreciones mucosas que cumplen una serie de funciones. En las planarias semiterrestres e intertidale, el moco forma una cubierta húmeda que proporciona protección contra la desecación y ayuda en el intercambio gaseoso. La mayoría de los gusanos planos bentónicos poseen una concentración ventral de células de glándulas mucosas que segregan un lodo que ayuda a la locomoción. La secreción mucosa alrededor de la boca ayuda a la captura y la deglución de las presas. Otras células glándulares o complejos de células proporcionan adhesivos para uniones temporales a sustratos o anfitriones.

Figura 8.2. Epidermis
turbelárica y estructura de la pared corporal. (A) Epidermis del policlasdio Thysanozoon
brocchii. (B) Pared corporal y epidermis celular del tricladio Geoplana.
La mayoría de las planarias poseen las
estructuras epidérmicas llamadas rabdoides.
Estas inclusiones en forma de barra únicas generalmente se producen por
células epiteliales y luego se almacenan en paquetes dentro de la epidermis.
Tras la liberación, los rabdoides producen cantidades copiosas de moco que
pueden ayudar a proteger al animal de la desecación y de posibles depredadores.
Los rabdoides que son producidos por las células de la glándula en el
mesénquima se llaman rabditas. Estas estructuras pueden llegar a la superficie
corporal a través de los espacios intercelulares en la epidermis y también
contribuir a la producción de moco. Pueden ser responsables de la liberación de
productos químicos de defensa nocivos por algunas planarias. Algunos planarias
tienen tubérculos prominentes que cubren la superficie dorsal; estas
estructuras probablemente tienen un papel defensivo. En algunas especies, los
nematocistos sin digerir y activar de las presas cnidarias son transportados a
los tubérculos. En otros, como Thysanozoon, los tubérculos parecen liberar un
ácido potente que puede disuadir a los depredadores.
8.2 La piel
en tremátodos y cestodos
Las modificaciones de la cobertura del cuerpo exterior son comunes entre los parásitos, y los platelmintos no son una excepción. A diferencia de las planarias, los tremátodos y los céstodos poseen una cubierta externa llamada tegumento, formada por extensiones citoplasmáticas no ciliadas de células grandes cuyos cuerpos celulares se encuentran realmente en el mesénquima. El tegumento no sólo proporciona cierta protección sino que es un sitio importante de intercambio entre el cuerpo y el medio ambiente del anfitrión. Los gases y los desechos nitrogenados se mueven a través de esta superficie por difusión, y algunos nutrientes, especialmente aminoácidos, son capturados por pinocitosis. En las tenias “céstodos”, la absorción de nutrientes ocurre solamente a través de la pared del cuerpo, y la superficie del tegumento se encuentra aumentada por muchos pequeños pliegues llamados microtritos. Como una de las adaptaciones más notables de la naturaleza, estos pliegues se pueden interdigitar con los microvillos intestinales del huésped y ayudar en la absorción de nutrientes.

Figura 8.3. (A) El
tegumento y la pared corporal subyacente del trematodo (Fasciola
hepatica; sección longitudinal). (B) El tegumento y la pared
corporal de un cestodo (sección transversal).
Algunos zoólogos
consideran que la naturaleza del tegumento en los pliegues y las tenias es
única y de importancia filogenética mayor. Las larvas de estos gusanos
parásitos tienen una epidermis ciliada "normal" sobre al menos parte
de sus cuerpos. Sin embargo, esta epidermis es alterada al madurar, y las
etapas postlarvales desarrollan un nuevo cuerpo sincitial cubriendo la
neodermia, o tegumento. Este fenómeno no ocurre en ningún otro animal y debe
ser visto como una sinapomorfia clave uniendo a los Trematoda, Monogenea y Cestoda como un taxón monofilético
llamado Neodermata (en referencia a
la "piel nueva" de estos animales).
Actualmente, la monofilia de Neodermata
es indiscutible, apoyada por datos morfológicos y moleculares (Littlewood, Rohde, Bray, & Herniou,
1999).
Uno de los
atributos menos explorados, pero más interesantes de las tenias, y de hecho de
todos los parásitos intestinales, es su capacidad de prosperar en
un ambiente de enzimas hidrolíticas sin ser digerido. Una hipótesis popular es
que los parásitos intestinales producen inhibidores enzimáticos (a veces
llamados "antienzimas"). Un estudio mostró que Hymenolepis diminuta
(una tenia común en ratas y ratones) libera proteínas que parecen inhibir la
actividad de la tripsina. Esta tenia también puede regular el pH de su entorno
inmediato a aproximadamente 5,0 mediante la excreción de ácidos orgánicos; esta
salida ácida también puede inhibir la actividad de la tripsina. El estudio de
estas sustancias podría permitir el desarrollo me mejores antiácidos con las
úlceras pépticas intestinales.
9. La piel de los nematodos
|| Regresar al Índice ||
El cuerpo de los nematodos está cubierto por una cutícula bien desarrollada y con capas complejas segregada por la epidermis. La cutícula es responsable en parte de permitir la invasión de ambientes hostiles, tales como suelos terrestres secos y los tractos digestivos de anfitriones hostiles, ya que reduce drásticamente la permeabilidad de la pared del cuerpo. Los nemátodos predominantemente terrestres o parasitarios (clase Secernentea) suelen tener una capa cuticular interna fibrosa y densa, mientras que la mayoría de las formas marinas y de agua dulce de vida libre (clase Adenophorea) carecen de esta capa interna. La textura de la cutícula es muy variable entre los nematodos. Puede ser relativamente suave, o cubierta con setas sensoriales y protuberancias similares a las de las verrugas. La cutícula en muchos gusanos redondos está anillada o marcada con crestas y surcos longitudinales. En muchas formas marinas, la cutícula contiene varillas dispuestas radialmente u otras inclusiones de diversas formas. A medida que crece un nematodo, desprende su cutícula y crece una nueva a través de una serie de cuatro mudas durante su vida.

Figura 9.1. (A) Sección transversal estilizada a
través de un nematodo femenino como Ascaris
La epidermis varía entre los diferentes taxones de celular a
sincicial y a menudo se espesa como cordones longitudinales dorsales, ventrales
y laterales. Los espesores dorsales y ventrales albergan las cuerdas nerviosas
longitudinales; los espesamientos laterales contienen canales excretores
(cuando están presentes, como son en algunos secernentios) y neuronas. Interna
a la epidermis hay una capa relativamente gruesa de músculo longitudinal
oblicuamente estriado dispuesto en cuatro cuadrantes. Los músculos están
conectados a las cuerdas nerviosas dorsal y ventral por extensiones únicas
llamadas brazos musculares. Esta disposición es diferente de las uniones
neuromusculares habituales en la mayoría de los otros animales; en los
nematodos las conexiones son producidas por extensiones de las células
musculares en lugar de por las neuronas.
10. La piel de los anélidos
10.1 La piel
en los poliquetos
El cuerpo de un poliqueto está cubierto por una cutícula delgada de escleroproteína y fibras mucopolisacáridas depositadas por microvilli epidérmico. La epidermis es un epitelio columnar que a menudo es ciliado en ciertas partes del cuerpo. Debajo de la epidermis se encuentra una capa de tejido conectivo, músculos circulares y músculos longitudinales gruesos, estos últimos a menudo dispuestos como cuatro bandas. Los músculos circulares no forman una vaina continua, sino que son interrumpidos al menos en las posiciones de los parapodios.

Figura 10.1. Corte
transversal de un oligoqueto (izquierda) y de un poliqueto (derecha).
El revestimiento interior de la pared del cuerpo es el
peritoneo, que rodea los espacios celómicos y las líneas de las superficies de
los órganos internos. El celoma de un poliqueto se dispone primitivamente como
espacios pareados lateralmente (es decir, derecha e izquierda), y dispuestos en
serie (segmentalmente) dentro del tronco. Los mesenterios dorsal y ventral
separan los miembros de cada par de celomas, y los septos intersegmentales
musculares aíslan cada par se segmentos a lo largo de la longitud del cuerpo.
En algunos poliquetos, los septos intersegmentales se han perdido
secundariamente o están perforados, por lo que en estos animales el líquido
celómico es continuo entre los segmentos. En muchos pequeños poliquetos, el
forro celómico se pierde enteramente. Tales condiciones alteran radicalmente
las cualidades hidráulicas del cuerpo.
Además de la pared principal y los músculos septales, otros
músculos funcionan para retraer partes del cuerpo protrusibles y eversibles
(por ejemplo, branquias, faringes) y para operar los parapodios. Cada parapodio
es una evaginación de la pared del cuerpo y contiene una variedad de músculos.
Los parapodios movibles son operados principalmente por conjuntos de músculos
diagonales (oblicuos), que tienen su origen cerca de la línea media del cuerpo
ventral. Estos músculos se ramifican y se insertan en varios puntos dentro del
parapodio. Los parapodios grandes típicamente contienen un par de varillas de
soporte quitínicas y escleroproteináceas llamadas acículas, en las cuales
algunos músculos se insertan y operan. Los quetos también son apoyados por los
músculos y por lo general se pueden retraer y extender (muy a diferencia de las
setas de los artrópodos).
10.2 La piel
en los oligoquetos
La pared del cuerpo de los oligoquetos se construye sobre el
mismo plan que el descrito para los poliquetos. Una cutícula delgada cubre la
epidermis, que suele ser un epitelio columnar que contiene varias células
glándulares secretoras de moco. Los cilios a veces se producen en ciertas
partes de la epidermis, alrededor del prostomium
y las branquias de algunas pequeñas formas de agua dulce. Debajo de la
epidermis están las capas musculares circulares y longitudinales habituales,
estando limitada esta última internamente por el peritoneo. Ambas capas
musculares suelen ser bastante gruesas, especialmente en los oligoquetos
terrestres más grandes. Los septos intersegmentales son generalmente bien
desarrollados y musculosos. En su mayor parte son funcionalmente completos,
excepto en los extremos anterior y posterior del gusano, y perforaciones en los
septos están reguladas por los músculos especializados llamados esfínteres.
Este aislamiento eficaz de compartimentos celómicos entre sí juega un papel
importante en la locomoción de los oligoquetos. Muchos oligoquetos terrestres
llevan poros conectando espacios celómicos individuales al exterior. Estos
poros están protegidos por esfínteres y regulan el escape de los fluidos
celómicos sobre la superficie del cuerpo. Se supone que esta pérdida controlada
de fluidos corporales ayuda a mantener una película húmeda sobre el cuerpo,
facilitando el intercambio de gases y evitando la desecación. Los parapodios
están ausentes en los oligoquetos, pero los quetos ocurren en casi todas las
especies. Los quetos son movibles por varios músculos y juegan un papel en la
locomoción.
10.3 La piel
en las sanguijuelas
Una vista en sección transversal de la pared del cuerpo de
una sanguijuela es dramáticamente diferente de la de otros anélidos, en gran
parte debido a la presencia de una capa de tejido conjuntivo dérmica gruesa
debajo de la epidermis y la reducción del celoma. Una fina cutícula cubre una
sola capa de células epidérmicas simples. La epidermis contiene glándulas
mucosas, algunas de las cuales son bastante grandes y se extienden muy por
debajo de la superficie. Los músculos circulares y longitudinales habituales
están presentes, pero están organizados más flojamente que en poliquetos y
oligoquetos. Distintas bandas de músculos dorsoventrales también están
presentes, así como diagonales (oblicuso) entre las capas de músculos
circulares y longitudinales. La dermis densa llena las áreas entre las bandas
del músculo. La reducción del celoma se asocia con la proliferación de tejido
conectivo profundamente debajo de la superficie del cuerpo. Los compartimentos
coelómicos de los septos están presentes sólo en Acanthobdella (en los cinco primeros segmentos) y en la región del
cuerpo medio de los branchiobdelidos. En todos los demás miembros de esta
subclase, los espacios celómicos están representados por varios canales y
espacios que aumentan el sistema circulatorio en las sanguijuelas
rhynchobdellanias y que reemplazan completamente el sistema circulatorio en los
arhynchobdellanios. En la familia Hirudinidae,
los canales celómicos se reducen aún más por la producción de tejido botreoideo
de tipo mesénquima derivado del revestimiento celómico y proliferado hacia
fuera alrededor de los canales.
11. La piel de los moluscos
|| Regresar al Índice ||
El recubrimiento de los moluscos se divide en dos regiones,
la concha u el mando. Sin embargo, en este capítulo nos enfocaremos
exclusivamente en el manto, pues la información de la concha ya la discutimos
en la unidad del sistema esquelético. La pared corporal de los moluscos
comprende tres capas reconocibles: la cutícula, la epidermis y los músculos. La
cutícula se compone en gran parte de varios aminoácidos y proteínas
esclerotizadas (conchina), pero aparentemente no contiene quitina (excepto tal
vez en el caudofoveatanos). La epidermis es generalmente una sola capa de
células cuboidale columnares, que son ciliadas en la mayor parte del cuerpo.
Muchas de las células epidérmicas participan en la secreción de la cutícula,
mientras que otras parecen ser diferentes tipos de glándulas secretoras. La
función de la mayoría de estas glándulas no se conoce, pero algunas secretan
moco y son muy abundantes, especialmente en la superficie del cuerpo ventral.
Otras células epidérmicas especializadas se producen en la pared del cuerpo
dorsal, o manto. Estas células constituyen las glándulas de la cáscara de los
moluscos, que producen las espículas calcáreas o conchas características de
este filo. Aún otras células epidérmicas forman papilas epidérmicas sensoriales
u otros receptores. La epidermis y la capa muscular externa suelen estar
separadas por una membrana basal y ocasionalmente por una dermis. La pared del
cuerpo normalmente incluye tres capas distintas de fibras musculares lisas: una
capa circular externa, una capa diagonal media y una capa longitudinal interna.
Los músculos diagonales están a menudo en dos grupos con las fibras que se
organizan en ángulos rectos el uno al otro. El grado de desarrollo de cada una
de estas capas musculares difiere entre las clases (por ejemplo, en
solenogastros las capas diagonales están frecuentemente ausentes).
Ya hemos insinuado el significado de la cavidad del manto y
su importancia en el éxito del bauplan molusco. Aquí ofrecemos un breve resumen
de la naturaleza de la cavidad del manto, y su disposición en los grupos principales
de moluscos. El manto, como su nombre lo indica, es un órgano en forma de
lámina que forma la pared del cuerpo dorsal, y en la mayoría de los moluscos
crece durante el desarrollo en forma de uno o dos pliegues que contienen capas
musculares y canales hemocoelicos (Figura 20.14C). Este crecimiento externo
crea un espacio entre el pliegue del manto y el cuerpo propiamente dicho. Este
espacio, la cavidad del manto (o pálido), puede estar en forma de surcos poco
profundos, o una o dos grandes cámaras a través de las cuales el agua pasa por
acción ciliar o muscular. En general, la cavidad del manto alberga las
branquias, o ctenidia, y recibe el material fecal del ano y los productos de
los sistemas excretor y reproductivo. En algunos casos, el agua entrante
también transporta alimentos para la alimentación en suspensión.
11.1 Manto en
los quitones
La cavidad del manto de los quitones es un par de ranuras palliales largas situadas a lo largo de los lados del pie. El agua entra en estas ranuras desde el frente y los lados, pasando medialmente sobre los ctenidios y luego hacia atrás entre la ctenidios y el pie. Después de pasar por encima de los gonoporos y nefridióforos, el agua sale del extremo posterior de las ranuras y saca el material fecal del ano.

Figura 11.1. Anatomía
ventral de un quitón; manto, pie, branquias y boca.
11.2 Manto en
gasterópodos
La cavidad del manto único de los gasterópodos se origina durante el desarrollo como cámara situada posteriormente. A medida que avanza el desarrollo, sin embargo, la mayoría de los gasterópodos sufren torsión de la cáscara y masa visceral para llevar la cavidad del manto hacia delante, sobre la cabeza. Nuevamente, el agua que atraviesa esta cámara fluye sobre los ctenidios, el ano, los gonoporos y los nefridióforos. Un gran número de modificaciones secundarias en este plan han evolucionado en los miembros del clado Gasterópoda, incluyendo el cambio de los patrones actuales, la pérdida de ciertas estructuras asociadas, e incluso "detorsion".

Figura 11.2. Anatomía
general de un gasterópodo.
11.3 El manto
de los bivalvos
Los bivalvos poseen un par de grandes cavidades en el manto, una a cada lado del pie y la masa visceral. Los pliegues del manto alinean las cáscaras colocadas lateralmente, y a menudo se producen posteriormente como sifones inhalantes y exhalantes, a través de los cuales el agua entra y sale de la cavidad del manto. El agua pasa a través de las ctenidias, que extraen el material alimenticio suspendido y realizan el intercambio gaseoso, a través de los gonoporos y nefridióforos, y pasan el ano mientras sale a través del sifón exhalante.
Figura 11.3. Anatomía
interna de un bivalvo.
11.4 El manto
en los escafópodos
Los escafópodos llevan una concha cónica, tubular. El agua entra y sale de la cavidad del manto alargada a través de la pequeña abertura en la cáscara y fluye sobre la superficie del manto, que, en ausencia de ctenidia, es el sitio de intercambio gaseoso. El ano, los nefridióforos y los gonoporros también se vacían en la cavidad del manto. En todos los casos anteriores, el agua se mueve a través de la cavidad del manto por la acción de los cilios.

Figura 11.4. Anatomía general
de un pulpo.
En los cefalópodos, sin embargo, los músculos del manto
altamente desarrollados y altamente inervados realizan esta función. La
superficie corporal expuesta y carnosa de los calamares y los pulpos es, de
hecho, el propio manto. El manto de estos moluscos, que no está restringido por
una concha externa, se expande y se contrae para atraer agua hacia la cavidad
del manto y luego lo fuerza a través de un estrecho embudo muscular (= sifón).
Este chorro de agua exhalante proporciona un medio de locomoción rápida para la
mayoría de los cefalópodos. En la cavidad del manto el agua pasa sobre la
ctenidia, el ano, los poros reproductivos y las aberturas de excreción. Las
notables cualidades adaptativas del plan del cuerpo de los moluscos se manifiestan
en estas variaciones en la posición y función de la cavidad del manto y sus
estructuras asociadas. De hecho, incluso la naturaleza de muchas otras
estructuras está influenciada por la disposición de la cavidad del manto. El
hecho de que los moluscos hayan podido explotar con éxito una amplia gama de
hábitats y estilos de vida puede explicarse en parte por estas variaciones, que
son centrales en la historia de la evolución de los moluscos.
12. La piel de los artrópodos
|| Regresar al Índice ||
Los artrópodos (clase Athropoda) están cubiertos con un tegumento resistente, resiliente
o exoesqueleto de quitina. Generalmente, el exoesqueleto tendrá áreas
engrosadas en las que la quitina se refuerza o se endurece con materiales tales
como minerales o proteínas endurecidas. Esto ocurre en partes del cuerpo donde
hay una necesidad de rigidez o elasticidad. Típicamente los cristales
minerales, principalmente carbonato de calcio, se depositan entre la quitina y
las moléculas de la proteína en un proceso llamado biomineralization. Los
cristales y las fibras se interpenetran y se refuerzan mutuamente, los
minerales suministran la dureza y la resistencia a la compresión, mientras que
la quitina proporciona la resistencia a la tracción. La biomineralización
ocurre principalmente en los crustáceos; en los insectos y arácnidos los
principales materiales de refuerzo son diversas proteínas endurecidas por la
vinculación de las fibras en procesos llamados esclerización y las proteínas
endurecidas se llaman esclerotina. En cualquier caso, en contraste con el caparazón
de una tortuga o el cráneo de un vertebrado, el exoesqueleto tiene poca
capacidad para crecer o cambiar su forma una vez que ha madurado. Excepto en
casos especiales, cada vez que el animal necesita crecer, se muda, derramando
la piel vieja después de crecer una nueva piel desde abajo.
12.1 Cutícula
y pared corporal
Una sección transversal a través de un
segmento corporal de un artrópodo revela mucho sobre su arquitectura general. Como
se ha indicado anteriormente, la cavidad corporal es un hemocele abierto, y los
órganos se bañan directamente en el fluido hemocelico o hemolinfa, o sangre. La
pared del cuerpo está compuesta por una cutícula compleja y estratificada por
una epidermis subyacente. La epidermis, a menudo referida en los artrópodos
como la hipodermis, es típicamente un epitelio cuboidal simple. En general, cada
segmento del cuerpo (o somita) es "encajonado" por placas
esqueléticas llamadas escleritas. Cada somita tiene típicamente una esclerita
dorsal y ventral grande, llamadas tergita y el esternita respectivamente. Las
piernas (y las alas) de los artrópodos se articulan en esta región pleural.
Existen numerosas desviaciones secundarias de este plan, como la fusión o
pérdida de escleritas adyacentes. Las bandas musculares se unen en puntos donde
las superficies internas de las escleritas se proyectan hacia el interior como
crestas o tubérculos, llamados apodemas.
12.2 Esclerotización
Debajo de la epicuticula está la procuticula relativamente gruesa, que se puede subdividir en una exocuticula exterior y una endocuticula interna (figura anterior A, B) La procuticula consiste principalmente de capas de la proteína y quitina (pero no colágeno). Es intrínsecamentedura pero flexible. De hecho, ciertos artrópodos poseen exoesqueletos algo blandos y flexibles (por ejemplo, muchas larvas de insectos, partes de arañas, y algunos pequeños crustáceos). Sin embargo, en la mayoría de los artrópodos, la cutícula es dura e inflexible, excepto en las articulaciones, una condición provocada por uno o ambos procesos: esclerotización y mineralización. El endurecimiento cuticular por esclerotización ocurre en diversos grados en todos los artrópodos. La disposición en capas de las proteínas no esclerotizadas proporciona una estructura flexible. Para producir una estructura rígida esclerizada, las moléculas proteicas están unidas entre sí por enlaces ortoquinona. El agente de unión se produce típicamente a partir de polifenoles y se cataliza por polifenol oxidasas presentes en las capas de proteína de la cutícula. La esclerótización generalmente comienza en la capa de cuticulina de la epicucula y progresa en grados, donde se asocia con un oscurecimiento distintivo.

Figura 12.1. Anatomía general de la cutícula de los artrópodos.
12.3 Mineralización
La mineralización
del esqueleto es en gran parte un fenómeno de los crustáceos, y se logra
mediante la deposición de carbonato de calcio en la región externa de la
procutícula. La epidermis es responsable de la secreción de la cutícula, y como
tal contiene varias glándulas unicelulares (Figura anterior C), algunas de las
cuales llevan conductos a la superficie de la cutícula. Debido a que la
cutícula es secretada por las células de la epidermis, a menudo lleva sus
impresiones en forma de patrones geométricos microscópicos. La epidermis se
sustenta en una membrana basal distinta que forma el límite exterior de la
cavidad corporal o hemoceloma.
12.4 Propiedades
mecánicas
Las dos capas de la cutícula tienen
propiedades diferentes. La capa externa es donde ocurre la mayor parte del
engrosamiento, biomineralización y esclerotización, y su material tiende a ser
fuerte bajo esfuerzos de compresión, aunque más débil bajo tensión. Cuando una
región rígida falla bajo estrés, lo hace por agrietamiento. La capa interior no
está tan altamente esclerotizada, y es correspondientemente más blanda; resiste
las tensiones, pero es susceptible al fallo bajo compresión. Esta combinación
es especialmente eficaz para resistir la depredación, ya que los depredadores
tienden a ejercer compresión sobre la capa externa y la tensión en el interior.
Su grado de esclerotización o mineralización determina cómo la cutícula
responde a la deformación. Por debajo de cierto grado de deformación, los
cambios de forma o de dimensión de la cutícula son elásticos y la forma
original vuelve después de que se elimina la tensión. Más allá de ese nivel de
deformación, no reversible, la deformación plástica se produce hasta que
finalmente la cutícula se agrieta o se divide generando el sonido de craqueo
típico. Generalmente, cuanto menos esclerótica sea la cutícula, mayor será la
deformación necesaria para dañar irremediablemente la cutícula. Por otro lado,
cuanto más pesadamente la cutícula está blindada, mayor es la tensión requerida
para deformarla.
12.5 Segmentación
Por regla general, el exoesqueleto de artrópodo se divide en diferentes unidades funcionales, cada una de las cuales comprende una serie de segmentos agrupados.

Figura 12.2. La segmentación
y la especialización de los segmentos es una característica común en la
evolución de los artrópodos.
Este grupo se denomina tagma, y los tagmata se adaptan a diferentes funciones en un cuerpo de artrópodo dado. Por ejemplo, los tagmata de insectos incluyen la cabeza, que es una cápsula fusionada, el tórax que actúa casi como una cápsula fija, y el abdomen usualmente dividido en una serie de segmentos articulados. Cada segmento tiene escleritas según sus requisitos para la rigidez externa; por ejemplo, en la larva de algunas moscas, no hay ninguna y el exoesqueleto es efectivamente membranoso; el abdomen de una mosca adulta está cubierto de escleritas ligeras conectadas por articulaciones de cutícula membranosa. En algunos escarabajos la mayoría de las articulaciones están tan firmemente conectadas, que el cuerpo está prácticamente en una caja blindada, rígida. Sin embargo, en la mayoría de los artrópodos, los tagmata corporales están tan conectados y unidos con cutícula flexible y músculos que tienen al menos alguna libertad de movimiento, y muchos de estos animales, como los Chilopoda o las larvas de los mosquitos son muy móviles. Además, las extremidades de los artrópodos se unen, tan característico que el mismo nombre "Arthropoda" literalmente significa "piernas articuladas" en la reflexión del hecho.

Figura 12.3. Nombres de los segmentos de diferentes grupos
de artrópodos modernos.
12.6 Muda
La naturaleza química y física del exoesqueleto de los artrópodos limita su capacidad de estirar o cambiar de forma a medida que el animal crece. En algunos casos especiales, como el abdomen de reinas de termitas no hay placas duras de exoesqueleto, en todos los demás que están encerrados esto significa que el crecimiento continuo de artrópodos no es posible. Por lo tanto, el crecimiento es periódico y se concentra en un período de tiempo cuando se cambia el exoesqueleto, llamado muda o ecdisis, que está bajo el control de una hormona llamada ecdisona.

Figura 12.4. La muda o
ecdisis se produce debido a que el animal crece pero la coraza no, (YouTube)
(YouTube).
La muda es un proceso complejo que es
invariablemente peligroso para el artrópodo implicado. Antes de que el antiguo
exoesqueleto se cambie, la cutícula se separa de la epidermis a través de un
proceso llamado apolisis. La cutícula nueva es excretada por la epidermis
subyacente, y las sales minerales son usualmente retiradas de la vieja cutícula
para su reutilización. Después de que la vieja cutícula se desprenda, el
artrópodo típicamente infla su cuerpo por bombeo (por ejemplo, por aire o toma
de agua) para permitir que la nueva cutícula se expanda a un tamaño más grande:
el proceso de endurecimiento por deshidratación de la cutícula entonces tiene
lugar. Un artrópodo recién mudado típicamente es de color pálido; en ese estado
se dice que es teneral. Por lo general, el exoesqueleto se oscurece o gana
color a medida que su exoesqueleto se endurece.
13. La piel de algunos deuterostomos
|| Regresar al Índice ||
13.1 Equinodermos
Una epidermis cubre los cuerpos de todos los equinodermos y
se superpone a una dermis derivada del mesodermo, que contiene los elementos
esqueléticos, llamados osículos. Internos a la dermis y los osículos son las
fibras o capas musculares y el peritoneo del celoma. El grado de desarrollo del
esqueleto y los músculos varía mucho entre los grupos.
En erizos y dólares de arena, los osículos están firmemente unidos entre sí para formar un marco rígido, y los músculos de la pared del cuerpo están débilmente desarrollados.

Figura 13.1. Estructura de
la pared del cuerpo del equinodermo y algunos elementos esqueléticos. (A) La
pared del cuerpo de un erizo (sección compuesta).
En los pepinos de mar, sin embargo, los osículos están
separados y se encuentran dispersos en la dermis carnosa; aquí hay capas
musculares distintas. Entre estas condiciones extremas se encuentran casos en
los que las placas esqueléticas adyacentes se articulan en varios grados de
desarrollo. En los brazos de estrellas de mar, por ejemplo, los músculos de la
pared del cuerpo están dispuestos en bandas entre las placas, proporcionando
varios grados de movimiento del brazo.
En algunos grupos las placas esqueléticas se desarrollan hasta tal punto que casi reemplazan las cavidades internas. En las estrellas quebradizas, por ejemplo, cada "segmento" del brazo contiene un osculo central esquelético llamado vértebra (véase imagen siguiente), y los cólicos del brazo están reducidos a pequeños canales. Del mismo modo, el brazo celulares en los crinoides es muy reducido por placas esqueléticas.

Figura 13.2. Estructura de
la pared del cuerpo del equinodermo y algunos elementos esqueléticos. (B)
Espinas del dólar de arena Echinarachnius parma (SEM). Las flechas
apuntan a los tractos ciliares. La barra de escala representa 100 µm. (C)
Osículos esqueléticos de los discos centrales de cuatro especies de estrellas
frágiles (Ophiuroidea), que se muestran en las vistas superior (fila
superior), lateral (fila central) y basal (fila inferior). La barra de escala
representa 0.05 mm. (D) Osículos esqueléticos de los holoturianos Psolus
chintinoides. La estructura de stereom se muestra con dos aumentos. La
barra de escala representa 100 µm. (E) Tipos de pedicellaria equinoides que
rodean la base de una columna vertebral grande. (F, G) Pedicellaria elevada
utilizada para la captura de presas por la estrella de mar Stylasterias
forreri: F, pedicellaria abierta y extendida; G,pedicellariae
retraído. (H) Detalles de una pedicellaria generalizada. (I) Dos tipos de
sistemas musculares en pedicellariae. (J) Una columna vertebral móvil
(sección).
13.2 Tunicados
La pared del cuerpo de los tunicados
incluye un epitelio simple superpuesto por una túnica secretada de espesor y
consistencia variable. La túnica está más desarrollada en los ascidianos y
algunos thaliaceanos. Esta varía de suave y gelatinosa a dura y coriácea, y a
veces incluye espículas calcáreas. La matriz de la túnica contiene fibras y
está compuesta en gran parte de un carbohidrato similar a la celulosa llamado
tunicina. Sin embargo, la túnica no es una cutícula simple de naturaleza
inorgánica, ya que también contiene amebocitos y, en algunos casos, células
sanguíneas e incluso vasos sanguíneos. La túnica puede ser vista como un
exoesqueleto que proporciona apoyo y protección. Algunos ascidianos albergan
algas simbióticas en sus túnicas. Estas algas incluyen una variedad de especies
procariotas y eucariotas de varios grupos linajes diferentes.
13.3 Cefalocordados
El cuerpo de cefalocordatos está cubierto en todas partes por
una epidermis de epitelio columnar simple, subyacente por una delgada membrana
del tejido conjuntivo. Los músculos de la pared del cuerpo son claramente
parecidos a los de los vertebrados y se presentan como bloques en forma de
chevron llamados miotomos dispuestos longitudinalmente a lo largo de los
aspectos dorsolaterales del cuerpo. Estos bloques musculares son grandes y
ocupan gran parte del interior del cuerpo, reduciendo así el celoma a espacios
relativamente pequeños. El notocordio persiste en los adultos y proporciona el
soporte estructural principal para el cuerpo.
14. Generalidades del tegumento de los vertebrados
|| Regresar al Índice ||
El integumento (o piel) es un órgano compuesto. En la superficie está la epidermis, debajo de ella está la dermis, y entre ellos se encuentra la membrana basal (lámina basal y lámina reticular). La epidermis se deriva del ectodermo y produce la lámina basal (Figura 14.1). La dermis se desarrolla a partir del mesodermo y mesénquima, y produce la lámina reticular. Entre el tegumento y la musculatura profunda del cuerpo se encuentra una región subcutánea transicional compuesta de tejidos conectivos y adiposos muy sueltos. En el examen microscópico, esta región se denomina hipodermis. En la disección anatómica bruta, la hipodermis se conoce como la fascia superficial Figura 14.1b).

Figura 14.1. Desarrollo embrionario de la piel. (a) Sección transversal de un
embrión de vertebrado representativo. El ectodermo se diferencia inicialmente
en un estrato basal profundo, que repone el peridermo externo. El dermatoma se
deposita debajo de la epidermis para diferenciarse en la capa de tejido
conectivo de la dermis. A medida que las células de la cresta neural migratoria
pasan entre la dermis y la epidermis, algunas se depositan entre estas capas
para convertirse en cromatóforos. (b) La epidermis se diferencia aún más en una
capa estratificada que a menudo tiene una capa mucosa o cutícula en la
superficie. Dentro de la dermis, el colágeno forma capas (capas) distintivas
que constituyen el estrato compacto. La membrana basal se encuentra entre la
epidermis y la dermis. Debajo de la dermis y la capa más profunda de la
musculatura se encuentra la hipodermis, una colección de tejidos conectivos y
adiposos sueltos.
El integumento es uno de los órganos más grandes del cuerpo,
constituyendo alrededor del 15% del peso corporal humano. La epidermis y la
dermis juntos forman algunas de las estructuras más variadas que se encuentran
dentro de los vertebrados. La epidermis produce pelo, plumas, barbas de
filtración de ballenas, garras, cuernos, picos y algunos tipos de escamas. La
dermis da lugar a huesos dérmicos y osteodermos de reptiles. Colectivamente, la
epidermis y la dermis forman dientes, dentículos y escamas de peces. De hecho,
los destinos de desarrollo de la dermis y la epidermis están tan estrechamente
unidos a través de la membrana basal que, en ausencia de uno, el otro por sí
mismo es incapaz de producir estas estructuras especializadas. En términos de
desarrollo embrionario, entonces, la epidermis y la dermis están fuertemente acopladas
y son mutuamente necesarias.
Como el límite
crítico entre el organismo y su entorno exterior, el integumento tiene una
variedad de funciones especializadas. Forma parte del exoesqueleto y se espesa
para resistir lesiones mecánicas. La barrera que establece impide la entrada de
patógenos. Los hisopos de piel de voluntarios humanos sanos identificaron más
de 200 géneros diferentes de bacterias residentes, incluyendo muchos que, si se
les da la oportunidad a través de una brecha en la piel, producirían
infecciones graves de estafilococos, acné y eczema, entre otras patologías. El
integumento también ayuda a sostener la forma de un organismo. La regulación
osmótica y el movimiento de gases e iones hacia y desde la circulación son
ayudados por el tegumento en conjunción con otros sistemas. La piel reúne el
calor necesario, o irradia el exceso, y alberga receptores sensoriales. Posee
plumas para la locomoción, pelo para aislamiento y cuernos para la defensa. Los
pigmentos de la piel bloquean la luz solar dañina y exhiben colores brillantes
durante el cortejo. La lista de funciones se puede ampliar fácilmente. La
notable variedad de estructuras y funciones de la piel hace difícil resumir
brevemente las formas y funciones del tegumento. Comencemos examinando el
origen embrionario y el desarrollo de la piel.
14.1 Origen
embrionario
Al final de la neurulación en el embrión, la mayoría de los precursores de la piel están delineados. El ectodermo superficial de una sola capa prolifera para dar lugar a la epidermis multicapa. La capa profunda de la epidermis, el estrato basal (estrato germinativo), descansa sobre la membrana basal. A través de la división celular activa, el estrato germinativo repone la capa única de células externas llamada periderma (figura anterior a). Las capas de piel adicionales se derivan de estos dos como producto de la diferenciación. La dermis surge de varias fuentes, principalmente del dermatoma. Los epímeros segmentales (somitas) se dividen, produciendo el esclerótomo medialmente, la fuente embrionaria de las vértebras, y el dermomiótomo lateralmente. Las células internas del dermomiótomo se reorganizan en el miotomo, la fuente principal del músculo esquelético. La pared exterior del dermomiótomo se extiende bajo el ectodermo como un dermatoma más o menos distinto que se diferencia en el componente del tejido conectivo de la dermis. El tejido conectivo dentro de la piel suele ser difuso e irregular, aunque en algunas especies los haces de colágeno están dispuestos en una capa distinta y ordenada dentro de la dermis. Esta capa se denomina estrato compacto (Figura 14.1b).

Figura 14.2. Especializaciones del tegumento. Los receptores sensoriales residen en la piel. Las glándulas exocrinas con conductos y las glándulas endocrinas sin conductos se forman a partir de las invaginaciones de la epidermis. A través de una interacción dérmica-epidérmica, surgen estructuras cutáneas especializadas como el cabello, las plumas y los dientes.

Figura 14.3. Derivados
de la piel. (a) A partir de la simple disposición de la epidermis y la
dermis, con una membrana basal entre ellas, se desarrolla una gran variedad de
integumentos vertebrados. La interacción de la epidermis y la dermis da lugar a
plumas en las aves (b), el pelo y las glándulas mamarias en los mamíferos (c y
d), los dientes en los vertebrados (e), las escamas placoides en los
condrictios (f), y las escamas cosmoides, ganoides y cicloidctenoides. en peces
óseos (g – i)
Las células del origen de la cresta neural migran hacia la
región entre la dermis y la epidermis, contribuyendo a la armadura ósea y a las
células pigmentarias de la piel llamadas cromatóforos (que significa
"color" y "transportar"). Normalmente los cromatóforos
residen en la dermis, aunque en algunas especies pueden enviar pseudópodos a la
epidermis o residir allí ellos mismos. A menudo, los cromatóforos se dispersan
dentro de la hipodermis. Los nervios y los vasos sanguíneos invaden el
tegumento para completar su composición estructural. Fundamentalmente, el
tegumento se compone de dos capas, epidermis y dermis, separadas por la
membrana basal. Se agregan la vascularización y la inervación, junto con las
contribuciones de la cresta neural. A partir de tales ingredientes
estructurales simples, surge una gran variedad de derivados tegumentarios. El
tegumento contiene órganos sensoriales que detectan los estímulos que llegan
del entorno externo. La invaginación de la epidermis superficial forma
glándulas cutáneas: exocrinas si retienen conductos, y endocrinas si se separan
de la superficie y liberan los productos directamente en los vasos sanguíneos (Figura
14.2). La interacción
entre la epidermis y la dermis estimula especializaciones como dientes, plumas,
cabello y escamas de varias variedades (Figura
14.3).
14.2 Características
generales de la dermis
La dermis de muchos vertebrados produce placas de hueso directamente a través de la osificación intramembranosa. Debido a su origen embrionario y su posición inicial dentro de la dermis, estos huesos se llaman huesos dérmicos. Son prominentes en los peces ostracodermos y placodermos donde conjuntamente reciben el nombre de dermatoesqueleto y/o dermatocráneo, pero aparecen secundariamente incluso en grupos derivados, como en algunas especies de mamíferos, reptiles y dinosaurios. El componente más conspicuo de la dermis es el tejido fibroso conectivo compuesto principalmente de fibras de colágeno. Las fibras de colágeno pueden tejerse en distintas capas llamadas capas. La dermis del amfioxus un protocordado que ha retenido caracteres ancestrales, presenta una disposición especialmente ordenada de colágeno dentro de cada capa (Figura 14.4).

Figura 14.4. Protocordato, piel de anfioxo. La epidermis
es una capa única de células cúbicas o columnares que secretan una mucosidad
que recubre la superficie y descansa sobre una lámina basal. La dermis consiste
en fibras de colágeno muy ordenadas dispuestas en capas alternas para formar
una "tela" que trae soporte estructural y flexibilidad a la pared
exterior del cuerpo. El pigmento es secretado por las propias células
epidérmicas.
A su vez, las capas se laminan juntas en una orientación muy regular, pero alternando. Estas capas alternas actúan como un patrón de hilos en un telar, dando cierta forma a la piel y evitando que se deshaga. En los vertebrados acuáticos, como los tiburones, los haces de colágeno se encuentran en ángulos entre sí, dando a la piel un sesgo, como el paño; es decir, la piel se estira cuando se tira de un ángulo oblicuo a la dirección de los haces.

Figura 14.5. Sesgo en material tejido. (a) La urdimbre
(hilos longitudinales) y la trama (hilos cruzados) componen las fibras de la
tela. Si la fuerza de tracción es paralela a los hilos (indicados por las
flechas), se produce una pequeña distorsión de la tela. (b) Sin embargo, la
tensión a lo largo del sesgo a 45 ° de los hilos da como resultado un cambio
sustancial en la forma. (c) Los diseñadores de moda aprovechan estas
características de la tela cuando diseñan ropa. En la dirección de polarización
suelta, la tela cae en pliegues, pero puede mantener su forma a lo largo de los
hilos de urdimbre y trama. (d) Las capas de colágeno del estrato compacto de la
piel del pez actúan de manera similar. El sesgo flexible de la piel se orienta
a 45 ° de la longitud del cuerpo, acomodando así la flexión lateral durante la
natación. Esta disposición mantiene la piel flexible pero tensa para que no se
produzcan arrugas en la superficie y no se induzca turbulencia en las líneas de
corriente que pasan sobre el cuerpo a medida que el pez nada, especialmente en
los tiburones.
Por ejemplo, si
usted toma un pedazo de tela, como un pañuelo, y lo tira a lo largo de hilos de
urdimbre o de trama, el paño se extiende muy poco bajo esta tensión paralela.
Pero si se tira en las esquinas opuestas, la tensión se aplica oblicuamente en
un ángulo de 45 ° con respecto a las roscas, y la tela se estira considerablemente
(Figura
14.5 a, b). Este principio parece gobernar el
estrechamente tejido de colágeno de la piel de tiburón. Su inclinación flexible
de la piel acomoda la flexión lateral del cuerpo, pero resiste simultáneamente
a las distorsiones en la forma del cuerpo. Como resultado, la piel se estira
sin arrugas. Debido a que no se arruga, el agua fluye suavemente y sin
turbulencias a través de la superficie del cuerpo (Figura
14.5d).
En los peces y
vertebrados acuáticos, incluidos los cetáceos y los lagartos acuáticos, las
fibras de colágeno de la dermis se disponen generalmente en capas ordenadas que
forman un estrato compacto reconocible. En los vertebrados terrestres, el
estrato compacto es menos obvio porque la locomoción en tierra depende más de
los miembros y menos del tronco. Y, por supuesto, cualquier arruga de la piel
es menos perjudicial para un vertebrado terrestre que se mueve a través del
aire, ya que es un medio que genera menos rozamiento de fluidos. En
consecuencia, las fibras de colágeno están presentes, incluso abundantes, en la
piel de los vertebrados terrestres, pero son mucho menos regularmente ordenadas
y generalmente no forman capas distintas.
14.3 Características
generales de la epidermis
La epidermis de
muchos vertebrados produce moco para humedecer la superficie de la piel. En los
peces, el moco parece proporcionar alguna protección contra las infecciones
bacterianas y ayuda a asegurar el flujo laminar de agua a través de la
superficie del cuerpo. En los anfibios, el moco probablemente cumple funciones
similares y, además, evita que la piel se seque durante las estancias del
animal en tierra. En los vertebrados terrestres amniotas, la epidermis que
cubre el cuerpo a menudo forma una capa externa queratinizada o cornificada,
llamada estrato córneo. Es una de las innovaciones de los tetrápodos amniotas
que les ayudan a abordar la vida en un ambiente terrestre seco y abrasivo.
Todas las células
del stratum corrneum son células muertas. Las nuevas células epidérmicas
se forman por división mitótica, principalmente en el estrato basal profundo.
Estas nuevas células epidérmicas empujan a las que están por encima de ellas
hasta que llegan hacia la superficie, donde tienden a autodestruirse de manera
ordenada. Durante su desaparición, varios productos proteínicos se acumulan y
forman colectivamente queratina en un proceso llamado queratinización. El estrato
superficial resultante es una capa no viva que sirve para reducir la pérdida de
agua a través de la piel en ambientes terrestres secos. Las células epidérmicas
que participan son queratinocitos. En los sauropsidos, la epidermis produce dos
tipos de queratinocitos -un tipo que contiene queratina alfa (blanda) y el
otro, queratina beta (dura). Las alfa-queratinas están presentes en la mayoría
de las capas epidérmicas flexibles donde se producen cambios de forma. Las
beta-queratinas son más comunes en especializaciones como escamas duras,
garras, pico y plumas. En los sinapsidos, sólo están presentes las
alfa-queratinas.
La queratinización y la formación de un estrato córneo también se producen cuando la fricción o la abrasión mecánica directa insultan el epitelio. Por ejemplo, la epidermis en la cavidad oral de los vertebrados acuáticos y terrestres a menudo exhibe una capa queratinizada, especialmente si el alimento que se come es inusualmente agudo o abrasivo. En las zonas del cuerpo donde la fricción es común, como las plantas de los pies o las palmas de las manos, la capa cornificada puede formar una gruesa capa protectora, o callo, para evitar daños mecánicos (Figura 14.6).

Figura 14.6. Queratinización,
formación de callo. En lugares donde aumenta la fricción mecánica, el
integumento responde aumentando la producción de un callo queratinizado
protector, y como resultado el estrato córneo se engrosa. Clave: E, epidermis;
D, dermis.
El estrato córneo
puede ser diferenciado en pelo, cascos, vainas de cuerno, u otras estructuras
cornificadas especializadas. El término sistema queratinizante se refiere a la
elaborada interacción de la epidermis y la dermis que produce la transformación
ordenada de los queratinocitos en tales estructuras cornificadas. Finalmente,
las escamas forman dentro del tegumento de muchos vertebrados acuáticos y
terrestres. Las escalas son básicamente pliegues en el integumento. Si
predominan las aportaciones dérmicas, especialmente en forma de hueso dérmico
osificado, el pliegue se denomina escala dérmica. Un pliegue epidérmico,
especialmente en forma de una capa queratinizada engrosada, produce una escala
epidérmica.
15. La piel de los peces
|| Regresar al Índice ||
Con pocas excepciones, la piel de la mayoría de los peces vivos no es queratinizada y en su lugar se encuentra cubierta por moco. Las excepciones incluyen especializaciones queratinizadas en algunos grupos como: Los "dientes" que recubren el disco oral de las lampreas, los revestimientos mandibulares de algunos peces minúsculos herbívoros y la superficie de fricción en la piel del vientre de algunos peces semiterrestres son todos derivados queratinizados. Sin embargo, en la mayoría de los peces vivos, la epidermis está viva y activa en la superficie del cuerpo, y no hay ninguna capa superficial prominente de células muertas, queratinizadas. Las células superficiales suelen estar modeladas con pequeños microrredios que tal vez contienen la capa superficial de moco. La capa mucosa se forma a partir de varias células individuales en la epidermis con contribuciones de glándulas multicelulares. Esta capa mucosa, denominada cutícula mucosa, resiste la penetración de bacterias infecciosas, probablemente contribuye al flujo laminar de agua a través de la superficie, hace que el pez sea resbaloso ante los depredadores y a menudo incluye productos químicos repugnantes, alarmantes o tóxicos para los enemigos. Dos tipos de células ocurren dentro de la epidermis de los peces: células epidérmicas y glándulas unicelulares especializadas. En los peces vivos, incluidos los ciclostomas, las células epidérmicas predominantes constituyen la epidermis estratificada. Las células epidérmicas superficiales están estrechamente conectadas a través de las uniones celulares y contienen numerosas vesículas secretoras que se liberan a la superficie donde contribuyen a la cutícula mucosa. Las células epidérmicas de la capa basal son cuboidales o columnares. La actividad mitótica está presente en la capa basal, pero no se limita a ella.

Figura 15.1. Piel
de lamprea. Entre las numerosas células epidérmicas se encuentran glándulas
unicelulares separadas, las células granulares y las células del club. Tenga en
cuenta la ausencia de queratinización. La dermis consiste en colágeno y
cromatóforos dispuestos regularmente.
Las glándulas unicelulares son únicas, especializadas e
intercaladas entre la población de células epidérmicas. Hay varios tipos de
glándulas unicelulares. La célula bastón “Club cell” es una glándula unicelular
alargada, a veces binucleada (Figura
15.1). Algunos
productos químicos dentro de las células bastón se excitan ante señales de
alarma o miedo. Se cree que son liberados por individuos vigías para advertir a
los demás de un peligro inminente. La célula granular es una célula diversa que
se encuentra en la piel de las lampreas y otros peces (Figura
15.1). Tanto las
células granulares como las bastón contribuyen a la cutícula mucosa, pero sus
otras funciones no se entienden completamente. La célula caliciforme (goblet
cell) es un tipo de glándula unicelular que está ausente de la piel de lamprea,
pero se encuentra generalmente en otros peces óseos y cartilaginosos. También
contribuye a la cutícula mucosa y es reconocida por su forma de tallo basal
estrechado y un extremo apical ancho que contiene secreciones. El microscopio
electrónico ha ayudado a distinguir un tipo adicional de glándula unicelular en
la epidermis, la célula sacciforme. Sostiene un gran producto secretor de
membrana que parece funcionar como un repelente o toxina contra los enemigos
una vez que se libera. A medida que se presta mayor atención al estudio de la
piel de los peces, se están reconociendo otros tipos de células. Esta creciente
lista de células especializadas en la epidermis revela una complejidad y variedad
de funciones que antes no se apreciaban.
El colágeno
dentro del estrato compacto se organiza regularmente en capas que se enrollan
en espiral alrededor del cuerpo del pez, permitiendo que la piel se doble sin
arrugarse. En algunos peces, la dermis tiene propiedades elásticas. Cuando un
pez nada dobla su cuerpo, la piel en el lado estirado almacena energía que
ayuda a desdoblar el cuerpo y barrer la cola en la dirección opuesta. La dermis
del pez a menudo da lugar a hueso dérmico, y el hueso dérmico da lugar a
escamas dérmicas y en linajes primitivos a un esqueleto dérmico. Además, la
superficie de las escamas de los pescados se cubre a veces con un esmalte duro,
acelular del origen epidérmico y una capa más profunda de dentina de origen
dérmico. Hasta hace poco, tanto el esmalte como la dentina eran reconocidos por
su apariencia, no por su composición química. A medida que la apariencia
superficial de las escamas cambiaba entre los grupos de peces, también lo hacía
la terminología. Se pensaba que el esmalte/enamel cedía filogenéticamente a la
"ganoina" y dentina a la"cosmina". Estos términos estaban
inspirados en la apariencia superficial de las escamas, no por su composición
química, ni siquiera por su organización histológica. Tal vez es mejor pensar
en la ganoina como una expresión morfológica diferente del esmalte/enamel, y la
cosmina como una expresión morfológica diferente de la dentina y estar
preparado para las sutiles diferencias químicas a medida que las encontramos.
15.1 Peces
primitivos
En ostracodermos y placodermos, el integumento produjo placas óseas prominentes de la armadura dérmica que encerraba sus cuerpos en un exoesqueleto. Los huesos dérmicos de la región craneal eran grandes, formando los escudos de la cabeza; pero más a posteriori a lo largo del cuerpo, los huesos dérmicos tendían a ser divididos en pedazos más pequeños, las escamas dérmicas. La superficie de estas escamas estaba adornada a menudo con minúsculos tubérculos en forma de setas. Estos tubérculos consistían en una capa superficial de esmalte o una sustancia similar al esmalte sobre una capa interna de dentina (Figura 15.2). Una o varias cavidades de pulpa irradiadas residían dentro de cada tubérculo. El hueso dérmico que sostiene estos tubérculos era lamelar, organizado en un patrón acodado.

Figura 15.2. Sección a través de una escama de
ostracodermo ampliada. La superficie consiste en tubérculos elevados cubiertos con
dentina y esmalte que encierra una cavidad pulpar en su interior. Estos
tubérculos descansan sobre una base de hueso dérmico, parte de la armadura
dérmica que cubre el cuerpo. Recuerde que se trata de una piel-esqueleto, una
armadura defensiva dura y pesada.
La piel de los
mixinos vivos y de las lampreas se aparta considerablemente de la de los
primitivos peces fósiles. El hueso dérmico está ausente, y la superficie de la
piel es lisa y sin escamas. La epidermis se compone de capas apiladas de
numerosas células epidérmicas vivas en todas partes. Entre ellas se encuentran
las glándulas unicelulares, a saber, las grandes células granulares y las
células alargadas de bastón. Además, la piel de mixinos incluye las células del
hilado que descargan las cuerdas gruesas de moco a la superficie de la piel
cuando el pez es irritado. La dermis está altamente organizada en capas
regulares de tejido conectivo fibroso. Las células de pigmento ocurren a través
de la dermis. La hipodermis incluye tejido adiposo. Dentro de la dermis, los
mixinos también poseen glándulas mucosas multicelulares que liberan sus
productos a través de conductos a la superficie.
15.2 Tiburones
y rayas
Aunque parece lo contrario la piel de tiburones y rayas no es lisa, de hecho, al tacto se siente como lija. En los peces cartilaginosos, el hueso dérmico está ausente, pero los dentículos superficiales, denominados escamas placóides, persisten, estas escamas son prácticamente invisibles. Estas escamas son las que dan la sensación áspera a la superficie de la piel (Figura 37 a). La evidencia reciente sugiere que estas diminutas escamas placóides afectan favorablemente al agua que fluye a través de la piel cuando el pez nada hacia delante para reducir el arrastre por fricción. Numerosas células secretoras están presentes en la epidermis, así como células epidérmicas estratificadas. La dermis está compuesta de tejido conectivo fibroso, especialmente elástico y fibras de colágeno, cuya disposición regular forma una urdimbre y una trama en forma de tejido en la dermis. Esto da la fuerza de la piel y previene arrugas durante la natación.

Figura 15.3. Piel de tiburón. (a)
Vista superficial de la piel que muestra la disposición regular de escamas
placoides salientes. (b) Sección a través de una escala placoide de un tiburón.
La escala sobresaliente consiste en esmalte y dentina alrededor de una cavidad
pulpar.
La escama placóide se desarrolla en la dermis, pero se
proyecta a través de la epidermis para alcanzar la superficie. Una tapa de esmalte
forma la punta, la dentina se encuentra debajo, y una cavidad de pulpa reside
dentro. Los cromatóforos ocurren en la parte inferior de la epidermis y en las
regiones superiores de la dermis.
15.3 Peces
óseos
La dermis de los peces óseos se subdivide en una capa superficial de tejido conectivo suelto y una capa más profunda de tejido conectivo fibroso denso.
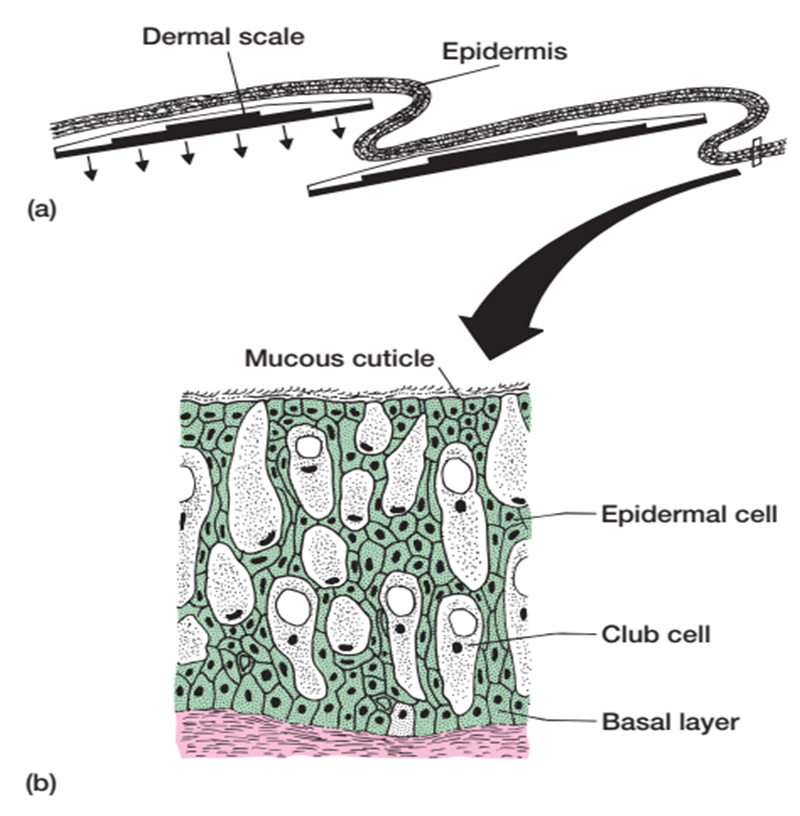
Figura 15.4. Piel de pez óseo. (a) Disposición de escamas dérmicas dentro de la
piel de un pez teleósteo (las flechas indican la dirección del crecimiento de
la escama). (b) Agrandamiento de la epidermis. Tenga en cuenta las células
epidérmicas y las células del club.
Cromatóforos se encuentran dentro de la
dermis. El producto estructural más importante de la dermis es la escama. En
los peces óseos, las escamas dérmicas no perforan la epidermis, pero están tan
cerca de la superficie que dan la impresión de que la piel es dura (Figura 15.4 a,
b).
La cubierta epidérmica incluye una capa
basal de células. Por encima de esta capa se encuentran las células epidérmicas
estratificadas. A medida que se mueven hacia la superficie, las células
epidérmicas experimentan transformación citoplásmica, pero no se vuelven
queratinizadas. Dentro de estas células epidérmicas en capas se producen
glándulas unicelulares individuales, llamadas células secretoras y de bastón.
Estas glándulas unicelulares, junto con las células epidérmicas, son la fuente
de la cutícula mucosa, o "limo" superficial.
Sobre la base de su aparición, varios tipos de escamas se reconocen entre los peces óseos. La escama cosmoide, observada en los sarcopterígios primitivos, reside en una doble capa de hueso, una capa vascular y otra lamelar. En la superficie exterior de este hueso hay una capa que ahora se reconoce generalmente como dentina y sobre esta se extiende superficialmente una capa que ahora se conoce como esmalte/enamel. La apariencia inusual de estos abrigos de esmalte y dentina inspiró en la literatura antigua los respectivos nombres de ganoina y cosmina, por la creencia errónea de que el ganoina era fundamentalmente un mineral diferente del esmalte/enamel y la cosmina como diferente de la dentina. Aunque la naturaleza química de estas capas es ahora clara, los nombres anteriores se han pegado por tradición para darnos los términos para los tipos de escama distintivos. En la escama cosmoide, hay una capa gruesa y bien desarrollada de dentina (cosmina) debajo de una delgada capa de esmalte (Figura 15.5).

Figura 15.5. Sección transversal de una
escama cosmoide.
La escama ganoide se caracteriza por la prevalencia de una gruesa capa superficial de esmalte (ganoina), sin una capa subyacente de dentina (Figura 15.6).

Figura 15.6. Sección transversal de una
escama ganoide.
El hueso dérmico forma la base de la escama ganoide, que aparece como una doble capa de hueso vascular y lamelar (en peces palaeoniscoides) o una sola capa de hueso lamelar (en otros actinopterygios ancestrales). Las escamas ganoides son brillantes (debido al esmalte), se superponen y se entrelazan. Los polipteriformes vivos y las gárgulas conservan las escamas ganoides. Sin embargo, en la mayoría de linajes de peces óseos, las escamas ganoides se reducen a través de la pérdida de la capa vascular del hueso y la pérdida de la superficie del esmalte. Esto produce, en los teleósteos, una escama bastante distintiva. La escama de los teleósteos carece de esmalte, dentina y una capa de hueso vascular. Sólo permanece el hueso lamelar, que es acelular y sobre todo no calcificado (Figura 15.7).

Figura 15.7. Sección transversal de una
escama de teleósteo (c). Vistas de superficie de los dos tipos de escama de
teleósteo, escamas cicloides y ctenoides (d).
Se reconocen dos
tipos de escalas de teleósteos. Una es la escala cicloide, compuesta de anillos
concéntricos, o circuli. La otra es la escala ctenóide, con una franja de
proyecciones a lo largo de su margen posterior (figura anterior d). Nuevos
círculos se establecen, como anillos en un árbol, como un pez teleósteo crece.
Los ciclos anuales son evidentes en los agrupamientos de estos circuli, y de
este patrón en las escalas podemos determinar la edad de los peces individuales.
16. La piel de los anfibios
|| Regresar al Índice ||
Los anfibios son de especial interés porque durante sus vidas usualmente realizan una metamorfosis de una forma acuática a una forma terrestre. En la mayoría de los anfibios modernos, la piel también se especializa como superficie respiratoria a través de la cual se produce el intercambio gaseoso, con los lechos capilares en la epidermis inferior y la dermis más profunda. De hecho, algunas salamandras carecen de pulmones y dependen enteramente de la respiración cutánea a través de la piel para satisfacer sus necesidades metabólicas. Los tetrápodos más primitivos tenían escamas como los peces de los que evolucionaron, lo cual implica que los anfibios modernos son formas derivadas y que no sirven tanto para imaginar cómo eran los anfibios primitivos. Entre los anfibios vivos, las escamas dérmicas están presentes sólo como vestigios en algunas especies de caecilias tropicales (Apoda). Las ranas y las salamandras carecen de rastros de escamas dérmicas (Figura 16.1a).

Figura 16.1. Piel de anfibios. (a) Corte a través de una piel de rana adulta.
Están presentes un estrato basal y un estrato córneo delgado y superficial. La
capa de transición entre ellos incluye un estrato espinoso y un estrato
granuloso. (b) Vista esquemática de la piel de anfibios que muestra glándulas
mucosas y venenosas que vacían sus secreciones a través de conductos cortos
hacia la superficie de la epidermis.
En las
salamandras, la piel de las larvas acuáticas incluye una dermis de tejido
conjuntivo fibroso, que consiste en tejido suelto superficial sobre una capa
profunda compacta. Dentro de la epidermis están las células basales profundas y
las células apicales superficiales. Esparcidas por todo son grandes células de
Leydig que se cree que segregan sustancias que resisten la entrada de bacterias
o virus (Figura 16.1b). En los adultos terrestres, la dermis
se compone de forma similar de tejido conectivo fibroso. En la epidermis, las
células de Leydig están ahora ausentes, pero se pueden reconocer regiones
distintas, como los estratos basales, espinosos, granulosos y corneum. La
presencia de un estrato córneo delgado proporciona cierta protección contra la
abrasión mecánica y retarda la pérdida de humedad del cuerpo sin interrumpir indebidamente
el intercambio de gases cutáneos. Durante la temporada de cría, las
almohadillas nupciales pueden formarse en dígitos o extremidades de salamandras
o ranas macho. Las almohadillas nupciales son calos levantados de la epidermis
cornified que ayudan al varón que sostiene a la hembra durante el acoplamiento.
Generalmente, la
piel de las ranas y salamandras incluye dos tipos de glándulas multicelulares:
glándulas mucosas y venenosas. Ambos se encuentran en la dermis y se abren a la
superficie a través de conductos de conexión (Figura 16.1b). Las glándulas mucosas tienden a ser
más pequeñas, cada una de ellas formada por un pequeño grupo de células que
liberan su producto en un conducto común. Las glándulas venenosas (glándulas
granulares) tienden a ser más grandes y a menudo contienen secreciones
almacenadas dentro del lumen de cada glándula. Las secreciones de las glándulas
venenosas tienden a ser desagradables o incluso tóxicas para los depredadores.
Sin embargo, pocas personas que manipulan anfibios se molestan por esta
secreción, porque es potencialmente dañino sólo si se come o se inyecta en el
torrente sanguíneo, pero no siempre. Los cromatóforos pueden encontrarse
ocasionalmente en la epidermis anfibia, pero la mayoría reside en la dermis.
Los lechos capilares, restringidos a la dermis en la mayoría de los
vertebrados, llegan a la parte inferior de la epidermis en los anfibios, una
característica que sirve de respiración cutánea.
La piel de la mayoría de los anfibios contiene glándulas que
segregan productos que son desagradables o incluso tóxicos para los
depredadores. A menudo estas toxinas cutáneas no son fabricadas por las propias
ranas, sino que son cooptadas de toxinas en sus presas, como las hormigas u
otros artrópodos. Algunos especialistas de hormigas sugieren que los insectos,
a su vez, pueden obtener las toxinas o los precursores de toxinas de su dieta.
En las regiones tropicales del Nuevo Mundo vive un grupo de ranas, las ranas de
dardo venenoso, con secreciones de piel especialmente tóxicas. Los pueblos
nativos de la región a menudo recogen estas ranas, las mantienen en palos sobre
un fuego para estimular la liberación de estas secreciones, y luego recoger las
secreciones en las puntas de sus dardos.
Una de las ranas dardo más peligrosas es la rana dardo dorada, Phyllobates terribilis endémica de la costa pacífica colombiana. La piel de la rana dardo dorada está densamente revestida de una toxina alcaloide, uno de los venenos comunes a las ranas de dardo (batrachotoxinas). Este veneno impide que los nervios de la víctima transmitan impulsos, dejando los músculos en un estado inactivo de contracción, que puede conducir a insuficiencia cardíaca o fibrilación. Las batrachotoxinas alcaloideas pueden ser almacenadas por las ranas durante años después de que la rana se vea privada de una fuente alimentaria, y esas toxinas no se deterioran fácilmente, incluso cuando se transfieren a otra superficie.

Figura 16.2. Parafraseando
al diablo en la película “El Abogado del Diablo”: mira… pero no toques (YouTube).
La dosis media transportada variará entre los lugares, y la
consiguiente dieta local, pero para la P.
terribilis salvaje promedio generalmente se estima que contiene
aproximadamente un miligramo de veneno, suficiente para matar a unos 10.000
ratones. Esta estimación variará a su vez, pero la mayoría está de acuerdo en
que esta dosis es suficiente para matar entre 10 y 20 humanos, lo que se
correlaciona con hasta dos elefantes africanos. Este veneno extraordinariamente letal es muy
raro. La Batrachotoxina sólo se encuentra en tres ranas venenosas de Colombia
(género Phyllobates), tres aves
venenosas de Papua Nueva Guinea; Pitohui
dichrous, Pitohui kirhocephalus e
Ifrita kowaldi y cuatro escarabajos
papúes del género Choresine en la
familia Melyridae; C. pulchra, C. semiopaca, y C. rugiceps.
Otras toxinas relacionadas, histrionicotoxina y pumiliotoxina, se encuentran en
especies de ranas del género Dendrobates.
La rana dardo dorada, como la mayoría de las otras ranas venenosas, almacena su
veneno en las glándulas de la piel. Generalmente debido a su veneno, las ranas
saben mal a los depredadores; El veneno de P.
terribilis no solo da mal sabor, este, mata a quien lo pruebe,
excepto por una especie de serpiente, Liophis
epinephelus. Esta serpiente es resistente al veneno de la rana, pero no es
completamente inmune. Las propias ranas venenosas son quizás las únicas
criaturas que son inmunes a este veneno. La batrachotoxina ataca los canales de
sodio de las células nerviosas, pero la rana tiene canales especiales de sodio que
el veneno no puede dañar.
Dado que los
alimentos que se compran fácilmente en los laboratorios no son ricos en
alcaloides necesarios para producir batracotoxinas, las ranas cautivas no
producen toxinas y eventualmente pierden su toxicidad en cautiverio. De hecho,
muchos aficionados y herpetólogos han informado de que la mayoría de las ranas
de dardos no consumen hormigas en cautiverio, aunque las hormigas constituyen
la mayor parte de sus dietas en la naturaleza.
Aunque todas las ranas venenosas pierden su toxicidad cuando se les
priva de ciertos alimentos, y las ranas dardo doradas criadas en cautividad
nacen inofensivas, una rana venenosa capturada puede retener alcaloides durante
años. No está claro qué tipo de presa suministra el potente alcaloide que da a
las ranas dardo doradas sus niveles excepcionalmente altos de toxicidad, o si
las ranas modifican otra toxina disponible para producir una variante más
eficiente, como lo hacen algunas de las ranas del género Dendrobates. Por lo tanto, la alta toxicidad de P. terribilis parece ser debido al
consumo de pequeños insectos u otros artrópodos, y uno de ellos puede ser
verdaderamente la criatura más venenosa en la Tierra. Los científicos han
sugerido que el insecto crucial puede ser un pequeño escarabajo de la familia Melyridae. Al menos una especie de estos
escarabajos produce la misma toxina que se encuentra en P. terribilis. Sus parientes en las selvas tropicales colombianas
podrían ser la fuente de las batrachotoxinas encontradas en las ranas Phyllobates altamente tóxicas de esa
región.
17. La piel de los reptiles
|| Regresar al Índice ||
Aunque la queratinización ocurre en los peces, entre los vertebrados terrestres se convierte en una característica principal del tegumento. La queratinización extensa produce una capa cornificada externa prominente, el estrato córneo, que resiste la abrasión mecánica. Los lípidos a menudo se añaden durante el proceso de queratinización o se extienden a través de la superficie de las glándulas especializadas. La capa cornificada junto con estos lípidos aumenta la resistencia de la piel del tetrápodo a la desecación. Las glándulas multicelulares son más comunes en la piel de los tetrápodos que en la piel de los peces. En los peces, la cutícula mucosa y las secreciones de las glándulas unicelulares en o cerca de la superficie de la piel lo cubren. En contraste, entre los tetrápodos, las glándulas multicelulares habitualmente residen en la dermis y llegan a la superficie a través de conductos comunes que perforan la capa cornificada. Así, el estrato córneo que protege la piel y evita la desecación también controla la liberación de secreciones directamente a la superficie. Si no fuera por estas aberturas en el estrato córneo, la superficie de la piel no podría ser recubierta o lubricada por estas secreciones.

Figura 17.1. Piel de reptil. (a) Escamas epidérmicas. El alcance de la
proyección y la superposición de las escamas epidérmicas varía entre los
reptiles e incluso a lo largo del cuerpo del mismo individuo. Se ilustran
escamas del cuerpo de la serpiente (arriba) y escamas tuberculares de muchos
lagartos (abajo). Entre las escamas hay un área adelgazada de la epidermis, una
"bisagra" que permite la flexibilidad de la piel. (b) desprendimiento
de la piel. Justo antes de que la vieja capa externa de la epidermis se
desprenda, las células basales producen una generación epidérmica interna. Los
glóbulos blancos se acumulan en la zona de división para promover la separación
de la epidermis externa nueva de la vieja, y para evitar infecciones durante el
proceso.
La piel de los reptiles refleja su mayor compromiso con una
existencia terrestre. La queratinización es mucho más extensa, y las glándulas
de la piel son menos comunes que en los anfibios. Las escamas están presentes,
pero éstas son fundamentalmente diferentes de las escamas dérmicas de los
peces, que se construyen alrededor del hueso de origen dérmico. La escala
reptiliana generalmente carece del soporte óseo o de cualquier contribución
estructural significativa de la dermis. En cambio, es un pliegue en la
epidermis superficial, por lo tanto, una escama epidérmica. La unión entre las
escamas epidérmicas adyacentes es una bisagra flexible (Figura 17.1a). Si la escama
epidérmica es grande y similar a una placa, a veces se la denomina sutura.
Adicionalmente, las escamas epidérmicas pueden ser modificadas en crestas,
espinas o procesos similares a cuernos.
17.1 Las
gastralias
Aunque no suele asociarse con las escamas, el hueso dérmico está presente en muchos reptiles. Las gastralia, una colección de huesos en el área abdominal, son ejemplos. Cuando los huesos dérmicos sostienen la epidermis, se les llama osteodermos, placas de hueso dérmico situadas bajo las escamas epidérmicas. Los osteodermos se encuentran en cocodrilos, algunos lagartos y algunos reptiles extintos. Algunos huesos de la concha de la tortuga son probablemente osteodermos modificados. La dermis de piel reptiliana está compuesta de tejido conectivo fibroso. La epidermis se delinea generalmente en tres regiones: estrato basal, estrato granuloso y estrato córneo. Sin embargo, esto cambia antes de la muda en los reptiles que cambian grandes piezas de la capa de piel cornificada.

Figura 17.2. Las gastralias
son estructuras óseas de muchos reptiles y dinosaurios de origen
integumentario, vulgarmente se las llama costillas ventrales, y sirven para
proteger los órganos del vientre como el estómago.
En las tortugas y
los cocodrilos, el desprendimiento de la piel es modesto, comparable a las aves
y los mamíferos, en los que los pequeños copos caen a intervalos irregulares.
Pero en los lagartos, y especialmente en las serpientes, el desprendimiento de
la capa cornificada, se denomina muda o ecdisis, y da lugar a la eliminación de
extensas secciones de la epidermis superficial. A medida que comienza la muda,
el estrato basal, que ha dado origen a los estratos granuloso (interior) y
corneo (exterior), duplica las capas más profundas de granuloso y corneo,
empujando por debajo de las capas antiguas. Los glóbulos blancos invaden el
estrato intermedio, una capa temporal entre la piel vieja y la nueva (Figura 17.1b). Se cree que estos glóbulos blancos
promueven la separación y pérdida de la capa superficial vieja de la piel.
Las glándulas
integumentales de los reptiles suelen estar restringidas a ciertas áreas del
cuerpo. Muchos lagartos poseen filas de glándulas femorales a lo largo de la
parte inferior del miembro posterior en la región del muslo. Los cocodrilos y
algunas tortugas tienen glándulas olfativas. En caimanes de ambos sexos, un par
de glándulas olfativas se abre en la cloaca, otro par se abre en los márgenes
de la mandíbula inferior. En algunas tortugas, las glándulas olfativas pueden
producir olores muy picantes, especialmente cuando el animal está alarmado por
la manipulación. Se piensa que la mayoría de las glándulas integumentales de
reptiles desempeñan un papel en el comportamiento reproductivo o para
desalentar a los depredadores, pero las glándulas y sus papeles sociales no se
comprenden bien.
17.2 Las
garras
Una garra es un apéndice curvado y puntiagudo, que se encuentra al final de los dedos en la mayoría de los amniotas (mamíferos, reptiles, pájaros). Una verdadera garra está hecha de proteína dura llamada queratina. Las garras se usan para atrapar y retener presas en mamíferos carnívoros como gatos y perros, pero también pueden usarse para propósitos tales como excavar, trepar a los árboles, autodefensa y arreglarse, en esas y otras especies. Apéndices similares que son planos y no llegan a un punto agudo se llaman uñas. Las proyecciones parecidas a garras que no se forman al final de los dígitos, se denominan espuelas. En los tetrápodos, las garras están hechas de queratina y constan de dos capas. El unguis es la capa externa más dura, que consiste en fibras queratínicas dispuestas perpendicularmente a la dirección del crecimiento y en capas en un ángulo oblicuo. El subunguis es la capa inferior más suave y escamosa cuyo grano es paralelo a la dirección del crecimiento.

Figura 17.3. Las garras y
las uñas son estructuras análogas, pero anatómicamente muy distintas, la garra
es una estructura tridimensional, sólida y muy dura, con movimiento
independiente por medio de un tendón asociado, cortar una garra es algo
irreversible y muy doloroso.
La garra crece hacia fuera de la matriz en la base de la unguis
y el subunguis crece más gruesa mientras viaja a través de la matriz. El unguis
crece hacia fuera más rápidamente que el subunguis para producir una curva y
los lados más finos de la garra se desgastan más rápidamente que su centro más
grueso, produciendo un punto más o menos agudo. Los tetrápodos usan sus garras
de muchas maneras, comúnmente para agarrar o matar presas, para cavar y para
trepar y para colgar. La mayoría de los reptiles tienen garras bien
desarrolladas. La mayoría de los lagartos tienen puntas que terminan en gruesas
garras, formando desde la última escama en el dedo del pie. En las serpientes,
los pies y las garras están ausentes, pero en muchos boas, restos de
extremidades traseras muy reducidas emergen con una sola garra como "espuelas"
a cada lado de la abertura anal. Las garras del lagarto se utilizan como ayudas
en la escalada, y en la retención de presas en especies carnívoras.
17.3 La concha
de las tortugas
La concha de las tortugas es un derivado del tegumento y del esqueleto. El aspecto de la tortuga se puede identificar fácilmente por su concha, esta desempeña un papel vital en la protección. No ha cambiado mucho en más de 200 millones de años de evolución. La forma y la función de la concha de tortuga sigue siendo la misma. Se divide en dos partes. Parte superior se llama el caparazón y el plastrón inferior. En la parte lateral de la tortuga, el plastrón y el caparazón se unen con estructuras óseas llamadas puentes. La capa interna de la cáscara está hecha de huesos. Incluyendo la columna vertebral y las costillas fusionadas. Eso básicamente significa que la tortuga no puede salir de la cáscara como erróneamente puede llegar a creerse. La cáscara de la tortuga se cubre con las escmas óseas (también llamadas escudos). Las escamas están hechas de queratina y por lo tanto son un derivado del sistema tegumentario. La queratina es una sustancia que se encuentra en el cabello y uñas de otros animales. La melanina en los escudos es la razón de diversos pigmentos y diferentes diseños.
Figura 17.4. Las costillas y
gastralias en las tortugas se expanden, fusionan y migran al exterior formando
la concha, recuerde que estas estructuras originalmente evolucionaron y se
originan de la epidermis.
Pero no todas las tortugas tienen escamas
córneas. Las tortugas laúd y la cáscara de tortuga de caparazón blando están
cubiertas con piel coriácea. La forma, el soporte y la protección de la cáscara
son proporcionados por la capa ósea debajo de la capa externa tegumentaria.
Cuando las tortugas eclosionan, sus placas no están conectadas de ninguna
manera. Pero con el tiempo las plancas crecerán lentamente y se
fusionarán. Las tortugas acuáticas
tienen conchas ligeras adaptadas para la natación. Las conchas de tortugas de
agua también contienen grandes espacios entre los huesos de la concha. También
se llama fontanelas. Las Fontanelas además disminuyen el peso de la concha de
tortuga. Las tortugas terrestres tienen conchas pesadas para una mayor
protección.
17.4 La
armadura de los cocodrilos
El exoesqueleto del cocodrilo consiste en los componentes dermales y epidérmicos protectores del sistema tegumentario en animales del orden Crocodilia. Es una forma de armadura. El exoesqueleto epidérmico del caimán y parientes consiste en escamas oblongas córneas, dispuestas en hileras transversales; los ejes largos de las escamas son paralelos a la del cuerpo. En la cola, excepto a lo largo de la línea media-dorsal, y en el lado ventral del tronco y la cabeza de estas escalas son muy regulares en el contorno y la disposición, a los lados de la cabeza y del tronco y en las piernas son mucho más pequeños y menos regularmente dispuestos, mientras que, a lo largo de la línea media-dorsal de la cola, especialmente en su mitad posterior, se elevan en altas quillas que dan a la cola una gran superficie para la natación. Los tres primeros dígitos del manus (pie delantero) y pes (pie posterior) están armados con garras córneas, que también pertenecen a la parte epidérmica del exoesqueleto.

Figura 17.5. Esqueleto interno
y escamas de un cocodrilo, la piel es tan dura que regularmente se ha
encontrado como a prueba de balas excepto en una posición en la nuca.
El exoesqueleto dérmico consiste en
escudos óseos que subyacen a las escamas epidérmicas de la superficie dorsal
del tronco y la parte anterior de la cola. Las escamas que lo recubren, excepto
en animales muy jóvenes, siempre se frotan, de modo que las escamas óseas
quedan expuestas. La superficie ventral o interna de los escudos es plana,
mientras que la superficie exterior está fuertemente cubierta y en animales
viejos a menudo es áspera y deshuesada. Las placas son casi cuadradas en
contorno y están estrechamente unidas en la mayoría de los lugares. Los escudos
se agrupan en dos áreas bastante distintas, conocidas como escudos nucal y
dorsal. El primero se encuentra justo detrás de la cabeza, en la región de las
patas delanteras, y consta de cuatro placas más grandes y un número de placas
más pequeñas. Este último, o escudo dorsal, se extiende sobre la espalda en
filas longitudinales bastante regulares y filas transversales bastante
regulares. En la parte más ancha del tronco hay seis u ocho de estos escudos en
una fila transversal. Se hacen más pequeños hacia la cola. Los dientes son
estructuras exoesqueléticas, en parte ectodérmicas, en parte de origen dérmico.
Son de forma cónica, sin raíces, y se reemplazan cuando se pierden.
La armadura de los cocodrilos hace difícil
cazarlos, además dio origen a la leyenda de la piel super dura de los dragones
en casi la totalidad de obras de fantasía en la cultura occidental. Sin
embargo, su armadura no es perfecta, justo detrás del cráneo y antes de la
armadura de la nuca existe una región que no está protegida, y es allí donde
los cazadores tratan de disparar cuando cazan a estos animales.
18. Generalidades de las plumas
18.1 Escamas y
plumas, una homología compleja
Las plumas de las aves a veces se denominan como elaboradas escamas de reptil. Esto simplifica demasiado la homología. Ciertamente, la presencia de escamas epidérmicas a lo largo de las patas y los pies (Figura 18.1a) de las aves demuestra su deuda con los reptiles.

Figura 18.1. Escamas de pájaros. (a) Las
escamas epidérmicas están presentes en los pies y las patas de las aves. (b)
Sección de piel que muestra el estrato basal y la capa superficial
queratinizada, el estrato córneo. Las células que salen de la capa basal se
mueven primero a través del estrato intermedio y la capa de transición antes de
llegar a la superficie. Estas capas intermedias son equivalentes a las capas de
espinosa y granulosa de los mamíferos.
Si no es una escama reptiliana remodelada directamente, entonces la pluma es un ejemplo de otra homología más fundamental de la interacción subyacente de las capas epidérmicas-dérmicas que producen tal especialización de la piel. La dermis de la piel de las aves, especialmente cerca de los folículos de las plumas, está rica en vasos sanguíneos, terminaciones nerviosas sensoriales y músculos lisos. Durante la temporada de cría, la dermis en el pecho de algunas aves se vasculariza cada vez más, formando un parche de cría en el que la sangre tibia puede asociarse estrechamente con los huevos incubados. La epidermis comprende el estrato basal y el estrato córneo. Entre ellas se encuentra la capa transicional de células transformadas en la superficie queratinizada del córneo (Figura 18.1b).

Figura 18.2. Tractos de plumas y morfología de plumas.
(a) Las plumas surgen a lo largo de pterilos específicos o tractos de plumas.
(b) Morfología general del contorno de las plumas de vuelo y de plumón.
La piel del pájaro tiene pocas glándulas. La glándula
uropigial, ubicada en la base de la cola (Figura 18.2a), segrega un
producto de lípidos y proteínas que las aves recogen a los lados del pico y
luego se untan las plumas. Acicalarse las plumas con esta secreción,
haciéndolas repelentes al agua, y probablemente condiciona la queratina de la
que están compuestas. Después de una muda, el acicalamiento también ayuda a que
la pluma recién regenerada se despliegue y asuma su forma funcional. La otra
glándula, ubicada en la cabeza de algunas aves, es la glándula de sal, que está
bien desarrollada en aves marinas. Las glándulas de sal excretan el exceso de
sal obtenida cuando estas aves ingieren alimentos marinos y agua de mar.
18.2 Tipos de
plumas
Las plumas distinguen a las aves de todos los demás
vertebrados vivos. Las plumas pueden ser estructuralmente elaboradas y vienen
en una variedad de formas. Sin embargo, las plumas son productos no vasculares
y no nerviosos de la piel, principalmente de la epidermis y del sistema
queratinizante. Se colocan a lo largo de tractos distintivos, denominados pterylae, en la superficie del cuerpo (Figura 18.2a). A través de
una o varias mudas, se reemplazan cada año. En general, la pluma de pájaro
moderno se construye a partir de un eje central tubular, el raquis, que lleva a
cada lado una veleta, una serie de púas con conexiones entrelazadas denominadas
bárbulas (ganchos). El raquis y las paletas adjuntas constituyen la espata (Figura 18.2b). El raquis
continúa proximal como el cálamo o pluma sin púas, que ancla la pluma al cuerpo
y a menudo es movida por los músculos dérmicos unidos. En las aves modernas, las
plumas son de muchos tipos y cumplen diferentes funciones (Figura 18.2c). Las plumas de
vuelo son largas y las paletas asimétricas alrededor del rígido raquis; esas
plumas de vuelo en las alas son remiges (sing, remix) y aquellas en las
rectrices de la cola (sing, rectrix). Las plumas de contorno, o plumas
pennaceas, cubren el cuerpo y generalmente tienen paletas simétricas alrededor
de un raquis. Las plumas de plumón o plumulaceas carecen de un raquis
distintivo y las púas no entrelazadas se extienden desde el calamo como una
pluma esponjosa importante en el aislamiento (Figura 18.2b).
Hay varios tipos de plumas (Figura 18.3). Las plumas de contorno dan forma aerodinámica a la superficie del ave. Las plumas de plumón se encuentran cerca de la piel como aislamiento térmico. Los filoplumes a menudo se especializan para la exhibición, y las plumas de vuelo constituyen las principales superficies aerodinámicas. Las plumas de vuelo de las alas son un tipo de pluma de contorno. Se caracterizan por un largo raquis y paletas prominentes. Estas plumas tienen cierto valor como aislamiento, pero su función principal es la locomoción. La mayoría de las plumas reciben estímulos sensoriales y llevan colores para su exhibición o cortejo. Los cromatóforos se producen dentro de la epidermis, y sus pigmentos se llevan a las plumas para darles color. Pero la refracción de la luz en las púas y las bárbulas de plumas también crea algunos de los colores iridiscentes que muestran las plumas.

Figura 18.3. Tipos de plumas. Las plumas de
vuelo constituyen las principales superficies locomotoras. Las plumas de
contorno en el cuerpo dan forma aerodinámica a la superficie de un pájaro. Los
filoplumas suelen estar especializados para su visualización. Las plumas de
plumón se encuentran cerca de la piel como aislamiento térmico.
18.3 Desarrollo
de las plumas
Las plumas se desarrollan embriológicamente a partir de folículos de plumas, invaginaciones de la epidermis que se sumergen en la dermis subyacente. La raíz del folículo de la pluma, en asociación con la cavidad de la pulpa dérmica, comienza a formar la pluma.

Figura 18.4. Crecimiento de plumas.
Secuencia de muda y desarrollo del reemplazo de plumas. (a) La pluma vieja se
desprende (muda), y un nuevo filamento de plumas pronto crece fuera del
folículo como resultado de la proliferación celular en su base. (b, c) Etapas
sucesivas en el desarrollo. Tenga en cuenta que algunos tejidos necesarios para
el desarrollo inicial (vaina, tapas de pulpa, ahora pierden esta función y se
desprenden a medida que emerge la pluma madura. (D) Pluma nueva y madura en su
lugar.
La pluma vieja se desprende (muda), y el comienzo de una
nueva pluma, el filamento de plumas (o alfiler o pluma de sangre), pronto crece
fuera del folículo como consecuencia de la proliferación celular en la base
folicular (Figura 18.4a). Las nuevas
células epidérmicas forman tres tejidos distintos: una vaina de soporte, pero
luego desechable alrededor de la pluma en crecimiento; los principales tejidos
de plumas que luego se despliegan para asumir su forma final y funcional; y
tapas pulpares que protegen temporalmente el delicado núcleo dérmico. A medida
que el espato en crecimiento comienza a desplegarse (Figura 18.4b), se forman
nuevas tapas protectoras una debajo de la otra a medida que las tapas de pulpa
más viejas se desprenden junto con las partes superiores de la vaina que se
caen durante el acicalamiento. El crecimiento adicional alarga la pluma a
medida que su espata continúa desplegándose (Figura 18.4c). Cuando se
completa el desarrollo de la espata, el desarrollo del calamo comienza dentro
de la misma vaina en la base de la pluma. La pluma completamente formada,
abrazada sobre su base por el folículo de la pluma, ahora está en su lugar (Figura 18.4d).
18.4 Funciones
de las plumas
Ya sea que comiencen con una ectoterma o una endoterma, muchos aún sugieren que las plumas jugaron un papel en el aislamiento de la superficie cuando aparecieron por primera vez en los dinosaurios y en segundo lugar fueron cooptados para un papel en vuelo. Sin embargo, las primeras aves y sus ancestros dinosaurios inmediatos carecían de cornetes nasales, un diagnóstico de carácter de fisiología de sangre caliente. Si sirviera como aislamiento, estas primeras plumas habrían tenido un papel más complicado de lo que se pensaba anteriormente.

Figura 18.5. Función de vuelo de veleta de
plumas asimétrica. (a) El ala está extendida, ya que podría aparecer durante la
mitad del golpe descendente de potencia. Se quita una de las plumas primarias
(resaltada) para mostrar el eje de rotación alrededor de su calamo, donde se
adhiere a la extremidad. (b, c) Secciones transversales a través de tres plumas
de vuelo durante el golpe descendente (b) y el golpe ascendente (c). Durante el
golpe descendente de potencia, cada pluma experimentaría presión de aire contra
la parte inferior del ala a lo largo de su centro de presión en la línea media
anatómica de la pluma. Sin embargo, debido a que el raquis está desplazado,
este centro de presión obliga a la pluma a rotar alrededor de su eje, y las
plumas primarias forman temporalmente una superficie cerrada y uniforme. Durante
el golpe ascendente de recuperación, la presión del aire contra la parte
posterior del ala fuerza la rotación en la dirección opuesta, los espacios
abiertos entre las plumas y el aire se desliza entre las ranuras resultantes,
reduciendo así la resistencia a la recuperación del ala.
Una visión diferente se deriva completamente del argumento de
que las plumas evolucionaron inicialmente como ayudas al planeo “como la
membrana de pterosaurios y murciélagos” y luego al vuelo. Las plumas se
seleccionaron debido a su efecto favorable en la corriente de aire que pasa
sobre el cuerpo o las extremidades de un animal que se aerodesliza. Si la
extremidad protoaviana no estuviera aerodinámica organizada, se produciría un
arrastre de presión y la turbulencia habría reducido la eficiencia
aerodinámica. Sin embargo, las escamas superficiales que se proyectan desde el
borde posterior de la extremidad habrían simplificado la extremidad, reducido
el arrastre y, por lo tanto, favorecido por la selección.
Independientemente de si evolucionaron primero para el aerodeslizamiento o para el aislamiento, las plumas se modificaron a partir de escamas de reptil o al menos a partir de la interacción inductiva común entre la dermis y la epidermis. En las aves modernas, las plumas que sirven para volar están muy modificadas. Las púas y las bárbulas entrelazadas dan cierta integridad estructural a la pluma de vuelo flexible. En las plumas de vuelo, el raquis está desplazado, lo que hace que la paleta sea asimétrica (Figura 18.5). Este diseño afecta la acción de la pluma de vuelo durante los golpes de las alas. En el golpe descendente, la presión en la parte inferior de cada pluma actúa a lo largo de su línea media anatómica, el centro de presión. Pero debido a que el raquis está desplazado, el resultado es torcer la pluma ligeramente alrededor de su punto de unión a la extremidad, forzando a las plumas del ala a formar una superficie amplia que presiona contra el aire y empuja al ave hacia adelante (Figura 18.5b). En el golpe ascendente, el centro de presión está ahora en la parte superior de la pluma asimétrica y la obliga a girar en la dirección opuesta, abriendo un canal entre las plumas (Figura 18.5c). Esto reduce su resistencia a la corriente de aire y permite que el ala se recupere y se prepare para el siguiente golpe descendente de potencia.

Figura 18.6. Pluma de Archaeopteryx. Esta pluma del ala de Archaeopteryx
muestra el diseño asimétrico de la veleta “barbas más cortas de un lado”, lo
que sugiere que puede haber sido utilizada durante el vuelo o aerodislizamiento
como en las aves modernas.
Esta torsión controlada de las plumas de vuelo que responden
pasivamente al latido del ala depende del diseño asimétrico de la pluma y, por
lo tanto, de la acción de la presión del aire contra ella durante el vuelo con
motor. Una mirada cercana a las plumas del ala del Archaeopteryx también revela un raquis compensado y una veleta
asimétrica (Figura
18.6). Esto sugiere
que en el momento del vuelo propulsado por Archaeopteryx
ya había evolucionado.
19. Partes de una pluma
|| Regresar al Índice ||
Desde el radiante y brillante plumaje naranja del gallo andino de las rocas (Rupicola peruvianus) hasta los patrones de tonos tierra y exquisitamente filagreados en el ala de un gran Argus (Argusianus argus), el plumaje de aves es uno de los más llamativos y encantadores ropajes de animales.

Figura 19.1. Rupicola peruvianus.

Figura 19.2. Argusianus argus.
Pero incluso las plumas más simples del
pájaro más sencillo tienen historias fascinantes que contar: cómo las plumas
individuales están hechas de estructuras intrincadamente dispuestas; cómo
evolucionaron las plumas desde comienzos simples en dinosaurios pre-aviarios
hasta sus formas modernas durante millones de años; cómo las variaciones en el
tema básico de las plumas llevaron a cientos de formas expresadas entre las
casi 10,000 especies de aves del mundo; cómo diferentes tipos de plumas pueden
servir para muchos propósitos simultáneamente dentro de un solo pájaro; cómo
las aves son capaces de producir plumas de todos los colores que reconocemos y
muchos colores que no; cómo los plumajes completos se adaptan al camuflaje o se
seleccionan sexualmente. Claramente, las plumas son fundamentales para lo que
significa ser un pájaro, y representan un componente fascinante y cautivador de
la biología aviar.
Figura 19.3.Características básicas de una pluma.
Hablando anatómicamente, una pluma tiene un extremo proximal (cercano) y distal
(lejano) en relación con su sitio de unión en el ala y / o el cuerpo. Estos
términos también se aplican al describir las partes componentes de una pluma.
Cuando se les pide que dibujen una pluma,
incluso los niños pequeños ilustrarán tres características básicas. Primero,
está el eje, la porción delgada pero rígida, central, en forma de barra. En
segundo lugar, las paletas o aspas “vane” se extienden en direcciones opuestas
desde el eje, formando dos regiones anchas, planas y más flexibles de la pluma.
Tercero, en una inspección minuciosa, las paletas, en lugar de ser lisas y
sólidas como una simple lámina de plástico, tienen una subestructura tipo
peine. Para completar esta descripción general, un biólogo probablemente dirija
la atención a algunos detalles más de una pluma típica (Figura 19.3). El
eje puede subdividirse en dos regiones, la porción más medial (o cerca del
cuerpo) del eje no tiene paletas y define el área llamada cálamo.
19.1 El cálamo
El calamo es relativamente corto y libre
de veletas principalmente porque es la porción de la pluma que perfora la piel
del ave, anclando así la pluma al cuerpo.
19.2 El raquis
El resto del eje es distal (más alejado
del cuerpo) en relación con el cálamo, y las paletas se extienden desde él.
Esta región se llama raquis. Dado que el raquis con frecuencia comprende tanto
como 90-95% de la longitud del eje, los términos raquis y eje a menudo se usan
indistintamente.
19.3 Plumuláceas
vs penáceas
Cuando la mayoría de las personas imaginan
una pluma típica, tienden a pensar en plumas largas de ala o cola en lugar de
un pequeño mechón de plumón, que también es una pluma. Este contraste llama la
atención sobre otra característica estructural fundamental de las plumas, o,
más específicamente, de los álabes de plumas. Una pluma de ala o cola típica es
penácea, una pluma que tiene veletas
con una superficie bidimensional (plana) relativamente lisa y con una forma o
contorno distintivo. Una pluma de plumón es plumulácea,
una pluma con púas que son tan esponjosas que carecen de forma. Las plumas plumuláceas
son incapaces de sostener cualquier forma redondeada más delicada, y su forma
general cambia drásticamente en respuesta a la más leve brisa. Para comprender
qué subyace a esta diferencia en forma de veleta (o falta de ella), uno debe
comprender con más detalle los fundamentos anatómicos de la veleta de plumas
pennaceas más firme (Figura 19.3).
19.4 Las aspas
de las plumas penáceas
Las aspas penáceas son una malla compleja,
transpirable y flexible hecha de cientos de pequeñas fibras entrelazadas. En
una inspección minuciosa, se puede ver que estas aspas se componen de una fila
densa de cientos de filamentos largos, delgados pero rígidos, llamados púas,
que se ramifican desde el raquis uno tras otro, algo así como dientes en un
peine (Figura 19.4A). A
una escala aún más fina, las púas tienen un conjunto de estructuras más
pequeñas, apenas visibles a simple vista. Es decir, a lo largo de las púas, la
estructura general de las plumas (el eje central más grande con muchas fibras
más pequeñas que se ramifican en lados opuestos) se repite a menor escala: cada
púa consiste en un eje central llamado rama que tiene sus propias filas de
ramas delgadas, llamadas bárbulas, que se extienden desde ambos lados
Las bárbulas generalmente no son simétricas en los dos lados de la barba, y nos referimos a las bárbulas que se extienden lejos del cuerpo del ave (o lejos de la base de la pluma) como las bárbulas distales, y aquellas que se extienden hacia el cuerpo como el bárbulas proximales. Los dos lados suelen tener forma y función complementarias; las bárbulas distales a menudo tienen estructuras en forma de gancho a lo largo de sus superficies ventrales (o debajo), llamadas barbicelas o hooklets “enganchadoras” (Figura 19.4B). Los ganchos agarran y sostienen la superficie dorsal (o superior) de las bárbulas proximales de la púa adyacente. Por lo tanto, aunque las púas son largas, delgadas y pueden desconectarse físicamente entre sí, los ganchos en sus bárbulas distales las mantienen paralelas a las púas vecinas en toda su longitud. De esta manera, las bárbulas generan y mantienen la estructura continua de los álabes, lo que hace que la pluma parezca una superficie lisa y continua.

Figura 19.4.
Aspas de plumas y anatomía de la bárbula. (A) Detalle de las aspas de las
plumas con bárbulas proximales y distales que interactúan. (B) Los detalles que
muestran ganchos interactúan con las bárbulas en la púa adyacente. (C) Dos
bárbulas plumulaceas (i) de una paloma de roca (Columba livia), con tres
regiones desde la punta proximal a la distal (ii – iv) agrandadas debajo. En
(ii), la base y los nodos proximales son relativamente grandes en comparación
con la región central de la barbula (iii). Las puntas de la bárbula distal (iv)
tienen nodos sustancialmente reducidos (YouTube).
19.5 Las aspas
de las plumas plumuláceas
Las fibras que crean aspas plumuláceas
también son púas, pero con una estructura diferente. Las púas en las plumas plumuláceas
tienen ramios proporcionalmente más largos y menos rígidos, y sus bárbulas
generalmente se reducen a menos pelos, más delgados y simples (sin ganchos), o
incluso nódulos cortos fuera del ramillete (Figura 19.4C).
En comparación con las fibras densamente compactas, rígidas y enganchadas de
los álabes penáceos, la falta de estructura de bárbula libera a las ramas
largas y delgadas para que se muevan independientemente unas de otras, y crea
la forma suelta y copetuda de un álabe plumuláceo. Las diferentes funciones de
estas formas de paletas contrastantes se abordan más adelante en este capítulo.
19.6 Material
de construcción
Todas estas estructuras de plumas están hechas del mismo material: queratina. La queratina es un tejido conectivo proteináceo (el mismo material básico de las uñas humanas) producido dentro de células especializadas llamadas queratinocitos. Entre todas las especies en el reino animal, hay un conjunto diverso de genes que producen queratina, y en consecuencia existen diversas formas de proteínas de queratina. Dentro de los animales vertebrados, las queratinas pertenecen a una de dos formas estructurales fundamentalmente diferentes (Figura 19.5A). Las alfa-queratinas se encuentran en todos los vertebrados (incluidos nosotros mismos), son relativamente flexibles y constituyen gran parte de la piel, el cabello y las uñas de los mamíferos.

Figura 19.5. Estructura y uso de queratina. (A) Las diferentes
estructuras moleculares de las queratinas alfa y beta crean estructuras y propiedades
plegables (terciarias) distintivas. (B) Las beta-queratinas son exclusivas de
las aves y los reptiles y son el principal componente estructural de las plumas
y las garras, como en las garras de este águila calva (Haliaeetus
leucocephalus).
Las beta-queratinas son exclusivas de los
reptiles y las aves, tienden a ser más duras y se encuentran en escamas, garras
y plumas (Figura 19.5B).
La beta-queratina específica que se encuentra en las plumas es una forma
exclusiva de las aves vivas. La diversidad de las beta-queratinas dentro de las
aves todavía está en gran parte inexplorada y, dada la importancia de las
propiedades materiales de la queratina para la función de las plumas, y de la
función de las plumas para las aves en general, promete contar algunas
historias interesantes sobre la evolución de las plumas y las aves (Glenn et
al.2008).
20 Detalles sobre el crecimiento de las plumas
|| Regresar al Índice ||
Uno podría imaginar que la estructura de
doble ramificación de una pluma típica emerge a medida que la pluma crece, de
una manera análoga a la forma en que un árbol crece sus ramas: por pequeñas
puntas que envían brotes de una rama en crecimiento. En cambio, las plumas
tienen su propia forma sorprendente de crecer en su forma ramificada
distintiva.
20.1 Los
folículos
El desarrollo de las plumas es un proceso complejo que comienza y termina con los folículos de las plumas, las estructuras especializadas en la superficie de la piel que generan plumas. Si ha visto pollo crudo, probablemente haya notado los folículos de plumas como pequeños bultos y / o poros en la superficie de la piel (Figura 20.1). La clave para entender cómo un folículo hace una pluma es comprender su anatomía.

Figura 20.1. Folículos de plumas. Cada bulto elevado en la piel de este pollo desplumado es un folículo de plumas. Ocurriendo en distintos tramos llamados pterilos que siguen los contornos del ave, están separados por regiones de piel desnuda sin folículos.

Figura 20.2. Crecimiento de plumas y de la papila dérmica. (A) El brote temprano
de una nueva pluma, que se eleva desde la placoda epidérmica (no se muestra la
piel epidérmica engrosada) y las capas cutáneas dérmicas, forma la papila. (B)
Las células epidérmicas en la base de la papila crecen rápidamente. Estas
células eventualmente se doblan hacia atrás en la capa dérmica, formando una
capa doble de piel epidérmica invaginada: el folículo de la pluma. (C) Una
sección transversal de la pluma en crecimiento dentro del folículo revela la
disposición de la piel epidérmica, creando el collar epidérmico y su núcleo dérmico.
20.2 Desarrollo
de los folículos de plumas
La piel de un pájaro (como nuestra piel)
consta de dos capas principales, la dermis interna y la epidermis externa.
Durante el desarrollo dentro del huevo, la superficie de la piel del embrión
está visiblemente salpicada de pequeñas protuberancias llamadas capullos de
plumas o papilas. Cada papila se forma cuando una pequeña inflamación de la
dermis, el núcleo dérmico, empuja hacia su piel epidérmica engrosada
suprayacente (llamada placoda epidérmica) (Figura 20.2A). A
medida que el desarrollo progresa en el embrión, cada papila se transforma en
un folículo de plumas cuando las células epidérmicas en la base de la papila
comienzan a crecer nuevamente en la dermis. El crecimiento de esta capa de piel
en el perímetro de la papila es tan rápido que la epidermis se dobla sobre sí
misma, invadiendo la dermis y formando así una doble pared dentro de la piel
(la pared interna rodea el núcleo dérmico, la pared externa se conecta a la
superficie de la piel) (Figura 20.2 B).
Por lo tanto, una definición más formal de un folículo de plumas es una columna
de epidermis de doble pared con un núcleo dérmico a partir del cual se
desarrollan las plumas.
Una vez desarrollados en el embrión, los
folículos permanecen en la piel por el resto de la vida del ave. El crecimiento
celular complejo ocurre dentro del folículo cada vez que se desarrolla una
pluma. Las células de la capa epidérmica interna del folículo son el lugar de
este crecimiento y se denominan collar epidérmico (Figura 20.2C).
20.3 Reemplazando
las plumas
Dentro del collar epidérmico, los
queratinocitos se multiplican rápidamente y se transforman en células llenas de
la queratina que se convierte en la pluma. Cuando se completa el desarrollo de
una pluma individual, las células dejan de multiplicarse dentro del folículo y
quedan inactivas. Sin embargo, el folículo, con su núcleo dérmico y collar
epidérmico, todavía existe, listo para reiniciar el proceso cuando sea el
momento de producir una nueva pluma. Los nuevos queratinocitos, a través de su
nuevo crecimiento rápido, simplemente empujarán a sus progenitores hacia arriba
y hacia afuera del collar, empujando la pluma vieja y reemplazándola por la
siguiente. El mismo folículo puede producir varios tipos de plumas diferentes
durante la vida de un ave.
20.4 Si lo que
sale es un tubo ¿por qué la pluma no es un pelo?
Dos aspectos de la anatomía del folículo hacen que el desarrollo de una pluma sea un poco contradictorio.

Figura 20.3. Etapas de una pluma desplegada. (A)
A medida que emerge de la papila dérmica, una pluma se asemeja a una pajita con
una hendidura en un lado (línea punteada negra). Las superficies internas y
externas tienen diferentes curvaturas. (B) A medida que se despliega, el
interior del tubo forma la superficie ventral (parte inferior) de la pluma. (C)
Cuando una pluma se ha desplegado por completo, su superficie ventral es
cóncava y su superficie dorsal es convexa.
El primero es el hecho de que los
folículos de plumas son estructuras tubulares. ¿Cómo se convierte el tejido
extruido de una superficie de crecimiento circular en una pluma
"plana"? Esta transformación se describe con mayor detalle más
adelante en esta sección, pero como introducción, imagine que la pluma en
crecimiento dentro del folículo se expulsa de la piel como una columna hueca de
tejido con forma de paja (Figura 20.3A).
Dividir la pajita a lo largo de un lado y desenroscarla (Figura 20.3B)
produce una estructura larga, aplanada pero cóncava (Figura 20.3C).
En términos generales, las plumas crecen rizadas con los bordes exteriores de
sus veletas tocándose. Este tipo de despliegue es la base de cómo emerge una
pluma plana de su forma columnar inicial.
El próximo desafío es descubrir cómo el folículo, que permanece fijo en la superficie de la piel, puede desarrollar una estructura intrincadamente ramificada hacia atrás, desde las puntas externas hasta la base. Es decir, en lugar de brotar púas del raquis en crecimiento a medida que el raquis crece (y las bárbulas de las púas a medida que crecen las púas), las plumas se desarrollan y emergen de sus puntas distales primero. La estructura de ramificación se produce cuando las fibras de crecimiento más fino migran o se desplazan dentro del folículo cilíndrico con el tiempo, para fusionarse en fibras más grandes (como pequeñas corrientes que se funden en ríos más grandes) a medida que crecen. Por lo tanto, las plumas no son estructuras tan intrincadamente ramificadas como los árboles, sino que son redes de fibras intrincadamente fusionadas en las que las fibras más pequeñas se originan primero, luego se fusionan y finalmente emergen del interior del folículo de plumas (Figura 20.4).

Figura 20.4. Crecimiento de plumas penáceas. El crecimiento comienza dentro del
placoda epidérmica (A) con el inicio de crestas de púas de plumas (B). Dentro
de la vaina de plumas, las crestas dorsales de las lengüetas continúan creciendo
hacia abajo (C, D), y finalmente se encuentran con el collar epidérmico
alrededor de la base de la placoda. La base ventral de la pluma en crecimiento
forma otra cresta de púas (E, F), con un crecimiento helicoidal que conduce a
la formación de una cresta central de raquis de la pluma aún envainada (G, H,
I). La vaina se deteriora (J), permitiendo que las púas de la pluma pennacea se
desplieguen completamente (K, L).
Con estas dos generalizaciones en mente:
una pluma se cultiva como una paja de paredes delgadas que se despliega a lo
largo de su eje largo, y su estructura ramificada se crea mediante la fusión
intrincada de fibras finas. Examinemos la biología del desarrollo de una pluma
pennácea típica en más detalle.
20.5 Generando
las barbas
El collar epidérmico es donde todo sucede para generar la estructura ramificada de la pluma. La organización geométrica de las células en crecimiento y fusión dentro del collar es lo que crea la clásica forma de pluma de un eje central (el raquis) con ramas laterales simétricas (las púas). Debido a que la posición de los queratinocitos dentro del collar epidérmico determina en qué parte de la pluma final se convertirán las células, es importante comprender la relación entre la posición del crecimiento celular dentro del folículo y la posición de las estructuras terminadas en la pluma madura.

Figura 20.5.
Superficies dorsales y ventrales. Estas guacamayas rojas (Ara macao)
exponen ambos lados de sus plumas primarias durante el vuelo. El lado dorsal
(superior) de cada primario es azul; el lado ventral (abajo) es rojo.
De la misma manera que el ave entera tiene
una espalda, una barriga, una cabeza y una cola, el collar epidérmico también
tiene una orientación o lados. Lo más simple, cuando se despliega, las células
en el interior del collar (la superficie interior de la paja) se convierten en
la superficie ventral (o inferior) de la pluma, y las del exterior del collar
(la superficie exterior de la paja) se convierten en la pluma. superficie dorsal
(o superior) (Figura 20.5).
Otro eje básico de una pluma es proximal / distal (o cercano al cuerpo del ave
versus más alejado). A medida que crece una pluma, se generan células más
nuevas debajo de las más viejas, de modo que el nuevo crecimiento fuerza al
crecimiento más viejo hacia afuera, lejos del cuerpo (al igual que crecen
nuestras uñas). Así, las primeras células crecidas corresponden a la punta
distal de la pluma, y las últimas células, la base proximal del cálamo (Figura 19.3).
El collar epidérmico tiene otras regiones visiblemente diferenciadas que corresponden a regiones de la pluma madura. Un área es una inflamación distintiva o cresta de queratinocitos llamada cresta del raquis (Figura 20.6 A). Como es de esperar, las células que crecen dentro de la cresta del raquis producen el raquis. El tejido que forma el resto del collar está compuesto por crestas de púas: tiras de queratinocitos activos, agrupados en paquetes (un paquete de queratinocitos por cresta de lengüeta) (Figura 20.6 B) y creciendo en una ruta semicircular hacia la cresta del raquis. A medida que las crestas de las púas crecen, se convierten en púas alargadas y filamentosas que eventualmente convergen y se fusionan con el raquis que crece continuamente (Figura 20.7). Nuevas crestas de púas se forman continuamente en el collar epidérmico opuesto a la cresta del raquis, y cada cresta de púas se convierte exactamente en una púa.

Figura 20.6. Collar epidérmico y anatomía de la cresta de la lengüeta. (A) Sección transversal del collar epidérmico y la cresta del raquis, donde la queratina generada por los queratinocitos produce el raquis de una pluma. (B) Una ampliación de las crestas de púas, que crecen hacia la cresta del raquis.

Figura 20.7. Progresión del crecimiento de las plumas desde el collar epidérmico.
Contraintuitivamente, las púas de plumas distales (punta de la pluma) crecen
del collar epidérmico antes de fusionarse con la cresta central del raquis más
adelante durante el proceso de crecimiento.
Cuando el collar epidérmico se ve en
sección transversal, se ve como un anillo, con bloques de tejido activo, las
crestas de las barbas, que rebordean el perímetro y con la cresta del raquis
formando una porción ampliada en un lado del anillo (Figura 20.6 A).
En una escala más fina, las púas individuales se forman mediante un proceso
similar al que ocurre dentro del collar epidérmico en su conjunto, con la
migración celular durante el crecimiento que conduce a bárbulas delgadas que
finalmente se fusionan con las ramas de la púa (Figura 20.6B).
En resumen, aunque la estructura de
crecimiento de plumas, el folículo, es un cilindro hueco de tejido, las células
de crecimiento de queratina se distribuyen en tiras dentro de ese cilindro.
Estas células migran y se fusionan entre sí de maneras muy específicas,
creando, cuando se despliega, la estructura doblemente ramificada y
relativamente bidimensional de la pluma. Un resultado de este anillo de
crecimiento constantemente extruido es que todas las fibras en un punto
particular a lo largo de la pluma madura, ya sean las del raquis, las púas o
las bárbulas, se originaron al mismo tiempo (Figura 20.7) y,
por lo tanto, representan un Momento único de tiempo durante el crecimiento.
Por esta razón, las diferentes estructuras de plumas no se originan como una
serie simple. En cambio, la punta distal del raquis se origina en el momento
del inicio del desarrollo de cada pluma, mientras que las púas se originan una
y otra vez, a menudo en pares, comenzando en las puntas opuestas al raquis y
creciendo nuevamente para fusionarse con el raquis. Como el raquis la cresta
genera a menudo, docenas de púas que crecerán al mismo tiempo, cada una en
diferentes puntos a lo largo de su longitud, y cada una se unirá
secuencialmente al raquis.
El núcleo dérmico, la estructura que originalmente precipitó el desarrollo del folículo de la pluma, ocupa el espacio dentro del collar epidérmico durante el desarrollo de la pluma. Mientras se desarrolla una pluma, los vasos sanguíneos se extienden hacia la base de la pluma en crecimiento, proporcionando alimento para las células en crecimiento. A medida que las células se multiplican, y son empujadas cada vez más lejos del collar epidérmico, dejan de recibir alimento del núcleo dérmico y, en última instancia, se deshidratan y mueren, mientras que las células más jóvenes aún crecen en la base de la pluma. Cuando la pluma está completamente formada, el suministro de sangre a toda la pluma se corta a medida que los vasos se reabsorben en la papila a través de un orificio en el extremo del cálamo. En una pluma grande, este agujero a menudo se puede ver sin aumento.

Figura 20.8. Plumas de alfiler. Este polluelo
de cuitlacoche rojizo (Toxostoma rufum) tiene plumas de alfiler visibles
que emergen de sus vainas cerosas y azuladas mientras muda desde sus plumones
de nacimiento hasta su plumaje juvenil. Eventualmente, estas vainas de plumas
de alfiler se romperán o serán removidas por el ave durante el acicalamiento.
Cuando se observa un pájaro vivo, las plumas en crecimiento pueden reconocerse por la presencia de varillas extrañamente gruesas, de aspecto plástico, donde las plumas estarían de otra manera. Estas plumas de crecimiento distintivas se llaman plumas de alfiler: plumas recién crecidas aún encerradas dentro de una cubierta cerosa azulada o grisácea llamada vaina (Figura 20.8). Al desarrollarse independientemente del resto de la pluma, la capa externa del collar epidérmico produce la vaina a medida que la pluma crece. Funcionalmente, la vaina protege la pluma naciente a medida que crece y luego se seca y muere. También sostiene la pluma en su forma cilíndrica y curvada mientras continúa creciendo en su base. A medida que la pluma madura, y sus células se deshidratan y mueren, la vaina también se seca, se divide y se cae o el ave la corta.

Figura 20.9. Barras de fallas en plumas.
Cuando un ave experimenta altos niveles de estrés (como limitación severa de
alimentos) durante el desarrollo temprano o antes de una muda adulta, pueden
aparecer barras de falla, causadas por una reducción temporal en el crecimiento
de la fibra, en sus nuevas plumas. A menudo, estas barras de falla son más
evidentes en las rectrices (plumas de la cola).
Aunque el proceso general de desarrollo de
plumas descrito aquí procede de procesos estrictamente programados y regulados,
los factores ambientales externos pueden influir en el desarrollo de la
estructura de plumas. Por ejemplo, breves deficiencias nutricionales o
tensiones a corto plazo pueden conducir a una reducción repentina del área de
la sección transversal de las fibras de púas y bárbulas, creando líneas
visibles en la pluma madura conocidas como barras de falla (Figura 20.9).
21. Tipos de plumas en las aves modernas
|| Regresar al Índice ||
Los folículos de plumas son los sitios de crecimiento y desarrollo de las plumas. Al igual que los folículos pilosos y sus pelos, las plumas maduras permanecen unidas al cuerpo en el sitio del folículo, pero a diferencia de los pelos, las plumas se distribuyen en la superficie de un pájaro en patrones más espaciados pero estrictamente regulados. Esto se puede ver en la piel de un pollo fresco entero (Figura 21.1), pavo o pato, donde los folículos aparecen como poros pequeños, distintos y regulares, casi geométricamente espaciados.

Figura 21.1.Folículos de plumas. Cada
bulto elevado en la piel de este pollo desplumado es un folículo de plumas.
Ocurriendo en distintos tramos llamados pterilos que siguen los contornos del
ave, están separados por regiones de piel desnuda sin folículos.
En la mayoría de las aves, las plumas no se distribuyen uniformemente sobre el cuerpo, sino que se agrupan en tractos de plumas, más formalmente llamados pterilos, que son grupos definidos de plumas. Separando estos tractos hay regiones de piel desnuda (sin plumas) llamada apteria. Las plumas de los pterilos se extienden fácilmente sobre los espacios creados por la apteria, y las bases suaves de las plumas tienden a desprenderse, por lo que estos parches desnudos rara vez son visibles en un ave viva.

Figura 21.2. Regiones de crecimiento de plumas. En
aves jóvenes o desplumadas, los folículos de plumas están claramente dispuestos
en tractos de plumas, o pterilos. Las áreas sin plumas se llaman apteria.
La disposición detallada de los tractos de
plumas y los parches desnudos en todo el cuerpo, denominados colectivamente
pterilosis, varía entre los grupos de aves, y algunas aves tienen patrones
únicos de pterilosis. Por ejemplo, las aves de la familia Corvidae, que incluye cuervos, y arrendajos, tienen un apterio característico
en la espalda, a lo largo de la línea media de la columna vertebral. Por otro
lado, algunos tipos de aves, especialmente las aves acuáticas como los
pingüinos, tienen una pterilosis continua sin apteria, presumiblemente para
ayudar a evitar que el agua penetre en la piel de las aves y la enfríe.
Históricamente, tales patrones distintivos proporcionaron pistas importantes
para identificar algunos de los principales grupos evolutivos de aves.
Las plumas a menudo cumplen diferentes funciones en diferentes partes del cuerpo de un pájaro, y, por lo tanto, las plumas que pueblan las diversas pterilas vienen en una gran variedad de formas y tamaños (Figura 21.3A). Un ave típica podría tener una capa interna mullida para aislar; plumas que cubren el torso que actúan como protector solar, cortavientos o impermeable dependiendo del clima; cerdas sin vellos alrededor de la boca para funciones de alimentación; fuertes plumas de ala y cola que se usan para volar; y penachos ornamentales (que pueden ubicarse en cualquier número de lugares) para señalizar a posibles compañeros o rivales (Figura 21.3B).

Figura 21.3. Tipos
de plumas. (A) Ejemplos de seis tipos de plumas: plumas pennaceas del ala y la
cola, semipluma, pluma de contorno, plumón, filopluma y cerda. (B) Plumas
utilizadas para la exhibición: durante las exhibiciones de atracción de pareja,
el Pájaro del Paraíso del Rey de Sajonia (Pteridophora alberti) empuja
sus extraordinarias plumas de cabeza hacia adelante mientras trina
maravillosamente.
Desde el punto de vista de la ingeniería
adaptativa, los diversos tipos de plumas interactúan maravillosamente con los
dos entornos físicos básicos que encuentran: medios fluidos (en forma de aire y
agua) y radiación electromagnética (en forma de calor y luz). Al variar las
estructuras básicas de plumas del raquis, la púa y las bárbulas, las aves
pueden variar las propiedades aerodinámicas, hidrodinámicas, aislantes,
repelentes, acústicas y visuales de sus plumas. La diversidad resultante de
formas subyace a la riqueza de las funciones de plumas. La diversidad de plumas
existe dentro de un ave individual en un momento dado y en un contexto dado,
pero también a lo largo de la vida de un individuo, y ciertamente entre
especies.
21.1 Plumones
de aislamiento
Las plumas de plumón son completamente plumulaceas, definidas por tener un raquis que es más corto que las púas más largas, o con la falta del raquis por completo. Estructuralmente, son relativamente simples: sus púas emergen más o menos juntas como un penacho cerca de la base de la pluma, donde flotan unas de otras con poco o nada de raquis (Figura 21.4).

Figura 21.4. Anatomía del plumón. Es
un excelente aislante, ya que sus púas flotantes atrapan grandes cantidades de
aire estático, que aísla contra el flujo térmico. (A) Una pluma de plumón
típica; Tenga en cuenta que las púas se extienden sustancialmente más lejos que
el raquis. (B) Detalle de la estructura de plumas de plumón hacia abajo.
Como resultado, las plumas de plumón son
suaves y esponjosas. Como criaturas de sangre caliente, las aves deben ser
capaces de controlar sus temperaturas internas mediante termorregulación:
regulando la pérdida de calor y la ganancia de calor. Las plumas de plumón son
excelentes aislantes, en algunos aspectos superiores a los aislantes utilizados
en la ropa humana, como la lana y el poliéster. El papel del plumón como
aislante liviano, pero de alta calidad es especialmente importante para las
aves pequeñas que viven en climas fríos, así como para las aves que flotan, se
sumergen o nadan en el agua.
Las capacidades aislantes superiores se
deben a su estructura de fibra extremadamente fina, las púas y las bárbulas
forman pequeños enredos sueltos que atrapan el aire y su calor. La estructura
relativamente sin forma de las plumas suaves es uno de los factores que los
convierten en excelentes aislantes. Sin embargo, a la pequeña escala de las
púas y las bárbulas de plumas, el aire se comporta de manera viscosa, por lo que
el aire que se mueve entre las púas enredadas se puede imaginar como una
sustancia espesa, como el aceite o la miel, que se mueve a través de un tamiz
fino. Las plumas de plumón, en lugar de depender de una barrera cerrada para
reducir el flujo de aire, literalmente atrapan y enredan físicamente el aire
caliente en sus finas fibras.
Las aves tienen tres tipos distintos de
plumones: natales, corporales y polvorientos.
21.2 Plumones
natales
Los plumones de natales generalmente cubren todo el cuerpo del ave, son temporales (presentes solo en el momento de la eclosión) y surgen de los mismos folículos de plumas que luego producen plumas de contorno. De hecho, la continuidad del folículo de plumas (y su collar epidérmico) entre el crecimiento de las plumas posteriores se puede ver en algunas aves jóvenes cuyas plumas natales permanecen unidas a las puntas de las plumas de contorno que crecen posteriormente. Este apego débil entre el plumón natal y la posterior pluma de contorno da como resultado la apariencia esponjosa y descuidada de algunas aves juveniles como zorzales, petirrojos y pájaros azules.

Figura 21.5.
Plumaje suave en pichones precociales versus altriciales. (A) Dado que los
pollitos precociales deben mantener inmediatamente su propia temperatura
corporal, eclosionan con una gruesa capa de plumón y habilidades motoras
avanzadas. (B) Las crías altriciales son completamente indefensas, con poco
aislamiento (B flecha). Dependen de sus padres para brindarles calor durante
las primeras etapas de sus vidas. (C) Polluelos en una categoría intermedia,
que incluye muchas especies de crecimiento lento (como halcones, águilas y
búhos), eclosionan con poca movilidad, pero con plumones considerables, ya que
deben permanecer expuestos durante horas esperando sus comidas difíciles de
capturar, especialmente las que habitan climas fríos y templados.
Las aves precociales y altriciales suelen
tener distintas estrategias de termorregulación y, por lo tanto, pueden tener más
(en el caso de las aves precociales) o menos tiempo (en el caso de las aves
altriciales). La mayoría de las aves precociales abandonan el nido poco después
de la eclosión y deben mantener su propia temperatura corporal en forma de
aislamientos de nacimientos. Por ejemplo, los patitos o pollos recién nacidos
son conocidos por su aspecto suave y esponjoso (Figura 21.5A).
En contraste, muchas aves altriciales están mucho más indefensas después de salir
del cascarón y dependen del calor de un ave progenitora que se está
reproduciendo. Si bien el plumón protege a los pichones precociales al actuar
como un aislante entre la temperatura corporal del pichón y el medio ambiente,
también puede evitar la transferencia de calor deseable entre el pichón y su
progenitor, y en estos casos, demasiado plumón natal evitaría la transferencia
de calor del progenitor a los jovenes. Los pichones altriciales, por lo tanto,
a menudo solo tienen escamas natales dispersas, lo que permite una
transferencia de calor más eficiente a los pichones desde la piel cálida y
desnuda en el vientre de sus progenitores (Figura 21.5B).
En algunos casos, para las crías de desarrollo más lento que no pueden volar
durante semanas después de la eclosión pero que, sin embargo, pasan horas solas
en el nido, los polluelos pueden tener una cubierta aislante de plumas suaves
que dura gran parte de su período de cría. Los halcones y los búhos jóvenes,
por ejemplo, pueden tener una cubierta suave y prolongada (Figura 21.5C).
21.3 Plumones
corporales
Los plumones corporales son distintos de
los plumones natales, ya que los plumones corporales de las aves adultas surgen
de folículos que solo producen plumas. Estas plumas de plumón adultas a menudo
se distribuyen por todo el plumaje de las aves, y son más abundantes en aves
acuáticas como pingüinos, bribones, petreles, auks, gansos y patos (todos los
cuales, sin duda, se benefician del aislamiento térmico adicional). Muchas
aves, como las aves acuáticas hembra, arrancan sus plumones de sus vientres
para recubrir sus nidos. De estos, el más famoso es el Eider común (Somateria mollissima) (Figura 21.6).

Figura 21.6. El
eider común hace sus nidos con sus propias plumas de plumón corporales.
La ropa y cobikas de cama rellenas con
edredones de pluma de Eider son notoriamente caras porque el plumón
excepcionalmente suave se debe recoger a mano y cada nido generalmente proporciona
solo unos 15 gramos de plumón, aun así, al tratarse de una estructura que ha
evolucionado por millones de años para proteger del calor, las hacen muy
apetecidas.
21.4 Plumones polvorientos

Figura 21.7. Parche
de plumas polvorientas. En unos pocos grupos de pájaros, las puntas de las
plumas polvorientas, que se muestran aquí en una garceta nevada (Egretta
thula), se desintegran para producir un repelente de agua queratinoso que
se asemeja al polvo de talco.
A diferencia de los otros dos tipos de
plumones (y todos los demás tipos de plumas, de hecho), los plumones
polvorientos son distintivos porque nunca se mudan, sino que crecen
continuamente, desintegrándose en sus puntas para producir una fina sustancia
queratinosa que se asemeja a la textura del polvo de talco. El polvo es lo
suficientemente abundante como para impregnar todo el plumaje del ave,
probablemente ayudando a que sea resistente al agua. La apariencia de las
plumas polvorientas puede variar; se ven como plumas de contorno esponjosas en
algunas especies, o como pequeños plumones en otras. El polvo se produce solo
en ciertos grupos de aves, como garzas y palomas, y estos plumones pueden estar
dispersos por todo el plumaje o agrupados en parches (Figura 21.7).
21.5 Plumas de
contorno para toda ocasión
Llamadas plumas de contorno porque le dan a un pájaro su forma o contorno característico, estas plumas cubren la mayor parte de la superficie de un pájaro adulto. Las plumas de contorno se definen por tener un raquis más largo que la lengüeta más larga y al menos una porción del raquis que consiste en una veleta penácea (Figura 21.8).

Figura 21.8. Plumas de
contorno. Por definición, el raquis de una pluma de contorno debe ser más largo
que la púa más larga, y al menos una porción debe ser pennácea. Las plumas de
contorno crean la silueta característica de un pájaro. La mayoría de las plumas
de contorno son simétricas y por ende no están involucradas en generar
elevación.
Con frecuencia hay una porción plumulácea
de la veleta en la base de las plumas de contorno. Las plumas de contorno
tienen muchos roles, a menudo cumplen una amplia variedad de funciones
simultáneamente o en diferentes contextos. Colocadas una encima de otra, pero
compensadas como las tejas en un techo, las porciones pennaceas de los álabes
de plumas de contorno forman una barrera sorprendentemente efectiva. El plumaje
puede ayudar en la termorregulación al absorber o reflejar la luz solar
generadora de calor y desviar el viento y / o atrapar el aire caliente. En
términos de aerodinámica, las plumas de contorno racionalizan el cuerpo durante
el vuelo, disminuyendo las turbulencias, pero no generan empuje. Como lo
demuestran los contornos particularmente suaves de las aves que nadan, como las
alzas y los pingüinos, la función de racionalización es aún más crítica en el
agua, donde la resistencia es mucho mayor que en el aire.
La capacidad de las plumas para repeler el agua es otra propiedad importante que se observa mejor en las plumas de contorno, una que varía entre las especies de aves según su morfología de plumas. Las mismas plumas de contorno que perfeccionan los contornos de un pato zambullidor también repelen el agua bellamente, como lo ilustra el dicho idiomático "como el agua de la espalda de un pato". Se ha demostrado que los aceites de la glándula uropigial en la base de la cola mejoran la repelencia al agua, pero la forma y el espaciado de las púas y las bárbulas de una pluma son en realidad más importantes para conferir sus propiedades repelentes al agua. Esto se debe a que una propiedad física fundamental del agua es que, si bien tenderá a adherirse a la mayoría de los materiales sólidos, como la queratina, no puede adherirse a los espacios de aire en los materiales (incluso espacios muy pequeños). Entonces, al atravesar espacios de aire en un material, el agua formará gotitas.

Figura 21.9. Plumas
repelentes al agua. (A) Cuentas de agua en la superficie de la mayoría de las
plumas debido a los espacios de aire entre las púas y las bárbulas. (B) Cuando
las aves acuáticas como este pato de madera (Aix sponsa) resurgen de una
inmersión, el agua se acumula y se desliza de sus plumas con facilidad. (C) La
glándula uropigial (la pequeña protuberancia amarillenta en la base de la cola)
excreta aceites que las aves usan para mantener e impermeabilizar sus plumas.
Aquí una Crossbill de alas blancas (Loxia leucoptera) accede a sus
aceites uropigiales con su pico, que luego utilizará para extender los aceites
sobre su plumaje.
Por lo tanto, si las púas y las bárbulas
en una pluma están espaciadas exactamente a la derecha, el agua se ve obligada
a acumularse y rodar. Alternativamente, con púas y bárbulas espaciadas de
manera diferente más juntas, el agua puede ser arrastrada hacia la pluma,
saturándola o impregnándola. Algunas bajas (pero no todas), por lo tanto,
tienden a ser vulnerables a la saturación. Algunas aves de buceo, como los
cormoranes y las anhingas, tienen plumas que son relativamente fáciles de mojar.
La naturaleza más absorbente de agua de las plumas de los cormoranes les
permite bucear bajo el agua sin llevar burbujas de aire atrapadas en las
plumas, cuya flotabilidad contrarrestaría los esfuerzos de buceo de las aves a
la superficie.
La porción plumulácea de la veleta en la base de muchas plumas de contorno generalmente está protegida de los elementos por la porción penácea, y tiende a contribuir con una función aislante a las plumas de contorno. Algunas plumas de contorno tienen una pluma posterior (a veces también llamada aftershaft/pluma posterior), una pluma menor que brota del mismo folículo, pero es (generalmente) más pequeña que la pluma principal y (generalmente) completamente plumulacea (Figura 21.10 A). En el desarrollo, las plumas posteriores se forman dentro de un folículo de plumas al seccionar una porción del collar epidérmico y crear una segunda cresta de raquis, hacia la cual luego crece un subconjunto de las crestas de las púas. Las plumas posteriores proporcionan un aislamiento adicional y están especialmente bien desarrolladas en urogallos (la mayoría de los cuales viven en regiones estacionalmente frías o árticas). El Emu no volador (Dromaius novaehollandiae) de Australia es único en tener plumas posteriores que son tan grandes como las plumas de contorno principales (Figura 21.10B).

Figura 21.10. Plumas plumuláceas posteriores.
(A) Una pluma de contorno típica con una pluma posterior plumulácea conectada a
la derecha. Las plumas posteriores, que se desarrollan a partir del mismo
folículo, pueden servir para proteger la pluma principal. (B) Dos plumas
Plumuláceas de doble eje de un Emu (Dromaius novaehollandiae), la única
especie que desarrolla una pluma posterior hasta el contorno principal.
Las plumas de contorno, aunque relativamente largas, planas y anchas, se unen al cuerpo en un solo punto pequeño. Parecería que, como las hojas en el viento, tales plumas serían propensas a desaliñarse o desorganizarse entre sí. Para mantener su organización ordenada, cada pluma de contorno se une a un conjunto de músculos pequeños y especializados. Ubicada debajo de la superficie de la piel en un perímetro alrededor del folículo, la tensión en estos músculos mantiene la pluma posicionada correctamente. Estos músculos también permiten que un pájaro mueva sus plumas voluntariamente. En algunos casos, las láminas musculares inmediatamente debajo de la piel actúan junto con los músculos de plumas individuales para coordinar el movimiento de las plumas a lo largo de todo el tracto de plumas. La mayoría de las plumas no pueden moverse individualmente, y el movimiento de la mayoría de las plumas se limita a subir y bajar las plumas, esponjándolas y durmiéndolas alternativamente.

Figura 21.11. Pluma de vuelo
terminología. A medida que este Caracara con cresta (Caracara cheriway) se
desliza hacia la tierra, las plumas de vuelo en sus alas (remiges) y cola
(retrices) se extienden por completo. Observe que los bordes anteriores
(delanteros) (amarillo sombreado) están orientados hacia la dirección de vuelo,
mientras que los bordes posteriores más amplios siguen detrás (azul sombreado).
La capacidad de mover plumas de contorno es especialmente importante para la señalización en contextos sociales. La señalización que hacen las aves con sus movimientos de plumas puede ser conspicua y extrema, como en los movimientos de cresta de las cacatúas australianas, o relativamente sutil como se puede observar durante las interacciones entre pájaros cantores en comederos de pájaros en todo el mundo. La longitud, la coloración y / o la morfología de púas y pústulas de las plumas de contorno de algunas especies de aves se modifican para alterar su apariencia. En tales casos, estas plumas a menudo actúan como adornos, estructuras morfológicas cuya apariencia se modifica con el propósito de señalización social y / o exhibición. Las plumas del cuerpo rizado del Manucode con cresta rizada (Manucodia comrii) de Nueva Guinea, las plumas de la cresta eréctil de la cacatúa de palma (Probosciger aterrimus) de Australia, las plumas de la corona rígidas, con forma de paja, de los negros La grulla coronada (Balearica pavonina) de África, y las largas plumas elegantes de muchas garzas y garcetas en todo el mundo son solo algunos ejemplos. Los machos de las aves del paraíso han aprovechado al máximo sus plumas de contorno, utilizándolas para exhibirlas como alambres para la cabeza, protectores de pechos y capas para esconder siluetas.

Figura 21.12. Plumas de contorno modificadas. En algunas
especies, las plumas de contorno especiales sirven como adornos de
señalización, como (A) las plumas de la cresta de una cacatúa de palma (Probosciger
aterrimus), o (B) las plumas de la corona dorada y rígida de una grulla
coronada negra (Balearica pavonina).
21.6 Las
plumas de vuelo
Las plumas de vuelo son un grupo fácilmente definido e identificado, con sus propias ubicaciones específicas, modificaciones estructurales y funciones. Las plumas de vuelo forman la mayoría de las superficies de las alas y la cola de las aves; Por lo general, son las plumas más rígidas y grandes de un pájaro, y son casi completamente penáceas y asimétricas. El ancho de los dos lados de los álabes, lateral y medial, o anterior y posterior, respectivamente, a menudo son asimétricos, ya que el borde delantero (o borde anterior, el borde que corta el aire) es típicamente más delgado que el borde posterior.

Figura 21.13.
Perillas de pluma. Las plumas de vuelo secundarias de un buitre de Turquía (Cathartes aura) se adhieren directamente al cúbito
(A) a través de protuberancias óseas llamadas perillas de plumas (B), que se
muestran aquí sin las plumas adjuntas.
Esta es una adaptación que estabiliza la
pluma bajo las presiones de las corrientes de aire durante el vuelo. Hay dos
conjuntos de plumas de vuelo, las que se originan en el ala, llamadas remiges, y las que forman la cola,
llamadas rectrices (singular: rectrix). A diferencia de la mayoría de
las otras plumas, que se adhieren solo a la piel, las remiges se unen
firmemente a los huesos del ala, ya sea directa o indirectamente a través de
ligamentos, y entre sí mediante ligamentos que conectan sus calamo, por lo que
las huellas de las remiges pueden verse claramente en fósiles de aves y
dino-aves. Las rectrices forman el abanico de la cola, y también están
conectadas entre sí cerca de sus bases por ligamentos. El par más central de
retrices se adhiere directamente al hueso de la cola (el pigostilo).
El número de rectrices varía de 6 a 32 entre las aves voladoras, y los números más altos se producen en aves más grandes. Las remiges más externas (distales) del ala se denominan primarias, definidas por su unión al esqueleto de la "mano" o manus. Dependiendo de la especie, las aves suelen tener entre 9 y 12 primarias. El resto, conocidas como secundarias, se extienden desde el antebrazo (más específicamente el cúbito) del ala del pájaro (Figura 21.13A). Las protuberancias óseas en el cúbito, llamadas perillas de las plumas, indican los puntos de unión de las secundarias y, a menudo, son visibles en los esqueletos de las aves voladoras más grandes (Figura 21.13B). En todos los grupos de aves, el número de secundarias varía más que el número de primarias, que van de 8 a 32. Las aves con alas largas, como los albatros, tienen un mayor número de secundarias, lo que les da una mayor superficie de ala. Para identificar y discutir la biología de las plumas de vuelo importantes, los ornitólogos han desarrollado un sistema de numeración estandarizado para plumas individuales (Figura 21.14).

Figura 21.14. Sistema de numeración para remiges y
rectrices. Los ornitólogos subcategorizan las remiges según su
sitio de unión: las plumas primarias se extienden desde el hueso metacarpiano,
mientras que las plumas secundarias se unen al cúbito. A cada remige se le
asigna un número, comenzando en el punto de encuentro de los dos huesos del
ala; por lo tanto, las primarias están numeradas de derecha a izquierda, las secundarias
de izquierda a derecha. La numeración de las rectrices comienza en el centro de
la cola y se extiende hacia afuera en ambas direcciones. Tenga en cuenta que
los recuentos de plumas de vuelo varían ampliamente entre especies.
Bordeando y cubriendo las remiges y
rectrices en los lados dorsal y ventral del ala y la cola hay hileras de plumas
de contorno llamadas coberteras. Las coberteras son particularmente importantes
para contribuir a la forma aerodinámica del ala y la cola; La aerodinámica del ala
es extremadamente sensible a los detalles de la forma del ala.
Las remiges son las principales fuentes de
generación de elevación, y si bien las rectrices también generan elevación, son
particularmente importantes para estabilidad y control en vuelo. Más allá de su
papel en la aerodinámica del vuelo, vale la pena señalar las propiedades
específicas del material de las plumas de vuelo que las hacen tan excelentes
para manipular y manejar los medios fluidos tanto del aire como del agua.
Obviamente, para generar elevación en el medio muy delgado del aire, las plumas
del ala y la cola deben formar superficies largas y anchas que sean a la vez
firmes, flexibles y livianas. La queratina de plumas, como material, es firme y
flexible, y la estructura tejida de la vela penácea aumenta estas propiedades
al crear una forma muy específica al tiempo que reduce el peso y aporta una
flexibilidad aún mayor que la que podría proporcionar una lámina sólida de
queratina.
Un efecto incidental de las plumas de las
alas que se mueven por el aire es que sus movimientos tienden a generar
sonidos. Las modificaciones microestructurales que intensifican o minimizan la
producción de sonido durante el vuelo están muy extendidas.
En la mayoría de los búhos, por ejemplo, el borde de ataque de las primeras plumas primarias tiene un fleco suelto; La superficie dorsal de los álabes internos de la mayoría de las plumas de vuelo tiene una pila suave compuesta de bárbulas distales alargadas (Figura 21.15).

Figura 21.15. Los búhos tienen plumas de vuelo especializadas. Los búhos son
voladores extremadamente silenciosos, gracias a dos diferencias principales en
su microestructura de plumas de vuelo. Las paletas internas desarrollan una
pila suave de bárbulas distales alargadas (área en caja) que reduce el ruido de
las plumas que se mueven unas sobre otras. Los bordes de ataque de sus
primarios tienen una franja en forma de peine que reduce la turbulencia del
aire y el ruido asociado.
Estas modificaciones permiten a los búhos
volar muy silenciosamente, creando poco ruido incluso dentro del rango de alta
frecuencia que sus presas de roedores pueden escuchar bastante bien. En
contraste, muchas especies de papamoscas del Nuevo Mundo, así como palomas en
todo el mundo, tienen especializaciones de las plumas primarias que exageran
los sonidos producidos por los movimientos accidentales de las alas en sonidos
que funcionan como señales. Como señaló el propio Charles Darwin, los machos de
muchas especies de aves tienen modificaciones de plumas que han evolucionado
para la producción de sonido asociada con comportamientos de exhibición y
selección sexual (Darwin 1872).
21.7 Semiplumas
Los semiplumas representan una forma intermedia entre los plumones y plumas de contorno (Figura 21.16).

Figura 21.16. Los semiplumas son formas de
plumas intermedias. Los semiplumes carecen de ganchos de barbula como plumón,
pero tienen púas más cortas que el raquis, como en las plumas de contorno.
A diferencia de las plumas de contorno,
las bárbulas de los semiplumas a menudo carecen de ganchos y, por lo tanto, las
púas no se adhieren entre sí para formar una veleta penácea. Esto da como
resultado un álabe que no es completamente sin forma como el plumón, pero que,
sin embargo, no está bien conectado como las porciones peninosas de las plumas
de contorno. A diferencia de las plumas, los raquis de las semiplumas son más
largos que la púa más larga. Los semiplumas a menudo ocurren en el borde de los
tractos de plumas de contorno, a veces visibles y otras escondidas debajo de
las plumas de contorno, y en el interior de las alas (esencialmente en la axila
del pájaro) o de manera similar en el interior del muslo. Los semiplumas son
probablemente plumas de contorno que han perdido la porción penácea de sus
veletas, ya que están ubicadas en áreas relativamente protegidas de los
elementos por los contornos del cuerpo del ave. Es probable que complementen la
función aislante de plumón.
21.8 Cerdas
Las cerdas son un tipo de pluma altamente especializado en el que el raquis está relativamente rígido, más afilado y carece de púas en la mayor parte de su longitud. Las cerdas son esencialmente plumas de contorno que se han simplificado para fines funcionales, y se encuentran casi exclusivamente en la región de la cabeza aviar (Figura 21.17).

Figura 21.17. Variaciones de plumas de cerdas.
Los puntos de origen de las cerdas se muestran aquí en un pavo salvaje (Meleagris
gallopavo). Las cerdas a menudo tienen un eje y un eje posterior, ambos
utilizados para recopilar información sensorial.
Las cerdas más conocidas son las cerdas de
rictales, que se proyectan desde la base del pico. Muchas aves, especialmente
las que atrapan insectos en el aire, como los atrapamoscas y los nightjars,
tienen cerdas de rictales. Otras regiones de la cabeza también pueden tener
cerdas, y las cerdas mismas se clasifican por su ubicación, por ejemplo, como
cerdas faciales (desde los lados de la cabeza), cerdas narrales (en el cerebro
o detrás de las narinas) y cerdas lorales (desde los lores).
Algunos ornitólogos han sugerido que las cerdas podrían ayudar a las aves a canalizar insectos hacia la boca, pero se han realizado pocos estudios para probar esta suposición. En un estudio de cazadores de moscas del sauce (Empidonax traillii), Michael Conover y Don Miller (1980) descubrieron que tapar o quitar experimentalmente las cerdas de rictales no afectaba el éxito de las aves en la captura de insectos. Sin embargo, al usar un túnel de viento, estos investigadores demostraron que las cerdas protegen los ojos de las aves, ya que más partículas golpearon los ojos de las aves cuyas cerdas habían sido eliminadas. Las cerdas de rictales son particularmente prominentes en ciertos grupos (como los puffbirds neotropicales) que capturan regularmente polillas escamosas, mariposas y otros insectos grandes y nocivos; Estas cerdas pueden proteger la cara y los ojos de estas aves. Del mismo modo, las "pestañas" en las aves son a menudo cerdas, y estas están particularmente bien desarrolladas en aves como el avestruz (Struthio camelus) y el emú (Dromaius novaehollandiae), y en varias especies de rheas, halcones, pavos, cálaos y cucos (Figura 21.18). Otra función potencial posible pero básicamente no estudiada de las cerdas ricas es detectar el movimiento de la presa sostenida en el pico, proporcionando una función sensorial similar a los bigotes sensibles de algunos mamíferos.

Figura 21.18. Plumas de cerdas
("pestañas"). Presumiblemente, estas plumas de cerdas brindan
protección adicional a los ojos, lo que puede ser particularmente importante
para las aves de campo abierto como (A) el Hornbill de tierra del sur (Bucorvus
leadbeateri) y (B) el pájaro secretario (Sagittarius serpentarius).
¡ya quisieras tu tener estas pestañas!
21.9 Filoplumas
Esparcidos y generalmente ocultos por las plumas de contorno de la mayoría de las aves es un tipo de pluma distintivo pero raramente visible. Estas filoplumas son las plumas más pequeñas, con un raquis desnudo extremadamente delgado y un pequeño mechón de púas (si las hay) solo en la punta (Figura 21.19).

Figura 21.19. Filoplumas: el tipo de pluma más pequeño y más primitivo
actualmente reconocido. Los cambios sutiles en la posición del filopluma,
causados por los movimientos o el entorno de un ave en lugar de los músculos
cerca del folículo, pueden proporcionar información sensorial importante
relevante para procesos como la termorregulación.
De
lo contrario, recuerda a las cerdas mucho más grandes, las filoplumas son
distintas en la falta de músculos de plumas, en lugar tienen receptores
sensoriales distintivos en la piel, al lado de sus folículos. Se cree que estas
minúsculas plumas controlan el movimiento dentro del abrigo de plumas.
Presumiblemente, las aves usan esta información para monitorear y detectar
cambios en las posiciones de las plumas que podrían ser causadas por el viento
o los movimientos del cuerpo.
22. Evolución de las plumas 1, los dinosauros
|| Regresar al Índice ||
Entre los organismos vivos, solo las aves
tienen plumas. Como hemos empezado a ver, las plumas (y su desarrollo) son tan
complejas que plantean preguntas sobre cómo evolucionaron en los antepasados
de las aves modernas. Hasta este momento solo hemos visto puntos diseminados
en especies vivas y fósiles, pero la pregunta es ¿Cómo los unimos en un modelo
que sea predictivo?
No cabe duda que la pluma puede ser
considerada la estructura tegumentaria más compleja que existe, pero por mucho
tiempo su formas más ancestral registrada en la literatura científica era la de
Archaeopteryx, un organismo
transicional entre aves y dinosaurios, que mostraba una mezcla de caracteres
interesante, pues, aunque su esqueleto carecía de casi todas las
especializaciones de las aves modernas, siendo más cercano a los dinosaurios
dromeosaurios y deinonicosaurios, sus plumas eran marcadamente modernas en
apariencia. Esto implicaba necesariamente que los dinosaurios más basales
deberían tener estructuras integumentarias variadas, algunas de las cuales
deberían ser mucho más simples y escuetas que lo que se ve en Archaeopteryx.
22.1 El modelo
clásico
Una hipótesis moderna de los pasos evolutivos que condujeron a la estructura moderna de las plumas indicaba que las plumas eran esencialmente una versión encrespada o deshilachada de las escamas de reptil. El ornitólogo Richard Prum (Prum, 1999; Prum & Brush, 2002) ha propuesto un modelo de desarrollo para la evolución de las plumas modernas (Figura 22.1).

Figura 22.1. Etapas
hipotéticas de la evolución de las plumas. (A) Etapas de desarrollo del
crecimiento de las plumas desde una perspectiva anterior del collar epidérmico.
(B) Una transición hipotética de folículos de plumas.
Sugirió que la evolución de las plumas
probablemente ocurrió a través de una serie de seis etapas, cada una de las
cuales representa versiones cada vez más complejas del folículo de plumas aviar
y su pluma resultante. La última etapa evolutiva es el folículo de plumas
moderno y la pluma moderna (aunque como hemos visto, casi todos los tipos de
pluma han sido preservados en las aves modernas, el punto es ver cómo van
apareciendo en el registro fósil); Las primeras etapas representan sus posibles
predecesores evolutivos.
👉 La etapa I habría implicado el origen del folículo de plumas más simple, un
cilindro hueco de tejido no diferenciado y homogéneo. No estaría presente
ninguna subestructura dentro del collar, como una cresta de raquis o crestas de
púas, solo un tubo simple de células que crecen queratina. Podemos imaginar que
un collar epidérmico de esta estructura crecería una columna hueca de
queratina, o una pluma tubular.
👉 La novedad evolutiva que definió la etapa II habría sido la
creación de crestas de púas. Esto requeriría la diferenciación del tejido
dentro del collar epidérmico en tiras de queratinocitos que crecen células por
separado de otras tiras dentro del collar. Tal folículo crearía un mechón de
púas no ramificadas, muy parecido al mechón de tiras de papel ondeando en el
extremo de un cuerno de fiesta con flecos. Sin una cresta de raquis formada y,
por lo tanto, sin nada para fusionar las crestas de las púas, no se formaría
raquis y la longitud de la pluma se limitaría a la longitud de sus púas constituyentes.
👉 La etapa III implica dos pasos que deben tomarse antes
de que pueda ocurrir la siguiente etapa, generando aquí los dos niveles de
estructuras de ramificación que se encuentran en las plumas modernas: púas que
se ramifican desde (y fusionadas con) el raquis y bárbulas que se ramifican
desde (fusionadas con) la púa. Por lo tanto, en la etapa III, una cresta de
raquis se habría diferenciado de tal manera que podría haberse formado una
estructura ramificada por crestas de púas que se fusionan con la cresta de
raquis. Además, la subestructura de desarrollo que produce la barbula debe
haberse formado dentro de las crestas de las púas para crear la estructura de
ramificación de la púa. Estos dos avances en el desarrollo dan como resultado
dos niveles de jerarquía de ramificación (púas y bárbulas) en la propia pluma.
👉 La etapa IV implicó el avance de la mera presencia de bárbulas a la
especialización de la estructura de las bárbulas. Para funcionar de la manera
en que lo hacen en las plumas modernas del peine, las bárbulas distales deben
tener ganchos, o barbículas, que les permitan agarrar las bárbulas proximales
de las púas vecinas y crear ese tejido especial que cierra el álabe peninoso.
Esta es la novedad hipotética de la etapa IV.
👉 Finalmente, la etapa V
se considera la "etapa de especialización", varias modificaciones del
modelo general de desarrollo generan la diversidad en la morfología de las
plumas que vemos dentro y entre las aves modernas.
22.2 Analizando
la hipótesis de Prum
Sin embargo, la evidencia que desafía esta
hipótesis proviene de cómo crecen las plumas: en lugar de desarrollarse como
estructuras en forma de placas como escamas de reptil, las plumas son
producidas por folículos de plumas, que, como hemos visto, son órganos
tubulares de desarrollo en miniatura distribuidos sobre la piel. Aunque las
plumas tienen algunas características en común con la escama de reptil (como
distribuirse regularmente a través de la piel, crecer desde la base y estar
hecha de beta-queratina), la naturaleza columnar del folículo de la pluma en sí
es una característica única. Por lo tanto, desde el punto de vista del
desarrollo, desde la creación del folículo en adelante, todos los detalles del
crecimiento de las plumas también son novedosos, sin paralelos directos en otros
animales vivos. De hecho, la disposición única del folículo significa que una
pluma se puede definir simplemente como cualquier estructura producida por un
folículo de plumas.
Esta definición puede parecer trivial dado
que, en este punto de la historia, todas las aves y solo las aves tienen
plumas, pero se vuelve importante al mirar hacia atrás a través del registro
fósil de los orígenes de las plumas en los antepasados de las aves. En las
últimas décadas, los paleontólogos que trabajan en fósiles excepcionalmente
bien conservados de la provincia de Liaoning en China han completado muchos de
los detalles sobre cómo evolucionaron las diversas características anatómicas
de las aves, incluidas las plumas, en los dinosaurios terópodos. Se han
encontrado numerosos fósiles de terópodos no aviarios con huellas claras de los
llamados "apéndices integumentarios" que rodean el cuerpo,
estructuras que cubren la piel como escamas, plumas y cabello, presumiblemente
crecidas de la piel misma, formando halos pigmentados alrededor de extremidades
fosilizadas, torsos, cabezas, cuellos y colas. Cuando se verificó que estos
apéndices estaban compuestos de beta-queratina, que se sabe que ocurre solo en
plumas de aves y escamas de reptiles, los investigadores se sintieron más cómodos
al referirse a estas estructuras delgadas y alargadas como plumas.
22.3 La
teleología de la pluma
Las primeras hipótesis acerca del origen de las plumas se
basaron en la teleología o filosofía natural, los seres vivos están
perfectamente adaptados para X cosa a través de Y partes del cuerpo; y es muy
evidente que las plumas de las aves modernas están adaptadas para generar
empuje ascendente con un esfuerzo mínimo, de allí que se pensara como algo
evidente que las plumas fueron creadas para que las aves pudieran volar y san
se acabó, ¿Qué otras preguntas podrían plantearse? (van Dijk & Reydon, 2010). Sin embargo, estas afirmaciones empiezan a desdibujarse cuando
analizamos las aves no voladoras, las cuales resultan ser comunes aun en la
actualidad, existen muchas aves modernas con plumas asimétricas y esqueletos
modificados para el vuelo, que no vuelan, ¿Dónde queda entonces que las plumas
fueron hechas para volar?
La irrupción de las ideas darwinianas de la evolución no
cambió mucho esta noción, o por decirlo de otro modo, los primeros darwinistas
eran de algún modo neoteleológicos, solo que en lugar de decir que las plumas
fueron creadas para volar, se cambió por, las plumas evolucionaron para volar.
Este tipo de pensamiento se hizo más fuerte cuando las ideas naturalistas de la
evolución se separaron en dos frentes más claros, los ortogenesistas evolutivos
que planteaban que los seres vivos evolucionaban unilinealmente para hacerse
más complejos, y por ende la idea de una escala de perfección de plumas de la
menos desarrollada y apta para el vuelo, a la más adaptada para el vuelo tenía
mucho sentido. Los neodarwinistas que rechazaron totalmente la teleología por
otra parte plantearon que las plumas tenían muchas otras funciones que, aunque
eran accesorias al vuelo en la actualidad, tal vez fueron su función principal
anteriormente, y con el tiempo, estas plumas usadas para otras cosas fueron
cooptadas para el vuelo.
La pregunta pertinente ahora es ¿Cómo resolver la pregunta?
La respuesta viene de una sola fuente, los fósiles.
22.4 Los
dinopollos
Todas las aves son dinosaurios, aunque no todos los dinosaurios son aves, es un razonamiento típico de la taxonomía linneana basada en rangos incluyentes, por ejemplo, todos los roedores son mamíferos, pero no todos los mamíferos sopn roedores. Aunque hasta aquí parece fácil de diferenciar un ave de un dinosaurio, resulta que a partir del nuevo milenio se han descubierto fósiles nuevos, y especímenes más completos de viejos conocidos que retan el modo tajante en que diferenciamos un ave de un dinosaurio (Figura 22.2).

Figura 22.2. Dendrograma de los dinosaurios
saurisquios “caderas de dinosaurio o más correctamente, dinosaurios cuyos
ancestros tienen el hueso isquion hacia abajo”, que muestra las relaciones de
grupos de dinosaurios, especialmente terópodos “depredadores bípedos” y de
ellos los coelurosaurios “colas rígidas”. En este dendrograma se muestra la
diversidad y la gradualidad entre las formas fósiles, colocar una frontera
entre dinosaurio y ave en algún punto seria arbitraria (Zimmer, 2010).
Sin duda un punto de partida es el legendario Archaeopteryx, pero este espécimen aún se encuentra a mucho camino de ser un ave moderna
como la entendemos, y además sus plumas son ya en cierto sentido, modernas y
adaptadas para generar elevación al ser asimétricas. Sin embargo, él fue el
punto de partida para el entendimiento de la evolución de las plumas, y de una
u otra manera, nuestra narrativa inicia con él y sus parientes cercanos.

Figura 22.3. Las aves modernas como las entendemos e imaginamos son
las llamadas Neornithes “aves nuevas”, en este dendrograma que analiza a
las aves que hay entre Neornithes y Archaeopteryx, podemos ver
que la posición de este último es extremadamente basal, pues lo único
medianamente moderno en el son sus plumas, el resto de su esqueleto puede
denominarse de cierto modo, primitivo (Wang et al., 2015).
La visión funcional sobre la evolución de las plumas se ha
centrado tradicionalmente en el aislamiento, el vuelo y la selección sexual.
Los descubrimientos de dinosaurios emplumados del Cretáceo Tardío no voladores
en China (Brusatte, O’Connor, & Jarvis,
2015), sin embargo,
sugieren que el vuelo no pudo haber sido la función principal original ya que
las plumas simplemente no habrían sido capaces de proporcionar ninguna forma de
elevación al ser simétricas y endebles (Dimond, Cabin, & Brooks, 2011;
Sumida & Brochu, 2000).
Se han sugerido que las plumas pueden haber tenido su función original en
termorregulación, impermeabilización o incluso como sumideros de desechos
metabólicos como el azufre (Bock, 2000).
Se argumenta que los descubrimientos recientes respaldan una función termorreguladora, al menos en los dinosaurios más pequeños (Xu et al., 2012). Si bien se ha sugerido que las plumas evolucionaron a partir de escamas de reptil, hay numerosas objeciones a esa idea, y han surgido explicaciones más recientes del paradigma de la biología evolutiva del desarrollo (Prum & Brush, 2003, 2014). Las hipótesis de los orígenes de las plumas basadas en la escama sugieren que la estructura de escama plana se modificó para convertirse en plumas al dividirse para formar la cinta; sin embargo, ese proceso de desarrollo involucra una estructura tubular que surge de un folículo y el tubo se divide longitudinalmente para formar la cinta. El número de plumas por unidad de área de piel es mayor en aves más pequeñas que en aves más grandes, y esta tendencia apunta a su importante papel en el aislamiento térmico, ya que las aves más pequeñas pierden más calor debido a la superficie relativamente más grande en proporción a su peso corporal (Lovette & Fitzpatrick, 2016).

Figura 22.4. Representación
de Anchiornis huxleyi, empleando el esqueleto fósil que posee casi toda
la silueta de su integumento y los colores obtenidos mediante la identificación
de la estructura de sus melanosomas. Este es quizá uno de los dinosaurios cuya
imagen podemos reconstruir con mayor precisión a pesar de las eras que nos
separan de su especie.
Se cree que la
coloración de las plumas ha evolucionado principalmente en respuesta a la selección sexual. En un espécimen fósil de Parave Anchiornis huxleyi, las características
están tan bien conservadas que se puede observar la estructura del melanosoma
(células de pigmento). Al comparar la forma de los melanosomas fósiles con los
melanosomas de las aves existentes, se pudo determinar el color y el patrón de
las plumas en Anchiornis (Vinther, 2017, 2018). Se descubrió que Anchiornis
tenía plumas con dibujos en blanco y negro en las extremidades anteriores y
posteriores, con una cresta de color marrón rojizo. Este patrón es similar a la
coloración de muchas especies de aves existentes, que utilizan la coloración
del plumaje para su exhibición y comunicación, incluida la selección sexual y
el camuflaje. Es probable que las especies de dinosaurios no aviarios utilicen
patrones de plumaje para funciones similares a las aves modernas antes del
origen del vuelo. En muchos casos, la condición fisiológica de las aves
(especialmente los machos) está indicada por la calidad de sus plumas, y esto
es utilizado (por las hembras) en la elección de la pareja.
Varios dinosaurios no aviarios tenían plumas en sus extremidades que no habrían funcionado para el vuelo (Brusatte, O’Connor, & Jarvis, 2015). Una hipótesis sugiere que las plumas evolucionaron originalmente en los dinosaurios como resultado de sus propiedades de aislamiento, pero en las pequeñas especies de dinosaurios en que algunas de sus plumas crecieron más; estas pueden haberlas encontrado útiles para planear, lo que ha llevado a la evolución de protoaves como Archaeopteryx y Microraptor zhaoianus.

Figura 22.5. Yutyrannus huali es uno de los dinosaurios no aviares emplumados más grandes que conocemos, sus plumas eran estructuras mucho más simples, siendo más filamentosas.

Figura 22.6. Microraptor es un dromeosaurio que evolucionó en paralelo a las aves, de el también se conoce casi todo su tegumento y su color que era un azul oscuro metalizado, semejante al que tienen los machos de las palomas en su pecho.

Figura 22.7. Dilong
paraduxus posee en su fósil marcas tegumentarias de plumas filamentosas, se
trata de un pariente del ancestro del T.rex, por lo que es un dinosaurio
emplumado no aviar.
Otra hipótesis postula que la ventaja adaptativa original de
las primeras plumas era su pigmentación o iridiscencia, lo que contribuye a la
preferencia sexual en la selección de pareja (Dimond,
Cabin, & Brooks, 2011). Los dinosaurios
que tenían plumas o protoplumas incluyen Pedopenna
daohugouensis (Xu
& Zhang, 2005) y Dilong paradoxus, un tirannosauroide que
es de 60 a 70 millones de años más viejo que Tyrannosaurus rex (Xu
et al., 2004) aunque también
existieron dinosaurios grandes con plumas que habitaban regions templadas con
in viernos severos como el Yutyrannus
huali (Xu
et al., 2012).
22.5 Más allá
de los terópodos
La mayoría de los dinosaurios que se sabe
que han tenido plumas o protoplumas son terópodos, y por mucho tiempo se pensó
que la pluma era un carácter exclusivo de estos animales, lo cual planteaba
muchas paradojas, la principal era el hecho de que exhibían estructuras muy
avanzadas, por lo que seguía pareciendo como si la forma básica de la pluma
hubiera aparecido desde la nada, sin embargo, las "estructuras
integumentarias filamentosas" similares a plumas también se conocen por
los dinosaurios ornitisquios Tianyulong
y Psittacosaurus (Zheng, You, Xu, & Dong, 2009). La naturaleza exacta de estas estructuras aún está en estudio.
Sin embargo, se cree que las plumas de la etapa 1 “filamento simple” como las
que se ven en estos dos ornitisquios probablemente funcionaron en la exhibición
sexual (X Xu & Guo, 2009). En 2014, se informó que el ornitisquio Kulindadromeus tenía estructuras que se asemejaban a plumas en
etapa 3 “filamentos simples unidos a un raquis elemental” (Godefroit et al., 2014).
22.6 Clasificación
de las plumas por su complejidad evolutiva.
Varios estudios sobre el desarrollo de
plumas en los embriones de aves modernas, junto con la distribución de los
tipos de plumas entre varios precursores de aves prehistóricas, han permitido a
los científicos intentar una reconstrucción de la secuencia en la que las
plumas evolucionaron y se desarrollaron en los tipos encontrados en las aves
modernas. Esta clasificación está centrada en las aves modernas, pero no indica
que las plumas más recientes sean más perfectas. La evolución de las plumas fue
dividida en las siguientes etapas por Xu y Guo en 2009 (X Xu & Guo, 2009) (Figura 22.8):
👉 Filamento simple
👉 Múltiples filamentos unidos en su base
👉 Múltiples filamentos unidos en su
base a un filamento central (Filopluma).
👉 Múltiples filamentos a lo largo de
un filamento central
👉 Múltiples filamentos que surgen del borde de una estructura
membranosa
👉 Pluma pennacea con veleta de púas
y bárbulas y raquis central
👉 Pluma pennacea con un raquis
asimétrico
👉 Veleta indiferenciada con raquis
central
Sin embargo, Foth (2011) demostró que algunas de estas supuestas etapas (etapas 2 y 5 en particular) probablemente sean simplemente artefactos de conservación causados por la forma en que las plumas fósiles se aplastan y las plumas permanecen o se conservan las huellas. Foth reinterpretó las plumas de la etapa 2 como plumas trituradas o mal identificadas de al menos la etapa 3, y las plumas de la etapa 5 como plumas trituradas de la etapa 6.

Figura 22.8. Clasificación
de las plumas por su complejidad creciente, (Nota, las etapas 2 y 5 se
encuentran en debate, debido a que los fósiles que las presentan están mal
preservados y posiblemente son artefactos debido al aplastamiento de la pluma.
23. Evolución de las plumas 2, los pterosaurios
|| Regresar al Índice ||
Los pterosaurios no son dinosaurios, sino un linaje de reptiles que se especializaron en un modo diferente de vuelo, con un ala derivada de su dedo meñique alargado. ¿Por qué hablar de ellos? Resulta que el tegumento de sus pieles no era escamoso, como tienden a ser representados en el paleoarte anterior a 2010, estamos trat6ando con un terciopelo análogo al pelo de los mamíferos o posiblemente relacionado fundamentalmente con las plumas. Esta complicación de categorías se debe a que estamos tratando con evolución paralela y convergente, es posible que las escamas de los reptiles tengan la tendencia a desarrollar independientemente estructuras semejantes con anatomía muy parecida.

Figura 23.1. Reconstrucción
paleoartística de Scaphognathus crassirostris.
Al menos algunos pterosaurios tenían filamentos peludos conocidos como picnofibras en la cabeza y el cuerpo, similares, pero no homólogos (que comparten un origen común) con el pelo de los mamíferos. Goldfuss reportó por primera vez un tegumento difuso de un espécimen de Scaphognathus crassirostris en 1831 (Witton, 2013), y los hallazgos recientes de pterosaurios y la tecnología para el examen histológico y ultravioleta de especímenes de pterosaurios han proporcionado una prueba incontrovertible: los pterosaurios tenían capas de microfibra. Las picnofibras no eran el pelo verdadero como se ve en los mamíferos, sino una estructura única que desarrolló una apariencia similar “evolución convergente, analogía evolutiva”. Aunque, en algunos casos, las actinofibrillas (fibras estructurales internas) en la membrana del ala se han confundido con picnofibras o cabello verdadero, algunos fósiles, como los de Sordes pilosus (que se traduce como "demonio peludo") y Jeholopterus ninchengensis, muestran la huellas inconfundibles de picnofibras en la cabeza y el cuerpo, a diferencia de los murciélagos modernos, otro ejemplo de evolución convergente (Unwin & Bakhurina, 1994). Los abrigos para la cabeza no cubren las grandes mandíbulas del pterosaurio en muchos de los especímenes encontrados hasta ahora (Witton, 2013).

Figura 23.2. Fósil y reconstrucción paleoartística de Sordes pilosus.

Figura 23.3. Modelo del
fósil y reconstrucción paleoartística de Jeholopterus ninchengensis.
La presencia de picnofibras (y las
demandas de vuelo) implican que los pterosaurios eran endotérmicos (de sangre
caliente). La ausencia de picnofibras en las alas del pterosaurio sugiere que
el pelaje no tenía una función aerodinámica, lo que respalda la idea de que las
picnofibras evolucionaron para ayudar a la termorregulación del pterosaurio,
como es común en los animales de sangre caliente, siendo necesario el
aislamiento para conservar el calor creado por un metabolismo endotérmico (Witton, 2013). El
"pelo" del pterosaurio era tan claramente distinto del pelaje de los
mamíferos y otros integumentos de los animales, por lo que requería un nombre nuevo
e independiente. El término "picnofibra", que significa
"filamento denso", fue acuñado por primera vez en un documento sobre
las impresiones de tejidos blandos de Jeholopterus
por el paleontólogo Alexander W.A. Kellner y sus colegas en 2009 (Kellner et al., 2009). La investigación sobre el código genético de los embriones de
cocodrilo americano podría sugerir que las picnofibras, los escudos de
cocodrilo y las plumas de ave son homólogas en el desarrollo, según la
construcción de su beta-queratina (Alibardi, Knapp, & Sawyer, 2006), lo cual ratifica lo difícil que es distinguir un órgano homólogo
de uno análogo cuando este ha evolucionado de una misma estructura base en un
ancestro común cercano.
23.1¿Plumas
en pterosaurios?
Algunos investigadores (Czerkas y Ji, 2002)
han especulado que las picnofibras eran un antecedente de las protoplumas, pero
las impresiones disponibles de los integumentos de pterosaurios no son como las
"plumas" que se encuentran en muchas de las muestras de maniraptores
emplumados en el registro fósil (Witton, 2013). Las picnofibras de los pterosaurios se estructuraron de manera
similar a las protoplumas de los terópodos (Kellner et al., 2009). Las picnofibras eran filamentos cortos y flexibles, "solo de
5–7 mm en algunas muestras" y bastante simples, "aparentemente sin
ningún detalle interno aparte del canal central" (Witton, 2013). Las "pieles" de pterosaurio encontradas
"preservadas en esteras concentradas y densas de fibras, similares a las
que se encuentran alrededor de los mamíferos fosilizados" sugieren capas
con un espesor comparable al de muchos mamíferos mesozoicos, (Witton, 2013) al menos en las partes del pterosaurio cubiertas de picnofibras.
El grosor del pelaje y el área de superficie cubierta, definitivamente variaron
según las especies de pterosaurios.
Un estudio de 2018 de los restos de dos
pequeños pterosaurios de la edad jurásica de Mongolia Interior, China,
descubrió que los pterosaurios tenían una amplia gama de formas y estructuras
de fibras tegumentarias, a diferencia de la estructura homogénea que
generalmente se pensaba que poseían. El análisis encontró que al menos algunos
de estos tenían una estructura muy similar a 4 estructuras de plumas diferentes
comúnmente conocidas de aves y dinosaurios, pero casi nunca se conocían de
pterosaurios antes del estudio.
Sin embargo, Yang et al. (2011) presentan
nueva evidencia que sugiere que debemos considerar seriamente la posibilidad de
un origen evolutivo temprano de las plumas. La evidencia más convincente
presentada en su estudio es que las estructuras filamentosas integumentarias
preservadas en dos fósiles de pterosaurios muestran una característica clave de
las plumas: la ramificación en forma de pluma. Algunas estructuras
integumentarias fosilizadas en estas muestras de pterosaurios están compuestas
de múltiples filamentos unidos en su base, y algunas comprenden múltiples
filamentos unidos a un filamento central. Esas morfologías sugieren la
presencia de púas y un raquis (componentes principales de una pluma) fuera de Dinosauria. Además, los autores han
demostrado que estas estructuras integumentarias filamentosas de pterosaurios
probablemente también representan las primeras plumas desde una perspectiva
filogenética, apoyando así la hipótesis de origen profundo para la evolución de
las plumas (Xu, 2019).
Se desconoce si estas "plumas"
comparten una ascendencia común con las plumas de las aves “son homólogas” o si
evolucionaron independientemente de ellas “son análogas”, pero el autor
principal del estudio, Mike Benton, ha declarado que, a pesar del análisis
exhaustivo, prácticamente no hubo diferencias registradas entre las estructuras
de las aves o los dinosaurios, y por lo tanto es probable que compartan una
ascendencia común con ellos (Yang et al., 2019). Esto implicaría que la evolución de la pluma debe abarcar e
incluso ir más atrás que el ancestro común entre los pterosaurios, los
dinosaurios y las aves (Witton, 2013), aunque esta conclusión aun es tentativa, y tendremos que esperar
a los descubrimientos de la próxima década para ahondar más en esta revolucionaria
hipótesis.
23.2 La imagen
moderna de la evolución de las plumas
Juntos, estos fósiles contribuyen a una
imagen coherente de la evolución de las plumas que ocurre en varios pasos a
través de muchos millones de años. La mayor sorpresa ha sido cuán temprano ocurrieron
las etapas evolutivas / de desarrollo hipotéticas en relación con el origen de
las aves. Hasta hace poco, pocos investigadores cuestionaban la idea de que las
plumas están, y siempre han estado, limitadas a las aves. De hecho, las plumas
han sido consideradas durante mucho tiempo una característica definitoria de
las aves, como lo es el pelo para los mamíferos. Pero ahora, se han encontrado
plumas filamentosas relativamente simples, aquellas sin la estructura
secundaria y terciaria de púas y bárbulas, en algunos de los primeros
dinosaurios terópodos fósiles que vivieron mucho antes de que evolucionaran las
primeras aves, y también en dinosaurios no terópodos y hasta en no-dinosaurios,
por lo que la evolución de la pluma parece enterrarse cada vez más profundo en
el tiempo, lo cual da sentido al hecho de que la pluma del Archaeopteryx sea
tan moderna en apariencia.
Además del origen sorprendentemente antiguo de las plumas primitivas, las plumas que parecen completamente modernas también son anteriores al origen de las aves. Por ejemplo, Mark Norell et al. (2002) publicaron evidencia de una pluma pennacea moderna de un grupo de dinosaurios terópodos, el Microraptor zhaoianus (Figura 23.4). Se han encontrado otros dinosaurios que exhiben una diversidad de formas de plumas en todo el cuerpo, que se asemeja a la diversidad de formas de plumas que se encuentran en todo el cuerpo de un ave individual en la actualidad, con plumas distintivas de plumón, cuerpo y ala y / o cola. Un descubrimiento fósil particularmente emocionante fue el de un dinosaurio de "cuatro alas", Microraptor gui (Xu et al. 2003), en el que las plumas alargadas que aparecen como las plumas de vuelo modernas son visibles a lo largo de la pata trasera.

Figura 23.4. Este es el ya clásico fósil de
Microraptor, mostrando las huellas inconfundibles de plumas de vuelo.
Más evidencia de plumas de aspecto moderno
en dinosaurios pre-aviarios incluye un impresionante conjunto de publicaciones
que relacionan la apariencia visual de las aves y sus plumas con las de las
aves modernas. Los investigadores han encontrado evidencia fósil de que, como
en las aves modernas, las plumas antiguas tenían pigmentos marrones y negros en
su interior, incluidos los arreglos de estos pigmentos que podrían indicar la
presencia de coloración iridiscente (Vinther et al. 2008, 2010; Zhang et al.
2010). Estos descubrimientos se han utilizado para reconstruir los patrones de
plumaje de un puñado de dinosaurios emplumados. Por ejemplo, un estudio
reconstruyó un patrón de coloración sorprendente que podría haber existido en
el dinosaurio emplumado Anchiornis
huxleyi (Li et al. 2010).
La función original de las plumas, y su
papel en el origen del vuelo, ha sido un tema polémico durante más de un siglo.
Estos descubrimientos recientes de plumas en dinosaurios no aviarios han
anulado muchos de los argumentos más antiguos sobre por qué las plumas
evolucionaron en primer lugar, además del clásico argumento teleológico, ni la
pluma ni el brazo del ave fueron diseñados para volar originalmente. El origen
muy antiguo de las plumas “incluso más antiguo que los propios dinosaurios”, en
lo que parecen ser formas muy simples y sin ramificaciones, argumenta en contra
de las plumas que se originan para funciones aerodinámicas, ya que se requiere
una veleta pennácea para que las plumas ayuden en el vuelo. En cambio, la
variedad de formas de plumas vistas en muchos linajes de dinosaurios indica que
las plumas originalmente no evolucionaron para volar; en cambio, parecen
haberse diversificado rápidamente de cualquiera que sea su función original
para adaptarse a muchas funciones, y una de ellas casualmente le pegó a la aerodinámica,
la cual hizo muy exitoso a su portador. Es decir, como en las aves modernas,
las plumas probablemente desempeñaron una variedad de funciones que van desde
la cobertura protectora hasta la publicidad social sexual o el camuflaje, con
sus funciones aerodinámicas o de vuelo que solo evolucionan más tarde, más
cerca en el tiempo de la radiación evolutiva de las aves mismas con los
dromeosaurios y los deinonicosaurios.
24. La piel de los mamíferos
|| Regresar al Índice ||
Como en otros vertebrados, las dos capas principales de la piel de los mamíferos son la epidermis y la dermis, que se unen e interactúan a través de la membrana basal. Debajo se encuentra la hipodermis, o fascia superficial, compuesta de tejido conectivo y grasa.

Figura 24.1. Piel de mamífero. La epidermis se
diferencia en distintas capas. Como en otros vertebrados, el más profundo es el
estrato basal, que a través de la división mitótica produce células que a
medida que envejecen se convierten sucesivamente en parte del estrato espinoso,
el estrato granuloso, a menudo el estrato lúcido y finalmente el estrato córneo
superficial. La dermis resalta las papilas dérmicas que le dan a la epidermis
suprayacente una apariencia ondulante. Las glándulas sudoríparas, los folículos
pilosos y los receptores sensoriales se encuentran dentro de la dermis. Tenga
en cuenta que los conductos de sudor pasan a través de la epidermis
suprayacente para liberar sus secreciones acuosas en la superficie de la piel.
24.1 Epidermis
La epidermis puede estar localmente especializada
como cabello, uñas o glándulas. Las células epiteliales de la epidermis son
queratinocitos y pertenecen al sistema de queratinización que forma la capa
cornificada superficial y muerta de la piel. Las células queratinizadas de la
superficie se exfolian continuamente y se reemplazan por células que surgen
principalmente de la capa más profunda de la epidermis, el estrato basal. Las
células dentro de la base se dividen mitóticamente, produciendo algunas que
permanecen para mantener la población de células madre y otras que son
empujadas hacia afuera. A medida que se desplazan a niveles más altos, pasan
por etapas de queratinización exhibidas como capas distintas y sucesivas hacia
la superficie: estrato espinoso, estrato granuloso, a menudo un estrato lúcido
y un estrato córneo (Figura 24.1). El
proceso de queratinización es más distintivo en las regiones del cuerpo donde
la piel es más gruesa, como en las plantas de los pies. En otros lugares, estas
capas, especialmente el lúcido, pueden ser menos evidentes.
Los queratinocitos son el tipo celular más
prominente de la epidermis. Se reconocen otros tipos, aunque sus funciones son
menos conocidas. Las células de Langerhans son células estrelladas dispersas
individualmente en las partes superiores del estrato espinoso. La evidencia
actual sugiere que juegan un papel en las acciones del sistema inmune mediadas
por células. Se cree que las células de Merkel, que se originan en la cresta
neural y se asocian con los nervios sensoriales cercanos, responden a la
estimulación táctil (mecanorreceptores). Además de estos tipos de células
epiteliales, el otro tipo de célula prominente que se asocia secundariamente
con la epidermis es el cromatóforo. Los cromatóforos surgen de las células de
la cresta neural embrionaria y se pueden encontrar en casi cualquier parte del
cuerpo. Los que alcanzan la piel ocupan sitios dentro de las partes más
profundas de la epidermis. Secretan gránulos del pigmento melanina, que pasan
directamente a las células epiteliales y finalmente se transportan al estrato
córneo o a los tallos del cabello. El color de la piel resulta de una
combinación del estrato córneo amarillo, los vasos sanguíneos subyacentes rojos
y los gránulos de pigmento oscuro secretados por los cromatóforos.
24.2 Dermis
La dermis de los mamíferos es de doble capa. La capa papilar externa empuja proyecciones en forma de dedos, llamadas papilas dérmicas, hacia la epidermis suprayacente.

Figura 24.2. Tractos capilares. El
cabello crece con un grano, una dirección particular en la que se inclina.
Observe las diversas direcciones de crecimiento (flechas) en las que se
encuentran los pelos del bandicoot marsupial.
La capa reticular más profunda incluye
tejido conectivo fibroso dispuesto irregularmente y ancla la dermis a la fascia
subyacente. Los vasos sanguíneos, los nervios y el músculo liso ocupan la
dermis, pero no alcanzan la epidermis. La dermis de los mamíferos produce
huesos dérmicos, pero estos contribuyen al cráneo y la cintura escapular y rara
vez forman escamas dérmicas en la piel. Una excepción es Glyptodon, un mamífero fósil cuya epidermis estaba recubierta por
hueso dérmico. Una situación similar existe en el armadillo vivo. Estas
especies representan desarrollos secundarios del hueso dérmico en el
integumento mamífero. Los vasos sanguíneos y los nervios ingresan a la dermis.
Los folículos pilosos y las glándulas se proyectan hacia adentro desde la
epidermis (Figura 24.2). La
dermis generalmente está compuesta de tejido conectivo fibroso de disposición
irregular que a menudo está impregnado con fibras elásticas para darle algo de
estiramiento, pero devolverlo a su forma original. A medida que una persona
envejece, esta elasticidad se pierde y la piel se hunde.
24.3 Evolución
del cabello
Las impresiones de la piel fósil del
Jurásico Medio dan evidencia de la presencia de pelo, y presumiblemente de
mamíferos que lo poseían. Pero el valor adaptativo inicial del cabello sigue
siendo especulativo, es decir, sabemos que los protomamíferos tenían pelo, pero
no sabemos su función original.
Una visión sostiene que el cabello surgió
inicialmente como aislamiento de la superficie, reteniendo el calor del cuerpo
en los protomamíferos débilmente endotermos. La presencia de cornetes nasales
en los sinapsidos, antes del Pérmico Tardío, sugiere una endotermia temprana y,
por lo tanto, un papel para aislar para el cabello.
Una visión alternativa es que el cabello evolucionó primero como pequeñas barras salientes en las bisagras entre las escamas y sirvió como dispositivos táctiles. Estos "protopelos sensoriales" podrían ayudar a monitorear los datos sensoriales de la superficie cuando un animal se escondía de un enemigo o se retiraba del clima. Si tal papel aumentara en importancia, habría favorecido ejes más largos y tal vez la evolución de estructuras que se asemejan a vibrisas. Este protopelo sensorial podría haber evolucionado secundariamente en un pelaje aislante a medida que los mamíferos se volvieran endotérmicos. Aunque aislante en los mamíferos modernos, el cabello aún conserva una función sensorial.

Figura 24.3. Fauna del
pérmico, entre los que destaca Estemmenosuchus uralensis en el centro.
Al ser suave y descomponible, el cabello
no deja un rastro confiable en el registro fósil. Algunos terápsidos,
antepasados de los mamíferos, tienen pequeños hoyos en la región facial de
sus cráneos. Estos hoyos se parecen a los hoyos en los cráneos asociados con
vibrisas sensoriales en los mamíferos modernos como en grandes felinos. Algunos
han interpretado estos hoyos como evidencia indirecta de cabello en los
terápsidos. Pero los cráneos de algunos lagartos modernos con escamas tienen
hoyos similares y, por supuesto, los lagartos no tienen pelo. Por lo tanto,
estos pozos no son evidencia concluyente de la presencia de cabello. Además,
una impresión en la piel especialmente bien conservada de Estemmenosuchus, una terápsido del Pérmico Superior, no muestra
evidencia de pelo. La epidermis era lisa, sin escamas e indiferenciada, aunque
estaba provista de glándulas. En la vida, su piel era probablemente suave y
flexible. Por lo tanto, todavía no sabemos cuándo surgió el pelo por primera
vez en mamíferos primitivos o en sus ancestros terápsidos.
24.4 Glándulas
Principalmente hay tres tipos principales
de glándulas integumentales en mamíferos: sebáceas, ecrinas y apocrinas. El
olor, el sudor y las glándulas mamarias se derivan de ellos. Las glándulas
sebáceas son de forma globular o de saco; Las glándulas ecrinas y apocrinas son
invaginaciones largas y enrolladas de la epidermis que alcanzan las
profundidades de la dermis, pero mantienen la continuidad a través de la
superficie de la piel incluso a través del estrato córneo cornificado.
👉 Glándulas subcutáneas: Producen una secreción aceitosa, sebo, que se libera en los
folículos capilares para acondicionar y ayudar a la piel impermeable. Las
glándulas sebáceas están ausentes de las palmas de las manos y las plantas de
los pies, pero están presentes, sin vello asociado, en el ángulo de la boca, en
el pene, cerca de la vagina y al lado de los pezones mamarios. En estos sitios,
su secreción lubrica la superficie de la piel. Las glándulas de cera del canal
auditivo externo, que secretan cera, y las glándulas meibomianas del párpado,
que secretan una película aceitosa sobre la superficie del globo ocular,
probablemente derivan de las glándulas sebáceas.
👉 Glándulas Ecrinas: Producen fluidos acuosos y delgados, no están asociados con los
folículos capilares, comienzan a funcionar antes de la pubertad y están
inervados principalmente por nervios colinérgicos. En la mayoría de los
mamíferos, estas glándulas están asociadas con las plantas de los pies y las
manos, las colas prensiles y otros sitios en contacto con superficies
abrasivas. Los chimpancés y los humanos tienen la mayor cantidad de glándulas
ecrinas, incluidas algunas en las palmas y las plantas. En el ornitorrinco,
estas glándulas se limitan al hocico. En ratones, ratas y gatos, están
presentes en las patas y aparecen alrededor de los labios en conejos. En los
elefantes, las glándulas ecrinas y las glándulas sebáceas están ausentes por
completo.
👉 Glándulas apocrinas: Producen un fluido viscoso que contiene lípidos, están asociados
con los folículos pilosos, comienzan a funcionar en la pubertad y están inervados
principalmente por nervios adrenérgicos. Sus secreciones funcionan
principalmente en la señalización química. La evaporación de la superficie de
los productos de estas glándulas ayuda a disipar el calor, en donde se
denominan "glándulas sudoríparas". Sin embargo, las glándulas
sudoríparas no se encuentran en todos los mamíferos, y de hecho los humanos son
excepcionales en nuestro uso perfuso de enfriamiento evaporativo para
termorregular. Nuestras glándulas sudoríparas se derivan de las glándulas ecrinas,
pero las de los caballos derivan de las glándulas apocrinas, un ejemplo de
evolución convergente. El sudor también contiene productos de desecho; por lo
tanto, el tegumento representa una vía para la eliminación de subproductos
metabólicos.
👉 Glándulas de olor: Se derivan de las glándulas apocrinas y producen secreciones que juegan un papel en la comunicación social. Estas glándulas pueden ubicarse en casi cualquier parte del cuerpo, como en el mentón (algunos venados, conejos), la cara (venados, antílopes, murciélagos), la región temporal (elefantes), el pecho y los brazos (muchos carnívoros), la región anal (roedores, perros, gatos, mustelidos), vientre (ciervo almizclero), espalda (ratas canguro, pecaríes, camellos, ardillas de tierra) o patas y pies (muchos ungulados). Las secreciones de estas glándulas se utilizan para marcar el territorio, identificar al individuo y comunicarse durante el cortejo.

Figura 24.4. Glándulas mamárias. El tejido mamario
glandular derivado del tegumento se encuentra en la dermis, pero los conductos
llegan a la superficie a través de la epidermis. Las glándulas mamarias están
dispuestas en lóbulos, cada lóbulo es una colección de alvéolos y sus conductos
inmediatos. (a) En los monotremas, las glándulas mamarias se abren directamente
a la superficie de la piel no especializada, y las crías presionan su hocico en
forma del parche de piel donde se abren estas glándulas. (b) En algunos
marsupiales y en muchos mamíferos placentarios, los conductos mamarios se abren
a través de especializaciones del tegumento. La tetina es una especialización
tubular de la epidermis que se expande en su base hacia la cisterna, una cámara
que recibe leche de las glándulas mamarias. antes de pasarlo por el conducto
del pezón común al lactante. (c) El pezón es una papila epidérmica elevada
alrededor de la cual los suaves labios del bebé encajan directamente para beber
la leche liberada.
24.5 Glándulas
mamarias
Producen leche, una mezcla acuosa de
grasas, carbohidratos y proteínas que nutre a los jóvenes. Las crestas mamarias
ectodérmicas, dentro de las cuales se forman las glándulas mamarias, se
encuentran a lo largo del lado ventrolateral del embrión. El número de
glándulas mamarias varía entre especies. La liberación de leche a un lactante
es la lactancia. Se ha informado de lactancia en machos de una especie de
murciélago frugívoro de Malasia, lo que abre la posibilidad de que estos machos
murciélagos realmente puedan amamantar a las crías que crían. Fuera de este
murciélago frugívoro, la lactancia se ha informado solo en animales machos
domesticados, el resultado probable de endogamia anormal y en condiciones
patológicas. Con estas excepciones, las glándulas mamarias se vuelven
funcionales solo en las hembras. Las glándulas mamarias consisten en numerosos
lóbulos. Cada lóbulo es un grupo de alvéolos secretores en los que se produce
la leche. Los alvéolos pueden abrirse en un conducto común que, a su vez, puede
abrirse directamente a la superficie a través de una papila epidérmica elevada
o pezón. El pezón generalmente está rodeado por un área circular de piel
pigmentada llamada areola. Los conductos alveolares también pueden abrirse en
una cámara común, o cisterna, dentro de un largo collar de epidermis llamado
tetina. La tetina forma un conducto secundario que transporta la leche desde la
cisterna hasta la superficie (Figura 24.4). El
tejido adiposo puede acumularse debajo de las glándulas mamarias para producir
senos.
👉 Lactancia en
monotremas: En los monotremas, los pezones y
las tetas están ausentes, y no se forman senos. La leche se libera de los
conductos al parche de leche aplanado, o areola, en la superficie de la piel (Figura 24.4a).
La parte frontal del hocico del bebé está conformada para adaptarse a la
superficie, lo que permite una succión vigorosa. En breves períodos de
lactancia de 20 a 30 minutos, un equidna joven puede tomar leche equivalente a
aproximadamente el 10% de su peso corporal.
👉 Lactancia en los
terianos: Los terianos que agrupan a los
marsupiales y mamíferos euterianos, hay pezones y tetas (Figura 24.4b,
c). En la madurez sexual, el tejido adiposo se acumula debajo de la glándula
mamaria para producir el seno. El agrandamiento de las glándulas mamarias
ocurre bajo estimulación hormonal poco antes del nacimiento de las crías. La
succión estimula una respuesta neural al sistema nervioso que resulta en la
liberación de oxitocina, la hormona que estimula la contracción de las células
mioepiteliales que envuelven los alvéolos y, por lo tanto, se libera leche, así
como un comportamiento de apego por las crías.
👉 Evolución de la lactancia: El origen de la lactancia en los mamíferos sigue siendo un tema complejo. Los primeros mamíferos, con quizás solo una excepción (Sinoconodon), tenían dentición difiodonta (dientes de "leche" y dientes permanentes) como en los mamíferos lactantes modernos, en comparación con la dentición por polifodonta (reemplazo dental continuo) en la mayoría de los otros vertebrados. La difiodontia implica la presencia de glándulas mamarias y succión porque la succión para la nutrición, en lugar de buscar comida dura, permite un crecimiento sustancial del cráneo antes de que los dientes necesiten procesar la comida dura.

Figura 24.5. Reconstrucción
artística de Sinoconodon.
Las similitudes detalladas de las
glándulas mamarias en monotremas, marsupiales y eutherianos vivos defienden un
origen monofilético de estas glándulas, quizás por la combinación de partes de
glándulas sebáceas y apocrinas preexistentes. Esto ha llevado a Daniel
Blackburn a plantear la hipótesis de una serie de pasos en la evolución de la
lactancia de mamíferos, comenzando en los antepasados que incubaron huevos o
crías.
👉 Las secreciones de las glándulas
de la piel de las hembras que tenían propiedades antimicrobianas protegerían la
superficie de las crías de bacterias, hongos u otros patógenos. La ingestión de
estas secreciones en pequeñas cantidades por las crías reduciría los efectos
patológicos y los microorganismos en el tracto digestivo, esto aún sigue siendo
verdad con la lactancia mamífera, las crías de mamífero ingieren el sistema
inmune de la madre, y este les protege por un tiempo. Si esta secreción
incluyera inmunoglobulinas maternas, esto también entregaría inmunidad
directamente a la descendencia.
👉 Cualquier valor nutricional
incluido con estas secreciones de glándulas mamarias ancestrales sería de mayor
importancia adaptativa.
En tal escenario, las glándulas de la piel
materna en parche de incubación primero proporcionan protección contra los
patógenos y luego, en segundo lugar, se involucran en la nutrición de los
jóvenes. La evolución posterior incluyó la mejora de la secreción en un flujo
abundante que era mucho más nutritivo (es decir, leche). El aumento en el
contenido de nutrientes de la leche reemplazó el gran suministro de yema como
base para el soporte del embrión y, por ende, favorecería la simplificación de
los sacos embrionarios que se ve en los mamíferos. Esto se vio favorecido aún
más por una mayor eficiencia en la alimentación infantil (lactancia),
especializaciones anatómicas (tetinas en los terianos) y control fisiológico de
la producción y liberación de leche (hormonas).
25. El pelo de los mamíferos
|| Regresar al Índice ||
Los pelos son filamentos delgados y
queratinosos. La base de un cabello es la raíz. Su longitud restante constituye
el eje no vivo. La superficie externa del eje a menudo forma una cutícula
escamosa. Debajo de esto está la corteza del cabello, y en su núcleo está la
médula del cabello (Figura 24.1). El
tallo del cabello se proyecta sobre la superficie de la piel, pero se produce
dentro de un folículo epidérmico enraizado en la dermis. La superficie de la
epidermis se sumerge en la dermis para formar el folículo piloso. En su base
expandida, el folículo recibe un pequeño mechón de la dermis, la papila
capilar. Esta papila parece estar involucrada en la actividad estimulante de
las células de la matriz de la epidermis, pero no contribuye directamente al
tallo del cabello. El pequeño grupo de células matriciales vivas, como el resto
del estrato basal, es la región germinal que inicia el proceso de
queratinización para producir cabello dentro del folículo. A diferencia de la
queratinización dentro de la epidermis, que es general y continua, la
queratinización dentro del folículo piloso es localizada e intermitente.
25.1 El ciclo
de crecimiento del cabello
El tallo del pelo crece fuera del folículo piloso vivo, que pasa por un ciclo de actividad con tres etapas: crecimiento, degeneración, descanso. Durante el crecimiento, hay una proliferación activa de células en la papila del cabello en la base del cabello, produciendo una adición sucesiva al tallo del cabello, que emerge y continúa alargándose de la superficie de la piel. Al final de la etapa de crecimiento, las células productoras de cabello se vuelven inactivas y mueren, entrando en la etapa de degeneración.
Posteriormente, el folículo entra en la
etapa de reposo, que puede durar varias semanas o meses. Eventualmente, las
células madre en la papila producen un nuevo folículo y la etapa de crecimiento
comienza nuevamente. Aproximadamente en este momento, el viejo tallo del
cabello se cae para ser reemplazado por el nuevo tallo en crecimiento. El ciclo
es intrínseco y cortar el cabello no parece acelerar el crecimiento del
cabello.
25.2 Color del
cabello
Los cromatóforos en el folículo aportan
gránulos de pigmento al tallo del cabello para darle más color. A medida que
los humanos (y muchos otros mamíferos) envejecen, su cabello se vuelve gris,
sin importar su color juvenil original. Esto ocurre porque las células madre
especiales responsables del color del cabello dentro del folículo piloso
comienzan a morir. Al principio de la vida, estas células madre se diferencian
en melanocitos especializados que fabrican pigmentos de cabello y piel. Pero a
medida que un individuo envejece, estas células mueren, eliminando una fuente
de pigmentos y sin agregar color al tallo del cabello, dejando solo su color
gris o plateado intrínseco.
25.3 Control
cinético del cabello
El músculo retractor pili, una delgada banda de músculo liso anclado en la
dermis, se une al folículo y hace que el cabello se erija en respuesta al frío,
el miedo o la ira
25.4 Pelo y
cabello
Una cubierta gruesa de cabello, o pelaje,
generalmente se compone de pelos de protección y cabello. Los pelos de
protección, pelos más grandes y gruesos, son los más evidentes en la superficie
externa del pelaje. La parte inferior está estacionada debajo de los pelos
protectores y suele ser mucho más fina y más corta. Tanto los pelos protectores
como los bajos funcionan como aislantes. En la mayoría de los mamíferos
marinos, la parte inferior de la piel se reduce o se pierde por completo, y
solo unos pocos pelos protectores son evidentes. El cabello tiene grano; es
decir, se presenta en una dirección particular. Los golpes contra el grano son
resistidos (Figura 24.2). Se
produce una excepción en los lunares, que carecen de espacio de respuesta y
deben retroceder en sus túneles. El pelo de los lunares se puede peinar hacia
adelante o hacia atrás sin mucha diferencia en el grano.
25.5 Pelos
especializados
Algunos pelos son especializados. Los
nervios sensibles están asociados con las raíces de las vibrisas, o
"bigotes", alrededor de los hocicos de muchos mamíferos. Tal función
sensorial puede haber sido la primera función del cabello en forma de vibrisas,
apareciendo antes de que se desarrollara un pelaje aislante. No es sorprendente
que las vibrisas sean comunes hoy en día en mamíferos nocturnos y en mamíferos
que viven en madrigueras con luz limitada. Las púas de los puercoespines son
pelos rígidos y gruesos especializados para la defensa.
26. Estructuras especiales de la piel de los vertebrados
26.1 Unguis
tipo uña
Las uñas son placas de células epiteliales cornificadas
fuertemente compactadas en la superficie de los dedos de manos y pies; por lo
tanto, son productos del sistema queratinizante de la piel. La matriz de la uña
forma una uña nueva en la base de la uña empujando la uña existente hacia
adelante para reemplazar la que está desgastada o rota en el borde libre. Las
uñas protegen las puntas de los dígitos de lesiones mecánicas involuntarias.
También ayudan a estabilizar la piel en las puntas de los dedos de manos y
pies, de modo que en el lado opuesto la piel puede establecer un agarre seguro
por fricción en los objetos agarrados.
26.2 Unguis
tipo garra
Solo los primates tienen uñas (Figura 26.1a). En otros vertebrados, el sistema de queratinización en el extremo de cada dígito produce garras o pezuñas (Figura 26.1b, c). Las garras, o talones, son proyecciones queratinizadas curvadas, comprimidas lateralmente desde las puntas de los dígitos. Se ven en algunos anfibios y en la mayoría de las aves, reptiles y mamíferos.

Figura 26.1. Derivados epidérmicos. El unguis (uña,
garra o pezuña) es una placa de epitelio cornificado que crece hacia afuera
desde un lecho de células de matriz en proliferación en su base y desde un
subunguis más blando más cerca de su punta.
26.3 Unguis
tipo cascos o pezuñas
Las pezuñas son placas queratinizadas agrandadas en las puntas de los dígitos ungulados. El casco del caballo consiste en la pared del casco, la suela y la rana (Figura 26.2). La pared del casco tiene forma de U y está abierta en el talón, un derivado del tegumento. Consiste en un estrato externo queratinizado (tectorium), una capa superficial delgada y brillante; el estrato medio, más grueso y también queratinizado y permeado con canales tubulares enrollados; y un estrato interno interno (lamellatum), una capa inflada altamente laminada y regularmente que se interdigita con la dermis (corium) debajo.

Figura 26.2. Pezuña de caballo (a) Antepié del
caballo que muestra los huesos internos y la pared del casco. La sección
transversal ampliada muestra capas desde la pared externa hasta el hueso
interno. (b) Sección longitudinal a través del antepié del caballo.
La pared del casco crece desde su base, la región germinal
(células de la matriz), desde la dermis subyacente, a unos 6 mm por mes, y
tarda entre 9 y 12 meses en renovarse. La parte inferior del casco, la
superficie del suelo, incluye la rana en forma de cuña, un derivado mayormente
queratinizado del tegumento que llena la abertura en el talón de la pared del
casco. La suela llena el espacio de la superficie del suelo entre la pared y la
rana triangular (Figura 26.2b). Consiste en
epidermis y dermis engrosada, el corion de la planta del pie. En lo profundo de
la suela se encuentra el cojín digital graso, o almohadilla, un derivado de la
hipodermis. La pared del casco transfiere la energía del impacto a través de
las laminaciones al dígito. Gran parte de la energía del impacto también es
absorbida por el leve ensanchamiento de la pared del casco en forma de U. Las
fuerzas de la suela, la rana y el cojín digital, y el movimiento hacia afuera
de los cartílagos laterales, crean un componente hidráulico de absorción de
impactos. Breves sobretensiones en los plexos venosos en la pezuña alcanzan
valores altos durante el contacto de 600–800 mm Hg.
26.4 Cuernos y
cornamentas
Los lagartos "con cuernos" tienen procesos que se extienden desde detrás de la cabeza que parecen cuernos pero son escamas epidérmicas puntiagudas y especializadas. Los mamíferos, los dinosaurios y las tortugas extintas son los únicos vertebrados con verdaderos cuernos o astas. La piel, junto con el hueso subyacente, contribuye tanto a los cuernos como a las astas. A medida que estas estructuras toman forma, el hueso subyacente se eleva, llevando consigo el integumento suprayacente.

Figura 26.3. Cuernos y astas. (a) Los cuernos
aparecen como excrecencias del cráneo debajo del tegumento, que forma una vaina
queratinizada. Los cuernos ocurren en bóvidos de ambos sexos y generalmente se
retienen durante todo el año. (b) Las astas también aparecen como excrecencias
del cráneo debajo del tegumento suprayacente, que se conoce como
"terciopelo" debido a su apariencia. Finalmente, este terciopelo
suprayacente se seca y se cae, dejando las astas huesudas. Las astas están
restringidas a los miembros de la familia de los ciervos y, a excepción del
caribú (reno), están presentes solo en los machos. Las astas son arrojadas y
reemplazadas anualmente.
En los cuernos, el tegumento asociado produce una vaina
resistente y cornificada que se ajusta sobre el núcleo óseo que nunca se
ramifica (Figura 26.3a). En las
cornamentas, la piel viva suprayacente (llamada "terciopelo")
aparentemente da forma y proporciona suministro vascular al hueso en
crecimiento. Finalmente, el terciopelo se cae para desenvainar el hueso
desnudo, el material real de las astas terminadas que se ramifica (Figura 26.3b). La caía del
terciopelo no es placentera, pues los animales parecen apremiados por
quitársela una vez la cornamenta ha madurado.
👉 Cornamentas: Las astas verdaderas ocurren solo en miembros de los Cervidae (por ejemplo, ciervos, alces, alces).

Figura 26.4. Dendrograma sencillo de la familia Cervidae.
Por lo general, solo los machos tienen astas, que se ramifican y se desprenden anualmente. Hay excepciones notables. Entre los caribúes, ambos sexos tienen astas estacionales. En los ciervos, la cornamenta generalmente consiste en una viga principal, de la cual se ramifican dientes más cortos o puntos. El número de dientes tiende a aumentar con la edad, aunque no exactamente. En la vejez, las astas pueden incluso deformarse. En el caribú y especialmente en los alces, el haz de asta principal está comprimido y palmeado, o en forma de pala, con una serie de puntos que se proyectan desde el borde. El ciclo anual de crecimiento y pérdida de astas en el venado cola blanca, por ejemplo, está bajo control hormonal.
Figura 26.5. Crecimiento anual de cuernos de alce.
(a, b) Nuevas cornamentas comienzan a crecer en abril. (c) En mayo, las astas
están casi completamente formadas, aunque todavía están cubiertas por el
tegumento vivo (terciopelo). (d) A fines del verano, el terciopelo ha comenzado
a secarse y desprenderse. (e) Las astas óseas completamente formadas están en
su lugar.
En la primavera, el aumento de la luz del día estimula la
glándula pituitaria en la base del cerebro para liberar hormonas que estimulan
a las astas a brotar de los sitios en los huesos del cráneo. A finales de la
primavera, las astas en crecimiento están cubiertas por terciopelo. En otoño,
las hormonas producidas por los testículos inhiben la hipófisis y el terciopelo
se seca. Al golpear y frotar, pues parece que esa cosa pica mucho, el venado
limpia el terciopelo para exponer el hueso de las astas completamente formado,
pero ahora muerto.
Los machos usan sus astas durante los enfrentamientos con
otros machos para mantener el acceso a las hembras reproductivamente receptivas.
Después de esta breve temporada de apareamiento, otros cambios hormonales
conducen a un debilitamiento de la asta en su base, donde se une al hueso vivo
del cráneo. Las astas se desprenden, y durante un corto período de tiempo
durante el invierno, los ciervos no tienen astas.
👉 Cuernos: Entre los mamíferos, los verdaderos cuernos se encuentran entre los miembros de la familia Bóvidos (por ejemplo, bovinos, antílopes, ovejas, cabras, bisontes, ñus). Comúnmente, los cuernos ocurren tanto en machos como en hembras, se retienen durante todo el año y continúan creciendo durante toda la vida del individuo. La bocina no está ramificada y está formada por un núcleo óseo y una vaina queratinizada (Figura 26.6). Los de los machos están diseñados para resistir las fuerzas encontradas durante el combate de cabezazos. En las especies grandes, las hembras suelen tener cuernos también, aunque no son tan grandes y curvas como en los machos. En especies pequeñas, las hembras son a menudo sin cuernos.
Figura 26.6. Verdaderos cuernos de la oveja de montaña (bóvidos). La cubierta cornificada del cuerno de la oveja de montaña se retira del lado derecho del cráneo para revelar el núcleo óseo.

Figura 26.7. Otros tipos de cuernos. (a)
Antílope americano, el núcleo óseo de los cuernos no está ramificado, pero la
vaina cornificada está ramificada. (b) Los cuernos de jirafa son pequeñas
perillas osificadas cubiertas por el tegumento. (c) Los rinocerontes tienen
varios cuernos que descansan sobre un pomo bajo en el cráneo, pero estos
cuernos no tienen núcleo interno de hueso. Como consecuencia de la epidermis
sola, están compuestos principalmente de fibras queratinizadas compactadas.
A diferencia de los verdaderos cuernos de los bóvidos, los
cuernos del berrendo o antílope americano, la familia Antilocapridae, se bifurcan en los machos adultos. La vieja vaina
cornificada externa, pero no el núcleo óseo, se arroja anualmente a principios
del invierno (Figura 26.7a). La nueva
vaina debajo, ya en su lugar, se vuelve completamente desarrollada y bifurcada
para el verano. Los cuernos de las hembras también tienen cuernos cuya vaina
queratinizada se reemplaza anualmente, pero generalmente son mucho más pequeños
y solo ligeramente bifurcados. Los cuernos de las jirafas son aún más
diferentes. Se desarrollan a partir de procesos cartilaginosos separados que se
osifican, se fusionan en la parte superior del cráneo y permanecen cubiertos de
piel viva y no coronada (Figura 26.7b), pueden
parecer ridículos, pero pegan duro. El cuerno de rinoceronte no incluye un
núcleo óseo, por lo que es exclusivamente un producto del tegumento. Se forma a
partir de fibras queratinosas compactadas (Figura 26.7c).
26.5 Barbas de
filtrado
El integumento dentro de las bocas de las ballenas misticetas
forma placas de barbas que actúan como coladores para extraer el krill del agua
que se traga en la boca distendida. La barba no contiene hueso. Es una serie de
placas queratinizadas que surgen del tegumento. Durante su formación, grupos de
papilas dérmicas se extienden y alargan hacia afuera, llevando la epidermis
suprayacente. La epidermis forma una capa cornificada sobre la superficie de
estas papilas salientes. Colectivamente, estas papilas y su cubierta de
epidermis constituyen las placas de barba con volantes.

Figura 26.8. Las barbas de una ballena filtradora.

Figura 26.9. Barbas “baleen”
de una ballena. El revestimiento de la boca incluye un epitelio con la
capacidad de formar estructuras queratinizadas. Grupos de epitelio en
crecimiento se vuelven queratinizados y con volantes para formar la barba. Las
secciones longitudinales de la capa queratinizada y el volante se amplían
debajo del diagrama de la barba.
26.6 Escamas
Las escalas tienen muchas funciones. Tanto las escamas epidérmicas como las dérmicas son duras, por lo que cuando reciben daño mecánico y abrasión de la superficie, evitan daños a los tejidos blandos debajo. La densidad de las escamas también las convierte en una barrera contra la invasión de patógenos extraños, y retardan la pérdida de agua del cuerpo. En los tiburones y otros peces, las escamas amortiguan la turbulencia de la capa límite para aumentar la eficiencia de la natación. Algunos reptiles regulan la cantidad de calor de la superficie que absorben girando sus cuerpos hacia o lejos del sol. Esto determina si los rayos del sol se desvían de la cara completa de la escala o brillan debajo del borde posterior elevado de la escala para alcanzar la delgada epidermis debajo. Las escamas epidérmicas son el componente principal de la piel de los reptiles. También están presentes en pájaros a lo largo de sus patas, y en algunos mamíferos, como el castor, cubren la cola (Figura 26.10a).

Figura 26.10. Derivados epidérmicos y
dérmicos. (a) Las escamas epidérmicas ocurren en algunas estructuras de
mamíferos, como esta cola de castor (vista dorsal). (b) Aunque las escamas
dérmicas son raras, están presentes y fusionadas en la armadura del Glyptodon,
un mamífero fósil. (c) Caparazón de tortuga. El caparazón envolvente de una
tortuga se deriva de tres fuentes: las costillas y las vértebras del
endoesqueleto (punteado) y el hueso dérmico que surge en el tegumento (blanco).
La superficie de este caparazón óseo está cubierta por escamas epidérmicas
grandes y delgadas (no se muestran).
26.7 Armadura
dérmica
El hueso dérmico forma la armadura de los peces ostracodermos y placodermos. Al ser un producto de la dermis, el hueso dérmico encuentra alianzas con una gran variedad de estructuras. El hueso dérmico soporta las escamas de los peces óseos, pero tiende a perderse en los tetrápodos. Está ausente en la piel de las aves y la mayoría de los mamíferos. Se han observado excepciones anteriormente, a saber, en el mamífero fósil Glyptodon (Figura 26.10b) y en la piel del armadillo vivo.

Figura 26.11. Armadillo tatú
gigante Priodontes maximus.
Sin embargo, los huesos dérmicos seleccionados se instalan en
el cráneo de los peces y la cintura escapular y han persistido en grupos
modernos de vertebrados. La mayoría de los huesos dérmicos del cráneo y la
cintura escapular comenzaron todos filogenéticamente en la piel y luego se
hundieron hacia adentro para convertirse en partes del esqueleto. Este
intercambio de partes disponibles entre sistemas revela nuevamente el carácter
de remodelación de la evolución. El caparazón de las tortugas es una estructura
compuesta. La mitad dorsal del caparazón es el caparazón, formado por fusión
del hueso dérmico con costillas y vértebras expandidas (Figura 26.10c). Ventralmente,
el plastrón representa huesos dérmicos fusionados a lo largo del abdomen. En la
superficie del caparazón y del plastrón, las placas de epidermis queratinizadas
cubren este hueso subyacente.
26.8 Colores
El color de la piel resulta de interacciones complejas entre
las propiedades físicas, químicas y estructurales del integumento. Los cambios
en el suministro de sangre pueden enrojecer la piel, como en el rubor. La
dispersión diferencial de la luz, denominada dispersión de Tyndall, es la base
de mucho color en la naturaleza. Este es el fenómeno que hace que el cielo
despejado parezca azul. En las aves, las cavidades llenas de aire dentro de las
púas de las plumas aprovechan este fenómeno de dispersión para producir las
plumas azules de los martines pescadores, los arrendajos azules, los pájaros
azules y los racimos de añil. Muchos colores negro, marrón, rojo, naranja y
amarillo son el resultado de pigmentos que producen color por reflejo selectivo
de la luz. Los fenómenos de interferencia son responsables de los colores iridiscentes.
Como la luz se refleja de materiales con diferentes índices de refracción, la
interferencia entre diferentes longitudes de onda de luz produce colores
iridiscentes. En muchas aves, los colores iridiscentes son el resultado de la
interferencia de la luz reflejada en las pequeñas púas y las bárbulas de las
plumas, esto explica entre otras, porque el color de los mamíferos es más
limitado, el cabello mamífero carece de las estructuras de la pluma como las
bárbulas y sus ganchos, y por ende, el cabello y el pelo están más limitados en
sus colores.
27. La piel humana, dermis y epidermis
|| Regresar al Índice ||
La piel (tegumento) y sus apéndices son un órgano consiste en tejidos que trabajan juntos para realizar ciertas funciones. Aunque la piel es menos compleja que la mayoría de los otros órganos, sigue siendo una maravilla arquitectónica.

Figura 27.1. Estructura de la piel. Diagrama tridimensional generalizado de la piel, estructuras accesorias y la hipodermis subyacente.

Figura 27.2. Estructura gruesa de la piel y tejidos
subyacentes. Foto de la parte superior del brazo de un cadáver.
También es el más grande de todos los
órganos, representando aproximadamente el 7% del peso corporal total. El grosor
de la piel varía de 1,5 a 4 mm o más en diferentes regiones del cuerpo. Tiene
dos regiones distintas (Figura 27.1 y Figura 27.2): la
región superficial es un tejido epitelial grueso, la epidermis. En lo profundo
de la epidermis se encuentra la dermis, un tejido conectivo fibroso. Justo en
lo profundo de la piel se encuentra una capa de grasa llamada hipodermis,
compuesta de tejido conectivo areolar suelto y tejido adiposo. Aunque la
hipodermis no es parte del sistema tegumentario, comparte algunas de las
funciones de la piel y, por lo tanto, se describe en este capítulo. La piel
realiza una variedad de funciones:
👉 Protección.
La piel amortigua y aísla los órganos más profundos del cuerpo y protege al
cuerpo de golpes, rasguños y cortes. La piel también protege al cuerpo de
químicos y microorganismos invasores. La epidermis es resistente al agua, lo
que evita la pérdida innecesaria de agua en la superficie del cuerpo, y las
células en la capa epidérmica producen pigmento para proteger la piel de los
efectos nocivos de la radiación ultravioleta (UV).
👉 Regulación de la temperatura corporal. Las ricas redes capilares y las glándulas sudoríparas de la piel
regulan la pérdida de calor del cuerpo, ayudando a controlar la temperatura
corporal.
👉 Excreción.
La piel actúa como un sistema excretor en miniatura cuando la urea, las sales y
el agua se pierden a través del sudor.
👉 Producción de vitamina D. Las células epidérmicas usan radiación UV para sintetizar
vitamina D, una molécula necesaria para absorber el calcio del tracto
digestivo.
👉 Recepción sensorial. La piel contiene órganos sensoriales llamados receptores
sensoriales que están asociados con las terminaciones nerviosas. Al sentir el
tacto, la presión, la temperatura y el dolor, estos receptores nos mantienen al
tanto de las condiciones en la superficie del cuerpo
Explorarás las funciones de la piel con mayor detalle a medida que explore su anatomía.

Figura 27.3. Células epidérmicas y capas de la
epidermis. (a) Fotomicrografía de las cuatro capas epidérmicas principales
en piel delgada (500x). (b) Diagrama que muestra estas cuatro capas y la
distribución de diferentes tipos de células. El estrato lúcido, presente en la
piel gruesa, no se ilustra aquí.
27.1 La epidermis
La epidermis "en la piel" es un
epitelio escamoso estratificado queratinizado que contiene cuatro tipos
distintos de células: queratinocitos, melanocitos, células epiteliales táctiles
y células dendríticas (Figura 27.3).
Los queratinocitos, la célula epidérmica más abundante, producen queratina, una
proteína fibrosa resistente que le da a la epidermis sus propiedades
protectoras. Los queratinocitos proporcionan protección física y mecánica.
También producen antibióticos y enzimas que desintoxican los productos químicos
nocivos a los que está expuesta nuestra piel.
Los queratinocitos están estrechamente
conectados entre sí por una gran cantidad de desmosomas. Surgen en la parte más
profunda de la epidermis a partir de células que sufren mitosis casi continua.
A medida que estas células son empujadas hacia arriba por la producción de
nuevas células debajo de ellas, producen la queratina que eventualmente llena
su citoplasma. Cuando se acercan a la superficie de la piel, están muertos, los
sacos planos completamente llenos de queratina. Millones de estas células
muertas se eliminan todos los días, dándonos una epidermis completamente nueva
cada 35 a 45 días, el tiempo promedio desde el nacimiento de un queratinocito
hasta su desgaste final. Donde la piel experimenta fricción, tanto la
producción celular como la formación de queratina se aceleran.
Los otros tipos de células epidérmicas se
distribuyen escasamente entre los queratinocitos. Discutiremos estas células y
sus funciones mientras examinamos las capas de la epidermis. La variación en el
grosor de la epidermis determina si la piel es gruesa o delgada. En la piel
gruesa, que cubre las palmas y las plantas, la epidermis consta de cinco capas
o estratos. En la piel delgada, que cubre el resto del cuerpo, solo están
presentes cuatro estratos.
27.2 Estrato
basal
El estrato basal, la capa epidérmica más
profunda, está firmemente adherido a la dermis subyacente a lo largo de un borde
ondulado. También llamado estrato germinativo, consiste en una sola fila de
células, en su mayoría células madre que representan los queratinocitos más
jóvenes (Figura 27.3).
Estas células se dividen rápidamente y son visibles muchos núcleos mitóticos.
Las células epiteliales táctiles, o células de Merkel, se distribuyen
escasamente entre los queratinocitos. Cada célula epitelial táctil en forma de
hemisferio está íntimamente asociada con una terminación nerviosa sensorial
similar a un disco y funciona como un receptor para el tacto.
Alrededor del 10% al 25% de las células en
el estrato basal son melanocitos en forma de araña, que hacen que la piel se
oscurezca. La melanina se produce en gránulos con paredes de membrana y luego
se transfiere a través de los procesos celulares (las "patas de
araña") a los queratinocitos cercanos. En consecuencia, los queratinocitos
basales contienen más melanina que los melanocitos mismos. Esta melanina se
agrupa en el lado superficial de los queratinocitos entre la radiación entrante
y los núcleos celulares, protegiendo así los núcleos celulares de los rayos UV,
que pueden dañar el ADN y causar cáncer. En personas de piel clara, la melanina
es digerida por los lisosomas en las células a poca distancia por encima de la
capa basal. En las personas de piel oscura, no se produce dicha digestión, por
lo que la melanina ocupa los queratinocitos en toda la epidermis. Aunque las
personas de piel oscura tienen una melanina más oscura, más gránulos y más
pigmento en cada melanocito, no tienen más melanocitos en la piel. En todas las
personas, excepto en las más oscuras, los melanocitos responden a la radiación
ultravioleta (UVR) al aumentar la producción de melanina y aumentar su
transferencia a los queratinocitos, la respuesta protectora que conocemos como
bronceado.
El papel de los melanocitos en la
pigmentación de la piel ha sido reconocido desde hace mucho tiempo. Además, los
melanocitos también secretan una variedad de moléculas de señalización en
respuesta a la radiación ultravioleta que actúan para modular la respuesta
inmune en la piel. Estas moléculas de señalización influyen en la respuesta
inflamatoria y pueden tener otras funciones reguladoras.
27.3 Estrato
espinoso
El estrato espinoso tiene varias capas de
células gruesas (Figura 27.3). La
mitosis ocurre aquí, pero con menos frecuencia que en la capa basal. Esta capa
recibe su nombre de las muchas extensiones espinelas de sus queratinocitos,
como se ve en los portaobjetos histológicos típicos. Sin embargo, estas espinas
no existen en las células vivas: son artefactos creados durante la preparación
del tejido, cuando las células se encogen mientras se mantienen apretadas en
sus muchos desmosomas. Las células del estrato espinoso contienen gruesos haces
de flamentos intermedios, que consisten en una proteína pre-queratina
resistente a la tensión. Dispersos entre los queratinocitos del estrato
espinoso se encuentran las células dendríticas. Estas células en forma de
estrella son parte del sistema inmune. Las células dendríticas vigilan la
superficie externa del cuerpo, utilizando endocitosis mediada por receptor para
absorber proteínas extrañas (antígenos) que han invadido la epidermis. Luego
abandonan la piel y viajan a un ganglio linfático cercano e inician una
respuesta inmune a todas las células extrañas que transportan el antígeno.
27.4 Estrato
granuloso
El estrato granuloso delgado consta de una a cinco capas de queratinocitos planos. Junto con abundantes filamentos intermedios pre-queratina, estas células también contienen gránulos de queratohialina y gránulos lamelares, de ahí su nombre: capa granular. Los gránulos de queratohialina ayudan a formar queratina en las capas más superficiales, como se describe en breve. Los gránulos lamelares contienen un glicolípidos a prueba de agua que se secreta en el espacio extracelular. El glicolípido, junto con las uniones estrechas entre las células adyacentes, juega un papel importante en la disminución de la pérdida de agua a través de la epidermis. Además, las membranas plasmáticas de las células se engrosan para que se vuelvan más resistentes a la destrucción. Es como si los queratinocitos se "endurecieran" para hacer de la capa externa la región más fuerte de la piel. Como todos los epitelios, la epidermis se basa en capilares en el tejido conectivo subyacente (la dermis) para obtener sus nutrientes. Por encima del estrato granuloso, las células epidérmicas están demasiado lejos de los capilares dérmicos para recibir alimento, por lo que mueren, una ocurrencia completamente normal.

Figura 27.4.
Fotomicrografía de la epidermis y dermis de piel gruesa (185x).
27.5 Estrato
lúcido
El estrato lúcido ocurre en la piel gruesa
(Figura 27.4)
pero no en la piel delgada. Apareciendo a través del microscopio óptico como una
delgada banda translúcida, el estrato lúcido consta de unas pocas filas de
queratinocitos planos y muertos. La microscopía electrónica revela que sus
células son idénticas a las del fondo de la siguiente capa, el estrato córneo.
27.6 Estrato
córneo
La parte más externa de la epidermis, el
estrato córneo, tiene muchas células gruesas. Es mucho más grueso en piel
gruesa que en piel delgada (Figura 27.4).
Sus queratinocitos muertos son sacos planos completamente llenos de queratina
porque sus núcleos y orgánulos se desintegraron con la muerte celular. La
queratina consiste en los fragmentos intermedios pre-queratina incrustados en
un "pegamento" de los gránulos de queratohialina. Tanto la queratina
como las membranas plasmáticas engrosadas de las células en el estrato córneo
protegen la piel contra la abrasión y la penetración. Además, el glicolípido
entre sus células mantiene esta capa impermeable. ¡Es sorprendente que una capa
de células muertas pueda realizar tantas funciones!
Las células del estrato córneo se
desprenden regularmente. Estas células son la caspa que se desprende del cuero
cabelludo y los copos que salen de la piel seca. La persona promedio arroja 18
kg (40 libras) de estos copos de piel en la vida. Estas celdas desprendidas son
reemplazadas por celdas de las capas más profundas. La próxima vez que escuche
el dicho "La belleza es sólo superficial", ¡tenga en cuenta que casi
todo lo que ve cuando mira a alguien está muerto!
27.7 Dermis
La dermis, la segunda región principal de
la piel, es un tejido conectivo fuerte y flexible. Las células de la dermis son
típicas de cualquier tejido conectivo propiamente dicho: fibroblastos,
macrófagos, mastocitos y glóbulos blancos dispersos. Los tipos de fibra
(colágeno, elástico y reticular) también son típicos. La dermis une todo el
cuerpo como una media al pie. Es su "piel" y corresponde a las pieles
de animales utilizadas para hacer productos de cuero. La dermis tiene dos
regiones: la dermis papilar y la dermis reticular (Figura 27.4).
👉 Dermis papilar: La dermis papilar, el 20% superficial de la dermis, es un tejido
conectivo areolar que contiene colágeno muy delgado y fibras elásticas. Incluye
las papilas dérmicas, proyecciones similares a dedos que se extienden hacia la
epidermis suprayacente. Estas proyecciones de las papilas dérmicas hacia la
epidermis aumentan el área de superficie para el intercambio de gases,
nutrientes y productos de desecho entre estas capas. Recuerde que la epidermis
es avascular y depende de la difusión de estos materiales desde la dermis
subyacente. La interdigitación de estas capas también fortalece la unión
dermoepidérmica y, por lo tanto, reduce la formación de ampollas.
En las palmas de las manos y las plantas
de los pies, las papilas dérmicas se encuentran sobre montículos más grandes
llamados crestas dérmicas. Éstos elevan la epidermis suprayacente a crestas
epidérmicas o crestas de fricción, que crean huellas dactilares, huellas de
palmas y huellas (Figura 27.5a).
Las crestas epidérmicas aumentan la fricción y mejoran la capacidad de agarre
de las manos y los pies. Los patrones de estas crestas están genéticamente
determinados y son únicos para cada persona. Debido a que los poros de sudor se
abren a lo largo de las crestas de las crestas de fricción, dejan huellas
dactilares distintas en casi todo lo que tocan. Por lo tanto, las huellas
digitales son "películas de sudor".
👉 Dermis reticular: La dermis reticular más profunda, que representa aproximadamente
el 80% del grosor de la dermis, es tejido conectivo irregular denso. Su matriz
extracelular contiene gruesos haces de colágeno entrelazado y fibras elásticas
que se ejecutan en muchos planos diferentes. Sin embargo, la mayoría corre
paralela a la superficie de la piel. La capa reticular se llama así por sus
redes de fibras de colágeno (retículo = red); el nombre no implica ninguna
abundancia especial de fibras reticulares. Las separaciones o regiones menos
densas entre los haces de colágeno forman las líneas de escisión o líneas de
tensión de la piel (Figura 27.5b).
Estas líneas invisibles ocurren en todo el cuerpo: corren longitudinalmente en
la piel de las extremidades y la cabeza y en patrones circulares alrededor del
cuello y el tronco. El conocimiento de las líneas de escisión es importante
para los cirujanos. Las incisiones paralelas a estas líneas tienden a abrirse
menos y sanar más fácilmente que las incisiones realizadas a través de las
líneas de escisión.
Las fibras de colágeno de la dermis le dan a la piel su fuerza y resistencia. Por lo tanto, muchos golpes y rasguños no penetran esta capa resistente. Además, las fibras elásticas en la dermis proporcionan a la piel propiedades de estiramiento y retroceso. El estiramiento extremo de la piel, como ocurre en la obesidad y el embarazo, puede rasgar el colágeno en la dermis. Tal desgarro dérmico da como resultado cicatrices blancas plateadas llamadas "estrías". La dermis también es el sitio receptivo para los pigmentos utilizados en los tatuajes.

Figura 27.5. Modificaciones dérmicas. (a)
Crestas de fricción. (b) Las líneas de escisión representan las separaciones
entre haces de fibras de colágeno en la capa reticular de la dermis. (c) Se
forman líneas de flexión donde la dermis está estrechamente unida a la fascia
subyacente.
De la parte profunda de la dermis surgen
las marcas de la superficie de la piel llamadas líneas de flexión.
Observe, por ejemplo, los pliegues profundos de la piel en la palma de su mano
(Figura 27.5c).
Estos resultan del plegamiento continuo de la piel, a menudo sobre las
articulaciones, donde la dermis se adhiere firmemente a las estructuras
subyacentes. Las líneas de flexión también son visibles en las muñecas, las
plantas, los dedos y los dedos de los pies. La dermis se suministra abundantemente
con fibras nerviosas y vasos sanguíneos. Los vasos sanguíneos dérmicos
consisten en dos plexos vasculares (un plexo es una red de vasos convergentes y
divergentes) (Figura 27.1). El
plexo dérmico profundo se encuentra entre la hipodermis y la dermis. Nutre la
hipodermis y las estructuras ubicadas dentro de las porciones más profundas de
la dermis. El plexo subpapilar más superficial, ubicado justo debajo de las
papilas dérmicas, suministra las estructuras dérmicas más superficiales, las
papilas dérmicas y la epidermis.
Los vasos sanguíneos dérmicos hacen más
que solo nutrir la dermis y la epidermis suprayacente; También desempeñan un
papel crítico en la regulación de la temperatura. Estos vasos son tan extensos
que pueden contener el 5% de toda la sangre del cuerpo. Cuando los órganos
internos necesitan más sangre o más calor, los nervios estimulan la
constricción de los vasos dérmicos, desviando más sangre hacia la circulación
general y haciéndola disponible para los órganos internos. Por el contrario, en
los días calurosos, los vasos dérmicos se llenan de sangre tibia, enfriando el
cuerpo irradiando calor lejos de él. Incrustado
dentro de la dermis hay glándulas y folículos pilosos. Estos apéndices de la
piel se derivan de la capa epidérmica y se extienden hacia la dermis profunda y
la hipodermis.
27.8 La
hipotermis
Muy cerca de la piel está la hipodermis
grasa, de las palabras griegas que significan "debajo de la piel" (Figura 27.1 y Figura 27.2).
Esta capa también se llama fascia superfcial y capa subcutánea. La hipodermis
consiste en tejido conjuntivo adiposo y areolar, pero predomina el tejido
adiposo. Además de almacenar grasa, la hipodermis ancla la piel a las
estructuras subyacentes (principalmente a los músculos), pero lo
suficientemente flojo como para que la piel pueda deslizarse con relativa
libertad sobre esas estructuras. Tal deslizamiento asegura que muchos golpes
simplemente sean irrelevantes a nuestros cuerpos. La hipodermis también es un
aislante: como la grasa es un mal conductor del calor, ayuda a prevenir la
pérdida de calor del cuerpo. La hipodermis se engrosa notablemente con el
aumento de peso, pero este engrosamiento ocurre en diferentes áreas del cuerpo
en los dos sexos. En las mujeres, la grasa subcutánea se acumula primero en los
muslos y los senos, mientras que en los hombres se acumula primero en el
abdomen generando la famosa "barriga cervecera".
28. El color de la piel humana
|| Regresar al Índice ||
Tres pigmentos contribuyen al color de la
piel: melanina, caroteno y hemoglobina.
28.1 Melanina
La melanina, la más importante, está hecha
de un aminoácido llamado tirosina. Presente en varias variedades, la melanina
varía de amarillo a rojizo a marrón a negro. Como se señaló anteriormente, la
melanina pasa de los melanocitos a los queratinocitos en el estrato basal de la
epidermis. Las variaciones en el color de la piel en humanos resultan de
diferencias tanto en la cantidad como en el tipo de melanina producida, sin
embargo, el tono resultante puede variar de manera paralela, convergente o
atavística, de allí que sea un muy malo carácter de clasificación taxonómica.
28.2 Caroteno
El caroteno es un pigmento amarillo
anaranjado que el cuerpo obtiene de fuentes vegetales como zanahorias y
tomates. Tiende a acumularse en el estrato córneo de la epidermis y en la grasa
de la hipodermis. El tono rosado de la piel blanca refleja el color carmesí de
la hemoglobina oxigenada en los capilares de la dermis. Debido a que la piel blanca
contiene poca melanina, la epidermis es casi transparente y permite que se vea
el color de la sangre. Las marcas negras y azules representan sangre
descolorida que es visible a través de la piel. Tales contusiones, generalmente
causadas por golpes, revelan sitios donde la sangre ha escapado de la
circulación y se ha coagulado debajo de la piel. El término general para una
masa coagulada de sangre escapada en cualquier parte del cuerpo es hematoma.
Las razones de las variaciones en los
colores de la piel de diferentes poblaciones humanas no se entienden
completamente. Las teorías que explican esta variación reflejan el hecho de que
los rayos UV del Sol son peligrosos y esenciales al mismo tiempo. Una hipótesis
propone que la coloración oscura de la piel evolucionó para eliminar el peligro
de cáncer de piel causado por la radiación UV. Una hipótesis alternativa
argumenta que los efectos de la luz solar sobre los niveles de ácido fólico en
la sangre fueron la presión selectiva para la evolución de la piel oscura en áreas
tropicales. En personas de piel clara, los altos niveles de luz solar
disminuyen los niveles de ácido fólico en la sangre. La disminución de los
niveles de ácido fólico en mujeres embarazadas aumenta el riesgo de defectos
del tubo neural en el feto en desarrollo. La evolución de la piel oscura en los
trópicos protege los niveles de ácido fólico en la sangre y promueve una
descendencia saludable.
28.3 Vitamina
D
El otro factor que influye en el color de
la piel es la producción de vitamina D. Los rayos UV estimulan la epidermis
profunda para producir vitamina D, una hormona vital necesaria para la
absorción de calcio de la dieta y esencial para la salud de los huesos. El
calcio, y por lo tanto la vitamina D adecuada, también es fundamental para el
feto en desarrollo. La producción de vitamina D no plantea ningún problema en
los trópicos soleados, pero los caucásicos en el norte de Europa reciben poca
luz solar durante el largo y oscuro invierno. Por lo tanto, su epidermis
incolora asegura que penetrarán suficientes rayos UV para la producción de
vitamina D. La exposición natural a los rayos UV en estas regiones templadas no
es suficiente para disminuir los niveles de ácido fólico. La mayoría de los
tipos de color de piel humana evolucionaron en latitudes intermedias (China,
Medio Oriente, etc.) y se caracterizan por tonos de piel moderadamente marrones
que no son "blancos" ni "negros". Dicha piel es lo
suficientemente oscura como para proporcionar cierta protección contra la luz
solar efectos negativos en el verano, especialmente cuando se broncea, pero lo
suficientemente ligero como para permitir la producción de vitamina D en los
inviernos moderados de latitudes intermedias. La compensación entre la
protección contra los efectos nocivos de la radiación UV y la producción de
vitamina D puede haber sido la presión selectiva detrás de la evolución de la
coloración de la piel a medida que los primeros humanos se alejaban de los
trópicos.
28.4 Evolución
Se supone que la pérdida de vello corporal en las especies de Hominini está relacionada con la aparición del bipedalismo hace unos 5 a 7 millones de años. El vello corporal homínido bípedo puede haber desaparecido gradualmente para permitir una mejor disipación del calor a través de la sudoración, el color de la piel de los primeros Hominini posiblemente era blanco como en los chimpancés, pues el pelo cumple las funciones protectoras (Jablonski & Chaplin, 2000). La aparición de la pigmentación de la piel data de hace aproximadamente 1,2 millones de años (Jablonski, 2004), en condiciones de una megamedaza que llevó a los primeros humanos a paisajes áridos y abiertos. Tales condiciones probablemente causaron un exceso de radiación UV-B. Esto favoreció la aparición de pigmentación de la piel oscura para proteger contra el agotamiento de ácido fólico.

Figura 28.1. El color de la
piel se encuentra adaptado a la función, sin importar la filogenia, es decir,
no se cumple la ley de Dollo, un estado de carácter de color de la piel oscura
puede recuperarse si una población de piel blanca permanece unas 100
generaciones en un ambiente con presión de selección a piel oscura.
Con la evolución de la piel sin pelo, las
glándulas sudoríparas abundantes y la piel rica en melanina, los primeros
humanos podían caminar, correr y buscar comida durante largos períodos de
tiempo bajo el Sol caliente sin daño cerebral debido al sobrecalentamiento, lo
que les da una ventaja evolutiva sobre otras especies (Jablonski, 2012).
Hace 1,2 millones de años, alrededor de la época del Homo ergaster, los humanos arcaicos (incluidos los ancestros del Homo sapiens) tenían exactamente la
misma proteína receptora que los africanos subsaharianos modernos (Eswaran, Harpending, &
McCullough, 2004; Rogers, Iltis, & Wooding, 2004). O sea, todos los humanos eran negritos.
Este fue el genotipo heredado por los
humanos anatómicamente modernos, pero retenido solo por parte de las
poblaciones existentes, formando así un aspecto de la variación genética
humana. Hace aproximadamente 100,000–70,000 años, algunos humanos
anatómicamente modernos (Homo sapiens)
comenzaron a emigrar desde los trópicos hacia el norte, donde fueron expuestos
a la luz solar menos intensa. Esto posiblemente se debió en parte a la
necesidad de un mayor uso de ropa para protegerse del clima más frío. En estas
condiciones, hubo menos fotodestrucción del folato y, por lo tanto, se redujo
la presión evolutiva contra la supervivencia de las variantes genéticas de piel
más clara. Además, la piel más clara es capaz de generar más vitamina D
(colecalciferol) que la piel más oscura, por lo que habría representado un
beneficio para la salud en la luz solar reducida si hubiera fuentes limitadas
de vitamina D. Por lo tanto, la hipótesis principal para la evolución del color
de la piel humana propone que:
👉 Desde hace aproximadamente 1,2
millones de años hasta hace menos de 100,000 años, los humanos arcaicos,
incluido el Homo sapiens arcaico,
eran de piel oscura.
👉 A medida que las poblaciones de Homo sapiens comenzaron a migrar, la
restricción evolutiva de mantener la piel oscura disminuyó proporcionalmente a
la distancia al norte que migró una población, lo que resultó en una gama de
tonos de piel dentro de las poblaciones del norte.
👉 En algún momento, algunas
poblaciones del norte experimentaron una selección positiva para la piel más
clara debido a la mayor producción de vitamina D a partir de la luz solar y los
genes para la piel más oscura desaparecieron de estas poblaciones.
👉 Las migraciones posteriores a diferentes
entornos UV y la mezcla entre poblaciones han dado como resultado la variada
gama de pigmentaciones de la piel que vemos hoy.
👉 Las mutaciones genéticas que
conducen a una piel clara, aunque parcialmente diferentes entre los asiáticos
orientales y los europeos occidentales (Norton et al., 2006), sugieren que los
dos grupos experimentaron una presión selectiva similar después del
asentamiento en las latitudes del norte (Juzeniene, Setlow, Porojnicu,
Steindal, & Moan, 2009).
La teoría está parcialmente respaldada por
un estudio sobre el gen SLC24A5 que encontró que el alelo asociado con la piel
clara en Europa "determinó [...] que
habían pasado 18000 años desde que el alelo de la piel clara fue fijado en los
europeos", pero puede haberse originado tan recientemente como
12,000–6,000 años atrás "dada la imprecisión del método" (Gibbons,
2007), que está en línea con la evidencia más temprana de la agricultura.
La investigación realizada por Nina
Jablonski sugiere que un tiempo estimado de aproximadamente 10,000 a 20,000
años es suficiente para que las poblaciones humanas logren una pigmentación
óptima de la piel en un área geográfica particular, pero que el desarrollo de
la coloración ideal de la piel puede ocurrir más rápido si la presión evolutiva
es más fuerte, incluso en tan poco como 100 generaciones (Jablonski, 2011). El período de tiempo también se ve afectado por prácticas
culturales como la ingesta de alimentos, la ropa, los revestimientos corporales
y el uso de refugios que pueden alterar las formas en que el medio ambiente
afecta a las poblaciones (Jablonski, 2012).
Uno de los impulsores propuestos más
recientemente de la evolución de la pigmentación de la piel en humanos se basa
en una investigación que muestra una función de barrera superior en la piel de
pigmentación oscura. La mayoría de las funciones protectoras de la piel, incluida
la barrera de permeabilidad y la barrera antimicrobiana, residen en el estrato
córneo (SC) y los investigadores suponen que la SC ha sufrido el mayor cambio
genético desde la pérdida del vello del cuerpo humano. La selección natural
habría favorecido las mutaciones que protegen esta barrera esencial; Una de
estas adaptaciones protectoras es la pigmentación de la epidermis
interfolicular, porque mejora la función de barrera en comparación con la piel
no pigmentada. Sin embargo, en las exuberantes selvas tropicales, donde la
radiación UV-B y el estrés xérico no eran excesivos, la pigmentación ligera no
habría sido tan perjudicial. Esto explica la residencia lado a lado de pueblos
ligeramente pigmentados y oscuros (Elias, Menon, Wetzel, & Williams,
2010).
Los estudios de población y mezclas
sugieren un modelo de tres vías para la evolución del color de la piel humana,
con la piel oscura evolucionando en los primeros homínidos en África y la piel
clara evolucionando en parte por separado al menos dos veces después de que los
humanos modernos se expandieron fuera de África (Beleza et al., 2012; Norton
et al., 2006). En su mayor parte, la evolución de la piel clara ha seguido
diferentes caminos genéticos en las poblaciones de Eurasia occidental y
oriental. Sin embargo, dos genes, KITLG y ASIP, tienen mutaciones asociadas con
la piel más clara que tienen altas frecuencias en las poblaciones euroasiáticas
y tienen fechas de origen estimadas después de que los humanos se extendieron
fuera de África pero antes de la divergencia de los dos linajes (Beleza et al., 2012).
29. Apéndices del tegumento humano, uñas y cabello
|| Regresar al Índice ||
Junto con la piel misma, el sistema
integumentario incluye varios derivados de la epidermis. Estos apéndices de la
piel incluyen uñas, cabello y folículos pilosos, glándulas sebáceas (de aceite)
y glándulas sudoríparas. Aunque derivan de las células epiteliales de la
epidermis, todas se extienden hacia la dermis.
29.1 Las uñas
Una uña es una modificación en escama de
la epidermis que corresponde al casco o la garra de otros mamíferos. Las uñas
son herramientas integradas que nos permiten recoger objetos pequeños y rascar
la piel cuando pica. Las uñas están hechas de células muertas queratinizadas.
La queratina dura que predomina en los pelos y las uñas difiere de la queratina
suave que se encuentra en las células epidérmicas típicas de dos maneras:
👉 es más resistente y duradera, y
👉 las células de la queratina dura no se desprenden.

Figura 29.1. Estructura
de una uña. (a) Vista de la superficie de la parte distal de un dedo. (b)
Sección sagital a través de la punta de un dedo.
Cada uña tiene un borde libre distal, una
placa ungueal (la parte unida visible) y una raíz (la parte proximal incrustada
en la piel). La uña descansa sobre un lecho de epidermis llamada lecho ungueal.
Este lecho contiene solo las capas más profundas de la epidermis, porque la uña
en sí corresponde a las capas queratinizadas superficiales.
Las uñas se ven rosadas debido a la rica
red de capilares en la dermis subyacente. En la raíz y el extremo proximal del
cuerpo de la uña, el lecho se engrosa para formar la matriz de la uña, la parte
de la uña que crece activamente. La matriz es tan gruesa que la dermis rosa no
puede mostrarse a través de ella. En cambio, vemos una media luna blanca, la
lunule, debajo de la región proximal de la uña. Los bordes laterales y
proximales de la uña se superponen con pliegues cutáneos llamados pliegues
ungueales. El pliegue ungueal proximal se proyecta sobre el cuerpo de la uña como
la cutícula o el eponiquio. Una uña encarnada es justo lo que su nombre
implica, una uña cuyo crecimiento la empuja dolorosamente hacia el pliegue
ungueal lateral. Esto sucede cuando la uña crece torcidamente, más comúnmente
debido a la presión de un zapato que no funciona bien.
29.2 El
cabello
Juntos, los pelos y sus folículos forman
unidades estructurales complejas (Figura 5.7). En estas unidades, los pelos son
los largos flamentos, y los folículos pilosos son invaginaciones tubulares de
la epidermis de donde crecen los pelos.
Aunque el cabello sirve para mantener calientes a otros mamíferos, el vello del cuerpo humano es mucho menos exuberante y menos útil para conservar la temperatura corporal. Aun así, el cabello se distribuye por toda la superficie de nuestra piel, excepto en las palmas, las plantas de los pies, los pezones y partes de los genitales externos (la cabeza del pene, por ejemplo). La función principal de este escaso vello corporal es detectar cosas que tocan ligeramente la piel.

Figura 29.2. Estructura
de un cabello y folículo piloso.
El cabello en el cuero cabelludo protege
la cabeza contra la luz solar directa en verano y contra la pérdida de calor en
los días fríos. Las pestañas protegen los ojos y el vello de la nariz arroja
partículas grandes, como insectos y pelusas, del aire inhalado. Un cabello es
una hebra flexible hecha de células muertas cubiertas de queratina dura. Las
partes principales de un cabello son la raíz, la parte incrustada en la piel y
el tallo, la parte que se proyecta sobre la superficie de la piel (Figura 29.2). Si
su eje es plano y en forma de cinta en sección transversal, el cabello es
rizado; si es de sección ovalada, el cabello es ondulado; Si es perfectamente
redondo, el cabello es liso.
Un cabello consta de tres capas
concéntricas de células queratinizadas (Figura 29.2a y b).
Su núcleo central, la médula, consiste en grandes células y espacios aéreos. La
médula está ausente en los pelos finos. La corteza, que rodea la médula, consta
de varias capas de células planas. La cutícula más externa es una sola capa de
células que se superponen entre sí desde abajo como tejas en un techo. Este
patrón de tejas ayuda a mantener separados los pelos vecinos para que el
cabello no se tape. Los acondicionadores para el cabello alisan la superficie
rugosa de la cutícula para que el cabello se vea más brillante. La cutícula es
la parte más queratinizada del cabello, brinda resistencia y mantiene las capas
internas bien compactadas. Debido a que se somete a la mayor abrasión, la parte
más antigua de la cutícula del cabello tiende a desgastarse en la punta del
tallo del cabello. La abrasión hace que los fráiles de queratina en la corteza
y la médula se encrespen, creando "puntas abiertas".
Los melanocitos forman el pigmento capilar
en la base del folículo piloso (Figura 29.2c) y
se transfieren a las células de la raíz del cabello. Las diferentes
proporciones de dos tipos de melanina (de color negro-marrón y amarillo-óxido)
se combinan para producir todos los colores comunes del cabello: negro, marrón,
rojo, rubio, blanco y todos sus intermedios. El encanecimiento o blanqueamiento
del cabello resulta de una disminución en la producción de melanina y del
reemplazo de melanina por burbujas de aire incoloras en el tallo del cabello.
29.3 Folículos
pilosos
Los folículos pilosos se extienden desde
la superficie epidérmica hacia la dermis. El extremo profundo del folículo se
expande, formando un bulbo piloso (Figura 29.2 c).
Un nudo de terminaciones nerviosas sensoriales se envuelve alrededor de cada
bulbo piloso para formar un receptor de folículo piloso o plexo piloso. Doblar
el cabello estimula estas terminaciones nerviosas. Los pelos son, por lo tanto,
exquisitos receptores táctiles. Pruebe esto pasando suavemente la mano sobre
los pelos de su antebrazo. Una parte de la dermis con forma de pezón, la papila
dérmica (papila capilar), sobresale en cada bulbo piloso. Esta papila contiene
un grupo de capilares que entregan sustancias que estimulan el crecimiento del
cabello y suministran nutrientes al cabello en crecimiento. Si la papila
capilar se destruye por un traumatismo, el folículo deja de producir pelo
permanentemente. Las células epiteliales en el bulbo piloso justo encima de la
papila forman la matriz capilar. Estas células matriciales proliferan para
formar el tallo piloso. La pared de un folículo piloso tiene un componente
dérmico y un componente epidérmico. Estos componentes se describen de externo a
interno (Figura 29.2a –
d).
👉 Vaina periférica del tejido conectivo
(vaina fibrosa). Esta vaina de tejido conectivo se deriva
de la dermis. Forma la capa externa de la pared del folículo.
👉 Membrana vidriosa. La membrana vítrea está en la unión de
la vaina fibrosa y la vaina de la raíz epitelial. Es en esencia, la membrana
basal del epitelio folicular.
👉 Vaina de la raíz epitelial. La vaina de la raíz epitelial se deriva
de la epidermis. Tiene dos componentes: la vaina externa de la raíz, una continuación
directa de la epidermis; y la vaina interna de la raíz, que se deriva de las
células de la matriz
29.4 Tipos de
crecimiento de cabello
Los pelos vienen en varios tamaños y
formas, pero por regla general pueden clasificarse como vellus o terminal. El
vello corporal de niños y mujeres es de la variedad fina, corta y vellosa. El
cabello más largo y grueso del cuero cabelludo de todos es cabello terminal.
Los pelos terminales también aparecen en la pubertad en las regiones axilar
(axila) y púbica en ambos sexos y en la cara, el pecho, los brazos y las
piernas de los hombres. Estos pelos terminales crecen bajo la influencia de las
hormonas sexuales masculinas llamadas andrógenos, de los cuales la testosterona
es el tipo más importante.
Los pelos crecen un promedio de 2 mm por
semana, aunque esta tasa varía ampliamente entre las regiones del cuerpo y con
el sexo y la edad. Cada folículo pasa por ciclos de crecimiento. En cada ciclo,
una fase de crecimiento activa es seguida por una fase de reposo, en la cual la
matriz del cabello está inactiva y el folículo se atrofia un poco. Al comienzo
de cada fase activa, el nuevo cabello en crecimiento expulsa el cabello viejo y
lo desprende. La vida útil de los pelos varía: en el cuero cabelludo, los
folículos permanecen activos durante un promedio de 4 años, por lo que los
pelos individuales crecen bastante tiempo antes de ser arrojados. En las cejas,
por el contrario, los folículos están activos durante unos pocos meses, por lo
que las cejas nunca crecen mucho. Afortunadamente, los ciclos de los folículos
pilosos adyacentes en el cuero cabelludo no están en sincronía; por lo tanto,
los humanos arrojan solo un pequeño porcentaje de los pelos de la cabeza en
cualquier momento.
29.5 Tiñendo
el cabello
Dadas las condiciones ideales, el cabello crece más rápido desde la adolescencia hasta los 40 años. Cuando los pelos ya no se reemplazan tan rápido como se desprenden, el cabello se adelgaza. Entre los 60 y los 65 años, ambos sexos generalmente experimentan cierto grado de calvicie. Los pelos terminales gruesos se reemplazan por vellos, y el cabello se vuelve cada vez más tenue. La verdadera calvicie es diferente. El tipo más común, la calvicie de patrón masculino, es una afección genéticamente determinada y con influencia de género. Se cree que es causado por un gen que no se expresa hasta la edad adulta, momento en el cual cambia la respuesta de los folículos capilares a los andrógenos. Los pelos responden a los andrógenos acortando cada vez más sus ciclos de crecimiento. Los ciclos se vuelven tan cortos que muchos pelos nunca emergen de sus folículos antes de ser arrojados, y los que sí emergen son vellos finos que parecen pelusa de melocotón en el área "calva". Los medicamentos utilizados para tratar la calvicie de patrón masculino inhiben la producción de andrógenos o aumentan el flujo sanguíneo a la piel y los folículos pilosos. Estos tratamientos son solo parcialmente exitosos.
30. Glándulas de la piel humana
30.1 Glándulas
cebáceas
Las glándulas sebáceas son las glándulas sebáceas de la piel. Ocurren en todo el cuerpo, excepto en las palmas y las plantas. Son glándulas alveolares simples con varios alvéolos que se abren en un solo conducto, pero los alvéolos en realidad están repletos de células, por lo que no hay luz (cavidad central). Su producto aceitoso, llamado sebo (sebo; "grasa animal"), se secreta de la manera más inusual: las células centrales en los alvéolos acumulan lípidos aceitosos hasta que se hinchan y se rompen. Este proceso se llama secreción holocrina, porque las células enteras se rompen para formar el producto. La mayoría de las glándulas sebáceas están asociadas con los folículos pilosos, vaciando su sebo en el tercio superior del folículo. A partir de ahí, el sebo fluye superficialmente para cubrir la piel. Además de hacer que nuestra piel y cabello sean grasosos pero brillantes y manejables, el sebo acumula suciedad, suaviza y lubrica el cabello y la piel, evita que el cabello se vuelva quebradizo y evita que la epidermis se agriete. También ayuda a frenar la pérdida de agua en la piel y a matar las bacterias. La secreción de sebo es estimulada por las hormonas, especialmente los andrógenos. Las glándulas sebáceas están relativamente inactivas durante la infancia, pero se activan en ambos sexos durante la pubertad, cuando la producción de andrógenos comienza a aumentar.

Figura 30.1. Glándulas
de la piel.
30.2 Glándulas
sudoríparas
La sudoración evita el sobrecalentamiento
del cuerpo, porque el sudor enfría la piel a medida que se evapora. Solo los
mamíferos tienen glándulas sudoríparas. Los humanos tienen más de 2.5 millones
de glándulas sudoríparas distribuidas por toda la superficie de la piel,
excepto en los pezones y partes de los genitales externos. Los humanos
normalmente producen alrededor de 500 ml de sudor por día, pero esta cantidad
puede aumentar a 12 litros (más de 3 galones) en días calurosos durante el
ejercicio vigoroso. El cabello interfiere con la evaporación del sudor y la
capacidad de enfriar el cuerpo, por lo que la necesidad de una mayor regulación
de la temperatura a través del sudor provocó una reducción de la vellosidad en
los humanos. Hay dos tipos de glándulas sudoríparas, que aumentan su secreción
en respuesta al estrés y al calor: glándulas ecrinas y apocrinas.
👉 Ecrinas: Las glándulas ecrinas son, con mucho, el tipo más numeroso (Figura 30.1).
Son más abundantes en las palmas, las plantas y la frente. Cada uno es una
versión en espiral de una glándula tubular simple. La base secretora en espiral
se encuentra en la dermis profunda y la hipodermis, y el conducto se extiende
superficialmente para abrirse en la superficie de la piel a través de un poro
en forma de embudo. (Aunque la mayoría de los poros en la superficie de la piel
son poros del sudor, los "poros" que se ven en la cara son aberturas
de los folículos pilosos). El sudor es un producto secretor inusual en el
sentido de que es principalmente una filtración de sangre que pasa a través de
las células secretoras de la piel, glándulas sudoríparas y se libera por
exocitosis. El sudor es 99% de agua, con algunas sales (principalmente cloruro
de sodio) y trazas de desechos metabólicos (urea, amoníaco, ácido úrico). Es
ácido, por lo que retarda el crecimiento de bacterias en la piel.
👉 Apocrinas: Las glándulas apocrinas se confinan principalmente a las áreas
axilar, anal y genital. Son más grandes que las glándulas ecrinas y sus
conductos se abren en los folículos pilosos. Las glándulas apocrinas producen
un tipo especial de sudor que consiste en sustancias grasas y proteínas. Por
esta razón, el sudor apocrino es viscoso y a veces tiene un color lechoso o
amarillo. Este producto es inodoro cuando se secreta por primera vez, pero a medida
que sus moléculas orgánicas se descomponen por bacterias en la piel, adquiere
un olor a almizcle. Esta es la fuente del olor corporal. Las glándulas
apocrinas comienzan a funcionar en la pubertad bajo la influencia de los
andrógenos. Su actividad sexual aumenta con los juegos sexuales previos, y
aumentan y retroceden con las fases del ciclo menstrual de una mujer. Las
secreciones de las glándulas apocrinas se identificaron como feromonas humanas
verdaderas (señales químicas que transmiten información a un miembro de la
misma especie) a fines de la década de 1990, cuando se demostró que son
responsables de la sincronía del ciclo menstrual que ocurre en las mujeres que
vivir juntas.
Las glándulas apocrinas están involucradas
con la señalización sexual y parecen funcionar en el atractivo y la selección
de pareja. Los genes que codifican para el sistema inmune, el complejo mayor de
histocompatibilidad (MHC), también influyen en las secreciones de las glándulas
apocrinas. Cada persona tiene un conjunto único de estos genes. En los
experimentos, los olores del olor corporal que las mujeres seleccionaron como
"sexy" o "atractivo" provenían de hombres que tenían genes
del sistema inmunitario más diferentes que los suyos. Las parejas con genes del
sistema inmunitario complementario proporcionan a sus descendientes una mayor
protección contra la enfermedad y una menor probabilidad de trastornos
recesivos.
La piel forma varios tipos de glándulas
sudoríparas modificadas. Las glándulas ceruminosas son glándulas apocrinas
modificadas en el revestimiento del canal auditivo externo. Su producto es un
componente de cerumen. Las glándulas mamarias son glándulas sudoríparas
especializadas altamente modificadas para secretar leche.
30.3 Las
glándulas mamarias
Las glándulas mamarias (senos) son
glándulas sudoríparas modificadas que están presentes en ambos sexos pero
funcionan solo en las mujeres lactantes (Figura 30.2)
cuando producen leche para alimentar a un bebé en respuesta a la estimulación
hormonal. Embrionariamente, las glándulas mamarias se forman como parte de la
piel, surgiendo a lo largo de líneas bilaterales que se extienden entre la
axila y la ingle en el tronco lateral del embrión. Estas líneas de leche
embrionaria persisten solo en el tórax medio, donde se forman los senos
definitivos. Aproximadamente 1 de cada 500 personas desarrolla pezones o senos
adicionales (accesorios), que pueden ocurrir en cualquier lugar a lo largo de
la línea de la leche. La base del seno en forma de cono en las mujeres se
extiende desde la segunda costilla hacia arriba hasta la sexta costilla hacia
abajo. Su borde medial es el esternón, y su límite lateral es la línea
midaxilar, una línea que se extiende en línea recta hacia abajo desde la mitad
de la axila. Los músculos profundos del seno son el pectoral mayor y menor y
partes del serrato anterior y oblicuo externo. Las arterias principales del
seno son la arteria torácica lateral y las ramas cutáneas de los intercostales
torácicos internos y posteriores. Los vasos linfáticos que drenan el seno
drenan hacia los ganglios linfáticos paraesternales (ganglios a lo largo de las
arterias torácicas internas) y hacia los ganglios linfáticos axilares en la
axila.
El pezón, el área sobresaliente central de la cual un bebé succiona leche, está rodeado por un anillo de piel pigmentada, la areola. Durante la lactancia, las glándulas sebáceas grandes en la areola producen un sebo aceitoso que minimiza la formación de grietas en la piel del pezón. Internamente, la glándula mamaria consta de 15 a 25 lóbulos (Figura 30.2), cada uno de los cuales es una glándula alveolar compuesta distinta que se abre en el pezón. Los lóbulos están separados entre sí por una gran cantidad de tejido adiposo y tiras de tejido conectivo interlobar que forman ligamentos suspensorios de los senos. Estos ligamentos se extienden desde los músculos esqueléticos subyacentes hasta la dermis suprayacente y proporcionan, como su nombre lo indica, soporte para los senos. Los lóbulos del seno consisten en unidades más pequeñas llamadas lóbulos, que están compuestas de pequeños alvéolos agrupados como un racimo de uvas. Las paredes de los alvéolos consisten en un epitelio cuboidal simple de células secretoras de leche. Desde los alvéolos, la leche pasa a través de conductos progresivamente más grandes hasta llegar a los conductos más grandes, llamados conductos lactíferos (lactíferos = transportadores de leche), que se encuentran muy adentro del pezón. Justo en el fondo de la areola, cada conducto lactífero tiene una región dilatada llamada seno lactífero, donde la leche se acumula durante la lactancia.

Figura 30.2.
Estructura de una glándula mamaria lactante (que secreta leche). (a) Vista
anterior de un seno parcialmente disecado. (b) Sección sagital de un seno.
La estructura glandular del seno no está
desarrollada en mujeres no embarazadas. Durante la infancia, los senos de una
niña consisten solo en conductos rudimentarios, al igual que los senos de los
hombres a lo largo de la vida. Cuando una niña alcanza la pubertad, estos
conductos crecen y se ramifican, pero los lóbulos y los alvéolos todavía están
ausentes. (El agrandamiento de los senos en la pubertad se debe a la deposición
de grasa). Los alvéolos glandulares se forman finalmente, desde los conductos más
pequeños, aproximadamente a la mitad del embarazo, y la producción de leche
comienza unos días después del parto. El cáncer de mama, que generalmente surge
del sistema de conductos, es una de las principales causas de muerte por cáncer
en las mujeres.
31. Enfermedades y modificaciones de la piel humana
31.1 Tatuajes
El tatuaje, usando una aguja para
depositar pigmento en la dermis de la piel, se practicó por primera vez
alrededor de 8000 a. C. Hoy en día, los tatuajes se aplican como arte corporal,
se utilizan para expresar individualidad y hacer declaraciones personales. Los
tatuajes pueden indicar membresía en un grupo, como el ejército, una pandilla
callejera o un equipo deportivo. También se utilizan cada vez más con fines
cosméticos, como la aplicación de delineador permanente, cejas y liplines.
Pero, ¿qué pasa si el tatuaje pasa de moda
o si el pigmento migra? Hasta hace muy poco, una vez que tenía un tatuaje,
estaba esencialmente atrapado con él porque los intentos de eliminación (dermoabrasión,
criocirugía (congelación del tejido) o aplicación de productos químicos
cáusticos) dejaron cicatrices desagradables, algunas peores que el tatuaje
original. Los láseres de hoy en día generalmente pueden eliminar los tatuajes
"más antiguos", es decir, los diseños azul y negro aplicados hace
décadas. Los tatuajes de colores más nuevos son otra cosa completamente
distinta. Requieren el uso de varios láseres diferentes que emiten luz a
diferentes frecuencias para eliminar cada uno de los diferentes pigmentos. La
eliminación de tatuajes requiere de siete a nueve tratamientos con un intervalo
de aproximadamente un mes, y puede doler tanto como obtener el tatuaje en
primer lugar. Incluso entonces, los tatuajes no siempre se pueden erradicar por
completo: los pigmentos verdes y amarillos son los más difíciles de vaporizar.
Los tatuajes presentan algunos otros
riesgos. La Administración de Alimentos y Medicamentos de EE. UU. Ejerce cierto
control sobre la composición de los pigmentos utilizados, pero están aprobados
para su aplicación solo en la superficie de la piel; la seguridad de
inyectarlos debajo de la piel no está bien establecida. Además, las leyes
estatales varían ampliamente. Algunos estados prohíben la práctica por
completo, y otros no tienen regulaciones en absoluto. El tatuaje implica agujas
y sangrado, los requisitos de capacitación para los profesionales son mínimos y
existe el riesgo de reacciones alérgicas y enfermedades infecciosas
transmitidas por la sangre, como la hepatitis. (Incluso si el profesional
utiliza agujas estériles, pueden sumergirse en pigmentos que están contaminados
con la sangre de clientes anteriores). Incluso con la disponibilidad de
técnicas de extracción con láser, tiene sentido considerar los riesgos de
hacerse un tatuaje.
31.2 Acné
En algunos adolescentes, se produce tanto
sebo que no puede ser conducido de las glándulas lo suficientemente rápido.
Cuando una glándula sebácea se bloquea por el sebo, aparece un punto blanco en
la superficie de la piel. Si el material se oxida y se seca, se oscurece para
formar un punto negro (el color oscuro de un punto negro no se debe a la
suciedad). Es probable que las glándulas sebáceas bloqueadas se infecten con
bacterias y produzcan un grano. Las bacterias descomponen el sebo en ácidos grasos
irritantes. Estos ácidos, junto con los productos bacterianos mismos, inducen
la inflamación, especialmente cuando la masa infectada estalla del folículo
hacia la dermis circundante. El acné resultante puede ser leve o extremadamente
severo, lo que lleva a cicatrices permanentes. El tratamiento del acné aborda
los diferentes factores causales: aumento de la producción de sebo y formación
de puntos blancos, inflamación e infección bacteriana. Los medicamentos tópicos
como el peróxido de benzoilo tratan el acné inflamatorio; los retinoides
tópicos, derivados de la vitamina A, actúan para prevenir la formación de
puntos blancos; y los antibióticos destruyen las bacterias que causan el acné.
A menudo se usan combinaciones de tratamientos para obtener resultados máximos.
31.3 Quemaduras
Expuesta directamente a los peligros y
gérmenes del mundo exterior, la piel puede desarrollar más de mil condiciones y
dolencias diferentes. Los trastornos cutáneos más comunes son infecciones
bacterianas, virales o de levaduras. Las amenazas más graves para la piel son
las quemaduras y el cáncer de piel.
Las quemaduras son una amenaza devastadora
para el cuerpo, principalmente debido a sus efectos sobre la piel. Una
quemadura es el daño tisular infligido por el calor, la electricidad, la
radiación, la fricción extrema o ciertos productos químicos nocivos. La amenaza
inmediata a la vida por quemaduras graves es una pérdida catastrófica de
fluidos corporales. El edema inflamatorio es severo. A medida que el fluido se
filtra de las superficies quemadas, el cuerpo pierde rápidamente agua y sales
esenciales. Esta deshidratación a su vez conduce a un shock circulatorio fatal;
es decir, circulación sanguínea inadecuada causada por la reducción en el
volumen sanguíneo. Para salvar al paciente, el equipo médico debe reemplazar
los líquidos perdidos de inmediato. Una vez que ha pasado la crisis inicial, la
infección se convierte en la principal amenaza. Los patógenos pueden invadir
fácilmente áreas donde se destruye la barrera cutánea.
Las quemaduras se clasifican por su
gravedad (profundidad) como quemaduras de primer, segundo o tercer grado (Figura 31.1).
Las quemaduras de tercer grado son las más graves.
👉 En las quemaduras de primer grado,
solo se daña la epidermis. Los síntomas incluyen enrojecimiento, hinchazón y
dolor: la reacción inflamatoria típica al daño tisular. En general, las
quemaduras de primer grado sanan en pocos días sin atención especial. Las
quemaduras solares suelen ser quemaduras de primer grado.
👉 Las quemaduras de segundo grado
implican lesiones en la epidermis y la parte superior de la dermis. Los
síntomas se parecen a los de las quemaduras de primer grado, pero las ampollas
también aparecen a medida que se acumula líquido entre las capas epidérmica y
dérmica. La piel se regenera con poca o ninguna cicatrización en 3 a 4 semanas
si se tiene cuidado para prevenir la infección. Las quemaduras de primer y
segundo grado se consideran quemaduras de espesor parcial.
👉 Las quemaduras de tercer grado consumen todo el grosor de la piel y, por lo tanto, son quemaduras de grosor completo. El área quemada aparece blanca, roja o ennegrecida. Aunque la piel podría eventualmente regenerarse, generalmente es imposible esperar esto debido a la pérdida de líquido y la infección. Por lo tanto, la piel de otras partes del cuerpo del paciente debe injertarse en el área quemada. Tal injerto, en el que un individuo es donante y receptor, se llama autoinjerto.

Figura 31.1. Quemaduras.
En general, las quemaduras se consideran
críticas si existe alguna de las siguientes condiciones:
👉 Más del 10% del cuerpo tiene
quemaduras de tercer grado;
👉 25% del cuerpo tiene quemaduras de
segundo grado;
👉 hay quemaduras de tercer grado en
la cara, manos o pies. Una forma rápida de estimar cuánta superficie se ve
afectada por una quemadura es usar la regla de los nueves (Figura 31.1c).
Este método divide la superficie del
cuerpo en 11 regiones, cada una de las cuales representa el 9% (o un múltiplo
del 9%) del área total del cuerpo. (Los valores que se muestran en la figura
humana en la Figura 31.1c
indican valores del área de superficie para las superficies del cuerpo
anterior; los valores totales para cada región se indican a la derecha de la
ilustración). Este método es solo aproximadamente exacto, por lo que se usan
tablas especiales cuando se necesita mayor precisión.
Para los pacientes cuyas quemaduras son
demasiado grandes o que, de otro modo, son malos candidatos para autoinjertos,
ahora están disponibles buenos revestimientos artificiales. La piel artificial
consiste en una "dermis" hecha de colágeno bovino y una
"epidermis" hecha de silicona. La propia dermis del paciente
reemplaza y reabsorbe gradualmente la artificial, y la lámina de silicona luego
se despega y se reemplaza con una red de células epidérmicas cultivadas de la
propia piel del paciente. La piel artificial no es rechazada por el cuerpo,
salva vidas y produce cicatrices mínimas. Sin embargo, es más probable que se
infecte que un autoinjerto.
31.4 Envejecimiento
de la piel
La epidermis se desarrolla a partir del
ectodermo embrionario, y la dermis y la hipodermis se desarrollan a partir del
mesodermo. Los melanocitos se derivan de las células de la cresta neural que
migran hacia el ectodermo durante los primeros 3 meses de desarrollo prenatal.
Al final del cuarto mes, la piel está bastante bien formada: la epidermis tiene
todas sus capas, las papilas dérmicas son evidentes y los apéndices epidérmicos
están presentes en forma rudimentaria. Durante el quinto y sexto mes, el feto
se cubre con una suave capa de pelos delicados llamada lanugo. Esta capa se
desprende al séptimo mes y aparecen vellos.
Cuando nace un bebé, su piel está cubierta
de vernix caseosa, una sustancia de aspecto cursi producida por las glándulas
sebáceas. Protege la piel del feto del contacto constante con el líquido
amniótico. La piel y la hipodermis de un recién nacido son muy delgadas, pero
se engrosan durante la infancia y la niñez. Con el inicio de la adolescencia,
las glándulas sebáceas se vuelven más activas y puede aparecer acné. El acné
generalmente disminuye en la edad adulta temprana, y la piel alcanza su
apariencia óptima en los 20 y 30 años. A partir de entonces, la piel comienza a
mostrar los efectos nocivos de los continuos ataques ambientales, como la
abrasión, el viento, el Sol y los productos químicos. Las incrustaciones y
diversos tipos de inflamación de la piel, llamada dermatitis, se vuelven más comunes.
En la mediana edad, las sustancias
lubricantes y suavizantes producidas por las glándulas comienzan a disminuir.
Como resultado, la piel se vuelve seca. Las personas con piel naturalmente
grasa pueden evitar esta sequedad, por lo que su piel a menudo envejece bien.
La mayoría de los aspectos del envejecimiento de la piel no son intrínsecos,
sino que son causados por la luz solar en un proceso llamado
"fotoenvejecimiento". La piel envejecida que ha sido protegida del Sol
ha perdido algo de elasticidad y es más delgada (al igual que la hipodermis
debajo de ella), pero permanece sin arrugas. y sin marcar. La piel expuesta al
sol, por el contrario, está arrugada, suelta, inelástica, correosa y tiene
manchas pigmentadas llamadas "manchas hepáticas". En la dermis de esa
piel envejecida por el Sol, la cantidad de colágeno ha disminuido y un material
anormal que contiene elastina se ha acumulado. Gran parte de este cambio se
debe a la activación inducida por los rayos UV de enzimas que degradan el
colágeno y otros componentes de la dermis.
Al bloquear los rayos UV, grandes
cantidades de melanina protegen la piel del fotoenvejecimiento. Esta es la
razón por la cual las personas de piel clara, que tienen poca melanina para
comenzar, muestran cambios relacionados con la edad a una edad más temprana que
las personas con piel y cabello más oscuros. Por la misma razón, las personas
de piel oscura parecen más jóvenes que los blancos.
32. Cáncer de piel
|| Regresar al Índice ||
Muchos tipos de tumores surgen en la piel. La mayoría son benignos (verrugas, por ejemplo), pero algunas son malignos. El cáncer de piel es el tipo más común de cáncer, con aproximadamente un millón de casos nuevos que aparecen cada año en los Estados Unidos. Como se mencionó anteriormente, el factor de riesgo más importante para el cáncer de piel es la sobreexposición a los rayos UV en la luz solar. Los datos recientes muestran un mayor riesgo de los tres tipos de cáncer de piel en personas con antecedentes de bronceado en interiores.
Figura 32.1. Fotografías
de cánceres de piel.
32.1 Carcinoma
de células basales
El carcinoma basocelular es el cáncer de
piel menos maligno y más común (Figura 32.1a).
¡Más del 30% de todos los blancos lo obtienen en sus vidas! Las células del
estrato basal proliferan, invaden la dermis y la hipodermis y causan erosiones tisulares
allí. Las lesiones más comunes de este cáncer son nódulos brillantes en forma
de cúpula en áreas de la cara expuestas al sol. Posteriormente, estos nódulos
desarrollan una úlcera central y un borde con cuentas "perlado". A
menudo sangran. El carcinoma basocelular crece de forma relativamente lenta y
rara vez se produce metástasis. La cura completa por escisión quirúrgica u
otros métodos de extracción es la regla en el 99% de los casos.
32.2 Carcinoma
de células escamosas
El carcinoma de células escamosas (Figura 32.1b)
surge de los queratinocitos del estrato espinoso. La lesión aparece como una
pápula enrojecida, irregular y escamosa (elevación pequeña y redondeada) que
tiende a crecer rápidamente y a hacer metástasis si no se elimina. Sin embargo,
si se trata temprano, la posibilidad de una cura completa es buena y la tasa de
curación general es del 99%. El carcinoma se puede extirpar mediante
radioterapia, cirugía o cremas para la piel que contienen medicamentos contra
el cáncer.
32.3 Melanoma
El melanoma, un cáncer de melanocitos, es
el tipo más peligroso de cáncer de piel (Figura 32.1c).
Los melanocitos se derivan de las células de la cresta neural, que deambulan
ampliamente durante el desarrollo embrionario. Esta predisposición a la
migración explica la naturaleza invasiva del melanoma. El melanoma representa
solo alrededor de 1 de cada 20 cánceres de piel, pero está aumentando
rápidamente en países con poblaciones de piel clara, de 3% a 8% por año en los
Estados Unidos. El melanoma puede originarse donde haya pigmento, pero a menudo
surge de lunares existentes, que generalmente aparecen como un parche oscuro en
expansión. Debido a que las células de melanoma hacen metástasis rápidamente en
los vasos circulatorios circundantes, la clave para sobrevivir al melanoma es
la detección temprana. Las tasas de supervivencia disminuyen con el aumento del
grosor del melanoma, el grado de afectación de los ganglios linfáticos cercanos
y el grado de metástasis. En casos avanzados, el tratamiento quirúrgico es
seguido por inmunoterapia, radioterapia o, más recientemente, terapia génica
dirigida con resultados prometedores para reducir los tumores y / o prolongar
la vida.
La Sociedad Estadounidense del Cáncer
sugiere que las personas con exposición frecuente al Sol examinen regularmente
su piel en busca de lunares y nuevas manchas de pigmento, aplicando la regla
ABCD para reconocer el melanoma:
👉 A, Asimetría: las dos mitades de
la mancha o lunar no coinciden;
👉 B, irregularidad del borde: los
bordes tienen muescas;
👉 C, Color: la mancha de pigmento
contiene varios colores, incluidos negros, marrones, morenos y, a veces, azules
y rojos;
👉 D, diámetro: más de 6 mm (más
grande que un borrador de lápiz).
Algunos expertos han descubierto que
agregar una
👉 E, para Elevación sobre la
superficie de la piel, mejora el diagnóstico, por lo que utilizan la regla ABCD
(E).
32.4 Cáncer de
seno
El cáncer de seno, la segunda causa más común de muerte por cáncer en mujeres y el asesino del 3% de las mujeres de EE. UU., Generalmente surge de los conductos más pequeños en los lóbulos del seno. Una de cada ocho mujeres desarrollará cáncer de seno en su vida. Se aconseja a las mujeres que realicen autoexámenes mensuales de los senos para detectar bultos en el seno, un cambio en la forma del seno, descamación en el pezón o areola, secreción del pezón, hoyuelos o una textura de piel de naranja en el seno. Los factores de riesgo para el cáncer de mama incluyen antecedentes familiares de la enfermedad, menopausia tardía, inicio temprano de la pubertad, primer nacimiento vivo después de los 30 años y terapia de reemplazo de estrógenos posmenopáusica. Con la posible excepción de los antecedentes familiares, todos estos factores de riesgo reflejan una mayor exposición de por vida a los estrógenos, que estimulan la división de las células del conducto y promueven el cáncer. El cáncer de seno es un cáncer de rápida propagación que hace metástasis desde el seno a través de los vasos linfáticos hasta los ganglios linfáticos axilares y paraesternales, y luego hacia la pared torácica, los pulmones, el hígado, el cerebro y los huesos. Si las metástasis alcanzan los pulmones o el hígado, el tiempo promedio de supervivencia es inferior a 6 meses.

Figura 32.2. Mamograma.
Las técnicas de detección para permitir la
detección temprana reducen efectivamente la mortalidad por cáncer de mama. Se
aconseja a las mujeres mayores de 40 años que se realicen una mamografía
(imagen de rayos X del seno) anualmente (Figura 32.2).
Las masas sospechosas se biopsian y se examinan microscópicamente para
determinar si son cancerosas. Si uno o más lo son, los ganglios linfáticos
axilares se extirpan y se examinan para detectar células cancerosas; El
porcentaje de estos ganglios axilares afectados es el mejor indicador de
metástasis y el predictor más confiable de supervivencia.
Referencias bibliográficas
|| Regresar al Índice ||
Alexopoulos, C. J., Mims, C. W., & Blackwell, M. (1996). Introductory
mycology. 1996. New York, John Wiley & Sons, Inc.
Alibardi, L., Knapp, L. W. and, & Sawyer, R. H. (2006). Beta-keratin
localization in developing alligator scales and feathers in relation to the
development and evolution of feathers. Journal of submicroscopic cytology
and pathology, 38(2–3), 175–192.
Artigas, M., Martnez, R. A., Glick, T. F., & Martínez, R. A. (2006). Negotiating
Darwin: the Vatican confronts evolution, 1877–1902. JHU Press.
Barsanti, L., & Gualtieri, P. (2014). Algae: anatomy, biochemistry,
and biotechnology. CRC press.
Becker, B. (2000). The cell surface of flagellates. Taylor and
Francis New York.
Beleza, S.,
Santos, A. M., McEvoy, B., Alves, I., Martinho, C., Cameron, E., … Rocha, J. (2012).
The timing of pigmentation lightening in Europeans. Molecular biology and
evolution, 30(1), 24–35.
Belk, C., & Maier, V. B. (2013). Biology Science for Life with
physiology. (Pearson, Ed.) (4a ed.).
Bender, J. M. M. K. S., Madigan, D. H. B., Michael, T., & Stahl, D. A.
(2015). Brock Biology of Microorganisms. Boston: Pearson.
Bergquist, P. R. (1985). Poriferan relationships. The origins and
relationships of lower invertebrates, 28, 14–27.
Bock, W. J. (2000). Explanatory history of the origin of feathers. American
Zoologist, 40(4), 478–485.
Bond, C. (1998). Keeping Up with the Sponges. Natural History, 106,
22–25.
Brusatte, S. L., O’Connor, J. K., & Jarvis, E. D. (2015). The origin
and diversification of birds. Current Biology, 25(19), R888–R898.
Brusca, R., Brusca, G. J., & Haver, N. J. (2003). Invertebrates
(2a ed.). Sinauer Associates.
Conover, M. R., & Miller, D. E. (1980). Rictal bristle function in
willow flycatcher. The Condor, 82(4), 469–471.
Czerkas, S. A., & Ji, Q. (2002). A new rhamphorhynchoid with a
headcrest and complex integumentary structures. Feathered dinosaurs and the
origin of flight, 1, 15–41.
Darwin, C. (1872). The descent of man, and selection in relation to sex
(Vol. 2). D. Appleton.
Dimond, C. C., Cabin, R. J., & Brooks, J. S. (2011). Feathers,
dinosaurs, and behavioral cues: defining the visual display hypothesis for the
adaptive function of feathers in non-avian theropods. Bios, 82(3),
58–64.
Egger, B., Lapraz, F., Tomiczek, B., Müller, S., Dessimoz, C., Girstmair,
J., … Beli, E. (2015). A transcriptomic-phylogenomic analysis of the
evolutionary relationships of flatworms. Current Biology, 25(10),
1347–1353.
Elias, P. M., Menon, G., Wetzel, B. K., & Williams, J. (Jack) W.
(2010). Barrier requirements as the evolutionary “driver” of epidermal
pigmentation in humans. American Journal of Human Biology: The Official
Journal of the Human Biology Association, 22(4), 526–537.
Eswaran, R. H., Harpending, H., & McCullough, J. (2004). Genetic
Variation at the MC1R Locus and the Time since Loss of Human Body Hair1.
Foth, C. (2012). On the identification of feather structures in stem-line
representatives of birds: evidence from fossils and actuopalaeontology. Paläontologische
Zeitschrift, 86(1), 91–102.
Furnas, M. J. (1990). In situ growth rates of marine phytoplankton:
approaches to measurement, community and species growth rates. Journal of
Plankton Research, 12(6), 1117–1151.
George, H. (1998). Magical mushrooms, mischievous molds. Princeton, NJ:
Princeton University Press.
Gibbons, A. (2007). European skin turned pale only recently, gene
suggests. Science, 316(5823), 364.
Glenn, T. C., French, J. O., Heincelman, T. J., Jones, K. L., &
Sawyer, R. H. (2008). Evolutionary relationships among copies of feather beta (β) keratin genes from several avian orders. Integrative
and comparative biology, 48(4), 463–475.
Godefroit, P., Sinitsa, S. M., Dhouailly, D., Bolotsky, Y. L., Sizov, A.
V, McNamara, M. E., … Spagna, P. (2014). A Jurassic ornithischian dinosaur from
Siberia with both feathers and scales. Science, 345(6195),
451–455.
Gram, C. (1884). Ueber die isolirte Farbung der Schizomyceten in
Schnitt-und Trockenpraparaten. Fortschritte der Medicin, 2,
185–189.
Grzesik, P., Pryce, E. N., Bhalala, A., Vij, M., Ahmed, R., Etienne, L., …
McCaffery, J. M. (2019). Functional Domains of the Herpes Simplex Virus Type 1
Tegument Protein pUL37: The Amino Terminus is Dispensable for Virus Replication
in Tissue Culture. Viruses, 11(9), 853.
Harrison, S. C. (2008). Viral membrane fusion. Nature structural &
molecular biology, 15(7), 690.
Heijenoort, J. van. (2001). Formation of the glycan chains in the
synthesis of bacterial peptidoglycan. Glycobiology, 11(3),
25R–36R.
Hoefnagels, M. (2015). Biology: concepts and investigations (3a
ed.). McGraw-Hill New York.
Howland, J. L. (2000). The surprising archaea: discovering another
domain of life. Oxford University Press on Demand.
Hugenholtz, P. (2002). Exploring prokaryotic diversity in the genomic era.
Genome biology, 3(2), reviews0003-1.
Hwang, S. H., NORELL, M. A., Qiang, J. I., & Keqin, G. A. O. (2002).
New specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from
northeastern China. American Museum Novitates, 1–44.
Jablonski, N. G. (2004). The evolution of human skin and skin color. Annu.
Rev. Anthropol., 33, 585–623.
Jablonski, N. G. (2011). Why human skin comes in colors. AnthroNotes.,
32(1).
Jablonski, N. G. (2012). Living color: The biological and social
meaning of skin color. Univ of California Press.
Jablonski, N. G., & Chaplin, G. (2000). The evolution of human skin
coloration. Journal of human evolution, 39(1), 57–106.
Joseleau-Petit, D., Liébart, J.-C., Ayala, J. A., & D’Ari, R. (2007).
Unstable Escherichia coli L forms revisited: growth requires peptidoglycan
synthesis. Journal of bacteriology, 189(18), 6512–6520.
Juzeniene, A., Setlow, R., Porojnicu, A., Steindal, A. H., & Moan, J.
(2009). Development of different human skin colors: a review highlighting
photobiological and photobiophysical aspects. Journal of Photochemistry and
Photobiology B: Biology, 96(2), 93–100.
Kardong, K. V. (2011). Vertebrates, comparative anatomy, function,
evolution (6a ed.). McGraw-Hill New York.
Karp, G. C. (2013). Cell and Molecular Biology, Concepts and
Experiments (7a ed.). USA: Wiley Online Library.
Kellner, A. W. A., Wang, X., Tischlinger, H., de Almeida Campos, D., Hone,
D. W. E., & Meng, X. (2009). The soft tissue of Jeholopterus (Pterosauria,
Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing
membrane. Proceedings of the Royal Society B: Biological Sciences, 277(1679),
321–329.
Koch, A. L. (2003). Bacterial wall as target for attack past, present, and
future research. Clinical microbiology reviews, 16(4), 673–687.
Koivikko, R., Loponen, J., Honkanen, T., & Jormalainen, V. (2005).
Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga
Fucus vesiculosus, with implications on their ecological functions. Journal
of chemical ecology, 31(1), 195–212.
Littlewood, D. T. J., Rohde, K., Bray, R. A., & Herniou, E. A. (1999).
Phylogeny of the Platyhelminthes and the evolution of parasitism. Biological
Journal of the Linnean Society, 68(1–2), 257–287.
Lovette, I. J., & Fitzpatrick, J. W. (2016). Handbook of bird
biology. John Wiley & Sons.
Mason, K. A., Losos, J. B., Singer, S. R., & Raven, P. H. (2014). Biology
(7a ed.). McGraw-Hill New York.
Moore, J. (2006). Introduction to the invertebrates (2a ed.).
Cambridge: Cambridge Univ Press.
Norton, H. L., Kittles, R. A., Parra, E., McKeigue, P., Mao, X., Cheng,
K., … Shriver, M. D. (2006). Genetic evidence for the convergent evolution of
light skin in Europeans and East Asians. Molecular biology and evolution,
24(3), 710–722.
Preisig, H. R., Anderson, O. R., Corliss, J. O., Moestrup, Ø., Powell, M.
J., Roberson, R. W., & Wetherbee, R. (1994). Terminology and nomenclature
of protist cell surface structures. En The Protistan Cell Surface (pp.
1–28). Springer.
Prum, R. O. (1999). Development and evolutionary origin of feathers. Journal
of Experimental Zoology, 285(4), 291–306.
Prum, R. O., & Brush, A. H. (2002). The evolutionary origin and
diversification of feathers. The Quarterly review of biology, 77(3),
261–295.
Prum, R. O., & Brush, A. H. (2003). Which came first, the feather or
the bird? Scientific American, 288(3), 84–93.
Prum, R. O., & Brush, A. H. (2014). Which came first, the feather or
the bird? Scientific American, 23, 76–85.
Raper, K. B. (2014). The dictyostelids. Princeton University Press.
RAVEN, J. A. (1983). The transport and function of silicon in plants. Biological
reviews, 58(2), 179–207.
Rhoades, R. A., & Bell, D. R. (2013). Medical Physiology,
Principles for Clinical Medicine (4a ed.). Baltimore: Lippincott Williams
& Wilkins.
Rogers, A., Iltis, D., & Wooding, S. (2004). Genetic variation at the
MC1R locus and the time since loss of human body hair. Current Anthropology,
45(1), 105–108.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of
Biology (10a ed.). Sinauer & MacMillian.
Sengbusch, P. V. (2003). Interactions between Plants and Fungi: the
Evolution of their Parasitic and Symbiotic Relations. biologie. uni-hamburg.
de.
Simon, E. J., Dickey, J. L., & Reece, J. B. (2013). Essential
biology (Pearson).
Solomon, E. P., Berg, L. R., & Martin, D. W. (2008). Biology 8th
edition. Thomsom Brooks/Cole, USA.
Starr, C., Evers, C., & Starr, L. (2013). Biology: Today and
Tomorrow With Physiology (4a ed.). Brooks/Cole.
Stern, K. R., Bidlack, J. E., & Jansky, S. H. (2008). Introductory
Plant Biology (11a ed.). McGraw-Hill New York.
Sumida, S. S., & Brochu, C. A. (2000). Phylogenetic context for the
origin of feathers. American Zoologist, 40(4), 486–503.
Unwin, D. M., & Bakhurina, N. N. (1994). Sordes pilosus and the nature
of the pterosaur flight apparatus. Nature, 371(6492), 62.
van Dijk, E. M., & Reydon, T. A. C. (2010). A conceptual analysis of
evolutionary theory for teacher education. Science & Education, 19(6–8),
655–677.
Vinther, J. (2017). The true colors of dinosaurs. Scientific American,
316(3), 50–57.
Vinther, J. (2018). The True Colors of Dinosaurs. Scientific American,
27, 12–19.
Vinther, J., Briggs, D. E. G., Prum, R. O., & Saranathan, V. (2008).
The colour of fossil feathers. Biology Letters, 4(5), 522–525.
Wang, M., Zheng, X., O’Connor, J. K., Lloyd, G. T., Wang, X., Wang, Y., …
Zhou, Z. (2015). The oldest record of ornithuromorpha from the early cretaceous
of China. Nature Communications, 6, 6987.
Wayne, R. (2009). Plant Cell Biology (1a ed.). San Diego: Elsevier.
Webster, J., & Weber, R. (2007). Introduction to fungi.
Cambridge University Press.
White, D., Drummond, J. T., & Fuqua, C. (1995). The physiology and
biochemistry of prokaryotes.
Witton, M. P. (2013). Pterosaurs: natural history, evolution, anatomy.
Princeton University Press.
Xu, X. (2019). Feather evolution: Looking up close and through deep time. Science
Bulletin, 64(9), 563–564.
Xu, X., & Guo, Y. (2009). The origin and early evolution of feathers:
insights from recent paleontological and neontological data. Vertebrata
PalAsiatica, 47(4), 311–329.
Xu, X., Norell, M. A., Kuang, X., Wang, X., Zhao, Q., & Jia, C.
(2004). Basal tyrannosauroids from China and evidence for protofeathers in
tyrannosauroids. Nature, 431(7009), 680–684.
Xu, X., Wang, K., Zhang, K., Ma, Q., Xing, L., Sullivan, C., … Wang, S.
(2012). A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature,
484(7392), 92–95.
Xu, X., & Zhang, F. (2005). A new maniraptoran dinosaur from China
with long feathers on the metatarsus. Naturwissenschaften, 92(4),
173–177.
Xu, X., Zhou, Z., Wang, X., Kuang, X., Zhang, F., & Du, X. (2003).
Four-winged dinosaurs from China. Nature, 421(6921), 335–340.
Yang, Z., Jiang, B., McNamara, M. E., Kearns, S. L., Pittman, M., Kaye, T.
G., … Benton, M. J. (2019). Pterosaur integumentary structures with complex
feather-like branching. Nature ecology & evolution, 3(1), 24.
York, A. (2017). Viral evolution: On the origin of capsids. Nature
Reviews Microbiology, 15(5), 256.
Yu, X., Shah, S., Lee, M., Dai, W., Lo, P., Britt, W., … Zhou, Z. H. (2011).
Biochemical and structural characterization of the capsid-bound tegument
proteins of human cytomegalovirus. Journal of structural biology, 174(3),
451–460.
Zhang, F., Kearns, S. L., Orr, P. J., Benton, M. J., Zhou, Z., Johnson,
D., … Wang, X. (2010). Fossilized melanosomes and the colour of Cretaceous
dinosaurs and birds. Nature, 463(7284), 1075–1078.
Zheng, X.-T., You, H.-L., Xu, X., & Dong, Z.-M. (2009). An Early
Cretaceous heterodontosaurid dinosaur with filamentous integumentary
structures. Nature, 458(7236), 333.
Zimmer, C. (2010). The tangled bank: an introduction to evolution.
No hay comentarios:
Publicar un comentario