
Índice
Portada

1. Introducción
|| Regresar al Índice ||
El esqueleto es la parte del cuerpo que
forma la estructura de soporte de un organismo. También puede verse como el
marco óseo del cuerpo que proporciona soporte, forma y protección a los tejidos
blandos y órganos delicados de los animales. Existen varios tipos diferentes de
esqueleto: el exoesqueleto, que es la capa externa estable de un organismo, el
endosqueleto, que forma la estructura de soporte dentro del cuerpo, el
hidroesqueleto, un esqueleto flexible soportado por la presión del fluido y el
citoesqueleto presente en el citoplasma. de todas las células, incluidas las
bacterias y las arqueas. El término proviene del griego σκελετός (skeletós),
que significa 'seco'.
1.1 Robert
Thomas Bakker
(1945-vivo a 2021) es un paleontólogo
estadounidense que ayudó a remodelar las teorías modernas sobre los
dinosaurios, particularmente al agregar apoyo a la teoría de que algunos
dinosaurios eran endotérmicos (de sangre caliente). Junto con su mentor John
Ostrom, Bakker fue responsable de iniciar el actual "renacimiento de los
dinosaurios" en los estudios paleontológicos, comenzando con el artículo
de Bakker "Dinosaur Renaissance" en la edición de abril de
1975 de Scientific American. Su especialidad es el contexto ecológico y
el comportamiento de los dinosaurios.

Bakker ha sido uno de los principales
defensores de la teoría de que los dinosaurios eran de sangre caliente,
inteligentes, rápidos y adaptables. Publicó su primer artículo sobre la
endotermia de los dinosaurios en 1968. Su obra fundamental, The Dinosaur
Heresies, se publicó en 1986. Reveló la primera evidencia del cuidado de
los padres en los sitios de anidación de Allosaurus. También observó
evidencia en apoyo de la teoría del equilibrio puntuado de Eldredge y Gould en
las poblaciones de dinosaurios. Bakker actualmente se desempeña como curador de
paleontología del Museo de Ciencias Naturales de Houston.
2. Tipos de esqueleto
|| Regresar al Índice ||
Todos los seres vivos deben interactuar
con su ambiente, y estas interacciones dependen de las formas que adquieren sus
cuerpos. El sistema encargado de dar forma al cuerpo de un ser vivo se denomina
esqueleto. Los esqueletos poseen funciones asociadas a la de dar forma, pues la
forma de un ser vivo es un marco que sirve como punto de anclaje a otros
sistemas que permiten el movimiento, el sostenimiento de otras estructuras para
funciones diversas o la formación de partes duras que sirven para proteger.
Estas funciones pueden realizarse ya sea
por partes duras, secreciones o incluso la turgencia de un tejido inflado por
líquidos (Brusca, Brusca, & Haver, 2003). Algunos tipos específicos de esqueletos pueden tener funciones
extra asociadas como el movimiento o la formación de otros tejidos no
esqueléticos. Nosotros los seres humanos somos vertebrados, eso significa que
tenemos un esqueleto de cierto tipo y tendemos a juzgar los demás esqueletos en
base a nuestro esqueleto. Para poder estudiar las funciones del esqueleto
debemos ampliar nuestra definición o nuestra visión de los tipos de esqueleto.
2.1 Esqueleto hidrostático
Los hidroesqueletos funcionan básicamente
como gatos hidráulicos y placas incompresibles. Su funcionamiento se debe a dos
propiedades generales del agua y las soluciones líquidas, la incompresibilidad
y la transmisión del trabajo mediante cambios de presión.

Figura 2.1. Poco más que
una bolsa de líquidos, el esqueleto hidrostático ha demostrado ser uno de los
esqueletos más versátiles en los animales.
El agua es incompresible bajo condiciones
normales, la presión debe crecer a cantidades astronómicas para generar su
cristalización, sin embargo, los seres vivos conviven bajo condiciones
relativamente normales de presión. Un esqueleto hidrostático adquiere su forma
gracias a los tejidos blandos que los contienen, y la presión causada en una
región transfiere movimiento a otra, lo cual los hace funcionar también como un
sistema muscular (Brusca et al., 2003).
2.2 Esqueleto sólido
Los esqueletos sólidos se basan en el
endurecimiento de algunos tejidos, lo cual los hace servir como puntos de
anclaje para el sistema muscular.

Figura 2.2. Algunos
esqueletos sólidos pueden dar forma a paisajes geográficos completos como la
madera o los arrecifes de coral. La madera es un tipo de tejido vegetal que
cumple con las funciones de un esqueleto sólido.
Los esqueletos sólidos generalmente
constan de dos partes, una fibra que sirve de marco, da forma y permite la flexibilidad
–semejante a los tubos de hierro en el concreto – así como de una sustancia que
se solidifique de forma muy firme que rodea al marco.
2.3 Endoesqueleto
Es un sistema esquelético que se encuentra
contenido por una capa suave como una membrana o la piel, todos los
hidroesqueletos son endoesqueletos. El esqueleto de los vertebrados
también es un endoesqueleto (Kardong, 2011).

Figura 2.3. Los
endoesqueletos y exoesqueletos duros son la principal evidencia del modo en que
ha cambiado la vida en la tierra.
Los endoesqueletos de los vertebrados son
el sistema que más fácilmente se preserva como un fósil y es una de las fuentes
de información más rica sobre la biología y ecología de un vertebrado,
especialmente el cráneo. Esta es la razón por la cual el capítulo sobre el
esqueleto es el segundo más extenso después del de reproducción, básicamente
todo lo que conocemos de la vida del pasado viene de esqueletos fósiles.
2.4 Exoesqueleto
Son esqueletos sólidos que forman conchas
o cápsulas alrededor del ser vivo, dándoles su forma y
protegiéndolos (Brusca et al., 2003). Algunos seres vivos poseen sus
esqueletos por fuera, los cuales sirven como armaduras protectoras, y de hecho
la analogía con la armadura es aún más pertinente, ya que se emplean trazas de
metales pesados para reforzar las placas del exoesqueleto.

Figura 2.4. Los
exoesqueletos tienden a ser pesados y limitar el crecimiento.
En algunas ocasiones un esqueleto puede
ser al mismo tiempo endoesqueleto y exoesqueleto, la forma generalmente depende
de estado de desarrollo del ser vivo, es decir, la estructura dura comienza a
ser segregada en el interior del ser vivo –endoesqueleto –pero a medida que el
ser vivo envejece la estructura crece y debe salir por una parte del animal
configurándose como una secreción dura –exoesqueleto.
En las siguientes secciones revisaremos
los principales tipos de estructuras que cumplen de forma análoga con la
función de esqueleto. Muchas de estas se desarrollaron de forma independiente
durante el curso de la evolución, para finalmente concentrarnos en los
vertebrados, donde los esqueletos son homólogos –descienden de un ancestro
común. Por tal razón, será en el caso de los vertebrados donde se analice al
esqueleto en función de un proceso evolutivo con mayor detalle.
2.5 El primer
esqueleto
A diferencia del sistema muscular, todos
los seres vivos presentan al menos algún tupo de esqueleto que rige la forma y
por lo tanto, muchas de las funciones internas del organismo. Es más, este
capítulo debería introducir como factor inicial al propio citoesqueleto
celular, pero este ya ha sido trabajado dentro del tema de la célula, así que
solo mencionaremos algunos esbozos de su funcionamiento (Karp, 2013).
Tanto procariotas como eucariotas dependen
de un sistema de proteínas rígidas que forman un andamio interno, el cual sirve
como sistema de anclaje, y sostén para la membrana celular denominado de forma
apropiada, citoesqueleto, que significa esqueleto de la célula (Sadava, Berenbaum, & Hillis,
2014). Por
muchos años se pensó que el citoesqueleto era una característica exclusiva de
las células eucariotas, pero estudios recientes han demostrado que las células
procariotas también poseen un sistema de citoesqueleto (Erb & Pogliano, 2013; Jékely,
2014; Wickstead & Gull, 2011). Algunos autores sostienen que los citoesqueletos anteceden a las
membranas celulares como las estructuras sobre las cuales, las primeras
sustancias químicas de la vida pudieron haberse desarrollado durante los
procesos de química prebiótica (De Duve & Pizano, 1995).
3. Forma y función biológica
|| Regresar al Índice ||
Para la gran mayoría de los casos, la
forma de un ser vivo se encuentra relacionada con las funciones que debe
desempeñar para mantenerse con vida, lo cual va desde el nivel molecular en las
proteínas, hasta la estructura del cuerpo de un rorcual azul, el animal más
grande que ha vivido en todos los tiempos que tengamos registros. Encontrar
explicaciones para la asociación entre forma y función se ha convertido en una
de las mayores tendencias teóricas al interior de la biología. Las primeras
explicaciones se basaban en la teleología o teología natural,
la cual argumentaba que Dios había creado a cada ser vivo para cumplir un
propósito y por eso les había infundido con formas de supervivencia específicas
dependientes de sus estructuras físicas (Ginsburg & Jablonka, 2015). En la
actualidad la explicación se basa en la Teoría Sintética de la Evolución,
la cual afirma que la asociación de forma y función depende de un proceso de
trasformación física, desde organismos poco especializados, poco específicos y
que cumplen una determinada función de forma muy limitada, hasta descendientes
con formas optimizadas que a partir de cierto punto de mejora ya no pueden
cambiar más, ya que han alcanzado un óptimo desarrollo evolutivo para una
función concreta (Kutschera & Niklas, 2004).

Figura 3.1. El oso panda es
un ejemplo de un organismo pobremente adatado debido a que ha cambiado de nicho
recientemente en su evolución, pero que sobrevive por ausencia de competencia.
La segunda perspectiva ayuda a explicar
porque algunas partes del cuerpo de los seres vivos no parecen ser óptimas para
la función que deberían desarrollar, y el ejemplo más relevante es el sistema
digestivo de los osos panda. Los osos panda carecen de un sistema digestivo
largo y complejo para procesar la celulosa del bambú, y por eso deben comer
todo el día, casi todas sus vidas sin descanso, lo cual se solucionaría con un
compartimento de rumia, que procese la celulosa y les permita comer menos y
nutrirse más. Esto se debe a que los osos panda evolucionaron recientemente de
ancestros depredadores, y por ende heredan un sistema digestivo adaptado para
comer carne, y deberán esperar algunos miles o millones de años hasta que
evolucionen algún sistema de rumia de la celulosa vegetal (Dierenfeld, Hintz,
Robertson, Van Soest, & Oftedal, 1982; Xue et al., 2015; Zhu, Wu, Dai,
Zhang, & Wei, 2011). Aunque las bacterias
que los pandas poseen pueden en efecto degradar celulosa, estas son diferentes
de las bacterias degradadoras de celulosa de los herbívoros, y de hecho, estas
bacterias descienden de bacterias de carnívoros, además el punto primordial es
que el sistema digestivo del panda es corto, y le da poco tiempo a estas
bacterias para degradar la celulosa.
Regresando sistema esquelético, este es
uno de los pocos sistemas que se fosilizan con alta facilidad, y por ende es
básicamente la única fuente de información sobre la forma de los seres vivos de
periodos antiguos, y por ende de sus funciones (Dahdul et al., 2012). Los organismos como los osos panda que no tienen una clara
correspondencia entre forma y función tienen una vida geológica corta, hasta
que su linaje logra adaptarse, en consecuencia, los fósiles que se obtienen son
los de formas optimas a sus ambientes en los que si se encuentra una asociación
entre forma y función (Eldredge, 2014). Los dientes de los tiranosaurios tienen formas cónicas y son
robustos como puntillas de rieles de trenes, por lo que podemos asumir con un
nivel de confianza adecuado que los empleaban para penetrar pieles gruesas como
armaduras más que para desgarrar la carne (DePalma, Burnham, Martin,
Rothschild, & Larson, 2013), en contraste los dientes de los giganotosaurios si tienen forma
de cuchillo para desgarrar la carne (Mazzetta, Blanco, & Cisilino,
2013).

Figura 3.2. Izquierda el
cráneo del tiranosaurio y a la derecha el del giganotosaurio, a pesar de las
similitudes superficiales, sus dientes son un ejemplo de las diferencias de
comportamiento y dienta de estos antiguos vertebrados.
Sin embargo, es notable que las
adaptaciones por fantásticas que estas sean jamás presentan una forma
infinitamente diversa, por ejemplo, no existen vertebrados que se desplacen
sobre ruedas, o que vuelen por medio de un rotor como los helicópteros
(Kardong, 2011). La selección natural por sí sola no
sirve para explicar la razón de estas limitantes. Sin embargo, en la
descendencia con modificación, toda adaptación está limitada a la forma que
tenían los ancestros, lo cual crea sesgos no aleatorios sobre las posibles
soluciones que puede tener una ruta evolutiva en sus descendientes, lo cual da
lugar a las conocidas convergencias de forma o a evoluciones en paralelo en
diferentes continentes (Bailey, Rodrigue, & Kassen,
2015; Lenormand, Chevin, & Bataillon, 2016; Stern, 2013). En ocasiones la estructura previa genera muchísimos
inconvenientes a la nueva función, como la ya mencionada relación del sistema
digestivo del panda con su actual dieta.
Es por esto por lo que para poder entender
la forma de los seres vivos se hace necesario tener una vista evolutiva, la
cual nos permite poner en contexto los límites anatómicos de linajes anteriores
y como estos se modifican en los linajes más recientes. En este orden de ideas,
el sistema esquelético será la mayor fuente de información evolutiva y
funcional de los organismos en el pasado, lo cual nos clarifica el estado presente.
Esto es lo que hace que este capítulo sea particularmente largo, aun cuando se
ha hecho un esfuerzo por resumir. Muchos otros sistemas de órganos nos son
inaccesibles para linajes fósiles, pero el esqueleto siempre queda allí.
4. Materiales de construcción de un esqueleto
|| Regresar al Índice ||
Los esqueletos pueden ser duros o blandos.
Los esqueletos blandos emplean la presión del agua y los fluidos internos del
animal, pero lo esqueletos duros requieren de materiales rígidos, los cuales
inicialmente pueden ser proteínas. El problema es que estas proteínas pueden
tener consistencias semejantes al plástico, como en el caso del cartílago, para
endurecer aún más es necesario que el organismo introduzca otras sustancias, ya
sea de naturaleza orgánica o inorgánica para aumentar la dureza. Las
principales sustancias para crear esqueletos duros son
4.1 Carbonato
de calcio y otras sales
El carbonato de calcio, es un material
particularmente duro, el cual se asocia a las proteínas de la matriz
extracelular para generar un endurecimiento, es común en las conchas de los
animales marinos, en las espículas de las esponjas, en los exoesqueletos de los
arrecifes de coral entre otros. Acompañando al carbonato de calcio, pueden
aparecer otras sales derivadas del silicio o del magnesio (Brusca et al., 2003).
4.2 Quitina
La quitina es un polisacárido, es decir
una sustancia de naturaleza orgánica de los carbohidratos que posee una enorme
dureza, especialmente a la hora de fabricar conchas pequeñas como las de los
artrópodos (Brusca et al., 2003; Karp, 2013).
4.3 Esclerotina
La esclerotina es otra sustancia empleada
por los artrópodos, pero de naturaleza proteínica. En general las conchas de
los artrópodos es una aleación o sustancia compuesta de quitina, esclerotina,
matriz extracelular, sales inorgánicas e inclusive metales pesados como el
hierro y el vanadio. Esto da lugar a una amplia diversidad de durezas, y
también al agradable sonido cuando aplastamos a algún bicho molesto, ahora
recuerde, cada crack diferente al aplastar un bicho se debe a una composición
diferente de su exoesqueleto (Stevenson, 1969).
4.4 Lignina
Es una sustancia orgánica, muy compleja,
que conforma las partes duras de las plantas terrestres, siendo el principal
componente de lo que denominamos como la madera. Se trata de un grupo diverso de sustancias
basadas en uniones cruzadas de fenoles, históricamente ha sido difícil de
descomponer por parte de los microorganismos, por lo que la madera tiene una
resistencia relativa a la biodegradación, aun cuando esta no es perfecta. De
hecho la aparición de la biodegradación de la lignina conforma uno de los
capítulos más importantes de la historia de la vida macroscópica en el planeta
Tierra (Floudas et al., 2012; K. R. Stern,
Bidlack, & Jansky, 2008; Wayne, 2009).
4.5 Cartílago
Al igual que la quitina se trata de una
sustancia orgánica basada en proteínas que se segregan a la matriz extracelular
de las células, generando un endurecimiento, sin embargo la dureza no es tan
alta, generando un material con una textura semejante a un plástico poco denso
y flexible (Kardong, 2011).
4.6 Hidroxiapatita
La hidroxiapatita es un material compuesto
de sales como el carbonato y el fosfato que compone la parte dura de los huesos
de los animales vertebrados (Engin & Tas, 1999).
5. Esqueleto en plantas 1, generalidades
|| Regresar al Índice ||
Aunque al nivel celular las plantas se
describen en términos de un citoesqueleto y una pared celular, cuando se
aglomeran en sistemas multicelulares, sus células deben poseer algo a lo cual
anclarse para poder crecer. De hecho, las plantas más primitivas como los
musgos carecen de un sistema de anclaje y sostén verdaderos. En esta sección
discutiremos las estructuras de anclaje, sostén y protección de las plantas,
que corresponden a las raíces/rizoides y a los tallos. No todas las plantas
poseen un sistema de sostenimiento, de hecho, el desarrollo gradual de los
sistemas de sostén es uno de los rasgos a través de su proceso evolutivo. Por
ejemplo, las plantas durante el periodo Cámbrico de la Era Paleozóica no eran
más que algas verdes que podían existir como entidades unicelulares, o como
conglomerados multicelulares no especializados (De Duve & Pizano, 1995).

Figura 5.1. En las dos
imágenes anteriores tenemos al sistema vascular -circulatorio - de las plantas.
Este sistema en las plantas vasculares también da lugar al sistema esquelético
gracias al endurecimiento de la pared celular con un polímero amorfo que rodea
a la celulosa llamado lignina (Floudas et al., 2012).
En la actualidad existen plantas como el
musgo que presentan cierto nivel de especialización, pero que carecen de un
sistema de sostén y de un sistema circulatorio, por lo que están muy limitadas,
generalmente no crecen más de unos cuantos milímetros formando un tapete de
terciopelo (De Duve & Pizano, 1995). Para el periodo Devónico, las plantas
que son ancestros de todas las plantas terrestres tenían las mismas
limitaciones que el musgo, el paisaje estaría cubierto por un interminable
terciopelo verde, los individuos estarían apeñuscados de forma muy densa
compitiendo por capturar la mayor cantidad de luz solar, y por alcanzar la
mayor altitud posible para la liberación de las esporas reproductivas (Boyce et al., 2007).
La siguiente gran ramificación en las
plantas terrestres, las embriofitas como los helechos y parientes cercanos
poseen los rasgos que fueron desarrollados por sus ancestros en el Devónico
tardío, la primera es el sistema circulatorio, el floema y el xilema que
permite extraer materiales del suelo en contra de la gravedad, esto permitió a
las plantas crecer a niveles de arbustos, posteriormente la evolución de un
nuevo material cambiaría todas las reglas, la lignina, el material que forma la
madera y que permitió que las plantas despegaran rápidamente del suelo (De Duve
& Pizano, 1995). Para finales del devónico las plantas como los helechos
leñosos ya alcanzaban alturas de 8 o más metros (Dannenhoffer &
Bonamo, 1989; G J Retallack, Catt, & Chaloner, 1985; Stein, Mannolini,
& Hernick, 2007).
5.1 Raíces y
rizoides como sistema de anclaje de una planta
Una de las funciones de cualquier sistema esquelético es la de anclar al ser vivo a un sustrato, en los animales, este anclaje es importante ya que permite el movimiento, no puedes moverte fácilmente si no puedes ejercer una fuerza contra algún tipo de medio. Las plantas prosiguieron una ruta evolutiva diferente, para ellas lo importante era permanecer quietas en lugares donde la fotosíntesis fuera fácil, el problema es que permanecer anclado a un sustrato puede ser igual de difícil que moverse sobre o entre un sustrato. Un ejemplo de esto es lo que le sucede a un bosque después de un huracán, a menos que el suelo sea delgado, arenoso o suave, relativamente pocos arboles sanos habrán sido arrancados del suelo. Esto se debe a un sistema estructura que ANCLA el organismo al suelo de forma rígida, y esa es la raíz (Stern et al., 2008). Las raíces son un sistema apendicular que emerge del troco formando una compleja red que puede llegar a constituir hasta un tercio del peso seco de la planta. La raíz de la mayoría de las plantas no se extiende más de 3 o 5 metros bajo tierra; la mayoría de los pastos poseen raíces confinadas a menos de medio metro bajo la superficie (Stern et al., 2008).

Figura 5.2. El anclaje de la
raíz o el rizoide al suelo depende de múltiples factores como la profundidad y
extensión de la raíz y del tipo de suelo sobre el cual se ancla.
Las raíces son una estructura propia de
las embriofitas o plantas con un sistema vascular altamente especializado, por
lo que evolucionaron durante el devónico medio o tardío –junto con una serie de
hongos parasíticos o mutualistas como las micorrizas (Brundrett, 2002; Gregory J
Retallack, 1997; Taylor, Remy, Hass, & Kerp, 1995).
Las plantas más ancestrales poseen
rizoides, las cuales poseen la misma función de anclaje como en el caso del
musgo. La diferencia entre una raíz y un rizoide es la cantidad de tipos de
células que los componen, es decir de la cantidad de tejidos diferentes; un
rizoide está compuesto por un tipo único de tejido, mientras que una raíz es un
órgano compuesto por varios tipos de tejido. La función de anclaje de las
raíces y rizoides es enteramente macroscópica y depende del rozamiento entre el
suelo y la red de raíces/rizoides, por lo que no es necesario adentrarnos en la
biología de las raíces en este punto.

Figura 5.3. Corte transversal de una raíz de sidra, las raíces son estructuras complejas compuestas por diferentes tipos de tejidos (son un órgano).

Figura 5.4. Los rizoides
son estructuras simples compuestas por un solo tipo de célula (son un tejido).
5.2 Los
tallos de una planta como sistema de sostén
Las raíces pueden ser el sistema de
anclaje de una planta, pero quien se encarga realmente de sostener la planta y
anclar las partes vivas de esta es el tallo. El Tallo como hoy lo conocemos es
un rasgo propio de las plantas embriofitas y posee dos características
fundamentales que permiten a las hojas realizar sus funciones metabólicas a
metros de distancia de la fuente de agua más cercana: el sistema circulatorio
del xilema y el floema, y un mecanismo de sostenimiento del peso. En este
sentido podemos distinguir dos tipos de tallos, los tallos leñosos y los tallos
herbáceos.
Los tallos herbáceos, aunque poseen un
sistema circulatorio que les permite a sus dueños crecer a una altura superior
de unos cuantos milímetros, no permiten que la planta crezca mucho antes de
colapsar. De hecho, algunas plantas de jardín colapsan cuando florecen debido a
la ausencia de partes duras, otras colapsan cuando no se les aplica agua. La
turgencia o dureza de estas plantas depende de un tipo de esqueleto llamado
esqueleto hidrostático, es decir por la presión de agua en su interior (Moulia, Coutand, & Lenne, 2006).

Figura 5.5. Los tallos
herbáceos son verdes, blandos y flexibles. Son capaces de realizar la fotosíntesis. Los tallos leñosos son duros e incapaces de
realizar la fotosíntesis. En esta sección examinaremos la composición del tallo
leñoso.
Estos esqueletos hidrostáticos tienen
límites de peso, por eso algunos jardineros atan estas plantas herbáceas a un
palo que ancla las hojas o la flor; adicionalmente el esqueleto hidrostático
pierde su turgencia si la disponibilidad de agua disminuye (Moulia et al., 2006). Los tallos leñosos por el contrario han desarrollado un material
que endurece su estructura y le permite soportar un mayor peso, y elevar las
estructuras mucho más, al ser un material muy versátil debido a su relación de
peso y resistencia les permite a las plantas soportar pesos considerables hasta
alturas de más de 100 metros de altura, estamos hablando de la madera (McPARLAND et al., 2007)
5.3 Estructura
de un tallo
Una ramita consiste en un eje sobre el cual las hojas se encuentran dispuestas (Stern et al., 2008). Dependiendo de la posición de las hojas ha sido desarrollada una nomenclatura para designarlas, evidentemente esto se trata de una tradición de los botánicos del renacimiento y la antigüedad, y puede o no tener algún componente taxonómico. Si las hojas se encuentran en una disposición alternante o en espiral, se las clasifica como alternas/alternantes. Si las hojas están unidas en parejas, se las clasifica como opuestas.

Figura 5.6. anatomía ideal de un tallo.

Figura 5.7. (Izquierda)
hojas alternantes; (Derecha) hojas opuestas.
Si se encuentran en nidos de tres o más
hojas, se las clasifica como verticiladas. El área o región –más no la
estructura –donde se une una hoja con el tallo se denomina nodo, y la región de
un tallo entre dos nodos se denomina internodo. La estructura que une al tallo
con la hoja se denomina peciolo. El eje o Angulo entre un peciolo y un tallo se
llama axila, en cada axila, así como en la punta de los tallos se localizan
estructuras de crecimiento potencial llamados brotes, de las cuales puede crecer
más ramas o permitir la formación de flores. Cada brote se encuentra protegido
por escamas leñosas hasta que se encuentra listo para crecer en la estructura
para la cual esté dispuesto (Stern et al., 2008).
A medida que la madera leñosa y la corteza
reemplazan a un tallo herbáceo las estructuras respiratorias también cambian,
los estomas son reemplazados por poros más grandes, y a veces macroscópicos
llamados lenticelas con una función semejante. Las escamas leñosas del brote
terminal de la rama son más grandes y generalmente dejan cicatrices que
permiten conocer la edad de la planta. En otras ocasiones pueden encontrarse
cicatrices provocadas por estructuras asociadas al peciolo y las hojas llamadas
estípulas, por lo que es fácil confundirse.
5.4 La
lignina como material de sostén
La lignina es un polímero aromático muy complejo y amorfo que puede encontrarse en la pared celular de las plantas leñosas. La función de la lignina es la de ser un cemento o pegamento que une las fibras de celulosa y hemicelulosa, como si se tratara del concreto en un rascacielos. La lignina es uno de los compuestos biológicos de más difícil biodegradación –prácticamente se lo puede considerar como basura toxica producida naturalmente–siendo capaz de secuestrar el dióxido de carbono de la atmósfera sin dejarlo regresar cíclicamente. De hecho, el humus es en su mayor parte lignina no degradada. La lignina es el material rosado que rodea a los filamentos de celulosa y hemicelulosa en café. La estructura combinada funciona como el concreto y es la base molecular del sistema esquelético de las plantas leñosas.

Figura 5.8. La lignina es
un material altamente complejo y difícil de degradar, comparte muchas
propiedades con los plásticos, y, de hecho, durante la era carbonífera, la
madera generó un problema al plástico actual, casi nada podía biodegradarla.
Solo las bacterias y los hongos son
capaces de biodegradar la lignina –y solo después de mucho tiempo y en las
condiciones adecuadas. Ninguna enzima animal es capaz de degradarla, por lo que
animales como las termitas dependen de relaciones simbióticas para poder correr
la madera. Adicionalmente la capacidad de degradar lignina parece ser una
habilidad que tomó tiempo, mucho tiempo geológico para alcanzarse, ya que el
secuestro del dióxido de carbono es una de las causas más probables para la
extinción en masa que separa el periodo Carbonífero del periodo Pérmico de la
era Paleozoica (Floudas et al., 2012) ¿Qué le sucedió a la madera y a la lignina que jamás fueron
biodegradadas? Bueno el periodo Carbonífero recibe su nombre por algo ¿no?
Carbonífero significa el que porta carbón, y ya hemos mencionado que la lignina
secuestró el dióxido de carbono de la época, por lo que es fácil concluir que
el carbón secuestrado de la atmósfera del Carbonífero fue convertido a través
de la fosilización de la lignina en carbón mineral (Ruiz-Dueñas, Fernández-Fueyo,
Barrasa, Hammel, & Martínez, 2014).
5.5 La madera
y sus propiedades
El uso del xilema secundario –mejor
conocido como madera –por parte de los seres humanos como combustible, refugio,
armamento y propósitos varios data desde la antigüedad, de hecho, si le podemos
poner un sobrenombre es la del plástico de la naturaleza, es ligero,
resistente, y se le puede dar la forma que se desee en las manos de un buen
carpintero (Documental
sobre la Tala / Documental
sobre cambio climático y deforestación). La madera de un árbol vivo posee
una gran cantidad de humedad, la cual puede llegar a representar poco menos del
50% de su peso, por lo que para poder emplearla generalmente se la debe secar,
de forma tal que su contenido de humedad relativa baje a un 10% o menos. Esto
se logra ventilando la madera con aire seco en pilas almacenadas en bodegas
especializadas. El proceso de secado debe realizarse gradualmente y bajo una
supervisión estricta, pues de lo contrario la madera puede deformarse o se
pueden crear grietas no deseadas a lo largo de los radios haciéndola poco apta
para la mayoría de los usos ordinarios.
La parte seca de la madera está compuesta
por: glucosa (70 ± 5)%; lignina (20 ± 5)% junto con otros componentes
secundarios que afectan el color y la textura como resinas, gomas, aceites,
colorantes, taninos y almidón. Las proporciones de estos materiales afectan los
usos que va a tener un tipo determinado de madera. La función de los dos
componentes se asemeja al concreto, el cual posee barras de hierro que hacen
que el cemento a su alrededor sea más flexible; en este caso la celulosa actúa
como la fibra o barra, mientras que la lignina actúa como el cemento.
La densidad de la madera se encuentra
entre sus propiedades más importantes, y se define como la relación entre su
peso y su volumen. En botánica esta propiedad es denominada gravidez
específica. La madera que posee una gravidez semejante a la del agua, es decir,
que posee la misma densidad que el agua se la designa con el valor de 1.0. Si
la densidad de la madera es menor a la del agua si gravidez es menor a 1.0 y
viceversa. Esto posee efectos, específicamente contra el agua, pues si
construyeras un barco con madera más densa que el agua, este se hundiría a
menos que tuviera algunas modificaciones de diseño como los barcos de acero.
Por lo general la gran cantidad de
espacios vacíos al interior de la madera asegura que posean una gravidez
específica de entre 0.04-1.40. Por lo general la madera menos densa posee una
gravidez específica de 1.2. Las maderas con una gravidez menor o igual a 0.5 se
consideran ligeras, mientras que las maderas con una gravidez específica igual
o mayor a 0.7 se consideran pesadas.
Entre las maderas más pesadas, aquellas
con una gravidez específica de 1.25 o mayor, poseen propiedades semejantes a
los metales, por lo cual algunos submarinos silenciosos aún son fabricados con
ella. Los árboles que producen estas maderas superduras provienen de América
del Sur y de la India, como en los árboles de Carnipus apodados maderas de
hierro. La madera tiene la propiedad de soportar el decaimiento del tiempo
provocado por invertebrados, eso se denomina durabilidad.
Los únicos linajes de seres vivos que
pueden degradar la lignina, que es el cemento que mantiene unido a la madera,
son las bacterias y los hongos, y ambos requieren de altos niveles de
disponibilidad de agua para proliferar, a la disponibilidad de agua en la
atmosfera se la denomina humedad. La celulosa, que funciona como las barras de
hierro que le dan su flexibilidad a la madera, también necesita humedad para
poder ser degradada por las enzimas de los hongos y las bacterias. Los
componentes minoritarios de la madera, como los alcaloides, los taninos, los
aceites y las resinas también pueden presentar antibióticos y fungicidas
naturales que retrasan la colonización por parte de los hongos y las bacterias.
6. Esqueleto en plantas 2,Origen y desarrollo de los tallos
|| Regresar al Índice ||
Los tallos crecen gracias a que en su en su punta –región apical –se encuentra un tejido meristemático. Los tejidos meristemáticos se caracterizan por estar compuestos por células en constante mitosis y por ende en rápido crecimiento. Debido a que este meristemo se agrupa en la punta de los tallos y ramas se lo denomina meristemo apical.
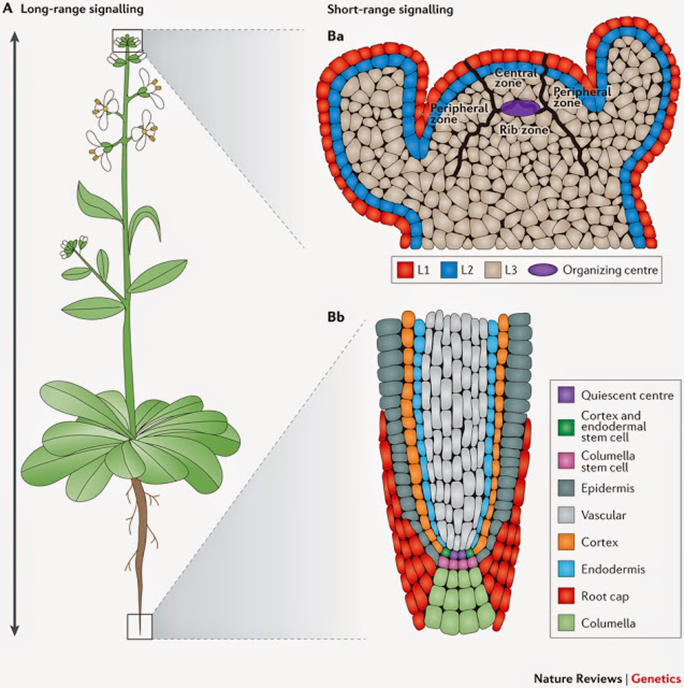
Figura 6.1. Modelo idealizado del meristemo apical.

Figura 6.2. Microfotografía
y modelo análogo del meristemo apical.
El meristemo apical se encuentra en
latencia antes de las estaciones de crecimiento –ya sea primavera y verano en
las regiones templadas, o las estaciones lluviosas en las regiones tropicales.
El brote se encuentra protegido por escamas leñosas, así como por los
primordios de los peciolos y las hojas. Cortar o dañar un brote significa que
la planta no podrá continuar con su crecimiento en dicha región, aunque no
siempre es el caso. En la siguiente imagen tenemos un corte longitudinal de la
punta de Coleus.
El meristemo apical del tallo embrionario de una semilla también se encuentra en latencia hasta que la semilla encuentra las condiciones de germinación. Cuando el brote comienza a expandirse o cuando la semilla germina, las células del meristemo apical empiezan una rápida mitosis y pronto tres meristemos primarios se desarrollan a partir de él. El más externo de los meristemos primarios se denomina protodermo y da lugar a la epidermis. A pesar de que existen plantas con una tendencia diferente, la epidermis es típicamente de una célula de espesor, y pronto empieza a segregar una sustancia dura y serosa llama cutícula que impide que el agua se evapore a través del tallo. Un cilindro de fibras que constituye el procambium aparece al interior de protodermo, a partir de estas fibras crecen los tejidos de conducción o sistema circulatorio de las embriofitas denominados floema y xilema.

Figura 6.3. En la
micrografía anterior tenemos el procambium, el cual es el tejido madre del cual
se diferencian el xilema y el floema primarios de un tallo/raíz joven.
El meristemo remanente denominado
meristemo de la capa base produce dos tejidos compuestos por células
parenquimatosas. Las células parenquimatosas son hasta cierto punto el
equivalente a las células madre en los animales, se trata de células poco
especializadas que rellenan muchos espacios y que bajo las condiciones
adecuadas pueden dar lugar a tejidos meristemáticos de crecimiento.
El tejido parenquimatoso en el centro de
un tallo se denomina Médula. El tejido parenquimatoso de un tallo que compone
la médula no es sólido, poco después de su desarrollo se rompe para dejar un
área hueca. Si no se rompe, con el tiempo es aplastado por nuevos tejidos
producidos por otros meristemos. El otro tejido producido por el meristemo de
la capa base es el córtex.
El córtex es una capa intermedia entre la epidermis y la médula, a menudo se encuentra salpicado por los ramilletes de tubos del sistema vascular. A pesar de que el córtex es más extenso que la médula, con el tiempo también es reemplazado por nuevos tejidos meristemáticos que crecen desde la médula, siendo aplastado hacía el centro del tallo. El parénquima de la médula y del córtex sirve no solo como tejido madre, también –y principalmente – como almacén de nutrientes y como tejido fotosintético –solo si está verde.

Figura 6.4. En la imagen
anterior tenemos un Corte transversal de un tallo joven. Todos los cinco
tejidos producidos por el complejo del meristemo apical: epidermis, xilema
primario, floema primario, médula y córtex; emergen cuando el tallo está
creciendo y se denominan tejidos primarios. Al mismo tiempo se forman los
brotes de las hojas, los cuales deben conectar sus primordios de floema y
xilema a sus contrapartes en el tallo en crecimiento.
6.1 Cambium
vascular y la madera
Ninguno de los tejidos embrionarios constituye la mayor parte del volumen de un tallo maduro. Justo en medio del floema y el xilema primarios se desarrolla una capa de tejido meristemático denominada cambium o más específicamente cambium vascular, aunque podría denominárselo más fácilmente como la capa vascular. Esta capa funciona como un tejido de células madre pluripotenciales que continuamente regenerar tejidos especializados y se regenera a sí misma.

Figura 6.5. El cambium
vascular es una capa de tejido que produce xilema hacia el interior y floema
hacia el exterior. El xilema se divide en funcional "que está en contacto
con el cambium" y xilema muerto. A las múltiples capas de xilema muerto le
llamamos madera o leña.
La capa vascular o cambium desarrolla una serie de tejidos como las traqueidas “células del xilema”, elementos de los vasos, fibras y otros componentes del xilema secundario, así como células filtro y otros componentes del floema. Sin embargo, la mayor parte del cambium vascular se dedica a la formación del xilema secundario, una serie de tubos que cumplen la misma función que el xilema primario “transportar agua y minerales de la raíz a las hojas. El xilema secundario se caracteriza por sintetizar un polímero complejo denominado lignina que se mezcla junto con la celulosa de la pared celular haciendo que los tejidos del xilema sean particularmente duros. Al conjunto del xilema secundario que ha sintetizado y saturado sus paredes celulares con lignina lo denominamos madera o leña. Su crecimiento se da hacia el interior del cambium vascular.

Figura 6.6. En el modelo
anterior podemos apreciar detalles de los tejidos que componen un tallo leñoso
de dos años. El xilema secundario funcional está cercano al cambium vascular,
mientras que el xilema secundario muerto o madera está cercano al xilema
primario.
El floema primario es posteriormente
separado del cambium vascular por el desarrollo del floema secundario, el cual
al igual que el floema primario tiene la función de transportar glucosa y otros
productos de la actividad celular desde las hojas a todos los tejidos vivos de
la planta, especialmente a las raíces. El floema secundario crece hacia el
exterior desde el cambium vascular.
6.2 Cambium
de corcho y la corteza
En muchas plantas, especialmente las especies leñosas, emerge una segunda capa de cubierta o cambium, este segundo cambium emerge de la epidermis o del floema primario. Recibe muchos nombres, la traducción literal desde el inglés es la de cambium de corcho, aunque en la literatura en español se lo puede encontrar como falógeno, cambium suberógeno o suberoso. El cambium suberoso está formado por células que segregan suberina, una sustancia aceitosa que hace al tronco impermeable. Esta capa suberosa constituye la corteza externa del árbol y protege las partes vivas de la intemperie y los parásitos. De hecho, tan pronto como la capa suberosa se forma, la epidermis primaria del tallo muere debido a que corta el flujo de agua y nutrientes desde el floema a la epidermis. De forma inversa, tiene el potencial de asfixiar los tejidos internos del tallo, pero al igual que las hojas, el tallo posee poros microscópicos denominados lenticelas que permiten en intercambio de gases de importancia metabólica.

Figura 6.7. Aunque
generalmente nos referimos a la corteza de un árbol como madera, su
constitución estructural es completamente diferente, la madera verdadera es el
xilema muerto de un árbol, la corteza es un producto segregado por el cambium
suberoso o de corcho y no está relacionado con el xilema.
6.3 Estela
El xilema primario, el floema primario y la médula, si están presentes, forman un cilindro central llamado estela, la cual tiende a estar presente solo en los especímenes más jóvenes. La estela es la forma desarrollada del procambium, en el cual el tejido del floema y el xilema primarios se han diferenciado, pero aun forman un anillo al rededor de la médula del tallo. La forma más simple de una estela se llama protoestela, consiste en un núcleo sólido de tejidos vasculares en los que el floema rodea al xilema. En este punto aún no ha crecido el cambium vascular, por lo que ambos tejidos vasculares primarios aún se encuentran en contacto. Las protéstelas en especímenes adultos son comunes en plantas productoras de semillas primitivas que se encuentran extintas en la actualidad, aunque también pueden encontrarse en algunos helechos y en los licopodios.

Figura 6.8. Las estelas
pueden organizarse de múltiples formas, pero en general el xilema
"rojo" se organiza al interior y el floema "azul" hacia el
exterior, esta tendencia se debe a que la mayoría de las plantas vasculares se organizan
como la eustela, con islotes vasculares formando un anillo.
La mayoría de las plantas productoras de semilla en la actualidad poseen euestelas, en los que el xilema y floema primarios se encuentran separados de forma discreta por haces vasculares que funcionan como muros de diferenciación; así como por todo el xilema secundario. Las plantas con flor poseen cierta diversidad en la forma en que presentan sus tallos, por lo que a continuación describiremos sus diferencias principales.
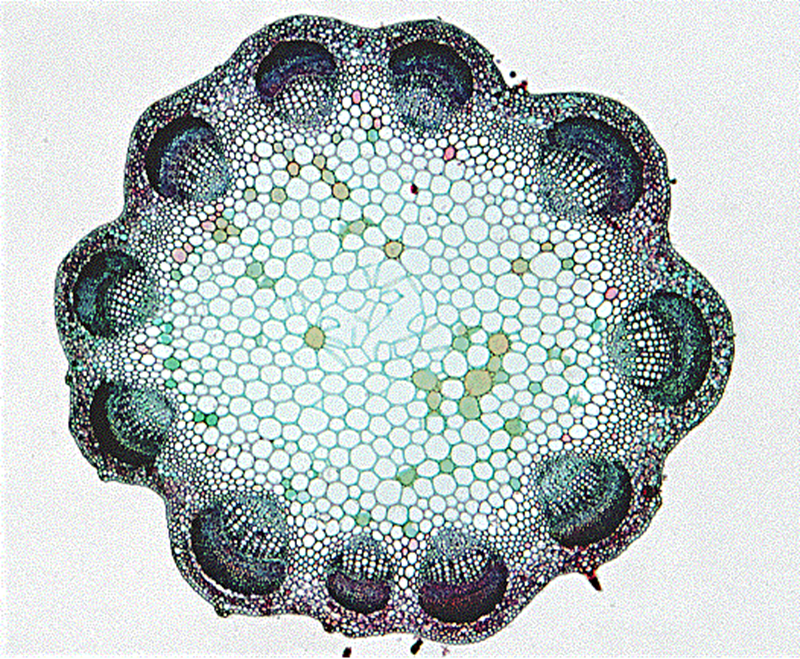
Figura 6.9. En la imagen
anterior podemos apreciar una micrografia de una eustela con los islotes
vasculares formando un anillo alrededor de la médula, en este caso no se ha
formado el cambium vascular ni el xilema o floema secundarios.
6.4 El xilema
de las gimnospermas
La nomenclatura empleada para describir la composición de la madera es un poco complicada si no tenemos claridad sobre las diferencias entre el xilema de las gimnospermas con respecto a las dicotiledóneas –recordemos que la madera es el xilema secundario muerto o vivo en un tallo y que la madera es la principal estructura de soporte y apoyo. Todos sabemos que el xilema transporta agua y minerales no procesados –denominados informalmente como sabia bruta. Lo que no se dice es que los componentes del xilema son dos entidades diferentes: las traqueidas y las traqueas.

Figura 6.10. En la imagen
anterior tenemos a la izquierda el xilema. El xilema puede estar compuesto por
la tráquea "en azul con flechas ascendentes" y por las traqueidas
"células en café alrededor de la tráquea". Las traqueidas son células
que forman las paredes de las tráqueas y en conjunto forman el xilema.
Una tráquea es un vaso o conducto hueco, mientras que una traqueida en un compartimento segmentado. En las angiospermas el xilema está compuesto por traquedas, que son tubos muy alargados que interactúan entre sí por medio de poros. Las traqueidas cumplen la misma función, pero estas no forman cilindros huecos, sino una serie de células más cortas que se interconectan entre sí para transportar la sabia bruta (Zimmermann, 1983). En las gimnospermas o plantas productoras de conos como los pinos, el xilema está compuesto mayoritariamente por traqueidas, mientras que en angiospermas como las dicotiledóneas el xilema está compuesto principalmente por tráqueas.

Figura 6.11. Modelo y
microfotografía del xilema
7. Esqueleto en plantas 3,Tallos como elemento de soporte
|| Regresar al Índice ||
Un tallo es uno de los dos ejes
estructurales principales de una planta vascular, el otro es la raíz. Sostiene
hojas, flores y frutos, transporta agua y sustancias disueltas entre las raíces
y los brotes en el xilema y el floema, almacena nutrientes y produce nuevo
tejido vivo. El tallo también se puede llamar halm o haulm o culmos. El tallo
normalmente se divide en nodos y entrenudos:
👉 Los
nodos sostienen una o más hojas, así como brotes que pueden convertirse en
ramas (con hojas, piñas de coníferas o flores). También se pueden producir
raíces adventicias a partir de los nudos. Las vides pueden producir zarcillos a
partir de los nudos.
👉 Los
entrenudos distancian un nodo de otro.
El término "brotes" a menudo se
confunde con "tallos"; "brotes" generalmente se refiere al
nuevo crecimiento de plantas frescas que incluye tanto tallos como otras
estructuras como hojas o flores. En la mayoría de las plantas, los tallos se
encuentran por encima de la superficie del suelo, pero algunas plantas tienen
tallos subterráneos. Los tallos tienen cuatro funciones principales que son:
👉 Soporte
y elevación de hojas, flores y frutos. Los tallos mantienen las hojas a la luz
y proporcionan un lugar para que la planta conserve sus flores y frutos.
👉 Transporte
de fluidos entre las raíces y los brotes en el xilema y el floema (ver más
abajo).
👉 Almacenamiento
de nutrientes.
👉 Producción
de nuevo tejido vivo. La vida normal de las células vegetales es de uno a tres años.
Los tallos tienen células llamadas meristemas que generan anualmente nuevo
tejido vivo.
Los tallos tienen dos tejidos en forma de
tubería llamados xilema y floema. El tejido del xilema transporta agua por la
acción de la transpiración, la acción capilar y la presión de la raíz. El
tejido del floema consta de tubos cribosos y sus células acompañantes. Los dos
tejidos están separados por cambium, un tejido que se divide para formar
células de xilema o floema.
7.1 Tallos de
dicotiledóneas herbáceas
Las dicotiledóneas herbáceas representan
todas aquellas dicotiledóneas que han perdido su capacidad para acumular
lignina en las paredes de las células que conforman el xilema secundario de los
tallos. Por lo general estas plantas no viven por mucho tiempo, teniendo ciclos
de vida anuales.
Los tejidos vasculares de estas
dicotiledóneas son principalmente primarios, esto se debe precisamente a que no
permanecen mucho tiempo vivas. Los tejidos vasculares secundarios se desarrollan
a medida que la planta crece y que el tronco envejece, pero una planta con un
periodo de vida limitada a un año o seis meses no desarrolla un cambium
vascular muy grueso.
En estas especies los tejidos vasculares
primarios tienden a asociarse en isletas discretas o parches que forman un
anillo alrededor de la médula del tallo que agrupa tanto al xilema como al
floema primarios. En algunas ocasiones un delgado cambium vascular puede
emerger, generando el xilema y el floema secundarios. En estos casos el xilema
y el floema secundarios no sobrepasan al xilema y el floema primarios,
simplemente se adicionan a los islotes vasculares de la planta, siendo en tal
caso un cambium vascular muy delgado.
7.2 Tallos de
dicotiledóneas leñosas
Las dicotiledóneas leñosas jóvenes inician su desarrollo de forma muy similar a una planta herbácea, con el xilema y el floema primarios y en contacto, formando un anillo en parches alrededor de la médula. Sin embargo, tan pronto como el cambium vascular y el cambium de corcho (corteza) empiezan a desarrollarse las diferencias aparecen a la vista. Las diferencias más importantes corresponden al xilema secundario o madera como es mejor conocido. A diferencia del xilema primario, el xilema secundario forma un cilindro continuo alrededor de la médula. Dependiendo del tipo de árbol y de sus condiciones, la apariencia del xilema secundario cambia.

Figura 7.1. En la imagen
anterior podemos apreciar una micrografia de una eustela dicotiledónea con los
islotes vasculares formando un anillo alrededor de la médula, en este caso no
se ha formado el cambium vascular ni el xilema o floema secundarios.
En los árboles de crecimiento continuo
como el ébano, el xilema secundario crece de forma relativamente continua
durante todo el año, lo cual le da una apariencia sin grano a su madera. Los
árboles que crecen en las regiones templadas deben afrontar las cuatro estaciones,
las cuales se dividen en estaciones de crecimiento –primavera y verano – y
estaciones de latencia –otoño e invierno.
Aun al interior de las estaciones de
crecimiento, el ritmo no es constante, el xilema secundario crece a un mayor
ritmo, sus vasos vasculares son más numerosos y amplios. Por el contrario, el
xilema secundario tiene dificultades para crecer durante el verano. Esto puede
deberse a que con una menor disponibilidad de agua en el verano, los tubos del
xilema deben hacerse más angostos para que la capilaridad sea mayor y en
consecuencia pueda elevarse suficiente agua a las hojas de la copa. Por tal
razón se genera una capa diferenciada de dos componentes: la madera de
primavera con tubos del xilema más amplios y la madera del verano con tubos del
xilema más angostos.
👉 Los anillos anuales: Un anillo anual consiste en una capa de xilema de vasos amplios y una capa de xilema de vasos angostos. Los intervalos entre uno y otro tipo de xilema es lo que crea un patrón de bandas a nivel macroscópico, esto se debe a que el grosor mancomunado de los vasos del xilema y sus demás componentes determinar su color relativo, por lo cual el xilema de verano y de primavera poseen colores diferentes. En las coníferas, la madera del xilema de los años anteriores pierde algunos componentes quedando solo las traqueidas. Los anillos anuales son todavía visibles debido a que al igual que sucede con los vasos, las traquidas de primavera son considerablemente más grandes que las de verano, lo cual altera la coloración de las capas del xilema producidas en una y otra estación. Tenga en cuenta que las dos capas de un anillo anual están compuestas por múltiples capas de células, de no ser así serian invisibles.

Figura 7.2. Los anillos
anuales permiten no solo contar los años, sino identificar cuales años fueron
más benignos que otros para el crecimiento del árbol.
El cambium vascular produce más xilema –en
su interior – que floema –en su exterior. El xilema tiene la tendencia a tener
paredes más rígidas, necesarias para una mayor capilaridad, esta es una de las
razones que pudieron impulsar la evolución de la lignina a mediados del
Devónico. Como resultado, la mayor parte del peso del tallo de un árbol
consiste en anillos anuales muertos –hacia el interior del cambium vascular en
crecimiento –siendo reemplazado capa por capa en cada una de las estaciones de
crecimiento. Una vez que la capa del xilema del verano pierde su actividad
–debido a los cambios en las condiciones del ambiente –su función es
reemplazada por el xilema de verano. Cuando pasa el periodo de latencia de
invierno y primavera, una nueva capa de primavera emerge reemplazando la
función del xilema del año anterior.
Si le ocurriera una lesión fuerte al árbol
que penetrara la corteza hasta el cambium en crecimiento, la cicatriz quedaría
registrada en la capa del año respectivo, mientras que el cambium crece sobre
la cicatriz y continua su crecimiento “en ocasiones alterando el patrón de
crecimiento del cambium vascular”. Adicionalmente, el grosor de la capa de
primavera y verano pueden indicar las condiciones ambientales de la respectiva
temporada, lo cual le permite al botánico realizar afirmaciones sobre el
pasado. A esta rama de la botánica se la conoce como dendrocronología.
👉 Dendrocronología: La dendrocronología (Video
de YouTube 1 / Video
de YouTube 2 / Video
de YouTube 3) es el estudio del tiempo por medio de la información
recolectada en los anillos anuales de los árboles. Por ejemplo, su la lluvia
durante un año particular es más alta, el anillo anual para ese año será más
grueso debido a que la lluvia favoreció su crecimiento. Las condiciones de un
ecosistema no están determinadas exclusivamente por los factores abióticos,
sino también por los factores bióticos. El forrajeo de los herbívoros también
puede afectar el grosor de los anillos anuales. Un ejemplo de esto es que
cuando hay una infestación de orugas particularmente álgida y estas devoran la
mayoría de las hojas del árbol, el anillo anual se hace más delgado debido a la
deficiencia en la fotosíntesis producto de tener pocas hojas en actividad.
Si un incendio quema por debajo de la corteza
del árbol, quedará la cicatriz para el anillo de crecimiento de este año en
particular, pero no quedará un hueco, el cambium vascular simplemente crecerá
sobre la cicatriz de forma que vuelve a generar un cilindro continuo para crear
el anillo del año próximo. Por lo anterior, simplemente se deben contar una a
una las capas de los anillos anuales hasta llegar a la cicatriz para determinar
el año en que el incendio tuvo lugar. No es necesario cortar el árbol para
determinar su edad. Los botánicos han desarrollado instrumentos que permiten
realizar cortes transversales cilíndricos de un árbol vivo. Se trata de un tubo
hueco que penetra desde la corteza hasta la médula del árbol o más, permitiendo
extraer un cilindro de madera. Este cilindro de madera es luego analizado.
El conteo de los anillos anuales ha
provocado la destrucción de mitos amados. Por ejemplo, el roble de Hooker, un
monumental árbol lo suficientemente grande como para dar sombra a 9000 personas
en un día de verano ha sido reverenciado por su gran antigüedad, la cual según
el mito y la placa puesta por la administración local tenía supuestamente 1000
años. En 1977 se demostró, por medio del conteo de los anillos anuales, que el
susodicho roble tenía menos de 300 años.

Figura 7.3. El roble de
Hooker
👉 La muerte del xilema y los tipos de madera: Como mencionamos anteriormente, a medida que el cambium vascular se engrosa, las capas o anillos anuales del xilema más internos mueren, lo cual implica que solo la capa más externa del xilema secundario realiza la función de transportar la sabia bruta, la pregunta en este punto es ¿Qué le sucede al xilema de las estaciones previas? A medida que el árbol envejece, los protoplastos del parénquima que se encuentra en la médula del árbol invade el lumen –cavidad hueca –del xilema, ya sea este tráquea o traqueida. En otras palabras, las células del tejido parenquimatoso invaden como un cáncer el xilema inactivo de las estaciones más viejas, y se convierte su volumen en un espacio de almacenamiento para metabolitos secundarios –productos del metabolismo que pueden tener funciones secundarias a la supervivencia o simplemente ser muy toxicas para dejarlas libres en la célula.

Figura 7.4. Algunos tipos
de madera (Video de YouTube).
Algunos de estos metabolitos secundarios
son gomas, resinas, taninos o alcaloides, así como pigmentos que provocan que
el xilema se haga más oscuro, en otras palabras, el xilema o madera vieja se
oscurece. A esta madera vieja se la denomina madera del corazón o duramen,
mientras que el xilema ubicado hacia el exterior del cambium vascular es más
ligero –porque aún sigue hueco –y se denomina albura/ madera blanca/ madera
joven. Diferenciar entre la albura y el duramen depende mucho de la especie con
la que se está trabajando.
Desde el punto de vista de la industria de
la madera, y especialmente de la carpintería, la albura es un factor a tener
muy en cuenta en cada una de las maderas, tanto es así que en cualquier informe
técnico sobre cualquier especie, lo primero que se refiere es al color del
duramen (parte central del tronco) y a la albura; hay algunas maderas que si su
albura fuera más resistente serían mucho más apreciadas, ya que la albura
normalmente no sirve para trabajarla o bien se pudre antes, incluso tiene un
color completamente diferente, que es casi siempre blanco.
La madera vieja o xilema viejo no realiza
la función de transporte de sabia bruta, y en consecuencia no se lo puede
considerar como parte del sistema circulatorio de la planta. Sin embargo, su
función es la de sostenimiento y anclaje, justo las funciones de un sistema
esquelético. La dureza de la madera no depende del xilema en sí, sino de
estructuras accesorias llamadas fibras, las gimnospermas como los pinos no
tienen fibras, por lo que sus maderas tienden a ser más blandas en comparación
con angiospermas como las dicotiledóneas leñosas, que al poseer fibras generan
maderas más duras.
👉 Resinas,
alcaloides y otros productos de los tallos leñosos: En muchas gimnospermas, los conductos que almacenan metabolitos
secundarios no solo residen en la madera vieja, sino que se encuentran
distribuidos en otros tejidos del tronco más externos. Estos canales con forma
de tubo dependiendo de la especie pueden ser lineales o ramificados. A pesar de
que los canales de resinas están asociados comúnmente a las gimnospermas,
algunas angiospermas también los presentan. Los arboles tropicales como el
olíbano y la mirra poseen conductos de resina cerca de la corteza, sus resinas
son conocidas respectivamente como el frankincienso y la mirra, conocidas desde
la antigüedad. Commiphora
myrrha (Amar, 2003) es la especie de
angiosperma que produce la mirra como una resina “imagen siguiente”.

Figura 7.5. Árboles de
mirra “Commiphora myrrha”, la mirra fue un material importante para el comercio del
mundo antiguo, siendo uno de los regalos dados por los Reyes Magos a Jesús.
A medida que el cambium vascular produce
nuevo xilema secundario constantemente en su interior, también se encuentra
produciendo floema secundario hacia su exterior. El término genérico de corcho
es aplicado a todos los tejidos por fuera del cambium vascular o corteza.
Algunos autores distinguen entre la corteza interna compuesta por el floema
primario y secundario; de la corteza externa que consiste en el cambium de
corcho que está en contacto con la intemperie. Al igual que sucede con el
xilema, el floema no funciona de forma permanente, esto se debe a que a medida
que el cambium vascular crece, este genera presión hacia afuera del árbol, lo
cual hace que el floema primario y secundario originales se aplasten. Cuando
esto sucede, la corteza interna inicia la producción de nuevo floema
secundario. Lo anterior provoca que la sabia procesada, que transporta gran
cantidad de azucares y otros metabolitos secundarios se encuentre muy próxima a
la corteza, por lo que solo se necesita una herramienta cortante para acceder a
esta rica fuente de nutrientes. Los nativos americanos se dieron cuenta de
esto, y emplearon por milenios la sabia del abeto de Douglas Pseudotsuga menziesii “imagen siguiente”
como una fuente de alimento en estaciones difíciles.

Figura 7.6. Abeto de
Douglas “Pseudotsuga menziesii”.
Debido a la facilidad con que se puede
acceder a su sangre nutritiva, es que los arboles poseen conductos de resinas
cerca de la superficie, cuando se produce una lesión, los conductos segregan
resinas en la herida, atrapando y matando a artrópodos y bacterias que intentan
infectar el floema o el xilema del tallo. La resina más común es el látex
vegetal, el cual está compuesto por gomas, proteínas azucares, aceites
alcaloides y otras sustancias con efectos biológicos. Algunas formas de látex
tienen un gran valor comercial (Video
de YouTube), pero entre ellas el caucho es el más importante de todos. Pero
otros tipos de látex sirven para producir gomas de mascar. Otros poseen efectos
neuronales en los animales como en las amapolas del opio de las cuales se
extrae la morfina y la heroína (Pareek, Srivastava, Maheshwari,
& Gupta, 2012). Otro tipo de látex con efecto neuronal es la
producida por el árbol del Sali, sause blanco o Salix alba que es un
tipo de sauce, su efecto analgésico permitió la síntesis de la droga más
exitosa de todos los tiempos, el humilde pero efectivo ácido acetil salicílico
“aspirina” (Harbourne, Marete, Jacquier, &
O’Riordan, 2009).
7.3 Tallos de
las monocotiledóneas
Las monocotiledóneas como los pastos y los lirios son plastas herbáceas, es decir, plantas en cuyos xilemas no se acumula lignina, por lo que son verdes y no adquieren grandes tamaños. Adicionalmente no poseen ni corteza y cambium vascular, por lo que sus islotes vasculares proceden de los tejidos embrionarios, en otras palabras no producen nuevo floema o xilema secundarios. Como en las dicotiledóneas herbáceas, la superficie en contacto con el ambiente es la epidermis, pero la principal diferencia es la ubicación de los islotes vasculares, estos se encuentran distribuidos sin formar el típico patrón de anillo de las dicotiledóneas. Los islotes vasculares en las monocotiledóneas se encuentran dispersos por el tallo.

Figura 7.7. Tejido vascular
de una monocotiledónea.
Cada islote está organizado de forma que
el xilema se encuentra orientado al eje del tallo, y el xilema se encuentra
orientado hacia la parte externa del tallo. Por lo general, los primeros islotes
vasculares en aparecer son aplastados a medida que la planta crece, por esta
razón tienden a desaparecer los que se encuentran en el centro del tallo. De
esta forma el patrón de distribución de islotes vasculares de floema y xilema
de una monocotiledónea como el maíz se basa en pocos hacia el centro y muchos
hacia el exterior, pero en un tallo delgado este patrón no es tan aparente.
8. Esqueleto en plantas 4,Tallos especializados
|| Regresar al Índice ||
Como cualquier estructura biológica, los
tallos pueden sufrir modificaciones secundarias especializándose en funciones
que ya realizaba de forma básica. Este proceso evolutivo se denomina
subfuncionalización, y hace parte de los procesos de especialización de
especies que evolucionan en ecosistemas estables y altamente competitivos. A
pesar de que la mayoría de las plantas poseen tallos erectos que cumplen las
funciones del sistema esquelético de sostener y anclar diversos órganos de las
plantas, muchas especies poseen tallos modificados para realizar diversas
funciones. Por lo general la especialización en una función determinada está
marcada por la adquisición de una forma diferente, sin embargo, sea cual sea la
modificación, los tallos pueden identificarse en base a la presencia de nodos,
internodos y brotes auxiliares. Estas características distinguen al tallo y al
tronco de otras estructuras como las hojas, los peciolos y las raíces. Las
hojas en los nodos de estos tallos y ramas especializadas tienden a ser
pequeños o con formas de escama, aunque raramente son verdes en algunas
ocasiones pueden presentarse en sus tamaños completos y con todas sus funciones
fotosintéticas. A continuación, describiremos algunas de estas funciones:
8.1 Rizomas
Los rizomas son tallos horizontales que crecen cerca de la superficie del suelo. Debido a esto se las puede confundir fácilmente con raíces, pero un examen detallado revela la presencia de hojas pequeñas en forma de escama y brotes auxiliares en cada nodo en al menos alguna etapa de su desarrollo.

Figura 8.1. Rizoma.
Raíces adventicias son producidas a todo
lo largo de un rizoma, especialmente cerca de la superficie. En botánica, la
palabra adventicia se refiere a órganos que crecen en lugares poco usuales,
como en este caso, una raíz creciendo a partir de una rama.
8.2 Estolones y tallos corredores
Son tallos horizontales que difieren de los rizomas en que crecen paralelos al suelo como en las fresas, y pueden ser lineales o ramificados.

Figura 8.2. Estolones.
8.3 Tubérculos
Son especializaciones de la punta de un estolón, cuando este madura se aísla del resto del estolón que le dio nacimiento. Se trata de almacenes nutrientes. Los tubérculos no se consideran raíces ya que poseen brotes, nodos e internodos igual que un tallo, y al igual que un tallo, sus brotes pueden germinar regenerando la planta.

Figura 8.3. Tubérculos
8.4 Bulbos
Se trata de brotes hipertrofiados cubiertos por muchas escamas y hojas carnosas con un tallo pequeño en la punta más baja.

Figura 8.4. Bulbos.
8.5 Cormos
Semejantes en apariencia a los bulbos, pero difieren de ellos al estar compuestos casi completamente de tejido del tallo.

Figura 8.5. Cormos.
No hay comentarios:
Publicar un comentario