
La célula // Ciencias de Joseleg // Biología

1. Introducción al estudio de la célula
La célula es la unidad fundamental de la
vida. De hecho, toda la vida está hecha de células. Sin embargo, las células
son extremadamente diversas en su forma y función. Nuestros propios cuerpos
están compuestos por varios cientos de tipos de células, y cada tipo está
presente en miles de millones de copias. Por ejemplo, hay células nerviosas
para conducir información, células musculares que permiten el movimiento,
células glandulares que secretan hormonas y células óseas para darles forma.
Como veremos, la estructura de cada uno de estos está especializada para
realizar su función particular.
Sin embargo, su estudio no vino de la
noche a la mañana, e incluso su propia existencia estuvo involucrada en arduos
debates científicos, así como en el foco de la era de los descubrimientos
gracias al desarrollo de nuevas tecnologías, especialmente en la rama de la
óptica y la electrónica. Sin embargo, hay que tener en cuenta que a pesar de
que la célula es la unidad fundamental que posee la propiedad de estar viva,
esta no siempre es el foco del estudio biológico.
2. Del vitalismo a la abiogénesis
La célula es la
unidad fundamental que posee la propiedad de estar viva, sin embargo, su
estudio no vino de la noche a la mañana, e incluso su propia existencia estuvo
involucrada en arduos debates científicos, así como en el foco de la era de los
descubrimientos gracias al desarrollo de nuevas tecnologías, especialmente en
la rama de la óptica. Sin embargo, hay que tener en cuenta que a pesar de que la
célula es la unidad fundamental que posee la propiedad de estar viva, esta no
siempre es el foco del estudio biológico.
Las ciencias de
la naturaleza se han ido construyendo mediante una serie de líneas de
investigación que han demostrado ser exitosas para el descubrimiento de muchos
fenómenos, por ejemplo, el fisicalismo newtoniano que ponía por encima de todas
las cosas, la posibilidad de deducir matemáticamente el comportamiento de los
fenómenos, y así expresar el funcionamiento del universo a través de ecuaciones
matemáticas simples. Otra línea de trabajo es el reduccionismo, tomar una
estructura y tratar de descubrir cómo funciona, mediante el estudio de sus
partes individuales. En años recientes, con el desarrollo de la computación ha
sido posible acceder al polo opuesto del reduccionismo, que es el holismo,
entender cómo interactúan las partes entre sí, generando propiedades que
emergen de las interacciones y que en consecuencia no aparecen al estudiarlas
por separado.
El estudio de la
célula es poco susceptible a la matematización, pero es posible plantear o
trasladar modelos físicos al ambiente celular, algo muy común en textos de
fisiología humana, y además muy útil, ya que eso ayuda a determinar
cuantitativamente el efecto de los fármacos en el cuerpo humano. El
reduccionismo por otro lado es lo que ha permitido entender cómo funcionan las
diferentes partes de una célula. En este sentido mucha de la información
consignada en los siguientes capítulos y secciones tendrá un enfoque
fuertemente reduccionista, ya que intenta meramente describir la anatomía y
fisiología de la célula individual y sus partes. Para tener un entendimiento
más holista es necesario abandonar el capítulo de la célula y adentrarnos ya
sea en la ecología o en la citología, donde ambas ramas estudian las
interacciones de la célula con sus ambientes circundantes.
2.1.
Vitalismo
y generación espontánea
Durante el
renacimiento europeo y el nuevo auge del estudio de las ciencias de la
naturaleza se desarrolló un debate antagónico que perduraría casi hasta finales
del siglo XIX, y fue la controversia entre el vitalismo y el fisicalismo, el
cual afectaría fuertemente el modo en que entendemos la existencia de las
células y demás entidades con vida, en oposición a los seres no vivos. En últimas,
se trata de dar una respuesta a la pregunta ¿Qué es un ser vivo?
👉 Reduccionismo vs
holismo: Mayr (1997) realiza una reconstrucción histórica del debe entre el fisicalismo
y el vitalismo en el siglo XIX. Se resalta como las afirmaciones fisicalistas
afirmaban que todos los seres vivos (menos el hombre) eran simplemente máquinas
que podían ser explicadas por las leyes de la física. Los fisicalistas se
oponían al uso de conceptos metafísicos no observables como la fuerza vital
para explicar los procesos de la naturaleza. En el texto Mayr resalta que lo
que los fisicalistas señalaban como el error en el vitalismo también lo hacían
ellos, ya que conceptos como gravedad o el calor también eran entidades no
observables. Mayr posteriormente comienza a discutir sobre el vitalismo, y como
al interior del contexto del siglo XIX se organiza como un modelo tan válido
como el de la gravedad, la energía o la afinidad para explicar aspectos de la
vida que no podía explicar el fisicalismo. El vitalismo según Mayr (1997) y otros autores (Gilbert & Sarkar, 2000; Henderson,
2012; Vlaardingerbroek, 2012) se
basa en dos ideas generales. La primera es que la vida es influida por una
especie de fuerza vital; y que esta solo se manifiesta en el organismo como un
todo. Como se había dicho anteriormente en el contexto del siglo XIX la
apelación a la fuerza vital, protoplasma o entelequia no es para nada
anticientífico ya que va en consonancia con otras fuerzas invisibles propuestas
precisamente por el fisicalismo.

Figura 2.1. Ernst Walter
Mayr (5 de julio de 1904, Kempten, Alemania – 3 de febrero de 2005, Bedford,
Massachusetts Estados Unidos), fue uno de los más notables biólogos evolutivos
del siglo XX. Fue, además un renombrado taxónomo, explorador de los trópicos,
ornitólogo, historiador de la ciencia y naturalista. Su obra contribuyó a la
revolución conceptual que llevó a la síntesis moderna de la teoría de la
evolución y al desarrollo del concepto biológico de especie.
En segunda instancia el postulado de que es necesario el todo del organismo para poder recibir esta fuerza sirve para hacer de la biología un campo de estudio independiente de la física o la química. En este último sentido la biología no quedaría relegada a emplear las leyes de la física o la química, sino que debería tomar conciencia de estudiar sus propias leyes. Es este segundo aspecto del vitalismo el que es rescatado por autores como Jacob (Jacob. F, 1973). Según Mayr el vitalismo decae bien entrado el siglo XX debido a la aceptación por parte de las comunidades científicas de la Selección Natural que permite explicar la adaptación en términos de un mecanismo y no en términos de una fuerza metafísica; así como de la genética que permite explicar la transmisión de la información desde los padres a los descendientes en un método definitivamente materialista (Blanco, 2012).


Figura 2.2. Georges
Canguilhem (Castelnaudary, 4 de junio de 1904 — Marly-le-Roi, 11 de septiembre
de 1995) fue un filósofo y médico francés, miembro del Collège de France,
especializado en epistemología e historia de la ciencia. Su obra se dirige
hacia las ciencias de la vida; es una de las más novedosas de la segunda mitad
del siglo XX, al conseguir desbordar, con sus intuiciones, ese campo estricto.
Se dice de él que fue el teórico de la normatividad, al considerar que las
normas visibles son siempre el producto de un poder regulador previo.
También se discute como a pesar de una
disminución de la aceptación del vitalismo por parte de los biólogos en el
siglo XX, fue acompañado de forma paralela por su aceptación en los círculos de
la física “lo cual es paradójico” con autores tan renombrados como Bohr o
Schrödinger (Hoyningen-Huene, 1991; Mayr, 1997;
Roll-Hansen, 2000).
Cabe destacar que el vitalismo no desapareció del todo de los autores en
biología, especialmente en el contexto francés donde cabe destacar a Georges
Canguilhem. En base a sus trabajos (Wolfe, 2007) realiza una extensa discusión sobre la concepción vitalista de
Canguilhem. A pesar de que las citas y la discusión sobre el vitalismo de
Canguilhem dejan entre ver que el concepto que el maneja es un poco diferente,
constantemente se ratifica que el vitalismo no es como el flogisto o el
geocentrismo, sería entonces que ¿no es una teoría científica susceptible de
ser falsada?
👉 El vitalismo moderno: Más allá de la discusión filosófica, lo que es importante
resaltar, es que el vitalismo de Canguilhem señala la necesidad de estudiar a
los seres vivos como todos integrados de forma heurística y no como partes
reducidas y aisladas del contexto. Esta afirmación va más en la línea de la
segunda idea del vitalismo planteada por Mayr (1997) en que la fuerza vital solo se manifiesta en la vida como un todo.
Sin embargo y lo que resulta más interesante para la concepción de vitalismo
que tienen numerosos autores es que no declara nada a cerca de fuerzas vitales,
entelequias o fluidos protoplasmáticos. De hecho, Gilbert y Sarkar (2000) definen
al vitalismo por esta vía doble, igual que Mayr: “En el vitalismo, la materia viviente es ontológicamente superior que la
suma de sus partes debido a alguna fuerza vital, que es adicionado o infundida
en las partes químicas”. Lo que se discute no es la parte en que la vida
sea más que la suma de sus partes, el problema recae sobre explicar esa
superioridad en base a un concepto como la Fuerza Vital. Las ciencias naturales
se construyen bajo el presupuesto de un materialismo metodológico (Blanco, 2012), por lo que la Fuerza Vital debe poderse traducir a algún tipo de
interacción material o propiedad emergente (Lazzerini-Ospri, Stano, Luisi, &
Marangoni, 2012; Morowitz, Srinivasan, & Smith, 2010). Mayr (1997) plantea que la resolución del debate entre el fisicalismo y el
vitalismo se logró mediante la adopción de un tercer modelo ecléctico, en el
cual se retoman consideraciones del fisicalismo y el vitalismo. En palabras de
Mayr tanto el vitalismo como el fisicalismo estaban parcialmente en lo correcto
y parcialmente equivocados.
El fisicalismo cometía el error de negar
que los seres vivos poseyeran propiedades que escapaban de las meras leyes de
la física o de la química, pero el vitalismo era problemático en buscar esas
leyes en entelequias abstractas o metafísicas. Se describe como problemático y
no como equivocado, el materialismo metodológico excluye toda fuerza metafísica
no porque se la conciba como una falsedad, sino porque no contribuyen a la
construcción de un proyecto de investigación ya sea en el campo o en el
laboratorio, es decir, aun cuando la Fuerza Vital sea una entidad metafísica
real, las ciencias de la naturaleza deben operar como si esta no existiera (Blanco, 2012), y aceptar esto tomó sus propias controversias en cada una de las
tres grandes ciencias de la naturaleza. El organicismo hace hincapié en el
concepto de propiedades emergentes y niveles de complejidad, conceptos que
gozan de gran popularidad al interior de las comunidades científicas
actualmente (Bergandi, 2011; Bersini, Stano, Luisi,
& Bedau, 2012; Damiano, 2012; Denton, Kumaramanickavel, & Legge, 2013;
Malaterre, 2013), y
que no se consideran metafísicas, la razón por la cual una propiedad desaparece
cuando reducimos el todo a sus partes, es porque al estudiar las partes por
separado no podemos ver el modo en que están actúan en conjunto, al acoplarse
se generan nuevas funciones y emerge una propiedad nueva, es decir leyes
netamente biológicas que no pueden reducirse a la física o a la química pero
igualmente naturalistas y matemáticamente describibles.
👉 La generación
espontánea: El fisicalismo newtoniano
encuentra muchas de sus raíces en el mecanicismo con autores tan influyentes
como el propio René Descartes “1596-1650”, el cual creó un fuerte debate sobre
eso que conformaba la estructura interna de los seres vivos. Por un lado los
fisicalistas-mecaninicistas que defienden la tesis de que los seres vivos se
comportan como máquinas muy complejas, pero como máquinas al fin y al cabo; y
por otro lado los vitalistas que defienden que la estructura interna de los
seres vivos dependen un algo más ininteligible al que se denominó, fuerza vital
entre otros muchos seudónimos que al final terminaron por englobarse en lo que
bien puede ser denomninado el primer paradigma de la pre-biología al cual
denominamos como generación espontánea. Lo llamamos pre-biología ya que la
superación o abandono de la generación espontánea fue uno de los pasos para
construir la biología tal cual la entendemos nosotros.
El punto para resaltar es que no todos los
defensores del vitalismo creían que la fuerza vital fuera ininteligible, de
hecho, la fuerza vital sería semejante a otras fuerzas “explicativas” empleadas
en otros campos del conocimiento, paradójicamente en la física, con su fuerza
de gravedad. En tal caso, la fuerza vital se podría estudiar dentro del marco
del naturalismo metodológico (Enlace)
del mismo modo que la fuerza de la gravedad, convirtiéndola en una explicación
naturalista del mundo natural. Aparte de las dificultades filosóficas también
se generaron controversias religiosas, por ejemplo para el filósofo inglés
Ralph Cudworth (Ruestow, 1984) “1617-1688” la sola idea de intentar explicar a los seres vivos y
su origen mediante mecanismos meramente materialistas ya fuera por la necesidad
o el movimiento fortuito de la materia no solo era irracional, también era una
idea peligrosamente atea.
La generación espontánea no solo estaba
apoyada por las ideas religiosas, sino también por la tradición popular y la
tradición académica. Para cualquier granjero era tangiblemente evidente que las
ratas emergían espontáneamente de las bodegas de grano, o que los gusanos
aparecían como de la nada a partir de un cuerpo en descomposición, esto revela
que desconocían mucho del ciclo de vida de los seres vivos. Un autor que
recopiló este modo de ver y lo plasmó de forma coherente y elegante fue
Aristóteles, quien se convertiría en la mayor autoridad en ciencias de la
naturaleza durante toda la edad media y parte del renacimiento. En este orden
de ideas
La generación espontánea es ciertamente el
primer método explicativo para el origen de la vida y su organización ecológica
y las ideas de Aristóteles al respecto serían el estándar sobre el asunto por
milenios. En su Historia Natural Aristoteles nos dice:
“Ahora existe una propiedad
de los animales que tienen en común con las plantas. Algunas plantas son
generadas por las semillas de otras plantas, mientras que otras se autogeneran
a través de la formación de algún principio elemental similar a una semilla; y
en estas segundas plantas algunas derivan sus nutrientes del suelo, mientras
que otras crecen dentro de otras plantas, como he mencionado por cierto en mi
tratado de Botánica. En los animales, algunos se originan de sus padres de
acuerdo a su clase, mientras que otros crece de forma espontánea y no desde su
misma clase; y en estos casos de generación espontánea algunos provienen de la
putrefacción del suelo o la materia vegetal, como es el caso de numerosos
insectos, mientras que otros son generados de forma espontánea al interior de
otros animales mediante las secreciones de varios órganos” (Aristotle, 2014a)
En otro texto, Aristóteles explica más a
cerca de este principio elemental:
“Los
animales y las plantas vienen a ser en lo sólido y lo líquido debido a que
existe agua en lo sólido y aire en lo líquido, y en todo aire existe el calor
vital en el sentido de que todas las cosas poseen un alma. Por lo tanto, los
seres vivos se forman rápidamente siempre que el aire y el calor vital se
encuentran encerrados en algo. Cuando están encerrados, los líquidos corpóreos
son calentados, y allí emergen como si fuera una burbuja” (Aristotle, 2014b)
Dado lo anterior el núcleo fuerte de la
generación espontánea es: la capacidad
que tienen todos los seres vivos de formarse en cualquier momento y cualquier
material que se encuentra en contacto con el aire que contiene la fuerza vital. Otra de las consecuencias de la
generación espontánea es que opera de forma independiente al tiempo, es decir la
generación espontánea opera de forma semejante seas que retrocedemos en el eje
del tiempo o se avanza en el eje del tiempo, en consecuencia, si la refutas
para el presente, esta se cae tanto en el pasado como en el futuro, ya que
debía funcionar en cualquier tiempo.
2.2.
Controversias por la
generación espontánea
👉 Redi y la biología
experimental: La muerte de la
generación espontánea fue lenta, y esto se debe a sus hipótesis auxiliares, es
decir explicaciones ad hoc empleadas
para proteger el núcleo fuerte. Un ejemplo clásico es el experimento realizado
por Francisco Redi “1626-1697” en el cual se empleaban gazas para evitar que
los insectos llegase a la carne en putrefacción Figura 2.3.

Figura 2.3. El experimento de Francisco Redi.
Colocó un trozo de carne en tres jarras
iguales, la 1º la dejó abierta, la 2º la tapó con un corcho, y la 3º la dejó
cubierta con un trozo de tela bien atada. Después de unas semanas Redi observó
que, en la 1º jarra, la que estaba abierta, habían crecido larvas. En la 2º
jarra y en la 3°, su interior estaba podrido y olía mal, pero no había crecido
ninguna larva. Por lo tanto, la carne de los animales muertos no puede
engendrar gusanos a menos que sean depositados en ella huevos de animales. Redi
pensó que la entrada de aire a los frascos cerrados podría haber influido en su
experimento, por lo que llevó a cabo otro. Puso carne y pescado en un frasco
cubierto con gasa o con un mosquitero; después de tiempo Redi se fijó y
descubrió que las moscas o gusanos dejaban sus huevos no en el frasco sino en
la gasa. Por eso la gente que creía en la generación espontánea; creían que
gracias a eso generaban vida. Los resultados fueron exactamente los mismos que
en el primer experimento. Aún con los resultados obtenidos y los de otros
autores, la gente seguía creyendo en la generación espontánea, y Francesco Redi
se vio obligado a admitir que en ciertas ocasiones sí se podía dar la
generación espontánea. Su obra más importante, donde expuso los resultados de
sus experiencias, la escribió en el año 1684. Suya es la frase Omne vivum ex ovum, ex vivo que se
traduce como todo lo vivo procede de un huevo y este de lo vivo (Gottdenker, 1979; Parke, 2014).
Este experimento en particular tenía como
objetivo demostrar la invalidez de la generación espontánea de los insectos, no
de la generación espontánea en general, pues el mismo Redi creía en la
generación espontánea en ciertos casos (Strick, 2009b). En este caso, una hipótesis ad
hoc que puede plantearse al experimento de Redi es que, si bien la gaza
permite la entrada del aire, podría potencialmente impedir el ingreso de la
Fuerza Vital. De este modo se desarrollaron hipótesis auxiliares que defendían
las ideas centrales de la generación espontánea, por ejemplo: La fuerza vital requiere de un espacio lo
suficientemente amplio para ingresar al material en putrefacción.
👉 Needham vs
Spallanzani: Una segunda hipótesis
auxiliar de la generación espontánea proviene de la controversia de defensa y
ataque a la generación espontánea. Desde la perspectiva de la defensa de la
generación espontánea se encuentran John Turberville Needham “1713-1781”, y
Georges Louis Leclerc conde de Buffon “1707-1788”; mientras que desde el ataque
se tiene a Charles Bonnet “1720-1793” y principalmente a Lazzaro Spallanzani
“1729-1799”. La controversia giro nuevamente en torno al diseño experimental (Strick, 2009b), en el caso de Needham el diseño experimental buscada esterilizar
el interior del material de cultivo mediante una ebullición cuidadosa, y
aislarlo del ambiente externo mediante el empleo de una sustancia cerosa. Por
su parte Spallanzani fue mucho más allá, aislando el sistema no con cera, sino
con el vidrio de la capsula de cultivo mediante calentamiento y realizando una
ebullición más vigorosa por más de una hora. Mientras que en el caso de Needham
se encontró crecimiento de protistos y el reporte de animáculos moviéndose, en
el diseño de Spallanzani no hubo crecimiento.

Figura 2.4. En el experimento de Needham se empleó corcho como barrera, sin embargo,
este material tiene poros pequeños que no aseguran un correcto aislamiento.
Nuevamente el núcleo firme de la
generación espontánea fue salvado por una nueva hipótesis ad hoc, en este caso propuesta por Needham:

Figura 2.5. En el
experimento de Spallanzani se emplea al vidrio como barrera, pero los
vitalistas argumentaron que el vidrio impedía el contacto con la fuerza vital.
La fuerza vital
es muy sensible a un proceso prolongado de ebullición. Adicionalmente a estas dos hipótesis auxiliares, la generación
espontánea poseía dos subhipótesis que pueden ser distinguidas (Farley, 1972) y son el concepto de abiogénesis y heterogénesis. La abiogénesis
hace referencia a la capacidad de las sustancias
inorgánicas de formar seres vivos; mientras que la heterogénesis
hace referencia a la generación
espontánea de un tipo de seres vivos a partir de otros seres vivos.
La segunda hipótesis le permite a la generación espontánea explicar el fenómeno
de los parásitos en una época en que el concepto de un ciclo de vida complejo
era desconocido para los historiadores naturales. Como se puede ver en el
trabajo de Aristóteles (Aristotle, 2014a, 2014b), ambas sub-hipótesis se encuentran ya contenidas en su propuesta
original.
👉 Luis Pasteur y los
cuellos de cisne: La situación seria
retomada por Luis Pasteur empleando el experimento de los frascos de cuello de
cisne. En su famoso experimento, Louis Pasteur usó un matraz especial cuyo
cuello tenía la forma de una S o el cuello de un cisne, de ahí el nombre
"Matraz de cuello de cisne". Puso un caldo rico en nutrientes en el
matraz, que llamó la "infusión”. Luego hirvió la infusión matando
cualquier microorganismo que ya estuviera presente. Luego permitió que la
infusión reposara (Nakazawa, 2018; Roll-Hansen, 1979).
Debido a la forma del matraz, la infusión se expuso al aire. Sin embargo, las partículas de polvo y otras cosas en el aire nunca llegaron a la infusión. Porque estaban atrapados en la curva del matraz de cuello de cisne. No importa cuánto tiempo dejó reposar el matraz, los microorganismos nunca aparecieron en la infusión. Sin embargo, si inclinaba el matraz y permitía que las cosas atrapadas en el cuello entraran en la infusión, entonces los microorgranismos comenzaron a aparecer en la infusión y se multiplicaron rápidamente. Esto demuestra que los microorganismos no aparecen como resultado de la generación espontánea. En cambio, se introducen en los alimentos a través de partículas de polvo y otras cosas que suceden en el aire y caen en los alimentos (Nakazawa, 2018; Roll-Hansen, 1979).

Figura 2.6. El experimento
de Pasteur dejaba un orificio por donde la fuerza vital debería penetrar, pero
no lo hacía. El problema radicó en que otros repicaron el experimento con
resultados diferentes.
A pesar de que la tradición dice que Luis
Pasteur “1822-1895” refutó contundentemente a la generación espontánea mediante
su trabajo con los frascos de cuello de cisne en 1864 (Cloutier, 1995), la historia es un poco más complicada que eso. La generación
espontánea era vista como una hipótesis científica aceptable por miembros
eminentes de la comunidad científica de la época (Benton, 1974). En cualquier caso, el experimento de Pasteur permitió eliminar
varias de las hipótesis auxiliares, así como la mitad del núcleo fuerte.
Para el caso de la hipótesis auxiliar (1)
Pasteur no utilizó gazas para bloquear el frasco, sino que lo dejó abierto con
una modificación, el cuello de la botella tenía forma de un cuello de cisne. El
frasco solo experimentó crecimiento si se rompía el cuello de cisne dejando un
cuello recto, o girando el frasco de forma tal que el líquido de cultivo
tuviera contacto con la parte más baja del cuello de cisne. De esta manera
Pasteur evitaba las afirmaciones sobre el “tamaño de la fuerza vital”. El mismo
procedimiento también evitaba la hipótesis auxiliar (2), aun cuando el caldo de
cultivo era vigorosamente esterilizado, aun permitía el crecimiento de
microorganismos cuando el cuello giraba o era roto. La estructura del cuello
del cisne permitía que el aire ingresara al frasco, pero que el material
particulado se terminara decantado en la parte inferior del cuello de cisne, lo
cual invalidaba el principio aristotélico de una fuerza vital transportada por
el aire.
👉 La biogénesis: Sin embargo no todos estuvieron satisfechos con esto, de hecho el
experimento de Pasteur fue seguido por otros trabajos como los de John Tyndall
“1820-1893” (Strick, 2009b) quien tuvo problemas con la esterilización vigorosa. Esto se debía
a la existencia de las esporas termoestables “desconocidas para él”, pero que
fueron del mismo modo una hipótesis auxiliar que el propuso para proteger el
núcleo fuerte de la propuesta de Pasteur de la biogénesis “toda vida proviene de una vida previa”. Por suerte para el programa
de investigación de la biogénesis, la hipótesis auxiliar de las formas resistentes al calor fue
corroborada en años posteriores por Ferdinand Cohn “1828-1898” (Drews, 2000) con su descripción de las esporas en 1876 -más de diez años después de los experimentos
de Pasteur.
3. El fin del vitalismo
En la sección anterior se ha visto como el
concepto de abiogénesis hace parte del programa de investigación de la
generación espontánea siendo una hipótesis auxiliar, pero no como parte del
núcleo fuerte, es decir aun cuando se refutó el concepto de abiogénesis, la
generación espontánea hubiera continuado mediante la hipótesis de heterogénesis
–hasta que fueran descritos los ciclos de vida complejos de los parásitos –lo
contrario también es válido. Sin embargo, es importante recalcar que el núcleo
fuerte de la generación espontánea hace referencia a dos ideas importantes, la
primera la de un tiempo reversible en la que la generación espontánea es la
misma en el pasado, presente y futuro; y la segunda el contacto con el aire o
fuerza vital.
3.1.
El darwinismo y el origen de
la vida
El pasado es irreversiblemente diferente
del presente y del futuro, el problema del tiempo irreversible es introducido
de forma primordial por Charles Darwin “1809-1882”, aunque las ideas evolutivas
ya estaban presentes en otros autores entre los que cabe destacar a
Jean-Baptiste-Pierre-Antoine de Monet caballero de Lamarck “1744-1829”. Tanto
la teoría de la evolución como la generación espontánea parecían estrechamente
relacionadas en el sentido de que ambas intentaban explicar al mundo natural en
base a las leyes de la naturaleza “naturalismo metodológico”. Sin embargo
Darwin era bastante perspicaz y cauteloso en ese sentido, por esa razón evadió
de forma magistral en su obra máxima El
Origen de las Especies (C. Darwin, 1859) a la generación espontánea o cualquier acercamiento al problema
del Origen de la Vida.
👉 Los darwinistas: Fueron otros autores quienes vincularon inicialmente las ideas
darwinianas la generación espontánea, entre ellos cabe destacar a uno de sus
más grandes seguidores, el biólogo alemán Ernst Haeckel “1834-1919”. A pesar de
que la evasión de Darwin sobre el Origen de la Vida puede verse como totalmente
intencional, Haeckel la percibía como una debilidad “El mayor defecto de la teoría darwiniana es que no da luces sobre el
origen del organismo primitivo –probablemente una célula simple –de la cual
todos los seres vivos descienden. En el momento en que Darwin asume un acto
creativo especial para el primer ser vivo, pierde consistencia” (Haeckel, 1862; Rieppel, 2011). La hipótesis de Darwin permitía una explicación materialista
aplicable a una metodología de investigación, por lo que si se asumía que el
Origen de la Vida tenía un origen no materialista (Blanco, 2012), su estructura de pensamiento no sería coherente “por lo menos en
lo metodológico” y eso era de lo que se quejaba Haekel.
Tal vez la referencia más directa que
tenemos del pensamiento de Charles Darwin sobre el Origen de la Vida no sea un
documento académico públicamente difundido y formalmente publicado, sino por el
contrario, una carta que escribió a su buen amigo Joseph Dalton Hooker
“1817-1911”. La hipótesis o serie de hipótesis expuestas en dicha carta han
venido a conocerse en su conjunto como la “Pequeña charca caliente” de Darwin
escrita en 1871, por decir casi una década después de que la teoría de la
generación espontánea fuera parcialmente refutada de Pasteur (Peretó, Bada, & Lazcano, 2009;
Spaargaren, 1985; Strick, 2009a) y al menos unos 5 años antes de la descripción de las esporas
termoestables, por lo que varios académicos aún seguían encontrando y
reportando crecimiento de microorganismos en caldos de cultivo vigorosamente
calentados. La carta así como otros documentos de Darwin fueron recolectados en
un solo texto por su hijo (Darwin, 1887) y su texto original completo es el siguiente:
👉 La opinión de Darwin
sobre el origen de la vida: “Será un
descubrimiento curioso si la observación del señor Lowe de que la ebullición no
mata ciertos mohos se demuestra; pero ¿cómo se puede explicar la ausencia de
todos los seres vivos en los experimentos de Pasteur? Siempre estoy encantado
de ver una palabra a favor de Pangenesis, que algún día, creo, tendrá una
resurrección…
… A menudo se dice que todas las
condiciones para la primera producción de un organismo vivo están presentes
ahora, igual que en todo momento pasado. Pero si (y es un enorme sí) pudiéramos
concebir en un pequeño estanque cálido con todo tipo de amoníaco y sales
fosfóricas, luz, calor, electricidad, etc. presente, que un compuesto de
proteína se formó químicamente, listo para sufrir cambios aún más complejos, en
la actualidad tal materia se devoraría o absorbería instantáneamente por la
vida ya presente, lo que no habría sido el caso antes de que se formaran las
criaturas vivientes.”(Darwin, 1887)

Figura 3.1. Darwin plantea
una idea simple pero potente, las condiciones para el origen de la vida no
están dadas en la actualidad, pues la vida se comería a la previda.
La carta resalta dos aspectos importantes
de la discusión sobre el origen de la vida y la generación espontánea, la
primera “contrario a la tradición”, los experimentos de Pasteur no eliminaron
por completo las hipótesis sobre abiogénesis, de hecho Darwin en el primer
párrafo de la carta se realiza una pregunta metodológica sobre el cómo Pasteur
determina la ausencia de desarrollo de seres vivos en los frascos de cuello de
cisne, el problema es que si existen seres vivos que son capaces de sobrevivir
a la ebullición, y estos causaban que no todos pudieran replicar los datos de
Pasteur. La aseveración de Darwin era ciertamente especulativa, dados los
diferentes datos en conflicto que iban y venían, aunque actualmente se sabe que
en efecto existen bacterias capaces de sobrevivir por encima de la temperatura
de ebullición (Takai et al., 2008). En cualquier caso, las dudas de Darwin sobre la refutación
definitiva de la generación espontánea eran compartidas por otros biólogos
ilustres como August Weismann “1834-1914”, Emil du Bois-Reymond “1818-1896”,
Karl von Nageli “1817-1891” y Hermann von Helmholtz “1821-1894” (Lazcano, 2010).
El segundo párrafo es el más famoso, y
representa uno de los primeros indicios no solo de la aproximación del
Origen de la Vida desde una perspectiva
química, sino también desde el punto de vista del tiempo irreversible (Benfey, 2007) y metodológicamente materialista (Blanco, 2012). Una vez que la vida surge, esta devora la previda a su alrededor,
pues la previda está
compuesta por materiales que sirven a la vida para crecer y reproducirse. En tal caso, toda práctica de
laboratorio que intente crear vida o condiciones antes del surgimiento de la
vida “prebióticas” debe asegurar que NO existan seres vivos que
contaminen/devoren los sistemas químicos en desarrollo.
👉 El tiempo
irreversible: Esta conclusión elimina
la otra mitad del núcleo fuerte de la generación espontánea que hace referencia
al tiempo reversible en el cual paso, presente y
futuro eran todos iguales; la
nueva hipótesis se basaba en un tiempo
irreversible, en donde las condiciones del pasado eran diferentes a las del
presente, y en consecuencia estas deberían ser deducidas y simuladas
experimentalmente. En base a esto el experimento de Pasteur refutaba la
generación espontánea en el presente, y en consecuencia lo refutaría en todos
los tiempos. Sin embargo, en una dimensión de tiempo
irreversible, donde se asumen condiciones diferentes para pasado,
presente y futuro, el hecho de que los experimentos de Pasteur y seguidores
refuten el presente no afectarían en nada al pasado (geológico) o al futuro
(diseños experimentales).
Sin duda la generación espontánea parte de
las ideas vitalistas, en la cual las estructuras biológicas están imbuidas con
algo más que sus constituyentes físico-mecánicos, pero se podían plantear
posturas vitalistas sin que se aceptara necesariamente la idea de la generación
espontánea, y Pasteur es un ejemplo de ello. Adicionalmente, como comenta Lazcano (2010), las ideas de la generación espontánea en Francia aun desde los
tiempos de “Buffon 1707-1708” habían sido asociadas con actitudes ideológicamente
polarizadas hacia filosofías materialistas de nivel ontológico y hacia un
secularismo radical (Farley, 1974; Henry, 1999). Es particularmente
evidente que las teorías darwinianas de la época eran materialistas –aunque
cada autor podría concebirla a nivel ontológico o meramente metodológico –el
problema es que, igual que sucedió siglos antes con la generación espontánea,
el mero hecho de ser materialista la convertía en una idea peligrosa, dentro de
un contexto fuertemente religioso, pues eran fácilmente tildadas de ideas
ateas, algo aun peligroso para la época.
👉 El antidarwinismo
religioso y el origen de la vida: Esta oposición
radical al pensamiento darwiniano era común dentro de la mayor parte de la
academia francesa (Farley, 1974), pues por ejemplo, uno de los más grandes opositores a las ideas
de Darwin también fue un francés, Louis Agassiz's “1807-1873”, quién realizó
sus críticas cerca de 1860 (Morris, 1997). En la perspectiva de la academia francesa, la generación
espontánea y el darwinismo eran tan materialistamente conectados, que
aparentaban ser uno, por lo que si se lograba refutar uno se eliminaba el otro (Farley, 1974), aun cuando para Darwin, Huxley y sus seguidores los problemas del
origen y el cambio de los seres vivos eran independientes entre sí (Strick, 2009a).
Farley (1974) menciona que esta oposición se debía a que la academia francesa
aún se encontraba dominada por viejas ideas románticas basadas en la filosofía
natural, en las cuales aún no se había generado una separación clara en el
método por el cual se desarrollaría la ciencia, es decir, la biología aún era
pensada como un mecanismo para comprobar la existencia de Dios “idealismo
metodológico”. Por el contrario, la academia de las naciones alemanas había ya
delimitado su campo de acción hacia una perspectiva materialista ya fuese
metodológica u ontológica. Este materialismo se basaba principalmente en la
escuela fisicalista del siglo XIX que se oponía al vitalismo y veía a los seres
vivos como máquinas creadas por sus componentes físicos.
3.2.
El vitalismo se arrinconó en
la química

Figura 3.2. La urea se
consideraba una sustancia que requería la fuerza vital al interior de los seres
vivos, sintetizarla de materiales no vivos en un frasco de vidrio fue un
descubrimiento revolucionario por parte de Wohler.
El problema de la estructura interna de
los seres vivos, fuera esta superveniente a sus componentes físicos, o basada
en alguna fuerza vital no sería resuelta en el siglo XIX. De hecho, el
vitalismo era verdaderamente arraigado dentro de la opinión de aquellos que
estudiaban los componentes de los seres vivos, es decir los químicos. Jöns
Jacob Berzelius “1779-1848” consideraba que las leyes de la química de los
seres vivos “química orgánica” era diferentes de los de la química inorgánica,
de forma tal que una síntesis inorgánica no podría producir moléculas orgánicas
(Miller & Lazcano, 2002).
Posteriormente las síntesis orgánicas
mediante sustancias inorgánicas darían su aparición. Estas síntesis fueron
exitosas en el sentido de lograr generar biomoléculas de importancia a partir
de moléculas inorgánicas simples, sin embargo, difieren en las síntesis
prebióticas en el siglo XX en un aspecto fundamental. Su diseño experimental no
tenía como objetivo tratar de reproducir condiciones abióticas-prebióticas, en
otras palabras, no se basaban en modelos geoquímicos de cómo debía ser la
Tierra primitiva (Benfey, 2007) por el contrario, trataban de encontrar los mecanismos por los
cuales las plantas actuales podían fijar ciertos componentes inorgánicos (Lazcano, 2010), es decir trabajaban en una escala de tiempo
reversible (Benfey, 2007). En cualquier caso, en esta época hay una reacción que se ha
vuelto relativamente celebre al interior de la comunidad de químicos
prebióticos y es la reacción de Formosa desarrollada por Aleksandr Butlerov
“1828-1886” en 1861 (Butlerow, 1861). La reacción de Formosa fue celebre inicialmente por ser una
síntesis inorgánica de carbohidratos, sin embargo, aún sigue llamando la
atención debido a sus mecanismos de reacción. Esto es debido a que el proceso
de la reacción de Formosa incluye la autocatálisis (Parmon, 2008; Parmon & Snytnikov,
2002), una de las propiedades
clave para los sistemas prebióticos (Hordijk, Hein, & Steel, 2010).
4. Del microscopio a la teoría celular
4.1.
Microscopios ópticos
Para poder descubrir la célula era
necesario un avance radical en la tecnología, el desarrollo de la óptica.
Durante el primer siglo antes de Cristo los romanos fueron capaces de fabricar
vidrio, y encontrando que a determinadas curvaturas del vidrio las imágenes se
deformaban o magnificaban. Durante el siglo XII Salvino D’Armate convirtió
estos lentes de aumento en las primeras gafas que mejoraban o corregían algunos
defectos de visión, el invento debió ser muy apreciado por los comerciantes de
telas ya que estas les permitirán apreciar mejor la calidad de los diferentes
tejidos (Bellis, 2009).
👉 El telescopio: No fue sino hasta la década de 1590 cuando el fabricante de lentes holandés Zacharias Jansen comenzó a experimentar con lentes múltiples para obtener una mayor magnificación de los objetos. Jansen logró obtener una magnificación de 9x, pero los objetos aparecían borrosos. Galileo Galilei retomó el diseño de los Jensen y lo mejoró en 1609 desarrollando un instrumento capaz de amplificar las imágenes muy distantes, había nacido el telescopio, que significa “mirar a lo lejos”. Basados en la idea del telescopio Giovanni Faber acuñó la idea del microscopio en 1625 aunque nunca pudo desarrollar uno el mismo (Schatzki, 2014).

Figura 4.1. Telescopio de Galileo
👉 El microscopio de lupa: Sin embargo, el individuo único a quien le adjudicamos los mayores honores e n el desarrollo de los primeros microscopios es sin duda el mercader de telas Antonie van Leeuwenhoek. Van Leeuwenhoek nació en una familia de clase media, burgueses que no eran muy ricos y por lo tanto no se le suministró una educación universitaria de alto nivel, por lo cual no se hubiera pensado que van Leeuwenhoek llegaría al pináculo de la historia de las ciencias, sin embargo, con habilidad y diligencia logró algo muy importante, el desarrollo de una nueva tecnología y en consecuencia de un nuevo universo por explorar. Cerca de 1668 van Leeuwenhoek logró aprender a fabricar lentes, lo cual era una habilidad de importancia para un comerciante de telas, ya que una buena lente era la diferencia entre comprar basura o la mejor de las telas por el mismo precio. Sin embargo, van Leeuwenhoek decidió perfeccionar la tecnología de lentes al máximo posible desarrollando una especie de lupa capaz de aumentar hasta 270x. Este fue sin duda un avance importante, pues la máxima amplificación anterior a la superlupa-microscopio de van Leeuwenhoek había sido de 50x. Después del propio van Leeuwenhoek no se realizaron mayores progresos en la magnificación, aunque si en la comodidad del diseño (Bellis, 2009).

Figura 4.2. El microscopio de lupa de Leeuwenhoek.
👉 El microscopio de
mesa: De hecho, otro naturalista había
estado trabajando en la misma idea, hablamos de Robert Hooke, quien en 1665
había diseñado un microscopio propio, aunque no tan potente pues amplificaba
solo hasta 50x.
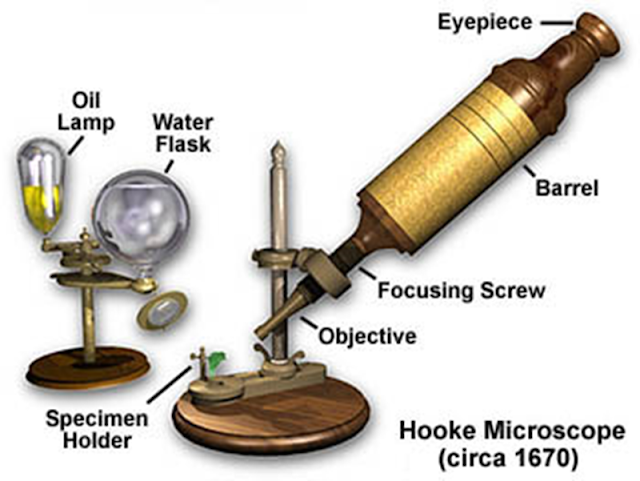
Figura 4.3. El microscopio de Hooke.
Sin embargo, el diseño de Hooke era mor mucho más cómodo para trabajar, y se convertiría a la postre en el diseño estándar de los microscopios ópticos. La calidad de la imagen de los microscopios no mejoró hasta que los alemanes se adueñaron del asunto, ya que eran los mejores en fabricación de lentes para todo tipo de aplicaciones científicas. Durante la década de 1850 Carl Zeiss un ingeniero alemán contrató a Otto Schott y Ernst Abbe para el mejoramiento de la tecnología del microscopio (Volkmann, 1966).

Figura 4.4. Microscopio
alemán del siglo XIX, con estos aparatos se estableció la primera teoría
celular.
Los microscopios de Schott y Abbe podían
enfocar objetos del tamaño de la longitud de onda o más grande, estableciendo
el primer límite de la tecnología microscopista, límites que no serían
superados hasta 1920 con el desarrollo de los microscopios electrónicos. Con
estos microscopios, los biólogos alemanes tuvieron la ventaja y en consecuencia
mucho del avance en el estudio de la estructura interna de la célula y de sus
procesos fisiológicos como la mitosis y la meiosis fueron hechos casi en su
totalidad por naturalistas germánicos.
4.2.
El descubrimiento de la célula
La célula fue descubierta en 1665 por
Robert Hooke (Hooke, 2003), en su libro Hooke dio 60 observaciones en detalle bajo su
microscopio compuesto. Varios de esos objetos fueron cortes muy delgados de
corcho, una estructura que se obtenía de las plantas, allí Hooke pudo observar
una serie de estructuras semejantes a las celdas de una colmena,
y las llamó tal cual como las celdas de una colmena de abejas en inglés
“cells”, sin embargo, la traducción al español no fue el de
celdas sino de células. Sin embargo, las células de un corcho
son solo las paredes celulares dado que están muertas, por lo que Hooke nunca
encontró evidencia de para que servían las celdas observadas bajo su
microscopio y ni siquiera pensó que estas estuvieran vivas en algún momento (Becker, Kleinsmith, & Hardin, 2005).

Figura 4.5. Células vegetales.
Anton van
Leeuwenhoek a diferencia de Hooke observó muestras vivas y se
dio cuenta de que estas estructuras poseían movimiento, y en consecuencia, es
el primero en adjudicarle la propiedad de vida a las células.

Figura 4.6. Anton van
Leeuwenhoek (24 de octubre de 1632, Países Bajos-26 de agosto de 1723),
conocido como el «padre de la microbiología», fue un comerciante neerlandés
que, además, sobresalió por ser el primero en realizar observaciones y
descubrimientos con microscopios cuya fabricación él mismo perfeccionó. La
historia de la biología lo considera precursor de la biología experimental, de
la biología celular y de la microbiología.
En 1676 van Leeuwenhoek logró describir
protozoos, bacterias y algunas células humanas como los glóbulos rojos, los
espermatozoides y el proceso de fertilización en animales con fecundación
externa, dando un golpe directo a la hipótesis de la generación espontánea,
aunque no mortal (Gest, 2004). Las células ancladas a tejidos en los animales fueron observadas
después de las de las plantas, debido a que los tejidos animales son más
frágiles. Durante la época los biólogos estaban buscando una unidad fundamental
para la vida, pero no estaban seguros de cual era. No fue sino hasta 100 años
más tarde que la idea de una unidad fundamental para la vida fue conectada con
la existencia de las células (Mazzarello, 1999).
4.3.
La teoría celular
Henri Dutrochet fue el primero en proponer
que las células eran la unidad estructural y funcional de los seres vivos (Nezelof, 2003). Sin embargo, la comunidad científica aún estaba dividida, pues se
pensaba que las células no estaban aisladas, sino que el fluido interno podía
transmitirse de una célula a otra. En 1804 Karl Rudolphi y J.H.F. Link
demostraron que las células poseían paredes celulares independientes y en
consecuencia que se encontraban aisladas funcionalmente (marquis Du Trochet, 1824).
👉 La primera teoría celular: Clásicamente el crédito por la teoría celular se les otorga a dos científicos: Theodor Schwann y Matthias Jakob Schleiden.

Figura 4.7. Friedrich Theodor
Schwann (Neuss, 7 de diciembre de 1810-Colonia, 11 de enero de 1882) fue un
naturalista, fisiólogo y anatomista prusiano, considerado uno de los fundadores
de la teoría celular. Además, las fermentaciones y las fibras nerviosas, en las
que describió la vaina de Schwann y contribuyó notablemente a la histología.
Mientras que Rudolf Virchow contribuyó a
esta, generalmente no se menciona en los libros de texto. En 1839 Schleiden
sugirió que cada parte estructura de una planta estaba hecha de células, o era
el producto de la acción de las células. El también sugirió que las células
estaban hechas por un proceso de cristalización desde el interior de otras
células o desde el exterior. De hecho, esta idea fue plagiada de Barthelemy
Dumortier. Este proceso de cristalización no se acepta en la actual teoría
celular.
En 1839 Theodor Schwann señaló que tanto
las plantas como los animales estaban compuestos por células o productos de
ellas (Schwann, 1839). Esto fue un avance importante, ya en aquel momento ya
era mucho más fácil observar las estructuras celulares de las plantas. Desde
estas conclusiones a cerca de las plantas y los animales, dos de los tres
principios fundamentales de la teoría celular fueron propuestos (Mazzarello,
1999).
- Todos los seres vivientes están compuestos por una o más
células.
- Las células son las unidades básicas de la vida
La hipótesis de Schleiden sobre la
cristalización fue refutada en la década de 1850 por Robert Remak, Rudolf
Virchow, y Albert Kolliker (Mazzarello, 1999). En 1855 Rudolf Virchow adicionó
el tercer postulado de la teoría celular:
- Todas las células emergen de células preexistentes.

Figura 4.8. Matthias Jacob
Schleiden (Hamburgo, 5 de abril de 1804-Fráncfort del Meno, 23 de junio de 1881)
fue un botánico alemán que, junto con su compatriota el fisiólogo Theodor
Schwann, formuló la teoría celular.
👉 Interpretación
moderna de la teoría celular: Actualmente
los científicos aceptan el hecho de que las células se originan de células
preexistentes para que la nueva vida pueda ser generada. Dado que todas las
células actuales comparten mecanismos esenciales y una relación de parentesco
que data de hace miles de millones de años, la moderna teoría celular posee los
siguientes enunciados.
Las células son las unidades estructurales y fisiológicas
de todos los seres vivos:
Las células son tanto entidades como unidades de
construcción de los seres vivos más complejos.
Todas las células provienen de células preexistentes.
Todas las células son similares en su composición química.
La mayoría de las reacciones químicas de la vida ocurren en
solución acuosa en el interior de las células.
La información genética es replicada y pasada a las
siguientes generaciones durante la división celular.
Los virus carecen de una estructura celular, pero dependen
de los seres vivos celulares.
5. Más que una bolsa de proteínas
Con la muerte del vitalismo en la química,
el camino quedó libre para que emergieran nuevos científicos que trabajarían en
la intersección de la bioquímica y la biología molecular experimental. Uno de
estos científicos fue Jacques Loeb (1859-1924). La fuerte convicción de Loeb de
que los fenómenos básicos de la vida pueden explicarse en principio por las
leyes físicas y químicas, estuvo acompañada de una preocupación por encontrar
explicaciones físicas y químicas para los fenómenos básicos de la vida. Esto
también lo convirtió en un pionero en los estudios relacionados con la
genética, la bioquímica, en particular la enzimología y el desarrollo. Los
bioquímicos esperaban que las funciones básicas de la célula y la vida pudieran
encontrar su explicación en las propiedades de estas enzimas. Loeb preveía que
"el carácter específico de cada célula posiblemente algún día se caracterice
por los fermentos específicos que contiene y produce" (Deichmann, 2011).
En una serie de artículos entre 1907 y
1915, Loeb, a través del experimento, desarrolló la idea de que los genes eran
los determinantes de las enzimas en desarrollo. Este razonamiento fue apreciado
por Thomas H. Morgan, quien en general no estaba dispuesto a especular sobre la
naturaleza química del gen y su acción y, en particular, rechazó las afirmaciones
de genes como enzimas, porque no tuvieron en cuenta la distinción crucialmente
importante de genotipo y fenotipo, propuesta por Wilhelm Johannsen. Morgan
consideró el punto de vista de Loeb y Chamberlain sobre genes y enzimas como "el
más correcto" (Deichmann, 2011).
Los intentos de Loeb de relacionar las
características centrales de la vida, como la herencia, con la química de las
macromoléculas contrastan fuertemente con el enfoque morfológico empleado por
la gran mayoría de los biólogos celulares de principios del siglo XX. Loeb fue
uno de los biólogos que más criticó al neodarwinismo de principios del siglo
XX, aunque no desde una base religiosa, sino con el interés de hacer de sus
hipótesis algo más manejable desde el punto de vista experimental (Deichmann, 2011).
El proyecto de investigación reduccionista
de Loeb unificó enfoques de fisiología, genética y embriología con el objetivo
de encontrar explicaciones fisicoquímicas (moleculares) para todos los procesos
de la vida, incluida la evolución. Este proyecto continuó más tarde en la
biología molecular, una síntesis de campos fisicoquímicos y biológicos. Aunque
el enfoque molecular ha sido altamente exitoso e indispensable para
investigaciones de nivel superior durante todo el siglo XX (Deichmann, 2011).
5.1.
Una bolsa de enzimas
Los grandes problemas de la biología del
siglo XIX eran de dos tipos conceptualmente bastante diferentes, y esta
diferencia se vería enormemente mejorada en el clima dentro del cual se
desarrolló la biología del siglo XIX y (especialmente) del siglo XX. Por un
lado, estaban los problemas "encapsulables", los del gen y la célula.
La comprensión aquí radica mucho en las partes. Por otro lado, estaban los
problemas holísticos, la evolución y la génesis y naturaleza de la forma
biológica, donde las partes no dan un sentido real del todo. La marcha de la
biología hacia el reduccionismo comenzó en serio con el
"redescubrimiento" del gen a principios del siglo XX. La disección
molecular de la célula, que había comenzado con al definir la función celular a
través de enzimas, realmente despegó con el advenimiento de la genética
molecular (Woese, 2004).
Iniciar con la genética de la célula era
lo más evidente desde el punto de vista técnico, los microscopios ópticos no
permitían ver al interior de las células nada más que los cromosomas durante la
mitosis, o el núcleo lleno de cromatina durante las etapas de crecimiento,
además, la acción más dramática que hacían las células eran sus procesos de
división celular, mitosis y meiosis. La pregunta radicaba en ¿Qué materiales
contenía la célula? La respuesta fue evidente cuando los análisis químicos
comenzaron a realizarse, la célula eran principalmente proteínas y enzimas, de
allí que muchos pensaran que la célula eran celdas o cubículos carentes de una
estructura interna que contenían una serie de componentes químicos que
permitían las reacciones químicas de los seres vivos como la fermentación. Sin
embargo, con el nacimiento de una nueva área interdisciplinar llamada
bioquímica, la confianza de los científicos sobre poder estudiar la naturaleza
de la vida solo Con base en causas fisicoquímicas comenzó a crecer de forma muy
rápida y en cierto sentido desproporcionada para lo que se sabía en el momento
(García-García, 2014).
Tal vez por esta visión carente de
estructura interna y extremadamente reduccionista de la naturaleza de la vida
hizo que varios investigadores intentaran realizar síntesis químicas para la
obtención de estructuras semejantes a bolsas o burbujas lipídicas –tal cual
eran consideradas las células en ese momento– como en el caso de Alfonso L.
Herrera “1868-1942”. Debido a la presunción a priori de la ausencia de una
estructura interna compleja de la célula, a no tomar en cuenta el tiempo
evolutivo –en este caso geológico– y a una línea de pensamiento
metodológicamente naturalista/materialista, Herrera razonó que sería
relativamente simple sintetizar una sustancia similar al protoplasma del
interior de la célula en una burbuja lipídica, para lo cual se valió de
sustancias como aceite de oliva, hidróxido de sodio, petróleo, gasolina,
tiocianato entre muchos otros (García-García, 2014, Herrera, 1932; Lazcano,
2010; Negrón-Mendoza, 1995; Aleksandr Ivanovich Oparin, 1957).
Este tipo de estudios sería denominado
plasmogenia y sería tan influyente como para permitir la creación de dos
revistas científicas: Gaceta de Plasmogamia “en español” y Laboratoire de
Plasmogenie “en francés” (Negrón-Mendoza, 1994), estas revistas fueron publicadas hasta 1942 –bien entrado el
siglo XX –cuando ya era claro que la estructura interna de la célula era algo
extremadamente complejo. Otra dimensión ausente en el trabajo de Herrera a
parte de la estructura interna de la célula es la noción del tiempo geológico “tiempo
irreversible”, sus síntesis no intentaban para nada ser un modelo de
simulación de condiciones antiguas. A pesar de esto, los trabajos de Herrera se
destacan por su enfoque basado en el materialismo metodológico experimental
(García-García, 2014).
Por el mismo periodo de tiempo los
bioquímicos comenzaban a avanzar sobre la naturaleza de la estructura interna
de los seres vivos, específicamente los componentes celulares. Aun cuando
muchas sustancias ya eran conocidas para los años 20s del siglo XX para los
biólogos y bioquímicos era bastante evidente que se sabía muy poco, en términos
del OdV esto tenía implicaciones ¿porque tratar de estudiar el origen de
algo que para empezar se entendía tan poco? (De Duve & Pizano, 1995;
García-García, 2014).
5.2.
Más que una bolsa de enzimas
Bajo el microscopio de luz, el citoplasma de una célula eucariota parece carente de estructura interna, sin embargo, incluso antes del comienzo del siglo XX, el examen cuidadoso de secciones pigmentadas de tejidos animales indicaba la existencia de un complejo sistema de estructuras internas. Sin embargo, no fue sino hasta la invención el microscopio electrónico en la década de 1940 que los biólogos comenzaron a apreciar la diversa gama de estructuras rodeadas por membranas que poseían la mayoría de las células eucariotas, e incluso, algunas procariotas.

Figura 5.1. Al microscopio
electrónico, resultó evidente que la célula era más que una bolsa, en su
interior tenía otras bolsas en una estructura compleja.
Los primeros usuarios del microscopio
electrónico vieron vesículas de diferentes tamaños, que transportaban
materiales de densidad variable, largos canales que radiaban en el citoplasma
formando redes como si fueran canales múltiples apretujados entre sí. Poco a
poco se hizo evidente que los mecanismos que permiten el funcionamiento celular
a nivel bioquímico ocurrían de forma especializada en estos compartimentos de
la estructura interna de las células. A medida que más células fueron
analizadas comenzaron a observarse patrones entre estas estructuras en
diferentes linajes, desde hongos unicelulares a plantas, desde amebas hasta el
ser humano.
5.3.
La estructura interna
Debido a que los organelos, sin importar
su fuente pueden ser clasificados en tipos más o menos generales, con funciones
consistentes, es que estos reciben nombres que son muy famosos, sin embargo,
podemos subdividir los organelos de la célula en membranosos y no membranosos.
Los organelos no membranosos no están rodeados por membranas, y en su lugar
están hechos de proteína o ARN, como el citoesqueleto, o los ribosomas. Los
organelos membranosos son definidos por las membranas biológicas y en general
son los siguientes: retículos cercanos al núcleo; retículos intermedios entre
la membrana externa y el núcleo; y vesículas de transporte o almacenamiento.
Estos organelos membranosos no son rígidos, se los debe visualizar como un
sistema de bombas de jabón dinámico, las vesículas se desprenden o fusionan con
los retículos más grandes, los cuales pueden crecer o hacerse más pequeños.
Tomados en su conjunto, estos organelos
conforman el sistema de membranas internas o endomembranas, en los que las
funciones individuales hacen parte de un todo coordinado. Debido a los
adelantos en biología evolutiva en la actualidad los organelos energéticos no
se clasifican como parte del sistema de membranas internas, pero eso es una
verdad a medias. Los organelos energéticos surgieron por endosimbiosis bacteriana,
por lo que las membranas más internas de estos no provienen de la célula
eucariota y en efecto no harían parte de sistema de membranas internas, pero la
última membrana que rodea a estos como la mitocondria y el cloroplasto proviene
evolutivamente de vesículas endocíticas, y por lo tanto si debiesen hacer parte
del sistema de membranas internas.
En el presente texto se los mencionará
como los organelos energéticos separados formalmente del sistema de membranas
internas. Para poder estudiar el sistema de membranas internas se requirió del
desarrollo de nuevas tecnologías que fueran más allá de la información que
puede proporcionar un microscopio óptico, y algunas de las más importantes
fueron las siguientes.
6. Como se estudia la célula
6.1.
El microscopio electrónico
El microscopio permite dos cosas
fundamentales, la primera es que permite ver la célula, y la segunda es que
permite ver las cosas que están en el interior de la célula. En primera
instancia, el microscopio incrementa el tamaño aparente de las cosas. Este
proceso se denomina magnificación. Sin embargo, el solo hecho de magnificar
algo no significa que puede verse con claridad.

{kind=link}
Figura 6.1. Las imágenes de
los microscopios electrónicos pasan directo a computadoras.
La segunda capacidad del microscopio es
que, al mismo tiempo que permite ver las cosas como si fueran más grandes,
también permite ver los detalles de sus formas, a esta propiedad la denominamos
resolución. Formalmente, la resolución es la distancia mínima en la que dos
objetos pueden alejarse y aun así seguirse viendo como dos cosas
independientes. La resolución del ojo humano es del alrededor de 0.2 milímetros
o (200 micrómetros).
La mayoría de las células son más pequeñas
que 200 micrómetros, y por lo tanto son invisibles al ojo humano. El
microscopio magnifica e incrementa la resolución de modo tal que las células y
sus estructuras internas pueden ser vistas. Existen dos tecnologías de
microscopio diferentes, el microscopio óptico y el microscopio electrónico.
El microscopio electrónico permite una
mayor resolución y magnificación, sin embargo, debe tenerse en cuenta de que lo
que se observa es un preparado especial, que requiere la deshidratación y por
lo tanto la muerte celular. Es decir, bajo el microscopio electrónico solo
pueden verse células muertas, por lo que los analistas deben tener eso en
cuenta a la hora de analizar los resultados. Los microscopios ópticos fueron
los primeros en aparecer, y su tecnología, aunque limitada para la amplificación
y la resolución, permite la observación de células vivas.
Antes de lidiar con los detalles de la
estructura celular es útil considerar los muchos usos que posee la microscopia.
Una rama completa de la medicina, la patología, hace uso de muchos métodos
diferentes de microscopia para ayudar en el análisis de las células en el
diagnóstico de las enfermedades.
Por ejemplo, un cirujano remueve una
muestra de tejido para que el patólogo determine si es tumoral o no. Para
realizar el esto, el patólogo deberá examinar rápidamente el tejido mediante
microscopia de contraste de fase o contraste de interface para determinar el
tamaño, la forma y la dispersión de las células. Usar colorantes
específicos para tinturar los tejidos en microscopia óptica para ver algunas
características como el núcleo, las características de la división celular o
algunos otros organelos que fijan los colorantes de manera densa. También se
pueden usar colorantes fluorescentes. Finalmente puede usarse el microscopio
electrónico para observar las estructuras internas en detalle como la
mitocondria y la cromatina.
6.2.
Autoradiografía
De todas las células del cuerpo, las
células ancinares poseen un sistema de membranas internas particularmente
grande. Estas células se especializan en la síntesis y secreción de las enzimas
digestivas. Después de haber sido segregadas por el páncreas, estas enzimas son
transportadas a través de varios conductos hasta el intestino delgado, donde se
encargan de la digestión química de los alimentos. ¿Dónde al interior de las
células ancinares se sintetizan las proteínas, y como es que estas sustancias
alcanzan el exterior de la célula sin digerirse esta misma?

{kind=link}
Figura 6.2. De todas las
células del cuerpo, las células ancinares poseen un sistema de membranas
internas particularmente grande. Estas células se especializan en la síntesis y
secreción de las enzimas digestivas.
Estas preguntas son inherentemente
difíciles de responder debido a que todos los pasos para la secreción ocurren de
forma simultánea en la célula. Para poder aislar los pasos en un solo ciclo
desde el ensamblaje del péptido hasta la maduración de la enzima activa,
James Jamieson y George Palade de la universidad de Rockefeller emplearon
una técnica denominada autoradiofragía.
La autoradiografía provee los medios
necesarios para visualizar los procesos bioquímicos permitiendo al investigador
determinar la localización de materiales marcados radioactivamente al interior
de una célula viva. En esta técnica, secciones de tejido que contienen isótopos
radiactivos son cubiertos por una delgada capa de emulsión fotográfica, la cual
es expuesta a la radiación que emana de los isótopos radiactivos del tejido.
Para determinar los lugares donde las
proteínas a segregar son sintetizadas, Palade y Jamieson incubaron placas de
tejido pancreático en una solución que contenía isótopos radioactivos por un
breve periodo de tiempo. Durante ese instante, los aminoácios marcados fueron
tomados por las células vivas e incorporados en sus sistemas metabólicos, lo
cual incluía la producción de enzimas a medida que estas iban siendo producidas
por los ribosomas.
A intervalos los tejidos fueron fijados,
es decir matar las células en momentos variantes para poder tener una imagen
estática que lo que sucede a lo largo de una secuencia de tiempo. Usando esta
técnica encontraron que el retículo endoplasmático rugoso era el principal
lugar de síntesis de los péptidos iniciales en la cadena de producción de las
enzimas activas. Para determinar la ruta intracelular los investigadores debían
encontrar una forma de eliminar el esceso de aminoácidos marcados, para esto
realizaron una segunda incubación con aminoácidos no marcados, de forma tal que
el retículo endoplasmático se liberara de los aminoácidos marcados y solo
pudiera verse las zonas en las cuales eran transferidos los péptidos después de
un tiempo.
Usando esta técnica fue posible
identificar la compleja ruta de las sustancias a través del sistema de
membranas internas. En resumen, hay dos grandes zonas de membranas, los
retículos cercanos al núcleo donde se sintetizan los péptidos, el aparato de
Golgi a medio camino donde se modifican y maduran los péptidos a las proteínas
funcionales o formas semiactivas, y las vesículas que transportan las
sustancias entre las membranas internas y la membrana externa.
6.3.
Proteína verde fluorescente
Técnicas empleando isotopos radioactivos han sido abandonadas paulatinamente por los biólogos. Una técnica alternativa requiere crear mutantes transgénicos empleando la inserción de un gen de medusa denominado gen de la proteína verde fluorescente. Esta proteína o por lo menos su dominio activo emite un color verde fluorescente que permite rastrear dicho dominio a medida que se mueve por la célula.

Figura 6.3. La microscopía
de fluoresencia permitió ver cosas que hasta entonces parecían agua, eran
invisibles.
Si se inserta el dominio de gen de la
proteína verde fluorescente en otra proteína, se genera una proteína quimérica
que además de sus dominios nativos posee el dominio de la proteína verde
fluorescente, esta proteína quimérica puede ser seguida de forma dinámica a
través del microscopio de fluorescencia para determinar la ruta de proteínas
especificas al interior de la célula.
Para insertar el gen de la proteína verde
fluorescente es necesario emplear retrovirus que inserten el gen en una
posición específica, uno de estos es el virus de la estomatitis vesicular. Los
virus como este son útiles ya que no solo insertan el gen deseado, sino que
también estimulan a que la célula produzca activamente el gen deseado
convirtiéndolas en fábricas muy eficientes del producto de interés.
6.4.
Microscopia electrónica de
fracciones subcelulares
Aunque las técnicas anteriores permiten
seguir la ruta biosintética, estas no permiten determinar la composición
molecular del sistema de membranas internas. En las décadas de 1950 y 1960
Albert Claude y Christian De Duve (Figura 20)
propusieron un nuevo método que consistía en el rompimiento de la célula por
homogenización. Cuando una célula se rompe por homogenización las membranas
internas y externa se fragmentan, los fragmentos se fusionan rápidamente entre
sí para formar vesículas de menos de 100 nm de diámetro. Las vesículas
derivadas de diferentes organelos poseen propiedades diferentes, lo cual
permite separarlas y estudiarlas independientemente, lo cual le da el nombre a la
técnica, fracciones subcelulares.

{kind=link}
Figura 6.4. Christian René Marie Joseph de
Duve (Thames Ditton, Surrey; 2 de octubre de 1917-Grez-Doiceau, Bélgica; 4 de
mayo de 2013) fue un citólogo y bioquímico inglés. Se le otorgó el Premio Nobel
de Fisiología o Medicina en 1974 compartido con Albert Claude y George Emil
Palade por haber descrito la estructura y funciones de los diferentes orgánulos
en el interior de las células.
Con el paso del tiempo las técnicas para
hacerlo se han mejorado, pero el principio es el mismo, separar las membranas
de diversos organelos y luego purificar las proteínas que les otorgan sus
propiedades específicas. Cientos de proteínas pueden ser identificadas
simultáneamente, proveyendo información de la composición molecular de
cualquier organelo. Un ejemplo de los resultados que otorga esta técnica es que
un simple fagosoma, que es una vesícula de menor tamaño puede contener más de
160 tipos de proteínas, muchas de las cuales ni siquiera se conocían antes o se
sospechara siquiera que estuvieran involucradas en la fagocitosis.
6.5.
Sistemas celulares libres
El fraccionamiento celular sirve para la
anatomía, pero también para la fisiología, ver de que son capaces las vesículas
fraccionadas representa un sistema celular libre. Cuando se realizaron los
procesos, los investigadores se dieron cuenta de que los sistemas subcelulares
aislados aun eran capaces de realizar actividades muy complejas, que no pueden
ser estudiadas con una sola enzima aislada. Durante la década de 1960 George
Palade, Philip Siekevitz,y sus colegas de la Universidad de Rockefeller
emplearon esta tecnología para estudiar fracciones del retículo endoplasmático,
obteniendo resultados como (1) la identificación de la función de los ribosomas
tanto de forma aislada –síntesis de proteínas liberadas al citosol –como al
interior de la vesícula, donde las proteínas eran almacenadas en el lumen de la
vesícula. Durante las décadas pasadas, la técnica de sistemas celulares libres
ha ayudado a determinar la función de muchas proteínas involucradas en el
tráfico de sustancias entre las membranas por medio de vesículas.
6.6.
Mutantes
El estudio de mutantes es un clásico en
genética molecular, y se basa en crear mutantes para genes de proteínas que se
conocen estructuralmente, pero cuya función en el todo no es clara. Un mutante
en este contexto es un organismo que genera una proteína que no pliega
adecuadamente para realizar su función, por lo que, al observar el sistema, el
mutante revela cual es la función que tenía al mostrar la carencia de dicha
función.
7. Las propiedades de las células
(YouTube) En base a la teoría celular, y a las propiedades emergentes de los diversos niveles de complejidad, podemos destacar la principal propiedad emergente de la célula, y no es otra que el de estar vivas. Todos los niveles de complejidad inferiores a la célula no pueden vivir, o no pueden realizar sus funciones de manera que podamos designarlas como algo vivo. Mientras que todos los niveles de complejidad biológica por encima de la célula los denominamos como vivos. A diferencia de un virus, un viroide o un prión, las células al estar aisladas pueden continuar realizando funciones vitales como las de alimentarse, mantener un ambiente interno regulado, y la capacidad de reproducirse. Es más, células de tejidos aisladas de sus seres vivos multicelulares poseen todas esas capacidades, si se las mantiene en medios de cultivo donde puedan crecer al igual que un cultivo de bacterias.

Figura 7.1. Cada una de las
líneas que vemos en esta fotografía de microscopia electrónica es una membrana.
Todas las células se definen en el sentido de que sus membranas las aíslan del
exterior y les dan su identidad. Toda vida conocida depende de una membrana
biológica, por lo que es justo decir que, las membranas biológicas son una
estructura que define la capacidad de estar vivo. Otras sustancias como el
genoma o las proteínas están presentes en cuerpos subcelulares como los virus,
los cuales no están vivos.
Los virus, viroides y priones simplemente
permanecen inertes cuando se encuentran fuera de las células, tanto viroides
como priones son macromoléculas sin ambiente interno, mientras que los virus
son incapaces de aprovechar las sustancias químicas a su alrededor en un
ambiente interno regulado, además ninguna de estas tres entidades es capaz de
reproducirse por cuenta propia en un ambiente apropiado, pero sin células, o
productos generados por las células.
Si bien, la vida es la propiedad celular
más importante, también debemos mencionar su opuesto, la muerte. Existen
células que literalmente nunca mueren si es que se mantienen en un ambiente con
las condiciones adecuadas. Al reproducirse una bacteria genera dos bacterias,
pero no muere, se duplica, y este fenómeno puede hacerlo de manera
indefinida. Las células de los seres vivos multicelulares si poseen una
característica de suicidio programado, este suicidio es generado por el propio
genoma, es decir, están programadas para morir. Por lo general la muerte
celular programada se da para evitar el desarrollo de líneas celulares
cancerígenas.
7.1.
Las células viven continuamente
La primera vez que se cultivaron células
humanas de manera artificial “in vitro”
fue en 1941 por los investigadores George y Martha Gey de la universidad
de Johns Hopkins. Las células fueron obtenidas de un tumor maligno y nombradas
células HeLa en honor a la donante Henrietta Lacks. Henrietta Lacks,
nacida en 1920 y murió en 1951, sin embargo, las células de su tumor siguen tan
vivas hoy, como lo estuvieron en los años 20s.

Figura 7.2. George Gey y Henrietta Lacks. Ella
nacida en 1920 y murió en 1951, sin embargo, las células de su tumor siguen tan
vivas hoy, como lo estuvieron en los años 20s, y de cierta manera han
evolucionado hasta convertirse en un ser vivo capaz de invadir y estropear
investigaciones (Brown, 2015; Lucey, Nelson-Rees, & Hutchins, 2009).
Las células HeLa aún siguen siendo
cultivadas en el laboratorio aun hoy, después de que la donante original
hubiera muerto muchos años atrás (Brown, 2015; Lucey, Nelson-Rees,
& Hutchins, 2009). Lo
que implica un fenómeno importante, una de las características de las células
cancerígenas es que no pueden morir. Es tan fuerte esta propiedad que incluso
se han convertido en contaminantes comunes de los medios de cultivo de biología
celular, en otras palabras las células HeLa se han convertido en un ser vivo
independiente en toda regla (Skloot & Turpin, 2010). Lo anterior si lo comparamos con las bacterias nos confiere una
característica básica de los seres vivos, la muerte no es una característica
esencial de la vida a nivel celular; a ese nivel la vida nunca envejece y
potencialmente puede existir para siempre.
7.2.
Las células son altamente
complejas y organizadas
La complejidad es una propiedad que es evidente cuando se la encuentra, pero difícil de describir en términos formales, claros y no ambiguos. Por el momento, podemos pensar en la complejidad en términos de orden y consistencia. Entre más compleja una estructura, mas será el número de partes de la componen, existirá una menor tolerancia a los errores en la naturaleza de estos componentes y en las interacciones de estos. Se requerirá de una mayor regulación para mantener al sistema en su estado.

Figura 7.3. (Izquierda)
Muchos cristales inorgánicos se organizan en estructuras muy complejas y
organizadas llamadas fractales, pero no por ello se dice que estén vivos.
(Derecha) A medida que la complejidad se incrementa, también la dificultad de
describir un suceso de manera matemática, y en ese sentido se dice que requiere
una mayor cantidad de información.
Sin embargo, hay varias cosas a tener en
cuenta, existen sistemas con múltiples componentes que son ordenados a pesar de
poseer múltiples componentes. Un ejemplo son los cristales y los fractales, los
cuales se componen de una misma unidad repetida de manera indefinida a través
de múltiples niveles de complejidad, y a pesar de todo pueden ser descritos con
ecuaciones matemáticas relativamente simples (Lewin, 1999; Mazzocchi, 2008; Zak,
Zbilut, & Meyers, 2008).

Figura 7.4. La célula debe
responder a los cambios ambientales, mediante cambios internos, un sistema
incapaz de responder al cambio, que no es dinámico no puede adaptarse a las
necesidades de un ambiente cambiante, ya sea en términos de un periodo de vida
celular, o en términos de una población de células variables a lo largo del
tiempo.
A medida que la complejidad se
incrementa, también la dificultad de describir un suceso de manera
matemática, y en ese sentido se dice que requiere una mayor cantidad de
información. Sin embargo un sistema complejo y dinámico no está vivo, al
carecer de un orden que le permite responder al ambiente y reproducirse (Lewin, 1999; Mazzocchi, 2008; Zak
et al., 2008).
La complejidad absoluta tampoco es la
complejidad a la que hacemos referencia, por ejemplo, si doblamos una cuerda de
manera que forme bucles de manera caótica, esta será increíblemente compleja,
pero tendrá una máxima tolerancia a cualquier cambio, ya que cualquier
microestado del sistema será igualmente valido. La complejidad biológica es una
que posee un orden, una flexibilidad ya que no es rígida, los microestados del
sistema cambian de un momento a otro, dentro de un rango límite de
posibilidades, es decir es una complejidad altamente dinámica. Estos detalles
son precisamente lo que hacen difícil de definir la complejidad biológica, ya
que a diferencia de la complejidad absoluta, la complejidad biológica es una
complejidad ordenada y cambiante dentro de ciertos límites sobre los cuales, el
error ya no se puede tolerar, pero dentro de cuyos límites el error simplemente
es diversidad (Lewin, 1999; Mazzocchi, 2008; Zak
et al., 2008).
Penemos por ejemplo en el código genético,
este es un sistema complejo, pero con un orden y una función, el cual se
encuentra regulado, pero posee cierta tolerancia a los cambios. Los cambios sin
embargo son minúsculos con respecto al todo del genoma. Por ejemplo, cuando el
genoma se duplica, la tasa de mutaciones son el de un cambio “error o
variabilidad” por cada diez millones de posiciones para equivocarse; esto se debe
a que los errores son corregidos la mayoría de las veces por los sistemas de
reparación del genoma de la célula, pero estos sistemas no son perfectos, y la
mayoría de las veces, los cambios en el genoma no sin ni letales ni
beneficiosos.
En el ejemplo anterior podemos ver todas
las características de la complejidad de lo vivo, una complejidad ordenada y
que cambia con el tiempo bajo ciertos límites establecidos.
7.3.
Las células poseen material
genético
Los seres vivos son construidos de acuerdo a la información recolectada en el genoma. Sin embargo, vale hacer una aclaración en este punto. A diferencia de la información en términos humanos, que es una abstracción de la realidad; la información biológica es una realidad física que se transmite de una molécula a otra y que tiene que ver con la conservación de las formas físicas reales, ubicadas en un lugar real de la célula.

Figura 7.5. El ADN con su
estructura de doble hélice almacena la información para generar todas las demás
partes de la célula, pero se requieren las demás partes de la célula para que
el ADN funcione. Sin la célula, el ADN es un polímero inerte, como cualquier
polímero sintético. A diferencia de lo que se creía anteriormente, el genoma no
solo está compuesto por el ADN si no por otras sustancias como el ARN y
proteínas reguladoras que lo controlan y le otorgan sus propiedades.
El programa genético humano contiene
suficiente información, si es convertida en palabras para llenar millones de
páginas de texto. Esta vasta cantidad de información es empaquetada en
estructuras macromoleculares denominadas cromosomas, las cuales contienen
diversas proteínas que regulan al material genético. Los cromosomas ocupan un
lugar en el espacio físico del núcleo de la célula, la cual es cientos de veces
más pequeña que la punta de un alfiler.
La información biológica almacenada en los
genes debe ser expresada a otras moléculas de alto peso molecular, las cuales
se encargan de realizar la verdadera labor, ya sea realizando funciones, sirviendo
como unidades estructurales, o bloqueando/impulsando la producción de más
sustancias a partir del genoma.
El genoma es un sistema complejo y
ordenado, pero también dinámico, esto implica que cambia con el tiempo dentro
de ciertas limitantes, estos cambios en el genoma confieren nuevas propiedades
a las células, lo cual conlleva al proceso de evolución. Sin embargo, la
evolución es un proceso que toma varias generaciones de células, y una gran
cantidad de estas, es decir, es una propiedad emergente de la población de
células, no de una célula individual.
Sobre el genoma, se encuentran una serie
de proteínas que actúan sobre el genoma, estas proteínas regulan sus
capacidades. Aunque no hacen parte del genoma, estas proteínas de
regulación, así como otros agentes como el ARN pueden afectar al genoma de
manera hereditaria, por lo cual los denominamos epigenoma.
7.4.
Las células poseen una función
reproductiva
Las células son capaces de producir más individuos a partir de sus propios cuerpos, a este proceso lo denominamos reproducción. A nivel celular existe una amplia gama de modos reproductivos, pero los principales son dos, la reproducción sexual y la reproducción asexual. En la reproducción asexual solo se requiere de un individuo para generar más individuos a partir de su propio cuerpo. En la reproducción sexual se requieren dos individuos para generar más, lo cual requiere la fusión y luego división de las células.

Figura 7.6. Una ameba se come a dos paramecios (YouTube).
La reproducción sexual es más compleja,
pero les confiere nuevas propiedades a las células desencintes vía ciertos
mecanismos que incrementan la diversidad de las hijas sin recurrir a la
mutación. Este proceso fue originalmente estudiado bajo el concepto de la
hipótesis de Weissman. Existen dos tipos básicos de reproducción asexual, la
bipartición de las células procariotas y la mitosis de las células eucariotas.
Existe un solo tipo de reproducción sexual regulado por los procesos de meiosis
y fecundación en las células eucariotas y algunos tipos de parasexualidad
“casi-sexualidad” en las células procariotas.
7.5.
Las células poseen la función
de nutrición
Todo proceso biológico, ya sea a nivel celular, o subcelular está relacionado con la transformación de la energía obtenida del ambiente externo. Virtualmente toda la energía empleada en la biosfera proviene del Sol a través de la forma de transporte de energía denominada radiación electromagnética. La energía electromagnética es luego transformada en energía química por medio de la absorción energética de los electrones de ciertos elementos como el magnesio ubicado en el cetro de la molécula llamada clorofila. Los electrones energizados luego son empleados para construir enlaces entre moléculas, los cuales almacenan energía.
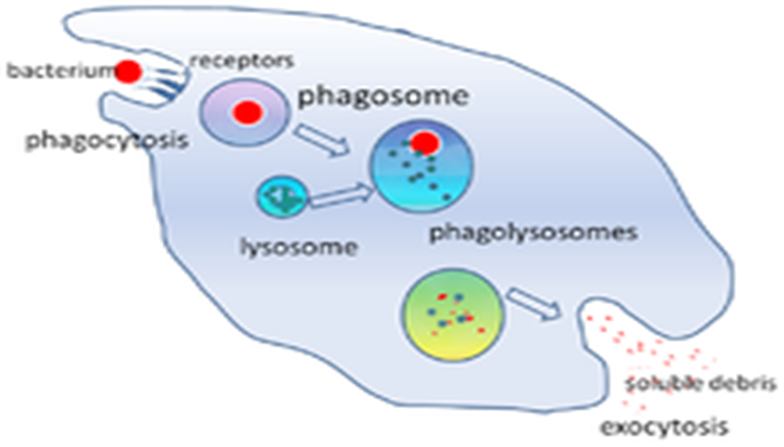
Figura 7.7. Las células,
cuando no hacen fotosíntesis, deben sustentarse mediante la obtención de
sustancias externas. Un modo es mediante la absorción de sustancias disueltas a
través de proteínas en la membrana llamadas receptores. Otra forma es mediante
la fagocitosis, este fenómeno ilustrado aquí requiere que la célula devore lo
que pretende como alimento, lo descomponga y finalmente libere las sustancias
de desecho.
Las moléculas cuyos enlaces almacenan
grandes cantidades de energía son muy diversas, algunas famosas son el adenosin
trifosfato ATP, los azucares y las grasas. Sin embargo, virtualmente toda
macromolécula se ha construido mediante la transformación de la energía solar.
Si el cuerpo requiere nuevamente de energía, los enlaces de estas moléculas se
romperán para realizar las transformaciones que la célula requiera.
La nutrición es un proceso que permite la
obtención ininterrumpida de energía del medio, ya sea por medio del Sol
“fotosíntesis” o por medio de la descomposición de sustancias que se encuentran
en el ambiente “heterotrofia”. Los modos de nutrición a nivel celular son
extremadamente diversos.
7.6.
Las células llevan a cabo
reacciones químicas
Los procesos de obtención de energía, se sincronizan con los de obtención de sustancias y con los de eliminación de sustancias, así como con los procesos de manutención del ambiente interno y de defensa de la célula. Todos estos procesos no son nada más que química en acción, todo proceso en la célula es una reacción química, controlada, acelerada, seleccionada, pero es a fin de cuentas un proceso químico.

Figura 7.8. Las reacciones
químicas llevadas a cabo por las células son numerosas y complejas,
generalmente requieren de un catalizador, es decir, una sustancia que acelera o
mejora la eficiencia de la reacción. En el esquema tenemos las reacciones del
ciclo de Krebs, encargadas de un paso de la obtención de energía por parte de
los alimentos en la respiración celular.
A la suma de todas las reacciones químicas
de la célula la denominaremos metabolismo. Existen dos tipos de metabolismo, el
metabolismo anabólico o constructivo, y el anabolismo o metabolismo
destructivo. Algunos de los procesos químicos más importantes a tener en cuenta
son la oxido-reducción en términos de la química orgánica, así como el
transporte de sustancias a través de membranas, y las propiedades de las
disoluciones acuosas.
Lo anterior se debe a que las reacciones
de reducción conllevan por lo general a la construcción de macromoléculas y por
ende al anabolismo, el cual requiere energía. Las reacciones de oxidación
generalmente conllevan al rompimiento de las macromoléculas, y por ende al
catabolismo, el cual libera energía. Por otro lado, la eficiencia de la
reacciones depende de las concentraciones de sustancias; mientras que el
transporte a través de la membrana es quien regula la concentración de las
sustancias en el agua. Después de todo, una célula es en su mayoría, agua.
7.7.
Las células realizan
actividades mecánicas
Las células son un ambiente de bulliciosa
actividad, los materiales son transportados de un lugar a otro, las estructuras
se ensamblan y luego rápidamente son degradadas, y en muchos casos, la célula
misma puede moverse de un lugar a otro.
Estos movimientos se basan en cambios dinámicos al interior de la
célula, los cuales dependen en últimas de reacciones químicas. Muchas células
poseen sustancias con diferentes formas y uniones, a estas sustancias las
llamamos proteínas “dada su capacidad de asumir una infinidad de formas”.
Algunas proteínas poseen forma de rotor, y giran de manera mecánica, algunas
agitando un largo filamento llamado flagelo, el cual las impulsa a través del
medio en el cual se encuentran.
7.8.
Las células responden a
estímulos
Algunas células responden a los estímulos de maneras obvias.

Figura 7.9. La quimiotaxis
es una respuesta al estímulo químico. Los receptores de membrana de la célula
captan los químicos y generan una respuesta mecánica, el movimiento de la
célula para acercarse a la fuente de las sustancias que activan a sus
receptores.
Algunas células complejas de vida libre
“llamadas de manera clásica como protistos” se mueven de un objeto a otro en
una ruta que los acerca a una posible fuente de nutrientes. Las células de los
seres vivos multicelulares como las plantas y los animales responden a los
estímulos de manera menos conspicua. Algunas células están cubiertas con
receptores que interactúan con las sustancias en el ambiente de maneras muy
específicas. Los receptores responden a hormonas, factores de crecimiento y
materiales extracelulares, así como a la superficie de otras células.
7.9.
Las células mantienen la
homeostasis
A demás de la reproducción la célula
quiere mantenerse viva, lo que en ultimas implica mantener un estado complejo
pero ordenado, y al mismo tiempo lo suficientemente flexible como para cambiar
en respuesta al ambiente lo más que sea posible. Este mantenimiento de un
estado complejo, ordenado, pero altamente cambiante al interior de una membrana
biológica como mínimo se denomina homeostasis. La homeostasis implica una
regulación la cual solo se hace evidente cuando se pierde.
Un ejemplo de ello son las mutaciones
deletéreas, las cuales se dan como resultado de la replicación del ADN durante
la reproducción celular. Las mutaciones deletéreas son debilitantes, o “en los
seres vivos multicelulares” puede conllevar a la destrucción de los limitantes
de reproducción, lo cual puede terminar en el desarrollo de un cáncer.
Gradualmente, los científicos han ido descubriendo como controlar estas
actividades, pero aún falta mucho por descubrir.
8. Generalidades de la célula y su estudio
8.1.
Tamaño celular
La mayoría de las células son minúsculas, uno de los primeros microscopistas llamado Robert Hooke estimó que una pulgada cuadrada de un corcho examinada con un microscopio de 1665 podía contener 1 259 721 000 células. El volumen de cada célula oscila entre 1 a 1000 micrómetros cúbicos. Existen algunas excepciones, células individuales de varios tipos de algas y de bacterias son los bastante grandes para poderlas ver a simple vista. Las neuronas a pesar de poseer un volumen celular “normal” poseen proyecciones del citoplasma que pueden extenderse por metros llevando señales de un lugar a otro del animal.

Figura 8.1. Esquema que
representa una escala logarítmica de tamaño de diferentes seres vivos. La
mayoría de las células rondan los tamaños de 100 micrómetros a 1 micrómetro. La
capacidad máxima de un humano para ver es de 200 micrómetros por lo que en la
práctica la mayoría de las células son invisibles.
Sin embargo, a pesar de que existe una
gran variedad de tamaños y volúmenes celulares, la mayoría de las células son
demasiado pequeñas. Este tamaño se debe a la relación entre tamaño y volumen. A
medida que un objeto su volumen, el área de contacto con el ambiente se hace
comparativamente más pequeña. Este fenómeno posee un gran impacto biológico por
dos razones.
El volumen de la célula determina la
cantidad de reacciones químicas que pueden darse en dilución por unidad de
tiempo, así como la cantidad de nutrientes que pueden almacenar. El área de
superficie determina la cantidad de sustancias que pueden entrar y salir de la
célula, así como la cantidad de reacciones que pueden llevarse a cabo
relacionadas con la membrana por unidad de tiempo.
A medida que la célula se hace más grande,
su actividad química y por lo tanto la necesidad de recursos y su tasa de
producción de desechos aumenta, pero la capacidad para obtener nutrientes y
eliminar desechos no crece al mismo tiempo, y de hecho en términos comparativos
disminuye. En otras palabras, entre más pequeña sea la célula, más eficiente
será un transporte de sustancias por la membrana y las reacciones que lleve en
el interior, asumiendo claro una forma rígida.
Existen dos modos de evadir este problema. El primero es mediante la modificación de la forma de la célula, si la célula no es esférica, sino más bien irregular, esta puede tener más área por unidad de volumen, pero esta solución es solo parcial, cuando la célula sigue creciendo llega a un límite en el que el cambio de forma aún sigue presentando el problema del área y el volumen.

Figura 8.2. Si dividimos el
área total en varias celas o "células" el área sumada de todas las
células supera en mucho al área del objeto único. En el caso presente, una célula grande posee
un área de 6 m2; mientras que con varias células pequeñas el área se incrementa
a 18 metros cuadrados. Si las células fueran microscópicas, el área total seria
enorme.
La segunda forma es que los organismos estén compuestos por varias células pequeñas, de este modo, aunque se tiene un volumen más grande, el área individual de cada célula representa un valor mucho mayor que el de solo una célula gigantesca.

Figura 8.3. Thiomargarita namibiensis al lado de una
abeja. Thiomargarita también bacteria
más grande y puede verse a simple vista, de hecho, varias células pueden verse
como "granitos" al lado de esta abeja. Visualizar el tamaño de una
célula es algo complicado, sin embargo, podemos realizarlo entendiendo algo del
sistema internacional de unidades.
La unidad básica es el metro, un centímetro
equivale a 0,01 metro (10-2). Un milímetro es la milésima parte de
un metro, lo cual equivale a 0,001 m (10-3). Ahora, imagine que un
milímetro tiene el tamaño de un metro, la milésima parte de ese milímetro sería
un micrómetro el cual se simboliza con el símbolo griego mu (µ) 1 µm
“micrómetro” equivale a (10-9) metros. La mayoría de las células
posee un tamaño entre 1 y 200 µm.
Su a su vez asumiéramos que l µm posee el
tamaño de un metro, la milésima parte de este sería un nanómetro (nm). Las
sustancias que miden su tamaño en nm son extremadamente pequeñas, como las
moléculas. Las ondas de luz también se separan entre sí por distancias medidas
en nm. Si asumiéramos de un (nm) es un metro, la décima parte de este sería un
Angstrom (Å) el cual equivale a (10-10) metros. El Å es generalmente
empleado para medir escalas atómicas. Por ejemplo, 1 Å es aproximadamente el
diametro de la nube de electrones del hidrogeno. Las moléculas y macromoléculas
pueden medirse en Å o en nm. Por ejemplo, la mioglobina, una proteína globular
típica posee unas medias de 4,5 nm X 3,5 nm X 2,5 nm. Proteínas altamente
alongadas como el colágeno o la miosina poseen hasta más de 100 nm de longitud.
El ADN posee alrededor de 2 nm de radio de giro. Macromoléculas complejas como
los ribosomas, los microtúbulos, los microfilamentos, poseen entre 5 y 25 nm de
diámetro.
A pesar de sus tamaños minúsculos, estas
moléculas realizan funciones mecánicas similares a las de las maquinas
macroscópicas “salvando las diferencias”, por lo que se las describe como
maquinas moleculares. Las células y los organelos son más fácilmente descritos
en micrómetros. El núcleo celular, por ejemplo, posee un tamaño de entre 5 a 10
µm de diámetro y la mitocondria es de aproximadamente 10 µm de largo. Mientras
que las células eucarióticas en promedio pueden tener entre 10 y 30 µm. Las
células procarióticas poseen un largo promedio de entre 1 y 5 µm.
8.2.
La unidad de la vida
Sin embargo, antes de irnos a las diferencias, hablaremos de las similitudes, las células procariotas y eucarióticas revelan una de las separaciones o clasificaciones más fundamentales en el mundo viviente, y aun así, a pesar de sus diferencias ambos tipos de células no son solo identificables como seres vivos, si no como seres vivos de un mismo tipo fundamental.

Figura 8.4. El ribosoma
"ilustrado en el video de color azul" es una estructura de alto peso
molecular que combina péptidos y secuencias de ARN estructural llamada ARN
ribosomal. La función del ribosoma es sintetizar cadenas de polipéptidos que
luego formaran las proteínas. La acción de la síntesis es llevada a cabo en una
región activa compuesta casi en su mayoría de ARNr, por lo que podemos decir
que el ribosoma es una enorme ribozima con péptidos accesorios (YouTube).
Lo anterior se debe a que, en mayor o
menor medida, aun con la transferencia horizontal de genes, la mayor parte del
genoma de los seres vivos, o sea de ambos tipos de célula descienden de un
ancestro común. Esto lo sabemos porque ambos tipos de célula usan el mismo tipo
de material genético, el mismo tipo de maquinaria molecular para formar
proteínas, los mismos códigos, e incluso, los mismos genes y las mismas
proteínas.
En términos de porcentajes, una célula
bacteriana muy estudiada Escherichia coli
comparte con el ser humano hasta el 60% de sus genes, lo cual revela como,
incluso a un nivel fundamental, los dos tipos de célula obedecen a un patrón
único, siendo un mismo tipo de vida. En la siguiente lista enunciaremos algunas
de las características que son similares entre eucariotes y procariotes.
👉 Membrana
celular de construcción similar
👉 Almacenamiento
de la información genética en ADN, el cual se "lee" del mismo modo,
es decir su código está compartido, una misma secuencia genera una misma
proteína sin importar que el gen esté en una célula procariota o eucariota.
👉 Mecanismos
similares de conversión de la información biológica almacenada "ADN"
a información biológica activa "proteínas", mediante diversos tipos
de moléculas mensajeras "ARN" y un mismo tipo de mecanismo de lectura
"ribosoma y código de codones".
👉 Mismas
rutas metabólicas como la descomposición de la glucosa en energía
"glucolisis".
👉 Mecanismos
de fotosíntesis idénticos.
👉 Mismo
mecanismo para la inserción de proteínas en la membrana
👉 Presencia
de proteosomas "estructuras digestoras de proteínas" similares entre
los eucariotes y cierto tipo de procariotes llamados arcaicos.
8.3.
¿Dos tipos de célula?
Todas las células se ven de manera
semejante a la luz de los microscopios ópticos primitivos, sin embargo, a
medida que la tecnología se fue perfeccionando, poco a poco empezaron a emerger
diferencias. Algunas células eran más grandes que otras, y presentaban una
estructura interna más compleja, al igual que una mayor diversidad en sus
formas. Otras células eran más pequeñas y relativamente más simples en su
composición interna.
La composición interna puede verse, porque
a ese grosor tan fino, la luz puede atravesar las paredes y las membranas
celulares permitiendo ver en su interior, mientras que estas membranas y
pareces apenas si pueden vislumbrarse como los bordes, en cierto sentido, a
pesar de que observamos una imagen de tres dimensiones, las imágenes de los
microscopios ópticos nos dan una percepción de dos dimensiones a menos que
muevas el objetivo; pero, algo como una foto de una microscopia dará
completamente la sensación de que observas algo plano, que realmente posee tres
dimensiones.
Una vez que una nueva tecnología fue
desarrollada y se convirtió en algo accesible para la mayoría de los grupos de
investigación, el microscopio electrónico, los biólogos fueron capaces de
examinar la estructura de una amplia variedad de células. Se volvió evidente a
partir de estos estudios que existían dos tipos de células, las cuales se
distinguían por sus tamaños, y el tipo de organización interna. La distinción
entre dos tipos celulares, sin ningún intermediario, representa una de las
divisiones evolutivas más fundamentales en el mundo biológico. De esta manera, comenzó a distinguirse entre
la célula eucariótica y la célula procariótica.
Existen seres vivos unicelulares con
células eucarióticas como las euglenas o las amebas; pero lo más importante es
que todos los seres vivos multicelulares poseen este tipo de células como su
unidad básica de composición. De esta manera, los hongos, las plantas y los
animales son todos clasificados como eucariotes en base a sus células. En
términos cronológicos no podemos saber cuándo fue que aparecieron las células
por primera vez, pero si podemos realizar algunas inferencias lógicas en base a
la información de la cual disponemos.
8.4. Registro fósil celular

Figura 8.5. Si dividimos la
historia de la Tierra en una torta, como en la imagen que presentamos, tenemos
que el origen del planeta está alrededor de 4.6 mil millones de años. La vida
aparece en el 3.8 mil millones de años, el oxígeno atmosférico y la
fotosíntesis del tipo I hace unos 3.6 mil millones de años, la fotosíntesis del
tipo II hace unos 2.6 mil millones de años, los eucariotes hace unos 2 mil
millones de años. Las algas, hace unos 1.2 mil millones de años, los
invertebrados, hace unos 0.6 mil millones de años.
El registro fósil de la vida unicelular se
extiende de manera prolongada y profunda en la edad geológica de la tierra.
Mientras que los animales complejos evolucionaron poco antes del cámbrico y se
diversificaron en la radiación del cámbrico alrededor de 600 a 500 millones de
años; las células han dejado registros fósiles hasta 3 500 millones de años
atrás “mda”.
Entre los fósiles celulares, podemos
distinguir las formas típicas de los eucariotes y de los procariotes. Mientras
que las células con características de eucariotas aparecen solo desde 2 000
millones de años, las células con características de procariotas aparecen desde
3 500 millones de años. En base a esta información podemos concluir que, las
primeras células en aparecer fueron las procariotas. Sin embargo, saber cuándo
se originaron es imposible, solo se puede afirmar que debe ser en algún momento
posterior a la lluvia de meteoritos de la Tierra primitiva y antes del 3 800
mda.
El cómo aparecieron las primeras células
resulta ser un misterio hasta ahora. La
característica emergente principal de la célula es el de estar viva, por lo que
estudiar como apareció desde sus unidades constituyentes requiere un estudio
desde lo no vivo. En otras palabras, el estudio del origen de la vida es
estudiado principalmente por las ramas de la bioquímica denominada química
prebiótica. Otro fenómeno de fecha importante, fue la gran proliferación de
oxígeno, fechada cerca de 2 400 mda; esta fecha coincide con el registro de
fósiles semejantes a un tipo de bacteria llamada cianobacteria, la cual es
capaz de realizar la fotosíntesis. A partir de esto, podemos inferir que el
proceso de la fotosíntesis evolucionó poco antes de 2 400 mda.
En base a los datos ofrecidos podemos ver
dos detalles importantes. La primera es que la vida aparece muy rápido en la
historia geológica de la Tierra; y la segunda es que la vida compleja tarda
mucho en desarrollarse.
9. La célula procariota
La distinción entre células procarióticas
y eucarióticas es basada en la complejidad y no en las relaciones
filogenéticas. A inicios del nuevo milenio, los avances en la investigación
bioquímica arrojaron lo que hasta ese entonces era un dato que alteraba los
fundamentos de la biología, la biología celular, la taxonomía, la evolución;
las células procariotas no pertenecían a un único grupo, no descendían de un
ancestro común. Más sorprendente a aun, de los dos grupos en los cuales se
dividió el antiguo grupo de procariotas, uno de ellos se encontraba más
relacionado a los eucariontes en ciertas características genéticas relevantes
que el otro.
Un nodo es un punto en que una línea
evolutiva se transforma en dos. Aunque actualmente la naturaleza de esta
separación es algo bastante complejo, en términos de clasificación es bastante
simple, pueden verse igual, pero existen dos grupos de procariotas, y uno de
ellos está más relacionado con las células eucariotas que con el otro grupo de
procariotas. Los dos grupos de bacterias fueron nombrados como bacterias
antiguas “archaeabacterias” y bacterias verdaderas “eubacterias”. Por
cuestiones de extensión, los dos grupos han venido a denominarse simplemente
como Archea “arcaicas” y Bacteria.
9.1. Las arcaicas

Figura 9.1. El árbol de la
vida, en verde las arcaicas, en azul las bacterias y en rojo los eucariotas.
Las especies mejor conocidas del demonio
de las arcaicas son aquellas que viven en ambientes extremadamente inhóspitos,
por lo que generalmente son denominados extremófilos. Incluidas entre las
archaeas se encuentran los metanógenos, procariotes capaces de convertir
metabólicamente el hidrógeno molecular y el dióxido de carbono en gases de
metano. También contamos a los halófilos
“amantes de las sales”. Los ambientes salinos en los que se encuentran son tan
extremos como los ambientes del mar muerto o en ciertas zonas del abismo
oceánico, donde las concentraciones de sal pueden llegar al 5 molar. Los
acidófilos son procariotes amantes de las sales, los cuales pueden llegar a
existir en pH de 0, los cuales se han encontrado en zonas de drenaje de minas
abandonadas. Los hipertermófilos son procariotes amantes del calor, los cuales
pueden vivir en ambientes hipercalentados. El agua hipercalentada es aquella
que se encuentra a una temperatura superior a su punto de ebullición. Tal
evento puede lograrse de dos maneras alternativas. La primera es que el agua se
encuentre pura e inmóvil. La segunda es que el agua se encuentre a una presión
mayor a la de una atmósfera. Existen
cepas capaces de vivir a temperaturas hipercalientes de 121°C lo cual implica
también que sobreviven a presiones más altas de las normales. Es interesante
que, la temperatura de 121°C es la que se emplea para esterilizar equipo
quirúrgico mediante el procedimiento del autoclave, básicamente una hoya a
presión que calienta al sistema a temperaturas hipercalientes con vapor de
agua.
9.2.
Las bacterias
Todos los demás procariotas conocidos pertenecen al grupo de las eubacterias “bacterias verdaderas” aunque la mayoría de los textos simplemente las nombran como bacterias.

Figura 9.2. Modelo de la
estructura interna de una bacteria.
El grupo de las bacterias incluye a las
células más pequeñas conocidas, el micoplasma (0,2 micrómetros de diámetro),
los cuales son los únicos procariotas en carecer de una pared celular. Su
genoma es igualmente pequeño, codificando cerca de 500 proteínas. Las bacterias
se encuentran presentes en todo ambiente concebible en el planeta Tierra, desde
los hielos permanentes del antártico, hasta los desiertos más secos de América
y África, e incluso hasta los confines del interior rocoso. Más interesante
aun, se encuentran, sobre, en el interior de los organismos, y a veces en el
interior de las mismas células de los eucariotas como las plantas y los
animales.
La estructura interna general de las eubacterias es simple, ADN, proteínas y otros bioelementos flotando en el citosol, sin un sistema de endomembranas. Sin embargo, esto es solo para la mayoría, no para todos. Las células bacterianas más complejas son las cianobacterias. Las cianobacterias contienen un sistema de MEMBRANAS INTERNAS llamadas tilacoides, los cuales contienen la clorofila necesaria para realizar la fotosíntesis. Los tilacoides de las cianobacterias son muy simulares a los tilacoides de los cloroplastos de las plantas. Tal como en los cloroplastos en las cianobacterias la fotosíntesis sigue en esencia las mismas rutas, usa las mismas moléculas, es guiada por las mismas proteínas y liberan finalmente el mismo oxígeno gaseoso.

Figura 9.3. Las bacterias
se cultivan generalmente en medios de cultivo semejantes a la gelatina, las
colonias son estructuras cerosas que crecen sobre o en el interior tal como se
muestra en la imagen. En este caso tenemos un cultivo de uno de los seres vivos
más estudiados y mejor conocidos entre todos, Escherichia coli.
Muchas cianobacterias no son capaces únicamente de la fotosíntesis, sino también de la fijación de nitrógeno. La fijación de nitrógeno es un proceso metabólico que permite capturar el gas de nitrógeno en moléculas útiles para los seres vivos como el amoniaco, el cual puede ser empleado subsecuentemente para sintetizar proteínas.

Figura 9.4. Las
cianobacterias son las eubacterias más complejas debido a que presentan estructuras
internas que flotan en la membrana diferentes a los biocompuestos y al ADN
flotando en el citosol, Los tilacoides son un verdadero sistema de membranas
internas en una bacteria, sin embargo, esta es la excepción y no la regla.
La atmósfera del planeta Tierra es en su
gran mayoría nitrógeno gaseoso “más del 70%”, pero la mayoría de los seres
vivos son incapaces de obtenerlo del aire. El nitrógeno es un elemento de vital
importancia para que los seres vivos puedan crecer y multiplicarse. La
capacidad de las cianobacterias que obtener recursos de fuentes inorgánicas, y
de convertir lo inorgánico a lo orgánico, hacen que posean la capacidad de ser
entre los primeros seres vivos en colonizar las rocas desnudas de un ambiente
nuevo o de ambientes extremos.
9.3.
Diversidad del mundo
procariótico, cultivables vs no cultivables
Para la mayoría de los microbiólogos los microorganismos familiares son aquellos que pueden ser cultivados en un medio controlado. Los medios de cultivo son sustancias semejantes a la gelatina, las cuales varían en su composición química. Algunas permiten el crecimiento de una amplia variedad de microorganismos, otras están diseñadas para dejar crecer solo unos cuantos microorganismos liberando en su crecimiento ciertas características como la forma en que crece la colonia y principalmente el color en que crece la colonia.

Figura 9.5. En la
microbiología clásica casi todo de reduce a poder cultivar el microorganismo y
a determinan sus propiedades bioquímicas mediante cambios de color de los
medios de cultivo específicos. Este acercamiento también permite determinar los
perfiles de proteínas y las estructuras complejas como los flagelos.
La microbiología clásica fue construida en
base a los medios de cultivo, y eso incluye a la microbiología médica. Por
ejemplo, cuando un paciente sufre de una infección urinaria, uno de os primeros
pasos que realiza en fisiólogo médico es el de cultivar al patógeno en un medio
de cultivo controlado para poder luego identificarlo. Una vez identificado
pueden ubicarse sus debilidades e iniciar el tratamiento más adecuado. Por los
azares del destino, se ha logrado cultivar la gran mayoría de los agentes
causantes de enfermedades, aunque algunos patógenos han demostrado ser más
retadores que otros como en el caso de la bacteria causante de la lepra.
Sin embargo, no puede decirse lo mismo de
la totalidad de la vida microbiana del planeta.
El problema se debe al hecho de que los procariotas pueden ser descritos
de manera muy rudimentaria por medio del microscopio, debido a que su
morfología no es muy distintiva “todos se ven muy parecidos”. Para identificar
a un microorganismo se deben hacer pruebas bioquímicas que nos dicen lo que
“pueden hacer”, y en base a ello se establecía una clasificación en base al
conocimiento acumulado por la microbiología clásica.
Las técnicas moleculares permiten
investigar la diversidad completa de una población microbiana, pero
difícilmente nos dice sobre las propiedades individuales de cada individuo, en
otras palabras, sabemos que existen muchas bacterias no cultivables, pero
desconocemos sus propiedades.
A la fecha, se han descrito
aproximadamente y en ocasiones de manera rudimentaria unas 6 000 especies de
procariotas mediante las técnicas tradicionales de la microbiología de cultivo,
lo cual representa menos de un décimo del uno por ciento de los millones de
especies de procariotas que existen en el planeta. Nuestra apreciación de la
diversidad de las comunidades procariotas se ha incrementado de manera
dramática en las recientes dos décadas mediante el uso de las técnicas
moleculares las cuales no requieren del aislamiento de un organismo particular.
Suponga que deseamos aprender a cerca de
la diversidad microbiana de las capas de sedimento más externas del océano
Pacifico en las costas de cualquiera nación que se desee. En lugar de intentar
cultivar a las bacterias, lo cual ha sido, de manera reiterativa un esfuerzo
fútil, los investigadores han optado por analizar el ADN contenido en las
muestras. Cada especie posee variantes únicas en su ADN aun en genes que
codifican para estructuras conservadas como el ribosoma, lo cual permite
generar una idea de la diversidad de una zona determinada.
Sin embargo, esta aproximación presenta
ciertos problemas, ya que nos hace ciegos ante los factores epigenéticos, y
ante el conocimiento de cómo es la diversidad morfológica de ciertas
características de importancia como lo es el flagelo bacteriano. Si no
conocemos la diversidad morfológica de la diversidad bacteriana que sabes que
está allí, nos perdemos de valiosas fuentes de información que pueden ayudarnos
a entender muchas cosas con respecto a la evolución de las especies sobre el
planeta Tierra.
9.4. Procariotas simbióticos y endosimbióticos

Figura 9.6. Modelo de la
estructura de una célula bacteriana típica de Escherichia coli.
Las técnicas de identificación molecular
han sido usadas para intentar dar un espectro de la diversidad que se encuentra
en el interior de los seres vivos, lo cual incluye a los seres humanos.
Los seres humanos poseen una serie de
pasajeros invisibles, sobre, dentro e inclusive en el interior de sus células.
La colección de microbios de un ser humano se denomina microbioma humano, y
actualmente es sujeto de muchas investigaciones, las cuales buscan relacionar
los cambios del microbioma humano con variables tales como la dieta, la edad,
ubicación geográfica y estados de salud. Con base a este tipo de
investigaciones ha llegado a demostrarse que, por ejemplo, los humanos obesos y
sedentarios poseen poblaciones de bacterias simbióticas marcadamente diferentes
en su sistema digestivo. A medida que una persona obesa pierde peso, la
constitución de las poblaciones microbianas de su tracto intestinal va cambiando
de manera marcada.
Los estudios en ratones sugieren que
algunas especies de bacterias que predominan en las personas obesas podrían
liberar más calorías desde la comida digerida que sus contrapartes en los
tractos digestivos de las personas delgadas, y por lo tanto, las bacterias de
las personas obesas harían que las personas ganen y mantengan su peso. Usando
las técnicas de secuenciación genética de la biología molecular se ha
descubierto que, lo que hasta hace unas décadas se pensaban que eran orgánulos
propios de las células, no eran más que polisones que se habían acostumbrado a
vivir en el interior de las células sirviendo a estas en una relación de
ganancia mutua, una endosimbiosis mutualista. Estos polisones evolucionaron
hacia las mitocondrias y los cloroplastos. La naturaleza y las pistas que
conllevaron a este descubrimiento serán explicadas en detalle en artículos
posteriores.
9.5.
Estructura interna de las
células procariotas
En las bacterias, el citoplasma está
rodeado por una membrana celular. La mayoría de las bacterias posee una pared
celular, y algunas veces también una cápsula. El citoplasma contiene una
variedad de enzimas. Las enzimas son proteínas especializadas en catalizar, es
decir en acelerar los muchos tipos de reacciones químicas que se requieren para
mantener un organismo. Como hemos discutido, las membranas plasmáticas de
procariotas y eucariotas tienen una estructura similar. Adicionalmente, poseen
una pared celular que mantiene la forma de la célula, incluso si el citoplasma
toma una gran cantidad de agua, el material de la pared es típico de las
bacterias y es conocido como peptidoglucano. A parte de la pared, algunas
bacterias poseen capas protectoras extra, una de ellas es la cápsula, esta es
una capa protectora de polisacáridos que se encuentra fuera de la pared
celular. En las bacterias, el ADN se encuentra en un único cromosoma circular
en espiral que reside en una región de la célula llamada nucleoide. Las muchas
proteínas formadas por los mensajes químicos del ADN bacteriano se sintetizan
en pequeñas estructuras llamadas ribosomas. El ADN bacteriano está organizado
secuencialmente, los genes se siguen linealmente sin que existan fragmentos no
codificantes, es decir, fragmentos de ADN incapaces de enviar mensajes químicos
para fabricar proteínas. Una célula bacteriana contiene miles de ribosomas. Algunas
bacterias tienen flagelos, que son apéndices en forma de cola que permiten que
las bacterias se impulsen. Un flagelo bacteriano no se mueve hacia adelante y
hacia atrás como un látigo. En cambio, mueve la celda en un movimiento
giratorio. Algunas veces, los flagelos ocurren solo en los extremos de una
célula, y otras veces se dispersan aleatoriamente sobre la superficie.
10. La célula eucariota
10.1.
Protistos
En muchos aspectos, las células eucariotas más complejas y capaces no se encuentran en el interior de las plantas y los animales. Estas células se encuentran en el interior de los muchos grupos de seres vivos unicelulares eucarióticos llamados protistos. Los protistos son un enorme grupo de seres vivos, los cuales en términos morfológicos aunque están compuestos por células eucariotas, no se los podía clasificar ni como un hongo, una planta o un animal, ellos son sus propios tipos de seres vivos eucariotas, tan diferentes a un animal, como lo es una planta a un hongo.

Figura 10.1. Protistas como Dictyostelium spp., se encuentran en la
transición entre la unicelular y lo multicelular (YouTube).
Giardia lamblia
expone la diversidad celular de los eucariotas, un
ser vivo que mantiene la mayor parte de su ciclo con dos núcleos haploides
independientes y activos. El estado de dicarionte como se describe a tener dos
núcleos que no se unen también se puede ver en los hongos y de hecho brevemente
en el cigoto humano. Entre los protistas unicelulares más comúnmente conocidos
contamos a las amebas, las euglenas y el volvox. Estas células se caracterizan
por realizar todas las funciones vitales, a diferencia de los eucariotes
multicelulares donde se da la tendencia a la especialización. Sin embargo,
existen casos en los que un protista puede presentar un estado de vida
unicelular, pero si las condiciones cambian pueden pasar a ser multicelulares
con una especialización celular relativamente poco avanzada. Los protistas no
son solo unicelulares, si no también multicelulares, tal como Dictyostelium spp., un ser vivo que en vida libre se comporta como una ameba,
pero cuando las condiciones se hacen duras se congrega sobre sí mismo formando
pequeñas babosas y luego una estructura fructificante, Sus células en la
estructura fructificante experimentan diferenciación, donde las de la punta
realizan la reproducción sexual, mientras que las del talo se sacrifican para
que la población sobreviva en otro lugar.
1.2.
La multicelularidad y la
especialización
Una forma alternativa de la evolución en el modo de vida individualista, fue el modo de vida grupal, en donde la célula individual pierde importancia en favor de la supervivencia del grupo. Los modos de multicelularidad no son igualmente complejos. Como ya mencionamos anteriormente, existen protistas generalmente unicelulares que se agrupan en ciertas eventualidades ambientales generando niveles de especialización celular parcial en donde solo algunos individuos lograran reproducirse, mientras que el resto se muere y no se reproduce, sacrificándose en favor del futuro.

Figura 10.2. La
multicelularidad ha evolucionado varias veces, una de las más recientes se ha
dado en el linaje del Volvox, un alga verde microscópica de la cual se
tienen los estadios morfológicos de varios puntos. Por ejemplo, dentro de Volvox
tenemos las Chlamidiomonas que son incelulares de vida libre, luego
tenemos a Gonium que es una agregación celular. Luego tenemos a Eudoria
que ya posee una matriz extracelular, lo que mantiene a las células unidas pero
a cierta distancia unas de otras. En Pleodorina ya tenemos un nivel
incipiente de especialización celular, mientras que Volvox como tal
posee tejidos tan especializados como el germinal.
Las esponjas también son agregados
celulares, las células presentan una pobre especialización y los tipos de
células apenas si pueden ser contados con los dedos de las manos, mientras que
los tipos de células de un ser humano pueden llegar a más de 200. En los seres
multicelulares donde la multicelularidad se va acentuando, el estado
multicelular se hace más prolongado, y se hace el estado en que la especie se
alimenta. En estos seres vivos, las células de la comunidad sufren
especializaciones muchísimo más marcadas. Esta especialización celular es
llevada a cabo mediante el proceso de diferenciación celular durante el
desarrollo embrionario de la especie.
Si
las esponjas son simples los placozoos lo son aún más, literalmente son una
masa de células con flagelos que le permite el movimiento, ostenta el título de
animal más simple y probablemente sea un ser vivo simplificado a partir de otro
con verdaderos tejidos. A diferencia de lo ocurre con los seres vivos como Dicyostelium spp., la diferenciación
celular en los eucariotes multicelulares es mucho más marcada, un solo embrión
humano puede dar lugar a 200 tipos de células diferentes, con formas, funciones
y bioquímica completamente diferentes, las cuales se coordinan para generar un
todo, una unidad coordinada, un ser vivo multicelular.
10.3.
Diferenciación celular y el
desarrollo
Como resultado de la diferenciación,
diferentes tipos de células adquieren una apariencia distintiva y contienen
materiales únicos, llevan a cabo reacciones químicas únicas y se encuentran en
zonas únicas de la anatomía del ser vivo. El cigoto puede reconocerse porque es
un dicarionte, a diferencia de lo que se esperaría, los núcleos del ovulo y el
espermatozoide no experimentan la cariogamia, en lugar de ello, iniciarla
primer mitosis o clivaje, y en el momento en que las membranas de los dos
núcleos se disuelven, el material genético se duplica y se reorganiza de modo
que las dos células del embrión si poseen el estado diploide, pero nunca se
fusionan los núcleos. A partir del cigoto se generan cientos de tipos de
células diferentes.
En los mamíferos por ejemplo tenemos las
células esqueléticas, musculares, glandulares, nerviosas, epiteliales,
sanguíneas, protectoras entre muchas otras más. Cuando el embrión alcanza el
nivel de gástrula, las células que lo componen ya se han diferenciado en tres
tipos generales, células del ectodermo, del mesodermo y del endodermo. Cada una
de estas tres dará lugar a un tipo de tejido y de órganos diferentes.
Una pregunta pertinente seria, ¿Cómo se
lleva a cabo la diferenciación celular? Anteriormente se pensaba que las
células eliminaban los genes innecesarios a medida que se iban diferenciando
unas de otras. Mientras que actualmente que más que eliminar, los genes se van
activando y desactivando. Prueba de esto es que, si se realiza una
transferencia de un núcleo somático a un ovulo, el ovulo activa nuevamente todos
los genes del núcleo reiniciando el programa y generando un embrión clonado de
manera somática.
10.4.
Estructura interna de las
células eucariotas
Las células eucariotas tienen una cantidad
limitada de rutas metabólicas, pero en compensación, poseen una gama casi
infinita de formas especializadas, a sea en los organismos de vida libre,
parásitos, o en organismos multicelulares, sin embargo, para simplificar las
cosas, por tradición se introducen solo dos, una célula animal idealizada, y
una célula vegetal idealizada, pero el lector deberá tener en cuenta que
existen muchos tipos y formas diferentes de las presentadas en este documento.

Figura 10.3. Estructura idealizada de la célula animal.
Deberías notar que las células eucariotas
están altamente compartimentadas. Estos compartimentos están formados por
membranas que crean espacios internos que dividen el trabajo necesario para
llevar a cabo las funciones de la vida. Los compartimentos de una célula
eucariota, típicamente llamados organelos u orgánulos, llevan a cabo funciones
especializadas que juntas permiten que la célula sea más eficiente y exitosa.
Casi todos los orgánulos están rodeados por una membrana con proteínas
integradas, muchas de las cuales son enzimas
(moléculas que aceleran las reacciones químicas). Estas enzimas hacen
productos específicos para ese orgánulo, pero su acción beneficia al sistema
celular completo.

Figura 10.5. Toxoplasma gondii es un parásito
intracelular que aprovecha el ciclo depredador presa, sus células son de tipo
eucariota al tener núcleo y un sistema de membranas internas, pero
adicionalmente posee organelos especializados que no son referenciados en los
cursos introductorios de biología celular.
La célula se puede ver como un sistema de
organelos interconectados que trabajan juntos para conducir y regular los
procesos de la vida. Por ejemplo, el núcleo es un compartimento que alberga el
material genético dentro de los cromosomas eucariotas, que contienen
información hereditaria. El núcleo se comunica con los ribosomas en el
citoplasma, y los orgánulos del sistema de endomembranas, especialmente el
retículo endoplásmico (RE) y el aparato de Golgi, se comunican entre sí.
La producción de moléculas específicas
tiene lugar dentro o sobre la superficie de los orgánulos. Estos productos son
luego transportados alrededor de la célula por vesículas de transporte, sacos
membranosos que encierran las moléculas y las mantienen separadas del
citoplasma. Por ejemplo, el retículo endoplásmico se comunica con el aparato de
Golgi por medio de vesículas de transporte.
Las vesículas se mueven por medio de una
extensa red de fibras de proteína llamada citoesqueleto, que también mantiene
la forma de la célula y ayuda con el movimiento de la célula. Estas fibras de
proteínas permiten que las vesículas se muevan de un orgánulo a otro. Los
organelos también se mueven de un lugar a otro usando este sistema de
transporte. Piense en el citoesqueleto como un sistema de vías tridimensional
dentro de las celdas utilizadas para transportar carga importante de un lugar a
otro. Los orgánulos relacionados con la energía -cloroplastos en plantas y
mitocondrias, tanto en plantas como en células animales- son responsables de
generar la mayor parte de la energía necesaria para realizar procesos
celulares.
11. Introducción a las partes de la célula
11.1.
Núcleo
Presente en: Eucariotas.
Debido a su gran tamaño, el núcleo es una de las estructuras más notables en la célula eucariótica. El núcleo contiene cromatina dentro de una matriz semifluida llamada nucleoplasma. La cromatina es una red de ADN, proteína y una pequeña cantidad de ARN. Justo antes de que la célula se divida, la cromatina se condensa y se enrolla en estructuras similares a varillas llamadas cromosomas. Todas las células de un organismo contienen el mismo número de cromosomas, excepto el óvulo y el esperma, que generalmente tienen la mitad de este número. Existen excepciones a esta regla como en los caballos, donde poblaciones aisladas pueden tener una cantidad de cromosomas diferente al resto de las poblaciones de la especie (Piras et al., 2009).

Figura 11.1. El núcleo
eucariota posee dos membranas, una interna que define su espacio interno, y
otra externa asociada al retículo endoplásmico rugoso (YouTube) (YouTube).
El ADN dentro de un cromosoma está
organizado en genes, cada uno de los cuales tiene una secuencia específica de
nucleótidos, que son los componentes químicos del ADN. Estos nucleótidos son
cuatro, y su secuencia es como un mensaje que sirve para codificar una
proteína, o algunas veces moléculas de ARN reguladoras. Por ahora, es
importante reconocer que la información en el ADN se procesa para producir ARN
mensajero (ARNm). Como su nombre lo sugiere, el ARNm actúa como un mensajero
entre el ADN y el ribosoma, donde se forman cadenas que darán lugar a las proteínas.
En las bacterias y arcaicas, no existe núcleo, sin embargo, parte de la
membrana única se pliega para formar una región semiespecial llamada nucleóide,
homóloga al núcleo real de las eucariotas.
Dentro del núcleo eucariota hay una estructura oscura llamada nucleolo. En el nucléolo, se produce un tipo de ARN llamado ARN ribosómico (ARNr). Las proteínas se unen con el ARNr para formar las subunidades de los ribosomas. Las subunidades ribosómicas ensambladas se envían desde el núcleo al citoplasma, donde se unen y asumen su papel en la síntesis de proteínas, en los eucariotas algunos los ribosomas se pegan a un organelo llamado retículo endoplasmático rugoso, que adquiere ese nombre precisamente porque su membrana está tachonada de ribosomas; en las arcaicas y bacterias, los ribosomas flotan en el citoplasma celular.

Figura 11.2. Las células de
los hongos pueden presentarse en dos formas, como hifas o como entidades
unicelulares “levaduras”. En cualquiera de las dos presentaciones hay un núcleo
y un sistema de membranas, por lo que son eucariotas, pero el forma de hifas,
las células pueden comunicarse entre sí a través de huecos en la pared celular
que las divide, en ocasiones algunas hifas pueden no tener separación entre sus
células, por lo que muchos núcleos pueden agruparse en la zona de crecimiento.
El núcleo eucariota está separado del
citoplasma por una membrana doble de fosfolípidos conocida como envoltura
nuclear. Localizados a través de la envoltura nuclear están los poros nucleares
que permiten que el núcleo se comunique con el citoplasma. Los poros nucleares
tienen un tamaño suficiente (100 nm) para permitir el paso de las subunidades
ribosómicas y las moléculas de ARN fuera del núcleo hacia el citoplasma, así
como el paso de proteínas desde el citoplasma al núcleo.
11.2.
Ribosomas
Presente en: Arcaicas,
Bacterias, Eucariotas.
Los ribosomas se componen de dos
subunidades, una grande y una pequeña. Cada subunidad tiene su propia
combinación de proteínas y ARNr.
El ribosoma actúa como una máquina de
escribir, y es aquí donde la información contenida en el ARNm del núcleo se
utiliza para sintetizar una proteína. Las proteínas pueden ser simples, o estar
hechas de dominios múltiples provenientes de mensajes diferentes, por lo que a
una proteína simple se la denomina a veces cadena polipeptídica o simplemente
polipéptido.
En las células eucariotas, algunos ribosomas se producen libremente dentro del citoplasma. Otros ribosomas están unidos al retículo endoplásmico (RE), un orgánulo del sistema de endomembrana. Después de que el ribosoma se une a un receptor en el ER, el polipéptido que se está sintetizando ingresa al lumen (interior) del RE, donde puede modificarse aún más y luego asumir su forma final.

Figura 11.3. El ribosoma (YouTube).
11.3.
Membranas celulares
Presente en: Bacterias, Arcaicas y Eucariotas
Son la barrera básica que se para a lo vivo de lo no vivo, están
compuesta por una capa doble de sustancias grasas conocidas como fosfolípidos,
esfingolípidos y colesterol que permiten un paso controlado de sustancias. El
paso de sustancias se da en puntos de control compuestos por proteínas que
pueden servir como canales o más comúnmente por sectores de paso mediado por
energía y cambio estructural o translocación. La membrana también tiene
proteínas receptoras que al unirse sustancias externas cambian su forma
interna, generando una reacción en cascada que envía información al núcleo de
la célula para que realice funciones específicas. Los organelos como los
retículos endoplásmicos, el aparato de Golgi, las vesículas, las mitocondrias y
cloroplastos también están definidos por membranas no muy diferentes de las
membranas biológicas externas, y muchas de sus funciones especiales dependen de
las proteínas que se encuentran insertadas en ellas.
11.4.
Sistema de membranas internas
Presente en:
Eucariotas.
El sistema de endomembranas consiste en la
envoltura nuclear, las membranas del retículo endoplásmico (RE) liso y rugoso,
el aparato de Golgi y numerosas vesículas de comunicación entre estos organelos
y con la membrana externa. Este sistema ayuda a compartimentar la célula, por
lo que las reacciones enzimáticas particulares se restringen a regiones
específicas, esto debido a que algunas reacciones se pueden obstruir entre sí
si no se aíslan de las demás, o incluso pueden ser peligrosas para el resto de
la célula si no se mantienen aisladas de las demás. Las vesículas de transporte
transportan moléculas de una parte del sistema a otra.
👉 Los retículos
endoplasmáticos: El retículo
endoplásmico (RE) consiste en un sistema interconectado de canales membranosos
y sáculos (vesículas aplanadas). Es físicamente continuo con la membrana
externa de la envoltura nuclear.
El retículo endoplásmico rugoso: Está tachonado con ribosomas en el lado de la membrana que está
frente al citoplasma; por lo tanto, el RE rugoso puede sintetizar polipéptidos
“proteínas inmaduras”. También modifica los polipéptidos después de que hayan
ingresado a la región central cerrada del RE, llamada lumen, donde las
proteínas toman forma. El RE rugoso forma vesículas de transporte, que llevan
las proteínas a otras partes de la célula. A menudo, las vesículas de
transporte están en camino hacia la membrana plasmática o el aparato de Golgi.
El retículo endoplásmico liso: Es continuo con el rugoso pero no tiene ribosomas unidos. El RE
liso sintetiza lípidos, como fosfolípidos y esteroides. Las funciones de RE
liso dependen de la celda particular. En los testículos, produce testosterona
y, en el hígado, ayuda a desintoxicar las toxinas. Independientemente de
cualquier función especializada, RE liso también forma vesículas de transporte
que transportan moléculas a otras partes de la célula, especialmente el aparato
de Golgi.
👉 El aparato de Golgi: El aparato de Golgi, llamado así por su
descubridor, Camillo Golgi, consiste en una pila de sáculos ligeramente
curvados y aplastados que se asemejan a panqueques. El aparato de Golgi puede
considerarse una estación de transferencia intermedia. En primer lugar, recibe
vesículas de transporte enviadas por los RE. Las moléculas dentro de las
vesículas se modifican a medida que se mueven entre los sáculos.
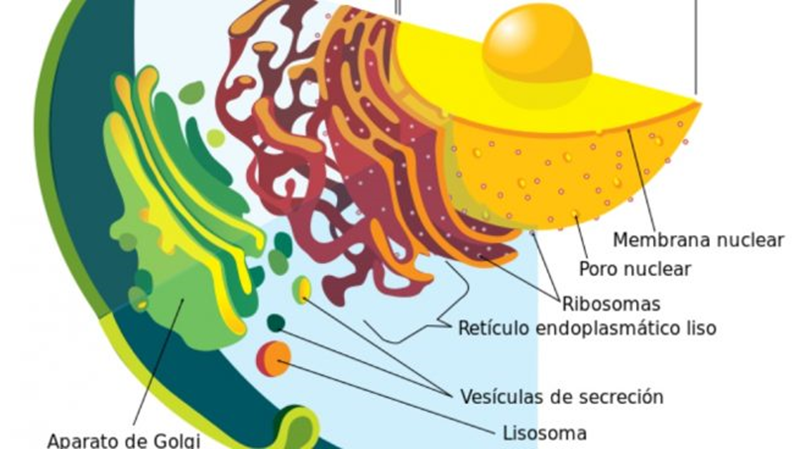
Figura 11.4. El aparato de Golgi (YouTube).
Por ejemplo, los azúcares pueden agregarse
o eliminarse de las proteínas dentro de los sáculos del Golgi. Finalmente, el
aparato de Golgi clasifica las moléculas modificadas y las empaqueta en nuevas
vesículas de transporte según sus destinos particulares. Las vesículas de
transporte salientes pueden regresar al RE, dirigirse a la membrana externa o
fusionarse con otras vesículas provenientes de la membrana externa.
👉 Vesículas: Las vesículas son fragmentos de membranas provenientes de la
membrana externa, de los retículos en doplasmáticos, o del aparato de Golgi que
transportan sustancias. Dependiendo del tipo de sustancias reciben nombres
especiales como lisosoma, liposoma, peroxisoma entre otros.
Los lisosomas son vesículas, producidas
por el aparato de Golgi, que digieren moléculas e incluso porciones de la
célula. A veces, después de englobar moléculas fuera de la célula, una vesícula
formada en la membrana plasmática se fusiona con un lisosoma. Las enzimas
lisosómicas luego digieren el contenido de la vesícula. Los liposomas
transportan sustancias grasas, y los peroxisomas transportan sustancias que se
encargan de oxidar las grasas. Existen muchos otros tipos de vesículas dependiendo
de la célula.
Las vacuolas, como las vesículas, son
sacos membranosos, pero las vacuolas son más grandes que las vesículas y están
más especializadas. Por ejemplo, las vacuolas contráctiles de los protistas
acuáticos están involucradas en la eliminación del exceso de agua de la célula.
Algunos protistas tienen grandes vacuolas digestivas para descomponer los
nutrientes. Las vacuolas de las plantas generalmente almacenan sustancias, como
nutrientes o iones. Estas vacuolas contienen no solo agua, azúcares y sales,
sino también pigmentos y moléculas tóxicas. Los pigmentos son responsables de
muchos de los colores rojo, azul y morado de las flores y algunas hojas. Las
sustancias tóxicas ayudan a proteger una planta de los animales herbívoros.
11.5.
Organelos generadores de
energía
Presente en: Eucariotas.
Los cloroplastos y las mitocondrias son
los dos orgánulos eucarióticos que se especializan en la conversión de energía.
Los cloroplastos usan energía solar para sintetizar carbohidratos. Las
mitocondrias descomponen los carbohidratos para producir moléculas de
adenosintrifosfato (ATP). La producción de ATP es de gran importancia porque el
ATP sirve como un portador de energía en las células. La energía del ATP se usa
siempre que una célula sintetiza moléculas, transporta moléculas o lleva a cabo
una función especial, como la contracción muscular o la conducción nerviosa.
Sin un suministro constante de ATP, ninguna célula podría existir por mucho
tiempo.
👉 Los cloroplastos: El cloroplasto, un orgánulo que se encuentra en las plantas y las
algas, es el lugar donde el dióxido de carbono, el gas, el agua y la energía
del sol se utilizan para producir carbohidratos mediante el proceso de
fotosíntesis. El cloroplasto es bastante grande, tiene el doble de ancho y
hasta cinco veces la longitud de una mitocondria. Los cloroplastos están unidos
por una membrana doble, que incluye una membrana externa y una membrana
interna.

Figura 11.5. El cloroplasto (YouTube).
El gran espacio interior, llamado estroma,
contiene una mezcla concentrada de enzimas y sacos en forma de discos llamados
tilacoides que forman una tercera membrana interna. Una pila de tilacoides “parecen
monedas apiladas” se llama un grano. Los lúmenes de los sacos tilacoideos
forman un gran compartimento interno denominado espacio tilacoidal. Los
pigmentos que capturan la energía solar se encuentran en la membrana de los
tilacoides, y las enzimas que sintetizan carbohidratos se encuentran en el
estroma. Los carbohidratos producidos por los cloroplastos sirven como
moléculas de nutrientes orgánicos para las plantas y, en última instancia, para
todos los organismos vivos en el planeta. El descubrimiento de que los
cloroplastos tienen su propio ADN y ribosomas respalda una teoría aceptada de
que los cloroplastos se derivan de bacterias fotosintéticas que ingresaron en
una célula eucariota en el pasado distante. Este proceso se llama
endosimbiosis.
👉 Las mitocondrias: Las mitocondrias son mucho más pequeñas que los cloroplastos y,
por lo general, son visibles solo con un microscopio electrónico. En las ilustraciones escolares o
introductorias visualizamos a las mitocondrias como si tuvieran forma de
frijol, pero en realidad a menudo cambian de forma, volviéndose más largas y
delgadas, o más cortas y más anchas. Las mitocondrias pueden formar cadenas
largas y móviles (como vagones en un tren), o pueden permanecer fijas en un
lugar (a menudo donde más se necesita energía). Por ejemplo, están empaquetadas
entre los elementos contráctiles de las células cardíacas (en el corazón) y
envueltas alrededor del flagelo de un espermatozoide. Al igual que los
cloroplastos, las mitocondrias están unidas por una doble membrana. La membrana
interna está plegada, y cada pliegue se denomina cresta, las crestas se
proyectan hacia el espacio interior, llamado matriz. Las cristas aumentan el
área de la superficie de la membrana interna tanto que, en una célula hepática,
representan aproximadamente un tercio de la membrana total en la célula.

Figura 11.6. La mitocondria (YouTube).
Las mitocondrias a menudo se llaman las
centrales energéticas de la célula porque producen la mayor parte del ATP que
utiliza la célula. La matriz contiene una mezcla altamente concentrada de
enzimas que ayuda a la descomposición de carbohidratos y otras moléculas de
nutrientes. Estas reacciones suministran la energía química que permite que la
síntesis de ATP tenga lugar en las crestas. La descomposición completa de los
carbohidratos, que también involucra el citoplasma, se llama respiración
celular porque se necesita oxígeno y se libera dióxido de carbono. La matriz
también contiene ADN mitocondrial y ribosomas mitocondriales independientes de
los nucleares. La presencia de ADN mitocondrial y ribosomas es evidencia de que
las mitocondrias y los cloroplastos tienen orígenes similares y se derivan de
las bacterias que se instalaron en una célula eucariota temprana. Al igual que
el origen de los cloroplastos, el origen de las mitocondrias es un ejemplo de
endosimbiosis. Todas las células eucariotas (con algunas raras excepciones)
tienen mitocondrias, pero solo los organismos fotosintéticos (plantas, algas y
varios protistas) tienen cloroplastos.
11.6.
El citoesqueleto
Presente en: Eucariotas
(principalmente).
El citoesqueleto es una red de filamentos
y túbulos proteicos interconectados que se extiende desde el núcleo hasta la
membrana plasmática en células eucariotas. De la misma manera que los huesos y
los músculos proporcionan una estructura animal y producen movimiento, los
elementos del citoesqueleto mantienen la forma de las células y, junto con las
proteínas motoras, permiten que la célula y sus orgánulos se muevan. Pero a
diferencia del esqueleto de un animal, el citoesqueleto es altamente dinámico:
sus elementos se pueden ensamblar y desmontar rápidamente según corresponda. El
citoesqueleto incluye microtúbulos, filamentos intermedios y filamentos de
actina.

Figura 11.7. El citoesqueleto (YouTube).
👉 Grúas moleculares: Las proteínas motoras asociadas con el citoesqueleto son
fundamentales para permitir los movimientos celulares. Las principales
proteínas motoras son la miosina, la quinesina y la dineína. La miosina a
menudo interactúa con los filamentos de actina cuando se produce movimiento.
Por ejemplo, la miosina está interactuando con los filamentos de actina cuando
las células se mueven de forma ameboide y / o envuelven partículas grandes en
la picnocitosis.

Figura 11.8. La quinesina es
una grúa molecular que permite el transporte de vesículas a través del
citoesqueleto.
Durante la división de células animales, la
actina, junto con la miosina, estrangulan la célula original en dos nuevas
células. Cuando una célula muscular se contrae, la miosina tira de los
filamentos de actina hacia el centro de la célula. La quinesina y la dineina se
mueven a lo largo de los microtúbulos tanto como una grúa viaja a lo largo de
una carretera jalando un material importante. En primer lugar, un orgánulo, tal
vez una vesícula, se combina con la proteína motora, y luego la proteína se
adhiere, se separa y se vuelve a unir a lo largo del microtúbulo, la estructura
se semejante a un gancho con patas que se mueven como caminando a lo largo del
citoesqueleto. De esta manera, el orgánulo se mueve de un lugar a otro en la
celda. La quinesin y dineína están de manera similar cuando las vesículas de
transporte llevan los materiales del aparato de Golgi a sus destinos finales.
👉 Microtúbulos: Los microtúbulos son pequeños cilindros huecos compuestos de 13
cadenas largas de tubulindímeros (dos moléculas de tubulina a la vez). Los
microtúbulos son dinámicos; pueden cambiar fácilmente su longitud eliminando
los dímeros de tubulina. Este proceso está controlado por el centrosoma, un
centro organizador de microtúbulos que se encuentra cerca del núcleo. Los
microtúbulos que irradian desde el centrosoma ayudan a mantener la forma de la
célula y actúan como pistas a lo largo de las cuales los orgánulos y otros
materiales pueden moverse jalados por la quonesina y la dineina.
👉 Filamentos
intermedios: Los filamentos
intermedios tienen un tamaño intermedio entre los filamentos de actina y los
microtúbulos. Son conjuntos de proteínas similares a los de las cuerdas que
normalmente se encuentran entre la envoltura nuclear y la membrana plasmática.
La red que forman es compatible tanto con el núcleo como con la membrana
plasmática. La proteína que compone los filamentos intermedios difiere según el
tipo de célula. Los filamentos intermedios hechos de proteína queratina
proporcionan una gran resistencia mecánica a las células de la piel por
ejemplo.
👉 Filamentos de actina: Cada filamento de actina consiste en dos cadenas de monómeros de
actina globulares retorcidos unos sobre otros de forma helicoidal para formar
un filamento largo. Los filamentos de actina sostienen la célula, formando una
red densa y compleja justo debajo de la membrana plasmática. Los filamentos de
actina también soportan proyecciones de la membrana plasmática, como las
microvellosidades.
1.7.
Centriolos
Presente en: Eucariotas
Ubicados en el centrosoma, los centriolos
son organelos cortos, en forma de barril, compuestos de microtúbulos. Es
posible que los centriolos den lugar a cuerpos basales, que se encuentran en la
base de cilios y flagelos eucarioticos y se cree que organizan los microtúbulos
en estas estructuras. Los centriolos también están involucrados en la
organización de los microtúbulos durante la división celular. Sin embargo,
algunos eucariotas, como las plantas y los hongos, carecen de centriolos
(aunque tienen centrosomas), lo que sugiere que los centriolos no son
necesarios para el ensamblaje de microtúbulos citoplásmicos.
1.8.
Cilios y flagelos
Presente en: Bacterias, Arcaicas
y Eucariotas
Los cilios y flagelos son proyecciones de
células en forma de látigo. Los cilios se mueven rígidamente, como un remo, los
flagelos eucarioticos se mueven de forma ondulada y en forma de serpiente,
mientras que en bacterias y arcaicas se mueven rotacionalmente. Los cilios son
cortos (2-10 μm) y los flagelos son más largos (generalmente no más de 200 μm).
Algunos protistas unicelulares utilizan cilios o flagelos para moverse. En
nuestros cuerpos, las células ciliadas son fundamentales para la salud
respiratoria y nuestra capacidad de reproducción. Las células ciliadas que
recubren nuestro tracto respiratorio barren los desechos atrapados dentro del
moco en la garganta, lo que ayuda a mantener los pulmones limpios. Del mismo
modo, las células ciliadas mueven al óvulo a lo largo del tubo uterino, donde
puede ser fertilizado por un espermatozoide flagelado.
1.9.
Paredes celulares
Presente en: Bacterias, Arcaicas
y Eucariotas
Una pared celular proporciona soporte y
forma a la célula. Las paredes de las células se encuentran en muchas células
eucariotas, incluidas las de las plantas, los hongos y la mayoría de los
protistas, pero no los de los animales. La composición de la pared celular
difiere entre los diferentes tipos de seres vivos, pero en esta sección nos
centraremos en la pared celular de la planta y bacterias.
👉 En plantas: Una pared celular primaria contiene fibrillas de celulosa y
sustancias no celulosas, y éstas permiten que la pared se estire cuando la
célula está creciendo. Las sustancias adhesivas son abundantes fuera de la
pared celular en la laminilla media, una capa que sostiene dos células
vegetales juntas. Para mayor resistencia, algunas células vegetales tienen una
pared celular secundaria que se forma dentro de la pared celular primaria. La
pared secundaria tiene una mayor cantidad de fibrillas de celulosa, que se
colocan en ángulo recto entre sí. La lignina, una sustancia que agrega fuerza,
es un ingrediente común de las paredes celulares secundarias.
👉 En bacterias: Como en otros organismos, la pared celular bacteriana proporciona
integridad estructural a la célula. En procariotas, la función principal de la
pared celular es proteger a la célula de la presión de turgencia interna
causada por las concentraciones mucho más altas de proteínas y otras moléculas
dentro de la célula en comparación con su entorno externo. La pared celular
bacteriana difiere de la de todos los demás organismos por la presencia de
peptidoglicano que se encuentra inmediatamente fuera de la membrana
citoplásmica. El peptidoglicano se compone de una cadena principal de
polisacáridos que consiste en la alternación de residuos de ácido
N-acetilmurámico (NAM) y N-acetilglucosamina (NAG) en cantidades iguales. El
peptidoglicano es responsable de la rigidez de la pared celular bacteriana y de
la determinación de la forma de la célula. Es relativamente poroso y no se
considera una barrera de permeabilidad para sustratos pequeños, y dado que sus
componentes no están presentes en los eucariotas es el blanco más común de los
antibióticos.
1.10.
La matriz extracelular
Presente en: Bacterias, Arcaicas
y Eucariotas
En biología, la matriz extracelular (MEC)
es una colección de moléculas extracelulares secretadas por células de soporte
que proporciona soporte estructural y bioquímico a las células circundantes. Debido
a que la multicelularidad evolucionó independientemente en diferentes linajes
multicelulares, la composición de la MEC varía entre las estructuras
multicelulares, e incluso, seres vivos que se clasifican como unicelulares como
las bacterias y arcaicas son capaces de formar una matriz extracelular llamada
biopelícula; Sin embargo, la adhesión celular, la comunicación y la
diferenciación célula a célula son funciones comunes de la ECM.
12. Referencias bibliográficas
Alberts,
B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P.
(2015). Molecular biology of the cell (5a ed.). Garland Science.
Aristotle. (2014a). History of animals. Recuperado a partir de
http://ebooks.adelaide.edu.au/a/aristotle/history/
Aristotle. (2014b). On the Generation of Animals. Recuperado a partir de
http://ebooks.adelaide.edu.au/a/aristotle/generation/index.html
Becker, W. M., Kleinsmith, L. J., & Hardin, J. (2005). The world of
the cell. Benjamin-Cummings Publishing Company.
Belk, C., & Maier, V. B. (2013). Biology Science for Life with
physiology. (Pearson, Ed.) (4a ed.).
Bellis, M. (2009). History of microscopes. Retrieved June, 29,
2009.
Benfey, O. T. (2007). The conceptual structure of the sciences. En Philosophy
of Chemistry: Synthesis of a New Discipline, Boston Studies in the Philosophy
and History of Science (pp. 95–117). Springer.
Benton, E. (1974). Vitalism in nineteenth-century scientific thought: A
typology and reassessment. Studies in History and Philosophy of Science Part
A, 5(1), 17–48.
Bergandi, D. (2011). Multifaceted ecology between organicism, emergentism
and reductionism. En Ecology Revisited (pp. 31–43). Springer.
Bersini, H., Stano, P., Luisi, P. L., & Bedau, M. A. (2012).
Philosophical and scientific perspectives on emergence. Synthese, 185(2), 165–169.
Blanco, D.
(2012). Aristas controversiales en la caja negra de Darwin. Lógoi, (8).
Breton, P. J. (1999). From microns to nanometers: early landmarks in the
science of scanning electron microscope imaging. Scanning Microsc, 13(1),
1–6.
Brown, J. (2015). Being cellular: Race, the inhuman, and the plasticity of
life. GLQ: A Journal of Lesbian and Gay Studies, 21(2–3),
321–341.
Butlerow, A. (1861). Formation synthétique d’une substance sucrée. CR
Acad. Sci, 53, 145–147.
Campbell, M. K., & Farrell, S. O. (2009). Biochemistry (6a
ed.). USA: Thomsom Brooks/Cole.
Cloutier, M. J. (1995). Antibiotics: Mechanisms of Action and Acquisition
of Resistance--When Magic Bullets Lose Their Magic. American Journal of
Pharmaceutical Education, 59(2), 167–172.
Cox, M. M., Doudna, J. A., & O’Donnell, M. (2012). Molecular
biology (1a ed.). Freeman.
Damiano, L. (2012). Co-emergences in life and science: a double proposal
for biological emergentism. Synthese, 185(2), 273–294.
Darwin, C. (1859). On the origins of species by means of natural
selection. London: Murray.
Darwin, F. (1887). The life and letters of Charles Darwin including an
autobiographical chapter edited by his son. London, Murray.
De Duve, C.,
& Pizano, M. (1995). Polvo vital: origen y evolución de la vida en la
tierra. Norma, Bogotá.
Deichmann, U. (2011). Early 20th-century research at the interfaces of
genetics, development, and evolution: Reflections on progress and dead ends. Developmental
biology, 357(1), 3–12.
Denton, M. J., Kumaramanickavel, G., & Legge, M. (2013). Cells as
irreducible wholes: the failure of mechanism and the possibility of an
organicist revival. Biology & Philosophy, 28(1), 31–52.
Drews, G. (2000). The roots of microbiology and the influence of Ferdinand
Cohn on microbiology of the 19th century. FEMS microbiology reviews, 24(3),
225–249.
Farley, J. (1972). The spontaneous generation controversy (1859–1880):
British and German reactions to the problem of abiogenesis. Journal of the
History of Biology, 5(2), 285–319.
Farley, J. (1974). The initial reactions of French biologists to Darwin’s
Origin of Species. Journal of the History of Biology, 7(2),
275–300.
García-García; J L. (2014). Un análisis histórico-epistemológico de tres aportes de la química
prebiótica. Universidad
pedagógica nacional de Colombia.
Gest, H. (2004). The discovery of microorganisms by Robert Hooke and
Antoni Van Leeuwenhoek, fellows of the Royal Society. Notes and Records of
the Royal Society, 58(2), 187–201.
Gilbert, S. F., & Sarkar, S. (2000). Embracing complexity: organicism
for the 21st century. Developmental dynamics, 219(1), 1–9.
Gottdenker, P. (1979). Francesco Redi and the fly experiments. Bulletin
of the History of Medicine, 53(4), 575.
Haeckel, E.
(1862). Die Radiolarien (Rhizopoda radiaria): eine Monographie (Vol. 1).
Reimer.
Henderson, J. (2012). Vitalism and the Darwin Debate. Science &
Education, 1–11.
Henry, F. G. (1999). Anti-Darwinism in France: Science and the Myth of
Nation. Nineteenth-century
French studies, 290–304.
Herrera, A. L.
(1932). La plasmogenia: nueva ciencia del origen de la vida. Cuadernos
de lectura. Recuperado a partir de http://www.uv.es/orilife/textos/Plasmogenia.pdf
Hoefnagels, M. (2015). Biology: concepts and investigations (3a
ed.). McGraw-Hill New York.
Hooke, R. (2003). 1665 Micrographia. London: Martyn and Allestry.
Hordijk, W., Hein, J., & Steel, M. (2010). Autocatalytic sets and the
origin of life. Entropy, 12(7), 1733–1742.
Hoyningen-Huene, P. (1991). Theory of antireductionist arguments: the Bohr
case study. En The Problem of Reductionism in Science (pp. 51–70).
Springer.
Jacob. F. (1973). La
lógica de lo viviente. Biblioteca científica Salvat.
Karp, G. C.
(2013). Cell and Molecular Biology, Concepts and Experiments (7a ed.). USA:
Wiley Online Library.
Lazcano, A. (2010). Historical development of origins research. Cold
Spring Harbor perspectives in biology, 2(11), a002089.
Lazzerini-Ospri,
L., Stano, P., Luisi, P., & Marangoni, R. (2012). Characterization of the
emergent properties of a synthetic quasi-cellular system. BMC bioinformatics,
13(Suppl 4), S9.
Lewin, R. (1999). Complexity: Life at the edge of chaos. University
of Chicago Press.
Lucey, B. P., Nelson-Rees, W. A., & Hutchins, G. M. (2009). Henrietta
Lacks, HeLa cells, and cell culture contamination. Archives of pathology
& laboratory medicine, 133(9), 1463–1467.
Mackean, D. G., & Hayward, D. (2014). Biology (3a ed.). IGCSE
Cambridge.
Mader, S. S. (2010). Biology (10a ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2015). Biology (12a ed.).
McGraw-Hill Education.
Malaterre, C. (2013). Life as an emergent phenomenon: From an alternative
to vitalism to an alternative to reductionism. En Vitalism and the
Scientific Image in Post-Enlightenment Life Science, 1800-2010 (pp.
155–178). Springer.
marquis Du Trochet, H. (1824). Recherches anatomiques et physiologiques
sur la structure intime des animaux et des végétaux, et sur leur motilité.
JB Baillière.
Mason, K. A., Losos, J. B., Singer, S. R., & Raven, P. H. (2014). Biology
(7a ed.). McGraw-Hill New York.
Mayr, E. (1997). This is biology: the science of the living world.
Universities Press.
Mazzarello, P. (1999). A unifying concept: the history of cell theory. Nature
Cell Biology, 1(1), E13–E15.
Mazzocchi, F. (2008). Complexity in biology. EMBO reports, 9(1),
10–14.
Miller, S. L., & Lazcano, A. (2002). Formation of the building blocks
of life. Life’s origin: the beginnings of biological evolution. California
University Press, Berkeley, 78–112.
Morowitz, H. J., Srinivasan, V., & Smith, E. (2010). Ligand field
theory and the origin of life as an emergent feature of the periodic table of
elements. The Biological Bulletin, 219(1), 1–6.
Morris, P. J. (1997). Louis Agassiz’s arguments against Darwinism in his
additions to the French translation of the Essay on Classification. Journal
of the History of Biology, 30(1), 121–134.
Murray, R. K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V.,
& Weil, A. (2012). Harpers Illustrated Biochemistry (29a ed.).
McGraw-Hill Medical.
Nakazawa, H. (2018). “Miller–Urey Experiment” in the Recent Picture of the
Early Earth. En Darwinian Evolution of Molecules (pp. 55–73). Springer.
Negrón-Mendoza, A. (1994). The Alexander Ivanovich Oparin 100th
Anniversary Conference. En The structure and model of the first cell (p.
7). Miramare, Italy: International Centre for Genetic Engineering and
Biotechnology.
Negrón-Mendoza, A. (1995). Alfonso L. Herrera: A Mexican pioneer in the
study of chemical evolution. Journal of biological physics, 20(1),
11–15.
Nezelof, C. (2003). Henri dutrochet (1776–1847): an unheralded discoverer
of the cell. Annals of diagnostic pathology, 7(4), 264–272.
Oparin, A. I. (1957). The origin of life on the earth. 3ed. The
origin of life on the Earth. Oliver & Boyd, Edinburgh & London.
Parke, E. C. (2014). Flies from meat and wasps from trees: Reevaluating
Francesco Redi’s spontaneous generation experiments. Studies in History and
Philosophy of Science Part C: Studies in History and Philosophy of Biological
and Biomedical Sciences, 45, 34–42.
Parmon, V. N. (2008). The Prebiotic Phase of the Origin of Life as Seen by
a Physical Chemist. En Biosphere Origin and Evolution (pp. 89–101).
Springer.
Parmon, V., & Snytnikov, V. (2002). The first Step of Evolution from
the View Point of chemical Catalysis. En 34th COSPAR Scientific Assembly
(Vol. 34, p. 489).
Peretó, J., Bada, J. L., & Lazcano, A. (2009). Charles Darwin and the
origin of life. Origins of Life and Evolution of Biospheres, 39(5),
395–406.
Piras, F. M., Nergadze, S. G., Poletto, V., Cerutti, F., Ryder, O. A.,
Leeb, T., … Giulotto, E. (2009). Phylogeny of horse chromosome 5q in the genus
Equus and centromere repositioning. Cytogenetic and genome research, 126(1–2),
165–172.
Pollard, T. D., Earnshaw, W. C., Lippincott-Schwartz, J., & Johnson,
G. T. (2017). Cell Biology (3a ed.). Elsevier.
Reece, J. B., Urry, L. A., Wasserman, S. A., Cain, M. L., Minorsky, P. V,
& Jackson, R. B. (2014). Campbell Biology (10a ed.). Pearson Higher
Ed.
Reisner, J. H. (1989). An early history of the electron microscope in the
United States. En Advances in Electronics and Electron Physics (Vol. 73,
pp. 133–231). Elsevier.
Rieppel, O. (2011). Ernst Haeckel (1834–1919) and the monophyly of life. Journal
of Zoological Systematics and Evolutionary Research, 49(1), 1–5.
Roll-Hansen, N. (1979). Experimental method and spontaneous generation:
the controversy between Pasteur and Pouchet, 1859–64. Journal of the history
of medicine and allied sciences, 34(3), 273–292.
Roll-Hansen, N. (2000). The application of complementarity to biology:
From Niels Bohr to Max Delbrück. Historical studies in the physical and
biological sciences, 417–442.
Ruestow, E. G. (1984). Leeuwenhoek and the campaign against spontaneous
generation. Journal of the History of Biology, 17(2), 225–248.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of
Biology (10a ed.). Sinauer & MacMillian.
Schatzki, S. C. (2014). The Microscope. American Journal of
Roentgenology, 202(1), 246–247.
Schwann, T. (1839). Mikroskopische Untersuchungen über die Uebereinstimmung
in der Struktur und dem Wachsthum der Thiere und Pflanzen: mit 4 Kupfertafeln.
Reimer.
Simon, E. J., Dickey, J. L., & Reece, J. B. (2013). Essential
biology (Pearson).
Skloot, R., & Turpin, B. (2010). The immortal life of Henrietta Lacks.
Solomon, E., Martin, C., Martin, D. W., & Berg, L. R. (2014). Biology
(10a ed.). Cengage Learning.
Spaargaren, D. H. (1985). Origin of life: Oceanic genesis, panspermia or
Darwin’s ‘warm little pond’? Experientia, 41(6), 719–727.
Starr, C., Evers, C., & Starr, L. (2013). Biology: Today and
Tomorrow With Physiology (4a ed.). Brooks/Cole.
Strick, J. E. (2009a). Darwin and the origin of life: public versus
private science. Endeavour, 33(4), 148–151.
Strick, J. E. (2009b). Sparks of life: Darwinism and the Victorian
debates over spontaneous generation. Harvard University Press.
Takai, K., Nakamura, K., Toki, T., Tsunogai, U., Miyazaki, M., Miyazaki,
J., … Horikoshi, K. (2008). Cell proliferation at 122 C and isotopically heavy
CH4 production by a hyperthermophilic methanogen under high-pressure
cultivation. Proceedings of the National Academy of Sciences, 105(31),
10949–10954.
Vlaardingerbroek, B. (2012). The Sorites Paradox,“Life,” and Abiogenesis. Evolution:
Education and Outreach, 1–3.
Volkmann, H. (1966). Ernst Abbe and his work. Applied optics, 5(11),
1720–1731.
Weaver, R. F. (2012). Molecular biology (5a ed.). McGraw-Hill
Education.
Woese, C. R. (2004). A new biology for a new century. Microbiol. Mol.
Biol. Rev., 68(2), 173–186.
Wolfe, C. T. (2007). The Return of Vitalism: Canguilhem and French
Biophilosophy in the 1960s.
Zak, M., Zbilut, J. P., & Meyers, R. E. (2008). From instability to
intelligence: complexity and predictability in nonlinear dynamics (Vol.
49). Springer Science & Business
Media.
No hay comentarios:
Publicar un comentario