
|| Principal || Cursos || Biología de la reproducción ||
Índice
Índice
|
||1|| Introducción ||2|| Generalidades de la reproducción ||3|| Historia de vida ||4|| Bacterias y arcaicas ||5|| Endosporas bacterianas ||6|| Pseudosexualidad ||7|| Reproducción asexual en hongos ||8|| Reproducción sexual | ||9|| Quitridiomicetos ||10|| Zigomicetos ||11|| Ascomicetos ||12|| Basidiomicetos ||13|| Protistas de vida libre ||14|| Falsos hongos ||15|| Algunos protistas parásitos |
Portada

Portada
1. Introducción
1. Introducción
La reproducción biológica es una de las funciones más importantes de los seres vivos, tanto así que siempre se encuentra en cualquier definición de vida. En los seres vivos la reproducción concierne a la habilidad de crear copias de sí mismo y que involucran a la duplicación celular. La habilidad de hacer copias de sí mismo puede realizarse por dos mecanismos generales, la reproducción asexual y la reproducción sexual, e involucrar individuos unicelulares y multicelulares. En la reproducción sexual un individuo crea copias exactas de sí mismo “excepto por algunas mutaciones de tipo SNP”, mientras que en la reproducción sexual se da el fenómeno de la recombinación meiótica. Toda reproducción sexual estará mediada por la meiosis y la fecundación generando lo que se conoce como ciclos de vida sexuales. Cuando la reproducción involucra individuos unicelulares, los únicos componentes importantes son las células que se dividen. En la reproducción sexual se requiere como mínimo de dos componentes, las células que son llamadas gametos y las hormonas o señalizadores químicos, que permiten a los gametos encontrarse mutuamente para poder realizar la fecundación. En seres vivos multicelulares, se adiciona un componente estructural, tejidos y órganos que no están involucrados con la fecundación directamente, pero que canalizan a los gametos y protegen al cigoto una vez formado.
1.1 Anton van Leeuwenhoek
Anton van Leeuwenhoek fue un científico y comerciante holandés, conocido por ser el padre de la microbiología. A lo largo de su vida, Leeuwenhoek desarrolló y perfeccionó técnicas para la fabricación de lentes microscópicas, que le permitieron observar organismos y estructuras microscópicas que nadie había visto antes. Sus descubrimientos, realizados a través de la observación de microorganismos en el agua, sedimentos y tejidos animales, sentaron las bases de la microbiología y revolucionaron la comprensión de la vida microscópica.

Contexto social
Anton van Leeuwenhoek nació en la ciudad holandesa de Delft en 1632, durante una época de grandes cambios políticos, económicos y culturales en Europa. En ese momento, los Países Bajos se encontraban en plena Guerra de los Ochenta Años contra España, que culminaría con la independencia holandesa en 1648. Además, los Países Bajos se habían convertido en una potencia comercial y marítima, lo que permitió un gran desarrollo económico y una creciente clase media.
En cuanto al contexto social, Leeuwenhoek pertenecía a la clase media, pero no tuvo una educación formal, ya que abandonó la escuela a una edad temprana para trabajar en el negocio de tejidos de su familia. Sin embargo, gracias a su curiosidad y su pasión por la ciencia, logró convertirse en uno de los científicos más destacados de su época.
En cuanto a la religión, los Países Bajos eran un país mayoritariamente protestante, pero también había una significativa comunidad católica. Leeuwenhoek, por su parte, era miembro de la Iglesia Reformada Holandesa, pero no se conocen detalles sobre sus creencias religiosas o su relación con la religión en general.
En cuanto al contexto cultural, el siglo XVII fue una época de grandes logros en el campo de la ciencia y la filosofía, conocida como la "Edad de la Razón". También fue una época de gran interés por el conocimiento y la exploración del mundo, lo que impulsó el desarrollo de nuevas tecnologías y herramientas para la observación y el estudio de la naturaleza.
Infancia
No se conocen muchos detalles específicos sobre la infancia de Anton van Leeuwenhoek, pero se sabe que nació el 24 de octubre de 1632 en Delft, una ciudad holandesa que en ese momento era un importante centro comercial y manufacturero. Era el quinto hijo de su familia y creció en un hogar modesto.
A pesar de su falta de educación formal, Leeuwenhoek tenía una mente curiosa y una pasión por la ciencia que lo llevó a realizar experimentos y observaciones por su cuenta. Por ejemplo, se sabe que construyó su primer microscopio a la edad de 20 años y comenzó a observar cosas como el ojo de una mosca y el aguijón de una abeja.
A pesar de que pasó la mayor parte de su vida en Delft, no se sabe mucho sobre su infancia en esa ciudad. Sin embargo, está claro que su pasión por la ciencia y la observación comenzó temprano y se convirtió en una de las fuerzas motrices de su vida.
Logros
Anton van Leeuwenhoek no recibió una educación formal, pero desarrolló una gran pasión por la ciencia y la observación de la naturaleza desde joven. Es conocido por sus notables contribuciones al campo de la biología, incluyendo la observación de microorganismos y la invención de microscopios de alta resolución que le permitieron ver detalles nunca antes vistos.
En 1674, Leeuwenhoek escribió su primera carta a la Royal Society de Londres, en la que describió la observación de organismos unicelulares en una muestra de agua. Esto fue un gran avance en la comprensión de la vida microscópica y allanó el camino para el descubrimiento de bacterias, protozoos y otros microorganismos.
Entre sus invenciones se encuentra un tipo de microscopio de alta resolución, que utilizaba una sola lente esférica para obtener una imagen extremadamente nítida. También diseñó un micrómetro para medir objetos vistos a través del microscopio, y fue uno de los primeros en utilizar iluminación artificial para mejorar las imágenes obtenidas.
Leeuwenhoek publicó sus observaciones en más de 200 cartas a la Royal Society de Londres y otros científicos europeos. Sus cartas incluyen descripciones detalladas de organismos como células sanguíneas, bacterias, protozoos y espermatozoides. También fue uno de los primeros en observar y describir la circulación de la sangre en los capilares.
En resumen, Anton van Leeuwenhoek es conocido por sus importantes contribuciones a la biología, incluyendo la observación y descripción de microorganismos y la invención de microscopios de alta resolución. Sus logros científicos incluyen la descripción detallada de células sanguíneas, bacterias, protozoos y espermatozoides, así como la observación y descripción de la circulación sanguínea en los capilares. Sus inventos incluyen microscopios de alta resolución, un micrómetro para medir objetos vistos a través del microscopio y la iluminación artificial para mejorar las imágenes obtenidas.
Comunidad científica
Anton van Leeuwenhoek no tuvo una comunidad científica formal, ya que trabajó de manera independiente como autodidacta y nunca obtuvo un título académico. Sin embargo, tuvo importantes colaboradores y amigos en el mundo científico, incluyendo a Reinier de Graaf, un médico y anatomista que lo introdujo en el mundo científico de Delft y lo recomendó a la Royal Society de Londres.
Leeuwenhoek también tuvo un gran apoyo por parte de Jan Swammerdam, un entomólogo y anatomista que también trabajó en la investigación microscópica. A pesar de que Swammerdam no estuvo de acuerdo con todas las teorías de Leeuwenhoek, sí lo apoyó y trabajó con él en varias investigaciones. Otros amigos y colaboradores incluyeron a Christiaan Huygens, un físico y matemático holandés que trabajó en la mejora de los lentes microscópicos, y a Johannes Vermeer, un famoso pintor que probablemente conocía a Leeuwenhoek y se interesó por su trabajo.
En cuanto a opositores y enemigos, no se conocen muchos casos específicos, pero Leeuwenhoek sí tuvo algunas disputas con otros científicos de su época. Uno de ellos fue Robert Hooke, un científico inglés que también trabajó en la observación microscópica. Hooke acusó a Leeuwenhoek de plagiar sus ideas y diseños de microscopios, aunque no hay evidencia concreta de que esto sea cierto. Otro oponente fue Jan Swammerdam, quien estuvo en desacuerdo con algunas de las teorías de Leeuwenhoek y criticó su método de investigación. A pesar de estas disputas, Leeuwenhoek mantuvo una reputación respetada en el mundo científico y fue ampliamente reconocido como uno de los fundadores de la microbiología.
Reconocimientos
Anton van Leeuwenhoek fue reconocido en su vida por diversos miembros de la comunidad científica y por figuras de la realeza. Aunque nunca obtuvo un título académico oficial, en 1680 fue elegido miembro de la Royal Society de Londres, lo que le permitió mantener correspondencia con otros científicos y compartir sus hallazgos. En 1684 fue elegido miembro de la Academia de Ciencias de París.
Leeuwenhoek fue reconocido por los monarcas de su época, y en 1695 recibió una invitación del rey Carlos II de España para visitar la corte. Además, fue honrado por su propia ciudad, Delft, donde fue nombrado consejero y luego presidente del tribunal local. En 1703, se le concedió el título de caballero por parte del rey Guillermo III de Inglaterra.
Después de su muerte, Leeuwenhoek fue recordado como uno de los más grandes microscopistas de la historia. En 1818, el botánico alemán Carl Sprengel acuñó el término "infusorio" en su honor. Además, su trabajo ha sido celebrado en la literatura y en la cultura popular, apareciendo en obras como "Micrographia" de Robert Hooke y "La montaña mágica" de Thomas Mann.
¿Por qué es importante?
El trabajo de Anton van Leeuwenhoek ha tenido un impacto significativo en nuestra vida cotidiana a través de su contribución a la microbiología. Al descubrir y estudiar microorganismos, Leeuwenhoek allanó el camino para el desarrollo de la microbiología moderna, lo que a su vez ha permitido importantes avances en la medicina, la agricultura y la biotecnología.
Gracias a los estudios de Leeuwenhoek sobre los microorganismos, se descubrieron enfermedades causadas por bacterias y otros microorganismos, lo que permitió a los científicos y médicos desarrollar tratamientos efectivos. También permitió el avance de la agricultura, ya que los estudios de Leeuwenhoek sobre las bacterias del suelo ayudaron a comprender cómo se nutren las plantas y cómo se pueden optimizar los cultivos. Además, el trabajo de Leeuwenhoek en la observación de microorganismos permitió avances en la producción de alimentos y bebidas, como la cerveza y el vino, al comprender mejor los procesos de fermentación.
¿Qué debemos aprender de él?
La vida de Anton van Leeuwenhoek es una fuente de inspiración para todos aquellos que buscan lograr grandes cosas en sus vidas. A pesar de ser un hombre común sin educación formal, Leeuwenhoek se convirtió en uno de los científicos más importantes de la historia al dedicarse apasionadamente a la observación de los microorganismos con un microscopio que él mismo construyó.
La vida de Leeuwenhoek nos enseña que la pasión, el trabajo duro y la perseverancia son ingredientes clave para el éxito. A pesar de las limitaciones y obstáculos que enfrentó, como la falta de educación formal y los recursos limitados, Leeuwenhoek siguió adelante con su trabajo, confiando en su propia curiosidad y perseverancia para alcanzar sus metas.
Además, la vida de Leeuwenhoek nos muestra la importancia de la observación y la curiosidad. A través de su trabajo en la observación de los microorganismos, Leeuwenhoek demostró que una mente curiosa y una atención detallada a la naturaleza pueden revelar cosas asombrosas sobre el mundo que nos rodea.
2. Generalidades de la reproducción
2. Generalidades de la reproducción
Al realizar un análisis filogenético de la función de reproducción me he encontrado con un problema, es muy extensa, tanto así que sería imposible tratarla en solo una unidad de menos de 20 capítulos con la profundidad mínima a la que estoy acostumbrado, por lo que he decidido dividir la unidad no en una sino en varias unidades independientes. En consecuencia, la presente introducción sirve de hecho como gran introducción, así como para la unidad de reproducción en unicelulares, protistas y hongos.
2.1 La reproducción celular no implica la función de reproducción
La mitosis (YouTube) o reproducción celular puede ser empleada para mantener la regularidad homeostática de un individuo, por ejemplo, cuando se sufre un corte, las células alrededor experimentan mitosis para recuperar a las que se perdieron. En consecuencia, es conveniente, para estos cuatro capítulos de reproducción, tener en cuenta que la mera reproducción celular no nos permite multiplicar la cantidad de individuos.
Lo anterior es precisamente el objeto de estudio y el objetivo de las adaptaciones que hemos de estudiar, la reproducción de los seres vivos hace referencia a la generación de nuevos individuos aumentando la cantidad de unidades en una población. Reproducir en este contexto es aumentar la cantidad de individuos somáticos. Los individuos somáticos son aquellas formas que se alimentan, y que de una forma u otra deben afrontar la mayor cantidad del peso de la selección natural. En algunas ocasiones esto se da manera relativamente directa, de un individuo somático salen otros más pequeños, o de manera indirecta, de un individuo somático nacen propágalos que luego crecen para generar nuevos individuos somáticos.
En ese sentido la mitosis posee tres funciones completamente diferentes.
(a) Como regenerador, la mitosis sirve para reemplazar células viejas, enfermas o dañadas para asegurar la homeostasis del individuo sin que se generen individuos nuevos independientes.
(b) Como desarrollador, la mitosis permite que la masa del individuo aumente y que sus estructuras se desarrollen, permitiendo a una célula desarrollar tejidos, órganos y sistemas altamente especializados, pero manteniendo la individualidad, es decir, por más que el individuo somático crezca por mitosis, este no se reproduce necesariamente.
(c) Como propagador/multiplicador, la mitosis permite la producción de propágalos que serán liberados al ambiente para colonizar nuevas regiones, estos propágulos pueden ser directos igualitos a su progenitor o estructuras de resistencia. Hay que anotar que la mitosis regeneradora es capaz de completar una reproducción solo si el individuo que ha sufrido el daño puede regenerarse desde fragmentos muy pequeños. Este tipo de reproducción por fraccionamiento y regeneración se denomina reproducción vegetativa.
2.2 Reproducción asexual y selección natural
El proceso de reproducción sexual no involucra la fusión de gametos, sino que descansa sobre la propia mitosis o sus equivalentes en los procariotas. En este sentido se descarta el proceso que discutimos anteriormente, la mitosis puede emplearse para regenerar, para crecer y para reproducirse. La división celular simple, es un modo común de reproducción asexual en los protistas, las plantas y muchos invertebrados. Sin embargo, también pueden ser involucrados procesos mitóticos más complejos, dentro de los cuales podemos referenciar la fragmentación de los cuerpos somáticos o el crecimiento de yemas. Estos procesos asexuales dependen principalmente en los mecanismos reproductivos de la especie o la capacidad regenerativa.
En ocasiones la reproducción asexual es solo una capacidad secundaria del individuo que se desprende de sus potentes capacidades regenerativas y que en condiciones “ideales” no representa una parte vital de su ciclo de vida. La principal ventaja de la reproducción asexual es la velocidad y el mantenimiento del fenotipo al ser una expansión clonal. Si el ambiente mantiene unas condiciones constantes y el organismo es apto para dichas condiciones, la reproducción asexual confiere dos ventajas. La primera es que al ser una reproducción asexual-clonal se mantiene el fenotipo y en consecuencia la aptitud de los descendientes de mantiene en un estado óptimo. El segundo es que la reproducción asexual requiere menos condiciones, como menos energía, menos procesos, menos tiempo y mayor cantidad de propágalos, lo cual permite que las colonias crezcan a gran velocidad (Kutschera & Niklas, 2004).
Haciendo una analogía con la hipótesis del equilibrio puntuado (Eldredge, 2014; Gould, 2014; Gould & Eldredge, 1977) a nivel micro, la reproducción asexual ralentizaría los ritmos evolutivos en la medida que las condiciones ambientales son constantes y favorables.
2.3 Reproducción sexual
A pesar de que la reproducción es una función primordial de los seres vivos, es al mismo tiempo una actividad que no posee efectos positivos en la homeostasis del individuo, es más, puede tener efectos perjudiciales o hasta deletéreos. Cuando las condiciones ambientales se hacen desfavorables muchas especies disminuyen sus capacidades reproductivas, aunque esto no es siempre el caso, muchos protistas ingresan en reproducción sexual o asexual y desarrollan estructuras de resistencia.
Ahora bien, en comparación con la reproducción asexual, la reproducción asexual es más costosa, en términos de energía, requerimientos, condiciones, procesos y tiempo, aun así, es un paso obligatorio en muchos ciclos de vida. Dadas las ventajas de la reproducción asexual, uno puede preguntarse porque la mayoría de los animales, las plantas y muchos protistas hacen más énfasis en la reproducción sexual. La explicación más comúnmente referida para la ubicuidad de la reproducción sexual son las ventajas de la recombinación genética con una perspectiva evolutiva, que se explica por medio de la hipótesis de la reina roja (Brockhurst et al., 2014; Gibson, 2015; Greenspoon & Mideo, 2017; Vergara, Jokela, & Lively, 2014; Zuccala, 2015).
Reproducción y sexo
Generalmente para nosotros los humanos ambas palabras son sinónimos, pero a nivel celular son ciertamente diferentes, pues en la reproducción el punto es aumentar la cantidad de células, mientras que cuando se completa el proceso sexual lo que se hace es fusionar células, y por lo tanto se disminuye. La fecundación/singamia puede ser de dos tipos: (1) plasmogámica y cariogámica en la que se fusionan tanto los citoplasmas como los núcleos de los gametos regenerando un cigoto completamente diploide; (2) plasmogámica y no cariogámica, en esta se fusionan solo los citoplasmas, mientras que los núcleos permanecen tan independientes que incluso inician una mitosis por separado, una célula con dos núcleos haploides es denominada como dicarionte. Cuando la célula dicarionte va a dividirse puede tener uno de dos destinos posibles: (1) que los dos núcleos se mantengan independientes generando una generación dicarionte o (2) que los cromosomas de los dos gametos se reorganicen para formar dos núcleos diploides en lugar de cuatro núcleos haploides, dando lugar a la generación diploide solo hasta que inicia la etapa de dos células.
Los pasos de la reproducción sexual
Nuevamente, para los humanos el termino sexual generalmente es sinónimo del coito, o momento en que los gametos masculinos son dejados al interior del cuerpo de la hembra, sin embargo, tal proceso es solo una adaptación fisiológica que ni siquiera es obligatoria, puede haber sexo sin penetración, es más, puede haber sexo sin que esté involucrado un cuerpo somático, lo peor de todo es que eso es lo más común entre los eucariotas. En biología entendemos por sexo o reproducción sexual la conjunción de los procesos de meiosis y singamia. El proceso general involucra una célula diploide (2n) que al pasar por la meiosis recombina sus cromosomas para aumentar la diversidad genética y en el proceso disminuye sus cromosomas a la mitad, estado llamado haploide. La Singamia es el proceso opuesto, siendo la fusión de dos células haploides que llamamos gametos para revertir al estado diploide. El chiste del proceso radica en la recombinación genética para aumentar la diversidad (Beukeboom & Perrin, 2014; Cavalier-Smith, 1995; Gorelick, 2012; Lee, Ni, Li, Shertz, & Heitman, 2010; Speijer, Lukeš, & Eliáš, 2015).
Hipótesis de la reina roja
La reproducción sexual asegura una alta variación de los propágulos a través del proceso de recombinación genética en la primera fase de la meiosis I. El proceso molecular es al azar, y produce mayor variación sin necesidad de que existan mutaciones SNP, lo cual provoca forzosamente que los propágulos sean todos diferentes de sus ancestros (Brockhurst et al., 2014; Gibson, 2015; Greenspoon & Mideo, 2017; Vergara et al., 2014; Zuccala, 2015).
Esta explicación es complementaria con respecto a la ventaja de la reproducción asexual. Cuando las condiciones ambientales no son favorables el pico de aptitud ya o es el mismo, por lo que es conveniente que los descendientes sean diferentes para que alguna variación pueda, al azar, encontrarse en mejores condiciones para las nuevas condiciones. Debido a que la constante de los ecosistemas es el cambio, la reproducción sexual asegura una constante respuesta a las nuevas condiciones ambientales. Sin embargo, para que esta hipótesis sea satisfactoria, uno debe tomar como ambiente no solo los factores abióticos, sino el nicho completo, todas las relaciones, incluyendo aquellas que son con otros factores abióticos que evolucionan constantemente para afectar negativamente la aptitud de los individuos. Depredadores, competidores y parásitos siempre se adaptan a las defensas nuevas, por lo que se hace imperioso, especialmente para los organismos multicelulares y más complejos, generar nuevas barreras defensivas generación tras generación (Brockhurst et al., 2014; Gibson, 2015; Greenspoon & Mideo, 2017; Vergara et al., 2014; Zuccala, 2015).
2.4 Estrategias reproductivas
Existen en general dos estrategias reproductivas que pueden seguir los individuos, y que se correlacionan con su tamaño y especialización. Estas se denominada la estrategia K y la estrategia r. estos nombres surgen de términos importantes en el modelo de crecimiento logístico exponencial, donde K es la capacidad de carga de un ecosistema y r es la tasa reproductiva de una especie. Adicionalmente esos términos se correlacionan a la tendencia de la curva en dos momentos diferentes del modelo logístico exponencial, siendo la etapa reproductiva donde la variable r es más relevante o siendo la etapa de equilibrio ambiental donde el valor K es más relevante.
Estrategia r
La estrategia r también se denomina como estrategia reproductiva. La idea es simple, producir la mayor cantidad de individuos nuevos que sea posible, aunque estos sean de baja calidad. La supervivencia de estas especies se enfoca en el momento y es a corto plazo. Al ser sus descendientes de baja calidad su posibilidad de supervivencia es baja, por lo que en general cada individuo posee una aptitud darwiniana muy baja, pero a cambio son capaces de evolucionar a gran velocidad.

Figura 2.1. Estrategia de vida r. La mayoría de microorganismos, protistas, artrópodos y muchos vertebrados se pueden clasificar como de estrategia reproductiva r, como por ejemplo las ratas. Otra característica de la estrategia reproductiva r es que sus respectivos individuos adultos son pequeños, débiles y con una expectativa de vida corta.
La estrategia de vida r es útil en momentos en que el ecosistema es colonizable y existe baja competencia, pero a medida que la cantidad de individuos satura los recursos del ecosistema y por lo tanto, la competencia por los recursos limitados se hace más fuerte, esta estrategia se ve limitada por la capacidad de carga del sistema, que no es potro que los límites para alimentarse, o la facilidad con que parásitos o depredadores pueden encontrar a los individuos numerosos de estrategia r.
Estrategia K
La estrategia de vida K es una estrategia de vida a largo plazo, esta emerge pensada en sistemas con condiciones estables y constantes, donde lo que es importante es adaptarse a las condiciones del sistema. Estas especies producen pocos individuos, pero a cambio son mayor calidad, más fuertes y, en consecuencia, más capaces de soportar las tensiones de la selecciona natural causadas por una población habitando un ecosistema en equilibrio, donde competidores, depredadores y parásitos habitan y se interrelacionan.

Figura 2.2. Modelo de crecimiento logístico exponencial. Los seres vivos pueden basar la supervivencia de su especie reproduciéndose rápido (estrategia r) o teniendo vidas largas y fuertes para adaptarse al ambiente a largo plazo (estrategia K). Estas se relacionan con el modelo de crecimiento logístico, un modelo matemático que describe el crecimiento de una población (n) en el tiempo (t) hasta ser equilibrado por fuerzas del ecosistema que llamamos selección natural. La curva se divide en dos momentos, la etapa reproductiva y la etapa de equilibrio.
Los seres humanos somos una particularidad ya que al interior de una misma población puede haber familias con una estrategia de vida r en la cual hay muchos hermanos, pero en los que se invierte poco cariño y esfuerzo, o familias con una estrategia de vida K en la que se tienen uno o dos hijos en los que se invierte mucho.

Figura 2.3. Estrategia de vida K. Solo los seres vivos más grandes y complejos descansan en una estrategia de vida K, por lo que ejemplo de esto son los grandes árboles que producen pocas semillas o los animales como los elefantes o incluso, los seres humanos.
Las consecuencias éticas de cada estrategia de vida son solo aplicables al ser humano en el sentido de que somos la única especie capaz de elegir a conciencia cual estrategia reproductiva seguir para nuestra propia familia. En términos generales una estrategia de vida r limita la cantidad de atención, cuidados y recursos se distribuyen entre cada hijo, mientras que en una estrategia K se arriesga todo a un único o dos descendientes.
3. Historia de vida
3. Historia de vida
Los seres vivos nacen, crecen, se reproducen y mueren, la definición más inocente de ser vivo es en realidad una descripción básica de la historia de vida. La historia de vida es toda descripción que podemos hacer de las etapas sucesivas por las que pasa un ser vivo para completar su función reproductiva, y en general toma la forma de los ciclos de vida.
3.1 Ciclos de vida
Los ciclos de vida son modelos que nos permiten representar por medio de dibujos procesos y estructuras, de forma tal que podemos comprimir una gran cantidad de información acerca del desarrollo e historia de vida de los seres vivos. Como su nombre implica, al ser ciclos debemos comenzar y terminar en un mismo punto, describiendo los principales procesos de la historia de vida de una especie en particular.
El estudio de los ciclos de vida representa en grado sumo, el estudio de la función de reproducción de los seres vivos, y no solo involucra el estudio de las estructuras reproductoras, sino de todas aquellas adaptaciones que le permiten a las especies propagarse y prosperar. Los ciclos de vida son particularmente importante en el estudio de las enfermedades infeccionas, ya que al conocer el ciclo de vida de un patógeno, es posible señalar los pasos cruciales donde es posible detener su propagación, disminuyendo así la virulencia del patógeno.
Por lo general hay dos estructuras básicas en todo ciclo de vida, el cuerpo somático y el propágulo. El cuerpo somático es aquella estructura que interactúa con el medioambiente desarrollando las demás funciones de un ser vivo, como alimentarse y relacionarse con su medio ambiente, en ultimas es aquella estructura que afronta el mayor peso de la selección natural. Por el contrario, el propágulo es una estructura de reproducción y colonización, incluso es una estructura de resistencia. Su función es la de aumentar la cantidad de individuos de forma tal que pueden regenerar por si mismos un individuo somático completo.
Los cuerpos somáticos y sus correspondientes propágulos pueden ser tanto haploides como diploides, y organizarse de maneras complejas a lo largo de los diferentes ciclos vitales. Por ejemplo, en el ser humano, los propágulos no pueden generar por sí mismos un nuevo individuo, sino que deben fecundarse sexualmente. Adicionalmente los propágulos no portan el mismo contenido genético ya que son producidos a través de meiosis. Sin embargo, para aclarar mejor que son los ciclos de vida y cómo funcionan, mejor comenzaremos su estudio por los dos más simples, el ciclo de vida asexual teórico y el ciclo de vida eucariótico completo que también es conocido como alternancia de generaciones.
3.2 Ciclo de vida asexual
El ciclo de vida asexual se caracteriza porque los propágulos, es decir todas las partículas reproductivas que en teoría deben producir más individuos nuevos y completos, poseen la misma carga genética completa. Si el cuerpo somático es diploide sus propágulos serán diploides, pero si el cuerpo somático es haploide sus propágulos serán haploides. Es raro encontrar especies que dependan únicamente de la reproducción por ciclo de vida asexual en los eucariotas, pero algunos hongos y protistas han perdido su capacidad sexual en favor de una capacidad de colonización acelerada.

Figura 3.1. Ciclo de vida asexual. El ciclo de vida asexual tiene dos subciclos, el menor o mitótico y el mayor en el que se generan esporas asexuales, las cuales normalmente son estructuras de resistencia.
En términos generales el ciclo de vida asexual posee dos alternativas, la primera y más compleja involucra a una célula vegetativa (1) que al enfrentar condiciones adversas ingresa en un proceso de formación de propágulos de resistencia (2) que se denominan esporas o quistes (3), las cuales al encontrar mejores condiciones se reactivan y por bipartición (4) pueden reconstruir una población de células vegetativas. A su vez la segunda ruta de la reproducción asexual es mucho más simple, y radica en el incremento de individuos unicelulares por bipartición (5).
3.3 Ciclo de vida de alternancia de generaciones
Existen varias formas de ciclos de vida sexuales, pero el más completo y complejo, y posiblemente el que nos sirve mejor para entender todos los demás ciclos de vida es el de alternancia de generaciones.
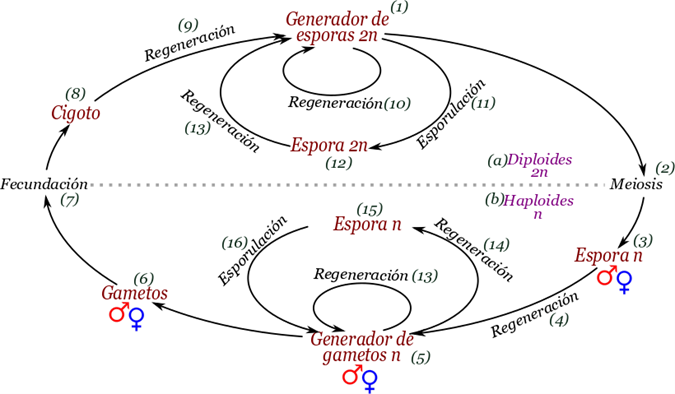
Figura 3.2. Ciclo de vida sexual con alternancia de generaciones. El ciclo de vida sexual completo tiene tres subciclos, el ciclo mayor o propiamente sexual, y dos subciclos asexuales que regeneran dos cuerpos diferentes, uno con células diploides denominado generador de esporas y otro con células haploides llamado generador de gametos. Algunos seres sexuales carecen del cuerpo generador de gametos o del cuerpo generador de esporas.
El ciclo de vida de alternancia de generaciones inicia con un cuerpo somático diploide generador de esporas (1). Este cuerpo especializa algunas células para realizar la meiosis (2) generando las esporas sexuales (3). Dichas esporas germinan y por medio de la mitosis en cualquiera de sus variedades regenera (4) un cuerpo somático haploide generador de gametos sexuales (5). El cuerpo generador de gametos puede estar dividido en dos sexos diferentes o ser hermafrodita, lo importante es que genera gametos sexuales (6) los cuales pueden ser anisogámicos o isogámicos. Para la representación esquemática sumimos que son gametos anisogámicos en los que podemos diferencia un gameto masculino o esperma y un gameto femenino u óvulo.
Los gametos se unen por medio de la fecundación que puede ser singámica, que es la unión de los citoplasmas formando un dicarionte o singámica y cariogámica y en tal caso el cigoto (8) sería un diploide. Ahora bien, el cigoto por cualquiera de los mecanismos mitóticos regenera (9) el cuerpo generador de esporas completando el ciclo sexual mayor (1).
A parte del ciclo mayor sexual, en teoría cada cuerpo somático debería ser capaz de realizar dos ciclos menores o epiciclos asexuales, ya sea para incrementar su número de individuos o para regenerar partes pérdidas por depredadores, este proceso puede llevarse a cabo por diferentes formas mitóticas y por lo tanto la denominamos simplemente como regeneración (10, 13). Sin embargo el epiciclo asexual también puede involucrar propágulos de resistencia generados por el proceso de esporulación o enquistado (11, 14) que genera una estructura reproductiva de resistencia y dormancia denominada espora o quiste (12, 15). Cuando la estructura de resistencia llega a condiciones favorables puede regenerar (13, 16) al individuo que la produjo sin recombinación genética (1, 5).
Hay que destacar que el ciclo que mostramos anteriormente es solo una aproximación teórica, en la naturaleza pode poseer simplificaciones, por ejemplo que el cuerpo generador de gametos sea incapaz de generar esporas asexuales, o que exista una tercera generación a parte de la generación haploide n (b) y la generación diploide 2n (a) llamada dicarionte. En el haploide solo hay un cromosoma de cada tipo, mientras que en el diploide hay dos cromosomas de cada tipo en un solo núcleo. Un dicarionte puede tener dos núcleos haploides no fusionados. Inclusive podemos tener estados más exóticos, como el dicarionte haploide/diploide en los ciliados, donde hay un núcleo haploide y otro diploide, o células multicariontes donde hay incontables núcleos.
3.4 Metamorfosis y etapas de desarrollo
En ocasiones el cuerpo somático no mantiene una estructura determinada a lo largo de la historia de vida de una especie, sino que sufre modificaciones, de forma tal que el cuerpo se adapta para diferentes funciones biológicas, y cada uno de esos cuerpos diferentes es separado por un evento de metamorfosis, cambio de piel o desarrollo de órganos. Por lo general podemos distinguir dos etapas, la etapa de crecimiento y la etapa madura.

Figura 3.3. Metamorfosis y desarrollo. Los cambios en el desarrollo muestran la enorme plasticidad del cuerpo de los seres vivos, y son uno de los aspectos que más se modifica en el desarrollo evolutivo.
La etapa de crecimiento, inmadurez se caracteriza por adaptaciones enfocadas a la alimentación, el organismo se adapta principalmente para alimentarse, crecer y desarrollarse, acumulando nutrientes necesarios para afrontar las siguientes etapas. La segunda etapa es denominada de madurez, pero dicha madurez se define en términos de la capacidad de reproducción del organismo, es una madurez sexual. Una vez que el organismo alcanza un tamaño crítico cambia y se hace capaz de reproducirse. Algunas especies poseen más de una sola etapa de inmadurez que se van alternando hasta llegar al individuo maduro. Por el contrario, otras especies solo tienen dos formas diferentes que se separan una de otra por el evento de metamorfosis. En otras las dos etapas apenas si pueden diferenciarse por el tamaño, y se entienden más por el desarrollo de los órganos reproductivos.
Aunque indudablemente toda especie debe pasar por transiciones estructurales en su historia de vida, algunas son más drásticas que otras. Cuando el juvenil tiene más o menos la misma forma del adulto decimos que el organismo posee un desarrollo directo. Pero si hay un momento de cambio drástico mediado por una metamorfosis, decimos que hay un desarrollo indirecto.
3.5 Partenogénesis
Es una estrategia reproductiva especial en la cual un ovulo no fertilizado puede desarrollar un individuo completo, y un ovulo fertilizado también. Las especies partenogenéticas son conocidas desde los invertebrados has vertebrados tan recientes como los lagartos. En la mayoría de las especies estudiadas, la partenogénesis es solo un modo de asegurar la procreación en casos en los que es complicado que los dos sexos opuestos se encuentren en el mismo lugar como en algunas especies de lagartos. Sin embargo, algunos invertebrados la partenogénesis se ha integrado de forma completa al ciclo de vida, permitiendo la diferenciación sexual. Por ejemplo, en las abejas, las hembras se producen por fecundación sexual típica, mientras que los machos son partenogénicos, y se generan solo cuando la reina decide que es momento de producirlos. Esto se debe a que la hembra recibe esperma solo durante el vuelo nupcial, y es capaz de almacenarlo en receptáculos especializados. Si decide que es momento de que existan machos, simplemente produce óvulos sin abrir las cámaras de esperma.

Figura 3.4. Partenogénesis. Algunos animales como los rotíferos emplean la partenogénesis como un mecanismo de propagación asexual hasta que las poblaciones lo bastante grande, y el ambiente lo bastante desventajoso como para inducir la aparición de machos y en consecuencia completar el ciclo sexual.
especies que dependen exclusivamente de la partenogénesis para su reproducción son raras, y se encuentran usualmente en ambientes altamente aislados, lo cual las libera parcialmente de lagunas presiones selectivas, sin embargo, pueden ser altamente vulnerables a especies invasoras con reproducción sexual, ya sean competidores, depredadores o parásitos zoonóticos. Sin embargo, aún es materia de debate si todas las especies partenógenas son igual de vulnerables, o si, por el contrario, poseen algún mecanismo de variabilidad desconocido que les permite lidiar con las presiones impuestas por los parásitos, depredadores y competidores.
3.6 Machos, hembras y otras cosas
La definición de macho y hembra puede aplicarse a un cuerpo somático haploide o diploide, pero se define en mase del concepto de anisogamia. La anisogamia es que los gametos sexuales que van a experimentar la plasmogamia poseen formas diferentes. Por lo general uno es más pequeño y se denomina microgameto y otro es más grande y se denomina macrogameto. En caso de que los microgametos deban moverse desde su lugar hacia donde está el macrogameto, generalmente por flagelos, los denominamos espermatozoides o espermas y el macrogameto lo llamamos óvulos.
Un cuerpo somático masculino produce espermas, uno femenino produce óvulos y uno hermafrodita produce ambos. En caso de que en la especie el cuerpo somático principal esté diferenciado en uno creador de gametos masculinos y otro formador de gametos femeninos como en el ser humano, decimos que la especie es dioica o dioecia, pero si hablamos de un hermafrodita decimos que tratamos con una especie monoica.
Sin embargo, nuevamente todo eso descansa el que los gametos no sean iguales, es decir de la anisogamia. Existen otros gametos que son isogámicos, son iguales los conjugantes plasmogámicos, por lo que los modelamos como compatibles con los símbolos (+) y (-) aunque las razones de conmutabilidad pueden ser bastante complejas y estar determinadas por complejos receptores de membrana y marcadores genéticos muy precisos que van más allá del mero tipo de cromosomas.
4. Bacterias y arcaicas
4. Bacterias y arcaicas
Los conceptos de haploide, diploide, y especialmente el de recombinación sexual no aplica para los procariotas. La estructura genética de los procariotas es diferente al de los eucariotas. Por lo general el ADN de los procariotas se encuentra almacenado en un único cromosoma circular que no está empaquetado de forma estricta ya que se encuentra transcribiendo “manifestando de forma práctica la información que almacena” la mayoría del tiempo.
4.1 Ciclo de vida
El ciclo de vida de las bacterias inicia con la célula vegetativa (1) la cual puede ser un individuo flotando en una suspensión acuosa o estar atada a un grupo de bacterias unidas por una matriz extracelular en una bioplaca o biopelícula.

Figura 4.1. Ciclo de vida de bacterias y arcaicas. El ciclo de vida de los procariotas es típicamente asexual, sin embargo, las bacterias deben entenderse en términos de colonias, más que de células individuales, con diferentes grados de expecialización.
Estas bacterias pueden experimentar dos ciclos, el ciclo asexual de bipartición, en la cual una célula se divide en dos por medio de la bipartición bacteriana (5) “que no debe confundirse con la mitosis por muchas diferencias en el proceso” y sirve para incrementar la cantidad de individuos. Adicionalmente en (1) las células vegetativas experimentan todas sus recombinaciones genéticas ya sea por conjugación, transducción o transformación sin alterar la cantidad de cromosomas bacterianos.

Figura 4.2. La bipartición. A pesar de sus similitudes, la fisión binaria es un proceso diferente de la mitosis, pues al no haber cromosomas, el proceso involucra pasos diferentes, y es mucho más rápida.
La segunda opción para el ciclo de vida de los procariotas bacterianos es la esporulación (2). En este se genera una estructura de resistencia denominada espora (3) en la cual hay un propágulo que se mantiene en una estasis metabólica y está protegido por más capas. Cuando las condiciones ambientales mejoran, la espora se reactiva y los individuos que emergen regeneran (4) por bipartición bacteriana a la población vegetativa (1), sea esa una población de células en suspensión o insertadas en una biopelícula.
4.2 La fisión binaria bacteriana por bipartición
En parasitología los términos fisión binaria y mitosis parasen ser altamente sinónimos, así que emplearemos el termino bipartición para tratar de aislar y diferenciar el mecanismo de reproducción en las bacterias. Y sí, evidentemente para entender de lo que estamos hablando tenemos que hacernos a la idea de un concepto fundamental, las bacterias no se reproducen por mitosis ya que no hay núcleo, ni huso mitótico, el cromosoma no posee centriolo y un largo etc. Es por esto que la bipartición bacteriana se denomina como la fisión binaria. Por lo general el ciclo de vida simplificado de los procariotes se describe como en la figura anterior, dando la impresión de que la síntesis de un nuevo anillo cromosomal se da al final de la citocinesis. De hecho, de manera simultánea también se está replicando, y lo vuelve hacer inmediatamente se divide en dos, lo cual hace que aun cuando la célula no se ha dividido totalmente su material genético ya se encuentra a medio camino de copiarse nuevamente.

Figura 4.3. Duplicación cromosomal compleja. En realidad, la síntesis del anillo cromosomal es más compleja, en b se muestra cómo se reproduce un cromosoma en una bacteria en fase de crecimiento exponencial, donde aún no ha acabado la síntesis de un nuevo anillo, cuando ya otros dos ya vienen en camino por cada una de las dos hebras originales que se estaban formando.
Evidentemente este afán replicador hace que el ciclo de vida de los procariotas sea mucho más rápido que el de cualquier eucariota. A una bacteria promedio le toma entre 20 y 30 minutos completar una división celular en comparación con las 20 horas que requiere una célula animal o vegetal de reproducción relativamente continua. Lo anterior implica que una sola bacteria podría cubrir la faz de la Tierra en dos días con sus descendientes, mientras que a una célula eucariota le tomaría alrededor de dos meses. Obviamente, las limitaciones ambientales y ecológicas limitan siempre la población de cualquier especie. De lo anterior se desprende que la noción de ciclo celular en los procariotas es más simple y rápida, las fases de crecimiento y síntesis son simultáneas, y la fase de crecimiento se demora lo que tarda en completarse una nueva síntesis.
4.3 Detalles de la bipartición
La reproducción celular de los procariotas se denomina fisión binaria por bipartición, la cual se clasifica como un mecanismo de reproducción asexual “no genera meiosis, fecundación o fases haploide y diploide”. Esto implica que durante la reproducción el único mecanismo de variabilidad de los procariotas es la mutación aleatora. Para solucionar esto los procariotas emplean mecanismos de recombinación que algunos autores denominan parasexualidad o pseudo-sexualidad.

Figura 4.4. La bipartición. El proceso se puede dividir en las siguientes partes: (1) Replicación del ADN, está ya viene a medio camino procedente del ciclo anterior, las bacterias duplican su genoma mucho más rápido que los eucariotas. (2) la fase de crecimiento celular llega a su cúspide incrementando el volumen de la bacteria. (3 y 4) la citocinesis separa la célula de gran volumen en dos células pequeñas iniciando la fase de crecimiento, en esta fase el genoma inicia su duplicación para las siguientes dos generaciones, de modo tal que cuando se dé la citocinesis completa (5) el genoma ya vaya a medio camino de la duplicación.
La fisión binaria es relativamente simple en comparación con la mitosis o la meiosis, en ella el material genético se inicia a duplicar antes de que se formen dos células hijas de la generación anterior. Luego cuando la célula ya se ha dividido la célula cuenta con cierta parte de su material genético duplicado. Una vez el material genético está en una cantidad doble la célula inicia la citocinesis, y al mismo tiempo el material genético en los polos de la célula inicia su siguiente fase de síntesis. Por lo general los modelos representan este mecanismo de forma simplificada omitiendo el verdadero punto de inicio de la síntesis de material genético.
Un detalle importante es que el ciclo celular bacteriano no diferencia etapa G ni etapa S, La fase de crecimiento y la fase de síntesis se dan de forma paralela y simultánea lo que asegura que al momento de iniciar la citocinesis una nueva replicación celular da inicio (De Duve & Pizano, 1995).
4.4 División instantánea en procariotas
Este tipo de división es típica de las bacterias con una pared celular relativamente gruesa como los bacilos Gram+. En ella al interior de la pared se forma una nueva en la mitad de la célula generando una pared doble en el punto de unión.
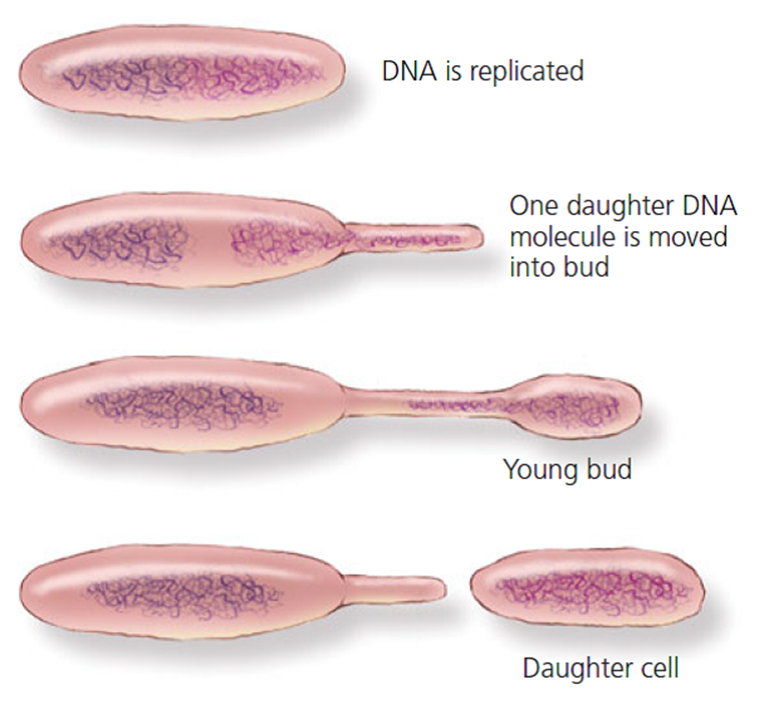
Figura 4.5. La gemación o brote. En el brote una parte de la célula madre experimenta un crecimiento parcial, como una burbuja que emerge o una hinchazón parcial, la cual recibe material celular y material genético mínimos para que una vez completada la citocinesis la célula hija pueda operar por sí misma.

Figura 4.6. Endosporas. Las endosporas constituyen un mecanismo de defensa contra condiciones hostiles o desfavorables.
En la división instantánea la pared celular se forma completamente antes de que las células hijas se separen, por lo que cuando las células hijas se separan lo hacen como si una célula madre se partiera a la mitad de manera instantánea. Con el crecimiento del material del citoplasma se crea una tensión que provoca que las células se dividan de manera instantánea justo en el punto más débil de la pared celular vieja. Las células hijas permanecerán pegadas e incluso con remanentes de la pared celular de la célula madre rota en el punto de fractura.
4.5 Esporulación y fragmentación bacteriana
Los procariotas pueden tener otros métodos o “variaciones” de la fisión binaria. Por lo general se trata de divisiones en la que la célula madre retiene su identidad durante y después de la división celular. Los actinomicetes por ejemplo producen unas células reproductivas en estado de resistencia denominadas esporas al final de sus células filamentosas. Cada espora tiene el potencial de generar un nuevo individuo una vez que encuentran las condiciones adecuadas para su crecimiento y proliferación. Algunas cianobacterias se reproducen por fragmentación produciendo filamentos móviles de se arrastran lejos del lugar de reposo de la célula ancestral para buscar nuevos territorios para colonizar.
4.6 Reproducción de procariotas por brote
Al igual que la reproducción por fragmentación y esporulación, en la reproducción por brote la célula madre mantiene su identidad durante y después de la división celular.
5. Endosporas bacterianas
5. Endosporas bacterianas
Algunas bacterias como las pertenecientes a Bacillus y Clostridium son reconocidas por la habilidad de formar estructuras únicas llamadas endosporas, las cuales son importantes por una gran variedad de razones, incluyendo su durabilidad y la potencial capacidad para inducir patogenicidad. Algunos autores prefieren llamarlas esporas, pero las endosporas no deben confundirse con las esporas reproductivas que ya hemos discutido anteriormente como las de las actinobacterias.
En la microfotografía anterior se puede ver que las endosporas son como raquetas, la razón de esto se entenderá cuando describamos el proceso de la esporulación. Una bacteria con metabolismo activo se denomina cuerpo vegetativo o célula vegetativa y se puede distinguir de una endospora debido a su actividad metabólica. A su vez las esporas reproductivas y las endosporas se distinguen debido a que las esporas reproductivas son producidas en grandes cantidades mientras que una endospora es una estructura única, una célula solo puede generar una endospora. Por lo tanto, las endosporas no son estructuras reproductivas, ¿Por qué analizarlas en la reproducción celular?, porque el proceso de la formación de una endospora es una variación de la fisión binaria.
5.1 Esporulación
Una célula vegetativa normalmente inicia su conversión a una endospora solo cuando las condiciones del ambiente se hacen adversas o cuando los recursos como fuentes de carbono o nitrógeno se hacen escasos. El proceso de formación de endosporas se denomina esporulación, y requiere entre 8 y 10 horas para completarse.Esencialmente inicia como una bipartición normal en la que la célula posee dos copias del material genético, pero no ocurre citocinesis, en su lugar una de las dos copias del material genético es destruida mientras que la otra es rodeada por una barrera de dos membranas y una pared gruesa de peptidoglicano.

Figura 5.1. Esporulación. La esporulación es semejante a la bipartición pues ambas inician con un proceso de duplicación del ADN, pero hasta allí van las diferencias. En la formación de endosporas uno de los materiales genéticos generados es destruido, mientras que el otro es protegido por varias capas y es deshidratado para soportar condiciones muy hostiles.
Una vez formada la barrera interna el citoplasma encerrado es despojado de la mayor cantidad de agua posible, mientras es suplementado con materiales de reserva. Como se dijo anteriormente la célula no experimenta citocinesis, con el tiempo la perdida de agua se hace extensiva al resto del citoplasma, haciendo que la membrana celular externa se adicione a la barrera como una capa protectora extra.
Dependiendo de la especie una célula que forma una endospora puede generarla en la parte central, en la parte subternimal “cerca de una punta” o en una región apical “en la punta”.
5.2 Capacidades de las endosporas como estructuras de resistencia
Las endosporas son extremadamente resistentes a la desecación, el calor, la radiación ultravioleta y los químicos letales. Por ejemplo, permanecen con vida en el agua hirviendo por varias horas: no son afectadas por los desinfectantes como el alcohol, el peróxido de hidrogeno, el blanqueador y otros químicos letales. Pueden permanecer con vida después de ser irradiadas con 400 rad, lo cual es más de 5 veces la cantidad necesaria para matar a un ser humano adulto.

Figura 5.2. Esporas de clostridios. Varias bacterias relacionadas con Clostridium tienen variación en la formación de sus esporas y su reproducción celular llegando a un híbrido donde las células hijas se forman al interior de la célula madre antes de que esta última explote dando a luz a su progenie.
Las endosporas son estructuras de resistencia que prácticamente no poseen metabolismo activo, se encuentran en un estado de animación suspendida esperando a que las condiciones del ambiente se tornen favorables. Los mecanismos de resistencia que les permiten a las endosporas sobrevivir condiciones tan adversas aún son desconocidas, pero aparentemente se debe a la presencia de una barrera compuesta por varias membranas y una pared celular gruesa, así como otros mecanismos de estabilización molecular como uniones de calcio y proteínas de fijación del material genético
5.3 Impactos de la habilidad para formar endosporas
La habilidad para sobrevivir condiciones difíciles hace de las endosporas las estructuras celulares más resistentes. Por ejemplo, las endosporas de Clostridium selladas en tubos de ensayo pueden sobrevivir con facilidad por 34 años. Este récord es pequeño en comparación con el reporte de haber reanimado endosporas de Bacillus al interior de un cristal de sal de más de 250 millones de años de antigüedad en un yacimiento cerca de Carlsbad en Nuevo Méjico. Otros cuestionan esta afirmación argumentando que puede ser contaminantes secundarios que ingresaron en el cristal a través de grietas microscópicas. En cualquier caso, existen pocas dudas de que las endosporas permanecen viables por periodos mínimos de décadas hasta milenios.
La formación de endosporas en una preocupación seria de los productores y procesadores de alimentos, así como de los prestadores de servicios de salud relacionados con la alimentación y la digestión. Las endosporas son resistentes a la mayoría de los mecanismos empleados para el control de otros microbios como el blanqueador, el alcohol, la radiación. Lo peor es que las endosporas son capaces de producir toxinas fatales como en el caso del ántrax, el tétano y la gangrena.
5.4 Variaciones de la formación de endosporas
Una variación de la formación de endosporas como mecanismo reproductivo es la que lleva a cabo Epulopiscium, una bacteria gigante simbiótica del pez cirujano, y muchos de sus parientes cercanos. En estas las barreras internas no forman endosporas, sino endocélulas, las cuales crecen rompiendo a la célula madre desde el interior.
Aproximadamente 12 células hijas son producidas a partir de una célula madre, y a este mecanismo se lo conoce como viviparismo en bacterias.
5.5 La espora en el zapato de Pasteur
A pesar de que la tradición dice que Luis Pasteur “1822-1895” refutó contundentemente a la generación espontánea mediante su trabajo con los frascos de cuello de cisne en 1864 (Cloutier, 1995), la verdad es que las esporas le jugaron una mala pasada haciendo que la historia es un poco más complicada que eso. La generación espontánea era vista como una hipótesis científica aceptable por miembros eminentes de la comunidad científica de la época (Benton, 1974). En cualquier caso, el experimento de Pasteur permitió eliminar varias de las hipótesis sobre la generación espontánea.
Para el caso de la hipótesis de que el principio vital (Benton, 1974; Henderson, 2012) era demasiado denso para atravesar un filtro de tela, Pasteur no utilizó gazas para bloquear el frasco, sino que lo dejó abierto con una modificación, el cuello de la botella tenía forma de un cuello de cisne. El frasco solo experimentó crecimiento si se rompía el cuello de cisne dejando un cuello recto, o girando el frasco de forma tal que el líquido de cultivo tuviera contacto con la parte más baja del cuello de cisne. De esta manera Pasteur evitaba las afirmaciones sobre el “tamaño de la fuerza vital”. El mismo procedimiento también evitaba la hipótesis de que el sobrecalentamiento de los nutrientes afectaba a la fuerza vital, aun cuando el caldo de cultivo era vigorosamente esterilizado, aun permitía el crecimiento de microorganismos cuando el cuello giraba o era roto.
Figura 5.3. Cuello de Cisne. El frasco de cuello de cisne fue empleado por Pasteur para poner a prueba el principio de la fuerza vital, aunque muchos piensan que fue un experimento clave, el problema radicaba en que los organismos esporulantes debieron ser ignorados.
La estructura del cuello del cisne permitía que el aire ingresara al frasco, pero que el material particulado se terminara decantado en la parte inferior del cuello de cisne, lo cual invalidaba el principio aristotélico de una fuerza vital transportada por el aire. Sin embargo no todos estuvieron satisfechos con esto, de hecho el experimento de Pasteur fue seguido por otros trabajos como los de John Tyndall “1820-1893” (Strick, 2009a, 2009b) quien tuvo problemas con la esterilización vigorosa. Esto se debía a la existencia de las esporas termoestables “desconocidas para él y para todo el mundo”, pero que fueron del mismo modo una hipótesis auxiliar que el propuso para proteger el núcleo fuerte de la propuesta de Pasteur de la biogénesis “toda vida proviene de una vida previa”, en otras palabras se generó una hipótesis ad hoc para salvar a la biogénesis sin tener evidencia directa en forma de la identificación o aislamiento de la espora, se trataba entonces de una predicción basada en información indirecta y razonamiento inductivo. Por suerte para el programa de investigación de la biogénesis, la hipótesis auxiliar de las formas resistentes al calor fue corroborada en años posteriores por Ferdinand Cohn “1828-1898” (Drews, 2000) con su descripción de las esporas en 1876 -más de diez años después de los experimentos de Pasteur –con lo cual se ponía punto final a la generación espotánea.
6. Pseudosexualidad
6. Pseudosexualidad
A pesar de que las bacterias carecen de la reproducción sexual, y por lo tanto de la posibilidad de recombinar mediante cromosomas homólogos, las bacterias pueden transferir genes unas a otras incluso entre linajes muy diferentes entre sí mediante lo que se ha denominado transferencia horizontal de genes. La transferencia horizontal de genes permite la recombinación de genes, y por lo tanto cumple una función homóloga a la reproducción sexual. El proceso de transferencia horizontal de genes puede darse por tres mecanismos que solo enunciaremos en el presente artículo, pero que serán tratados con total profundidad en temas futuros concernientes al genoma de las bacterias:
(a) Conjugación: mediante plásmidos y pilis una bacteria mutante puede alterar a otra.
(b) Transducción: algunos retrovirus transmiten genes de unas bacterias a otras permitiendo su recombinación.
(c) Transformación: una bacteria mutante muerta puede transferir su material genético a las que la rodean, estas absorben el material genético y lo integran al propio recombinando.
6.1 Conjugación
Es la tranferencia horizontal de material genético entre células bacterianas mediante el contacto directo célula a célula o por un contacto mediado pro estructuras especializadas. La principal diferencia con los otros mecanismos de transferencia horizontal de genes es que en la conjugación es necesario un contacto íntimo célula a célula. Dado lo anterior algunos autores plantean que la conjugación es el equivalente bacteriano de la reproducción sexual ya que involucra el contacto entre individuos con un patrimonio genético diferente, pero allí paran las diferencias.

Figura 6.1. Conjugación.
Otra de las características principales, y quizá la más importante para tener en la mente de cualquier estudiante de microbiología es que la conjugación involucra estructuras especializadas denominadas plásmidos, los cuales se pueden visualizar como cromosomas bacterianos en miniatura. Al igual que el cromosoma principal, los plásmidos son estructuras hechas de ADN con forma circular. Sin embargo los plásmidos no son lo único que puede transferirse durante una conjugación, ya que fragmentos de genes saltarines llamados transposones también pueden ser transferidos con independencia a un plásmido (Christie, 2016; Clewell, 2013). La mayoría de los plásmidos conjugativos poseen sistemas que aseguran que la célula receptora no contenga ya un elemento semejante.
La información genética transferida por lo general es benéfica para la célula receptora ya que contiene genes relacionados con el manejo de metabolitos secundarios externos como los antibióticos, y los xenobióticos, lo cual le permite a la nueva cepa adaptarse y sobrevivir a las nuevas presiones de selección, sean estas naturales o fabricadas por el hombre (Christie, 2016; Clewell, 2013).
El proceso de la conjugación (YouTube) involucra estructuras llamados pilus, los cuales actúan como receptores de membrana bastante grandes que reconocen a una bacteria que puede recibir el nuevo material genético. Una vez que se establece la conexión el pilus asegura que la membrana y la pared de ambas células se relajen permitiendo que parte del citoplasma de las dos células entren en contacto. En ese momento el plásmido se copia de forma tal que la hebra hija es transferida a la célula receptora, allí esta hebra replica su complemento e inicia la síntesis de sus respectivos genes.
6.2 Transducción
Es uno de los tres procesos de transferencia horizontal de genes, en el cual el ADN foráneo es transferido de una célula a otra por medio de retrovirus (YouTube).

Figura 6.2. Transducción. La transducción viral también puede darse en los eucariotas, siendo una forma menor de transferencia de genes entre diferentes linajes.
Hay que destacar que este mecanismo de transferencia horizontal de genes también es presentado por las células eucarióticas. La transducción no requiere un contacto íntimo entre células.
Es un mecanismo importante que aumenta la diversidad genética y no se restringe a los procariotas (Bannert & Kurth, 2004; Chalopin et al., 2014; Naville et al., 2016; Temin, 1985; Volff, 2006). Familias de genes como los de las globinas pueden ser explicadas como transposones y retrotransposones. La retrotrasnposición da origen a la copia del gen, sin que se requiera que todo el genoma se duplique. Posteriormente uno de los locus puede transponerse a otro cromosoma, y allí volver a copiarse por retrotransposición.
Esto convierte a los genes que pueden copiarse y moverse en entidades con cierta individualidad, de hecho, en las globinas dicha individualidad puede verse en que todos esos loci mantienen una estructura común. El hecho de que muchos virus denominados retrovirus puedan realizar proezas semejantes ha permitido generar hipótesis sobre la importancia de los virus en la evolución biológica, ya que sin la enzima transcriptasa reversa, muchas familias de genes no hubieran podido surgir.
Lo anterior implica que al menos los retrovirus al donar la enzima transcriptasa reversa han servido como parte del verdadero motor evolutivo que es la variación aleatoria, ya que al permitir una copia de genes más fácil, aceleran el potencial para la creación de nuevos rasgos al interior de los seres vivos. Personalmente, a diferencia de lo que proponen autores como Máximo Sandin, esto es perfectamente concordante con el estema de la síntesis evolutiva moderna, después de todo los loci nuevos creados por la retrotransposición deben medirse ante la selección natural.
6.3 Transformación
La transformación es el proceso de absorber material genético fragmentado a través de la membrana celular, en este proceso el donante debe morir y liberar su ADN al medio donde los receptores lo captan.

Figura 6.3. Transformación. En la transformación se transfiere ADN de una célula muerte y rota a otra célula viva.
Para que la transformación pueda ocurrir la bacteria receptora debe encontrarse en un estado de competencia, el cual es un estado poco común inducido por factores de estrés, y puede ser inducida en condiciones de laboratorio (Johnston, Martin, Fichant, Polard, & Claverys, 2014).
6.4 Fragmentación de una biopelícula
Las bacterias oscilan entre dos modos de vida, el de individuos disueltos en el medio acuoso y el de bioplacas. Las biplacas se forman cuando las condiciones se hacen adversas por medio del sistema de sensibilidad de densidad poblacional (Speziale & Geoghegan, 2015). En una bioplaca las bacterias se encuentran aisladas del medio externo por medio de la secreción de una sustancia viscosa y pegajosa que se denomina por homología o analogía como matriz extracelular MEC(Borlee et al., 2010; Bowen & Koo, 2011; Hawser, Baillie, & Douglas, 1998). En este sentido, la matriz extracelular funciona como el fluido interno al interior del cual los mensajeros químicos son segregados de forma interna, cumpliendo con la definición de la función endocrina. Las bacterias que generan biopelículas generalmente también poseen en sistemas de sensibilidad de densidad poblacional, por lo que segregan químicos mensajeros que coordinan sus esfuerzos para colonizar un ambiente.
El punto con esta breve introducción es que una biopelícula puede compartir un material hereditario común y multiplicarse como conjunto por medio del proceso de fragmentación. En este tipo de reproducción, algún efecto mecánico del ambiente rompe un fragmento de la bioplaca conteniendo matriz y algunos millones de células que se desplazaran con la corriente hasta llegar a un nuevo lugar, donde la colonia puede crecer como bioplaca y no como bacterias individuales (Battin et al., 2007; Claessen, Rozen, Kuipers, Søgaard-Andersen, & Van Wezel, 2014).
7. Reproducción asexual y parasexual en hongos
7. Reproducción asexual y parasexual en hongos
La reproducción es un proceso vital para la continuidad de la vida y los hongos no son la excepción. En este artículo, exploraremos los diferentes métodos que utilizan los hongos para reproducirse y cómo estos pueden variar según la especie y el ambiente en el que se encuentran. Desde la reproducción asexual hasta la sexual, pasando por la formación de esporas y la germinación, descubriremos la complejidad y diversidad de los procesos reproductivos en los hongos y su importancia en el ciclo de la vida de estos organismos. Además, analizaremos las aplicaciones prácticas de la investigación en reproducción de hongos, desde la producción de alimentos hasta el desarrollo de medicamentos.
7.1 Reproducción asexual
A diferencia de las funciones de nutrición, respiración y excreción, donde los hongos se describen con mecanismos relativamente simples todos relacionados con el transporte de sustancias a través de membranas, su función de reproducción es altamente compleja, ya que involucra todo tipo de reproducción imaginable: asexual unicelular, asexual multicelular, sexual unicelular, asexual multicelular. De hecho, la reproducción en los hongos sirve como un buen modelo de comparación de otros tipos de eucariotes no fúngicos. Por tal razón analizaremos en detalle la reproducción en los hongos y luego en comparación revisaremos la reproducción de otros eucariotes. Se ha estimado que más de un tercio de las especies de hongos son capaces de reproducirse con más de un método de propagación, en diferentes etapas del ciclo de vida y con dependencia a la favorabilidad u hostilidad del medioambiente.
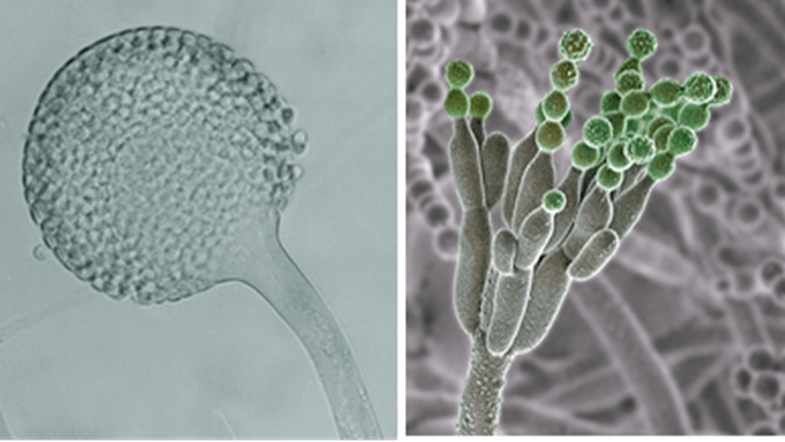
Figura 7.1. Conidios y conidióforos. (Izquierda) Los conidios se desarrollan a partir de las hifas normales o de hifas especializadas para producir los conidios, son producidas típicamente por los acomicota y algunos basidiomicotas. (Derecha) Conidióforo con conidios en verde, microfotografía al microscopio electrónico y coloreado digitalmente
La reproducción asexual de los hongos puede estar mediada por dos procesos, uno vegetativo y otro mediado por esporas. El proceso vegetativo involucra la reproducción simple de células, ya sea en colonias de individuos unicelulares como en las levaduras, o de hongos más complejos que forman hifas. Si sucede un fenómeno que traslade una parte de la colonia o un fragmento de una hifa de un lugar a otro, estas células son capaces de conformar una nueva colonia o un nuevo micelio.
Esto se debe a que los hongos poseen una estructura mucho más laxa que los animales y las plantas, separar un grupo de células de una colonia, o cortar una hifa de un micelio no afecta al resto de las células del hongo. En cualquiera de estos casos, la multiplicación celular se realiza por mitosis asexual, donde la tasa de variabilidad es relativamente baja.
Estructuras de propagación de tipo asexual
A parte del transporte mecánico vegetativo, los hongos también poseen mecanismos especializados para propagar células de tipo asexual, las cuales a su vez pueden servir como estructuras de resistencia. Las esporangiosporas por ejemplo, se forman por el clivaje del protoplasma de un esporangio multinucleado, y formando racimos de células listas para su propagación. Son producidas típicamente por los linajes de quitridiomicotas, oomicotas y zigomicotas.
Cabe resaltar que a diferencia de los cuerpos fructificantes de los basidios y los ascos, los esporangios y los conidióforos son microscópicos. Sin embargo, la estructura de conidióforos y esporangios permite la identificación rápida de los hongos en la micología clásica con un costo bajo en términos de equipo, aunque se requiere de un ojo bien entrenado y una clave “texto que permite identificar”.
Esporangios y esporangiosporas
Las esporangiosporas se forman usualmente al interior de un esporangioforo. Los esporangioforos son estructuras especializadas de las hifas donde se induce la formación de las esporas. Uno de los esporangios más conocidos es el del zigomicota llamado Mucor.

Figura 7.2. Esporangios y esporangiosporas. (A) Estructura básica de un esporangio. (B) Esporangioforos vistos al microscopio electrónico, no pueden ver más esporangiosporas debido a la membrana que cubre al esporangioforo.
Este esporangio se forma a partir de la punta de la hifa asemejándose a un globo que contienen las esporangiosporas. Dado que en los hongos existe el concepto de multi-nucleación, la punta de la hifa o esporangio es una sola célula con muchos núcleos, cuando se induce la formación de las esporas, esa gran célula experimenta clivajes, en la que el citoplasma no crece, pero si se divide para formar células cada vez más pequeñas. Posteriormente cuando cada uno de los núcleos ya cuenta con un citoplasma y membrana individualizados, una pared crece a su alrededor en el caso de los zigomicota, o se desarrollan flagelos motiles en el caso de los quitridiomicotas y oomicotas “que son falsos hongos”. Finalmente, las esporas son liberadas por una lisis controlada por parte de la pared general del esporangioforo.
Conidios y conidiosporas
¿Cuál es la diferencia entre una conodiospora y una esporangiospora? Su mecanismo de formación, o más precisamente, el lugar en donde se forman las esporas. En un esporangio, las esporas se forman al interior del esporangio, el cual posee una membrana que encierra a las esporas de forma general. En un conidio no existe dicha membrana, por lo que las membranas crecen expuesta al medio. Dado que la formación de conidios se da sin la intervención de una membrana, su estructura ha sido estudiada en detalle por medio de miucroscopios ópticos. Algunos tipos de conodios son: conidio único; burbujas secuenciales; extrusión de fialides; y picnidios.

Figura 7.3. Conidio único. Se observa en especies como Thermomyces lanuginosus en donde la punta de la hifa crece y se septa “separa de la hifa por un tabique” una única vez. A la izquierda el cultivo de T.lanuginosus en una caja de Petri vista desde arriba, a la derecha el conidio único, se diferencia de un esporangio al no presentar la columela.
Clasificación de conidios por tipo de desarrollo
Los conidios también pueden clasificarse por su tipo de desarrollo. Existen dos tipos de patrones de desarrollo básicos de un conidio, la gemación y la septación.

Figura 7.4. Burbujas secuenciales y extrusión de fialides. (A) En este caso la hifa genera estrangulaciones sucesivas, formando burbujas, que posteriormente se desarrollan hasta formar los conidiosporas. Este tipo de conidio puede encontrarse en Sclerotinia fructingena. (B) Extrusión por fialides: Una fialide es una célula semejante a un frasco, del cual emergen sucesivamente conidios en fila, lo cual forma una serie de esporas en fila. Este tipo de conidios es típico de mohos como Penicillium expansum. En algunos hongos, los conidios pueden formarse en estructuras aún más complejas como el sinema-coremium, el cual es una masa apretujada de conidioforos.
La formación basípeta de un conidio blastico o blastoconido se explica por medio de la mitosis asimétrica. La división celular, específicamente una mitosis puede ser realizada de manera simétrica o asimétrica. Por lo general, en los esquemas de los libros de texto, y en lo que se observa en la microscopia lo que se tiene son divisiones simétricas, en la que las dos células hijas poseen las mismas propiedades entre sí, es decir son células hermanas con propiedades semejantes. En la mitosis simétrica, por el contrario, una de las células hijas posee propiedades diferentes a su célula hermana. Esta propiedad de mitosis asimétrica es vital para las células madre, ya que permite que una de las células hijas retenga las propiedades de ser célula madre como tener mayor tamaño del citoplasma, mientras que la otra puede comenzar a reproducirse rápidamente para formar un tejido especializado.

Figura 7.5. Picnidios y acervulos. Otra estructura compleja es picnidio que se asemeja a un frasco cóncavo de apertura amplia donde se concentran muchos conodioforos. También existe una estructura opuesta, es decir de forma convexa donde la masa celular forma una especie de almuadilla en cuya punta crecen los conodioforos y se denomina acérvulo.
En la gemación una célula madre realiza mitosis por gemación produciendo una célula higa que sigue pegada a la célula madre, a este tipo de formación se la denomina basipeta. En la fragmentación "formación acrópeta" una hifa multinucleada se cliva formando varias células individualizadas. Cuando los conidios se forman por gemación se denominan blastoconidios, y cuando se originan por la fragmentación de una hifa se llaman taloconidios.

Figura 7.6. Tipos de desarrollo de los conidios. De izquierda a derecha, modelo y mocrofotografía de los blastoconidios; modelo y microfotografía de los taloconidios.
Utilidad evolutiva de la reproducción asexual de los hongos
La reproducción asexual de los hongos se caracteriza por una baja variabilidad de las estructuras reproductivas, es decir, las propiedades del hongo madre se transmiten a la de los hongos hijos sin que existan cambios sustanciales como los que se producen por medio de la recombinación genética. Esto es útil cuando el hongo se encuentra en un ambiente permisivo con múltiples recursos, ante los cuales el hongo ya está adaptado y calibrado para explotar con mayor eficiencia. En otras palabras, en un ambiente estable y cuando el hongo está en su pico de aptitud restringe sus mecanismos de variabilidad. Esto también implica que no genera estructuras de reproducción sexual, mientras que las condiciones del ambiente sean favorables.

Figura 7.7. August Weismann (1834, Fráncfort del Meno - 1914, Friburgo de Brisgovia) fue un biólogo alemán. Se formó en la Universidad de Gotinga y enseñó zoología en Friburgo (1866-1912). Ernst Mayr lo situó como el segundo más notable teórico evolucionista del s. XIX, detrás de Charles Darwin.
7.2 Reproducción parasexual
Los hongos también revelan un proceso que se aproxima a la sexualidad, pero que no involucra la recombinación meiótica. La parasexualidad en los hongos se denomina heterocariosis, y como su nombre da a entender es un proceso en el cual se generan células higas con números variables de cromosomas. Dos hifas de individuos diferentes se unen mediante un proceso llamado anastomosis, lo cual genera un micelio con una cantidad duplicada de cromosomas. En este punto se genera una recombinación mitótica. Finalmente, dado que hablamos de un micelio multicariótico, la cantidad de cromosomas de la especie se restaura por división nuclear directa, en contraste con la meiosis que involucra la división celular.
8. Reproducción sexual
8. Reproducción sexual
Cuando estudias la meiosis y una serie de teorías con respecto a la reproducción sexual y su utilidad evolutiva, especialmente relacionadas con la hipótesis de Weismann y la hipótesis de la reina roja, te das cuenta de que falta un tipo de ser vivo que ejemplifique todo. Ese ser vivo son los hongos, pues ellos son capaces de reproducirse sexual y asexualmente dependiendo de las condiciones predichas en dichas teorías. Peor aún, debido a esa sincronización, es posible realizar un puente entre el gradualismo filetico y el equilibrio puntuado solo analizando los mecanismos reproductivos de los hongos.
8.1 Los hongos se identifican por como realizan el acto sexual
Siendo menos teóricos y más concretos, los mecanismos de reproducción sexual de los hongos permiten algo más pragmático y es identificarlos.

Figura 8.1. Zigospora de los zigomicetos cuando la ves en un micelio puedes identificar a ese micelio como un zigomiceto.

Figura 8.2. Los ascomicetos pueden ser identificados por sus ascosporas.

Figura 8.3. Los basidios identificas a los basidiomicetos.
Muchos de los hongos llamados superiores poseen una estructura vegetativa llamada micelio, compuesta a su vez por múltiples fibras llamadas hifas.
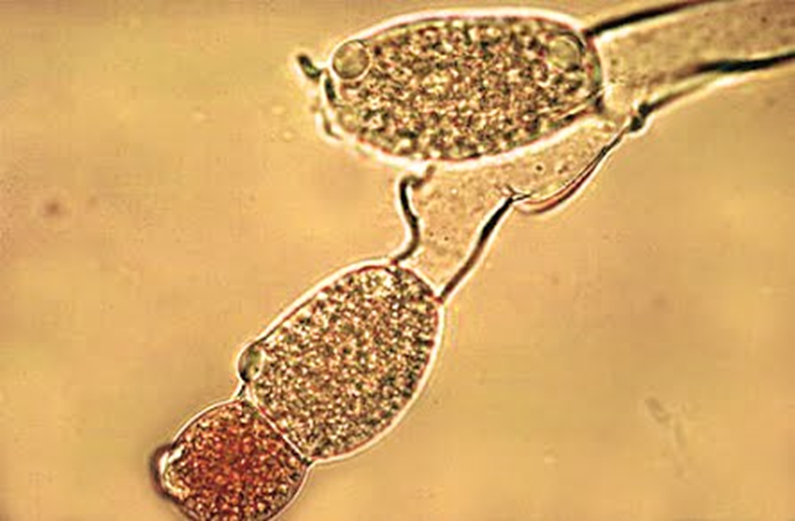
Figura 8.4. Los gametangios y las zoosporas identifican a los quitridiomicetos.
Las hifas pueden estar compuestas por múltiples células separadas entre sí por tabiques llamados septos, o ser una única masa con muchos núcleos agrupados en las puntas. En cualquier caso, la estructura de la hifa no sirve para hacer taxonomía, ya que dos hongos cercanamente emparentados genéticamente pueden tener hifas diferentes, mientras que dos hongos lejanamente emparentados pueden tener hifas semejantes. Puede darse el caso de que un hongo clasificado como superior no genere hifas, y viva en forma de colonias unicelulares. Es allí donde entra la espora sexual y su cuerpo fructificante en caso de generarlo, en otras palabras, los hongos se clasifican en sus grupos más generales “zigomicetos, ascomicetos y basidiomicetos”, de manera más general se puede decir que los hongos y otros eucariotas se identifican en base al modo en que tienen sus relaciones sexuales.
Los protistas multicelulares también se identifican de acuerdo al modo en que generan su espora sexual o generan su cuerpo fructificante como en los falsos hongos.
8.2 El ciclo de vida del hongo en comparación con un humano
En el ciclo de vida del ser humano tenemos que:
1- Los individuos adultos que se alimentan e inter-actúan con el ambiente están compuestos por células diploides. Cada uno en ciertas partes de sus cuerpos hacen la meiosis formando gametos sexuales haploides.
2- El acto sexual asocia a los gametos, los cuales pasan por varias etapas.
3- La plasmogoamia es la primera etapa de asociación de los gametos, en donde los citoplasmas de los dos gametos se fusionan.

Figura 8.5. Cigoto humano dicarionte, es decir, con dos núcleos haploides no fusionados.
4- Los humanos no poseen cariogamia, ya que en el cigoto los dos núcleos se disuelven, pero teóricamente otros seres vivos son capaces de fusionar sus núcleos para formar el núcleo diploide. En el cigoto humano los núcleos del espermatozoide y el óvulo retienen su identidad y son denominados pronucleos, es decir no es una célula diploide sino dicarionte, que el material genético no se fusiona sino hasta la etapa de dos células.

Figura 8.6. Los hongos pueden mantener el dicarionte por más tiempo y en muchas más células.
En los hongos por el contrario la plasmogamia puede generar una célula dicariotica que puede reproducirse para formar un micelio dicariotico, y solo después de muchísimas generaciones experimentarse la cariogamia. La sucesión meiosis, fecundación, plasmogamia y cariogamia cambian mucho dependiendo del hongo que se esté analizando.
En los hongos a veces trabajamos con tres generaciones celulares, la generación haploide (n) que comienza con la meiosis y termina con la plasmogamia; la generación dicarionte (n + n) en la que los dos núcleos permanecen individualizados en las hifas que va desde la plasmogamia y termina en la cariogamia; finalmente está la generación diploide (2n) en donde un núcleo contiene el doble de cromosomas y va desde la cariogamia y termina en la meiosis. Los hongos pueden llegar a tener ciclos de vida muy complejos o epicíclicos, llegando a tener inclusive sistemas de más de dos sexos y más de dos generaciones.
8.3 La sexualidad de un hongo, ¿existen machos y hembras?
Cuando un par de hifas van a realizar el acto sexual de los hongos, es bueno saber que existen diferentes tipos. Un hongo homotálico puede tener sexo consigo mismo, es algo semejante a un hermafrodita al tener ambos sexos, pero a diferencia del hermafrodita, el hongo homotálico es autofertil. Un hongo heterotalico es aquel que posee sexos diferenciados. Pero a diferencia de los animales y las plantas, donde existe un sexo masculino “muchos gametos de baja calidad” y un sexo femenino “pocos gametos de mejor calidad”, en los hongos no puede realizarse esta diferenciación.

Figura 8.7. Ciclo de vida de los hongos con alternancia de tres generaciones.
Y las cosas se pueden poner muy raras. Un caso normal es el de los hongos de compatibilidad bipolar en el cual existen dos tipos sexuales, que generalmente se denominan positivos y negativos en ausencia de poder designarlos como macho y hembra. Pero existen hongos de tipo basidio que pueden poseer una compatibilidad tetrapolar, es como si existieran cuatro sexos diferentes. En este caso la reproducción se logra si una hifa de un polo puede realizar el acto con la hifa de cualquier otro polo diferente a sí misma.
En la mayoría de los hongos los tipos reproductivos son controlados genéticamente, y eso se asocia a la producción de hormonas que permiten que los tipos reproductivos diferentes se atraigan y que los del mismo tipo se eviten. Algunos hongos como las levaduras del pan pueden presentar el mecanismo de cambio de tipo sexual, en el cual, un tipo sexual se cambia a otro por medio de la regulación genética.
Esto permite asegurarse de que la población pueda reproducirse en caso de que solo se encuentres células reproductoras de un solo tipo sexual. En palabras más simples, si en un grupo solo existen gametos de un tipo sexual, se transforman al otro, o sea cambian su sexo. Esto es un problema para un científico que desea llevar a cabo experimentos en estos hongos y requiere que no cambien el género. La solución ha sido el aislamiento de mutantes incapaces de realizar la transferencia de tipo sexual.
8.4 Utilidad evolutiva de la reproducción sexual en los hongos
La estructura sexual de un hongo solo se produce cuando el ambiente se hace hostil, por ejemplo, cuando los nutrientes se han consumido y en el ambiente solo quedan las toxinas o desechos metabólicos.

Figura 8.8. Los hongos cambian su estrategia reproductiva según el ambiente, especialmente la disponibilidad de alimento.
Adicionalmente a esto, la espora sexual es una estructura de resistencia, esto implica que es capaz de esperar hasta que las condiciones ambientales se tornen favorables. Algunas esporas sexuales están asociadas a cuerpos fructificantes, que permiten diseminaciones a largas distancias. Esto se asocia al hecho de que las esporas sexuales se producen por la meiosis, lo cual implica que su contenido genético es diferente del de sus progenitores, al haber realizado el proceso de recombinación genética.

Figura 8.9. El hongo se expande asexualmente sobre el alimento, mientras que las regiones viejas de la colonia que ya no tienen acceso a fuentes de alimento inician la reproducción sexual.
El modo en que crece una colonia de hongos ejemplifica el proceso. Mientras tienen muchos recursos los hongos generan estructuras asexuales microscópicas indiferenciables del micelio macroscópicamente, pero cuando el ambiente se torna problemático como por falta de recursos, se generan las estructuras sexuales, como en las zonas más viejas de la colonia en el centro. Esta variabilidad le permite al hongo adaptarse más raído a un ambiente desconocido, y una vez adaptado empieza a reproducirse asexualmente.
8.5 La hipótesis de la reina roja en los hongos
Weismann alguna vez se preguntó sobre el costo evolutivo de tener dos sexos, le hubiera dado una aneurisma al saber que algunos hongos pueden tener cuatro. ¿Cuál es su utilidad? En términos cuantitativos la reproducción asexual es más eficiente, pero en términos cualitativos, el sexo incrementa la diversidad al adicionar un mecanismo de mutagénesis, y es la recombinación durante la primera fase de la meiosis.
Esto va relacionado con la hipótesis de la reina roja, la cual establece que los seres vivos deben mutar para poder adaptarse a un ambiente cambiante y permanecer en su misma posición o nicho ecológico. En un ambiente cambiante los sistemas cambian y cambian solo para mantenerse vivos, es lo mismo para las empresas y para los seres vivos. Los hongos poseen cambios, cuando el ambiente no cambia ellos desaceleran, y cuando el ambiente es hostil aceleran. Los hongos restringen la reproducción sexual cuando el ambiente es permisivo y no requieren mutar para sobrevivir, pero emplean la reproducción sexual cuando deben colonizar ambientes a los cuales tal vez no estén adaptados.
8.6 Gradualismo filetico, equilibrio puntuado y la reproducción en los hongos
El gradualismo filetico establece que las mutaciones radicales son demasiado peligrosas para los organismos, por lo que es más probable que estos evolucionen de forma extremadamente continua y gradual, los cambios fisiológicos son muy lentos, más aún cuando la especie se aproxima al optimo teórico. El equilibrio puntuado hace referencia a que estos eventos de gradualismo se ven interrumpidos por eventos en los que la variabilidad se acelera, las especies evolucionan a mayor velocidad, esto generalmente asociado a cambios en el ambiente.
Cuando el ambiente es permisivo, los hongos evolucionan en base a la teoría del gradualismo filético, incluso hasta restringir los mecanismos mutagénicos más radicales como la recombinación meiótica al no permitirla “no realizar el acto sexual”. Cuando el ambiente se hace hostil, los hongos evolucionan de forma puntual, permitiendo la meiosis y por lo tanto elevando sus tasas de mutaciones genéticas, lo cual incrementa la variabilidad y la adaptabilidad de sus desdientes. Lo anterior acelera la tasa de evolución de la especie.
9. Reproducción y ciclo de vida de los quitridiomicetos
9. Reproducción y ciclo de vida de los quitridiomicetos
Los quitridiomicetes son hongos verdaderos, aunque con características bastante raras que podríamos describir como ancestrales con respecto a los más derivados grupos de zigomicetos, ascomicetos y basidiomicetos. Una de esas características es la presencia de zoosporas con flagelo en diferentes etapas de su ciclo de vida.

Figura 9.1. Mitosporangio asexual y meisoporangio sexual.
9.1 Reproducción asexual
A diferencia de otros hongos, los quitridios poseen un ciclo asexual que no está mediados por conidios, sino por esporas flageladas llamadas zoosporas. Cuatro serán las estructuras relevantes en el ciclo asexual, el micelio diploide, el zoosporangio mitótico, la zoospora y el quiste.

Figura 9.2. Zoospora.
Es importante resaltar que los quitridiomicetos poseen dos tipos de esporangios denominados indistintamente como zoosporangios, lo cual puede traer problemas a la hora de memorizar las estructuras del ciclo. Por esta razón el esporangio asexual mitótico será descrito en el presente documento como el mitosporangio, para distinguirlo del meiosporangio involucrado en el ciclo sexual.
En el mitosporangio se producen zoosporas de tipo mitótico asexual que también son llamadas zoosporas, pero como ocurre con los zoosporangios, este término se emplea casi que para cualquier propágulo de los quitridios que posee flagelos, así que las llamaremos mitosporas. Las mitosporas nadan por medio de un flagelo posterior, poseen una capa nuclear consistente en ribosomas encerrados en una membrana. Esta característica de los quitridios mejor conocidos y de cuyo ciclo de vida estamos hablando, llamado los Blastocladiales, presumiblemente facilita la síntesis de proteínas y un crecimiento rápido cuando la espora germina en un ambiente apropiado.

Figura 9.3. Gametangios.
Las zoosporas muestran atracción química por aminoácidos, proceso denominado quimiotaxis. Cuando llegan a un ambiente apropiado, la mitospora
pierde su flagelo y se denomina quiste. Posteriormente el quiste germina de manera bipolar, el micelio se abre paso al sustrato alimenticio y lo penetra a medida que se da la mitosis proliferativa para regenerar un nuevo cuerpo somático.
9.2 Reproducción sexual
El ciclo sexual inicia con la producción del segundo esporangio denominado de manera más específica como el meiosporangio, caracterizado por una pared más dura. Como su nombre indica, en el meiosporangio las esporas deben atravesar el proceso de la meiosis, para producir zoosporas haploides o meiosporas, las cuales también son zoosporas al poseer flagelos. Las meiosporas se comportan de manera similar a las mitosporas, pero después de perder su flagelo generan un micelio haploide que des denominado como micelio gametofítico. Cuando el micelio gametofitico detecta falta de nutrientes, genera estructuras sexuales llamados gametangios.
Los gametangios se desarrollan al final de las hifas, el gametangio masculino más pequeño se ubica por encima del gametangio femenino que es más grande, aunque en algunas especies se puede tener una organización inversa. El gametangio femenino no tiene color, y sus gametos también son zoosporas, de hecho, todas se parecen en tamaño y forma. El gametangio femenino son más pequeños y poseen una coloración anaranjada debido a que poseen altas cantidades de T-caroteno.
Los gametos masculinos, que son los primeros en ser liberados nadan alrededor del gametantgio femenino debido a una hormona liberada por el gametangio femenino denominada sirenina. Los gametos masculinos fecundan a los gametos femeninos a medida que estos salen del gametangio. La fecundación es plasmogánica y cariogámica, por lo que no existe una generación dicarionte. El cigoto formado obtiene en consecuencia dos flagelos. Una vez que el cigoto se ha formado, este se comportará muy semejante a las zoosporas, siendo atraído por aminoácidos, por lo que nada en su dirección. Cuando llega a un sustrato conveniente, al igual que las zoosporas monoflageladas, pierde sus flagelos e inicia el crecimiento del micelio esporofítico diploide.
9.3 Ciclo de vida
En resumen, tenemos un ciclo de vida con alternancia de generaciones haploide n (b) y diploide 2n (a).

Figura 9.4. Ciclo de vida de los quitridios.
El micelio diploide (1) es esporofítico es decir genera esporas, tanto sexuales como asexuales. Las esporas asexuales se generan a través de un meiosporangio (2a, 2b) caracterizado por una membrana gruesa. En el interior las esporas hacen la meiosis (3) para formar las meiosporas (4) que son de tipo zoospórico con un flagelo, la cual se mueve hasta encontrar un lugar apropiado, momento en el cual se enquista (5). El quiste haploide regenera (6) un micelio haploide (7) gametofítico que es capaz de realizar gametogénesis anisogámica para generar dos tipos de gametangios (8a, 8b). El gametangio más pequeño porta microgametos denominados masculinos y el más grande macrogametos denominados femeninos.
Los dos gametos (9) realizan una fecundación (10) plasmogámica y cariogámica, por lo que se pasa de inmediato a la generación diploide. El cigoto (11) es biflagelar, nada hasta encontrar un lugar apropiado para enquistarse (12). Allí germina y regenera (13) el micelio diploide (1). Dicho micelio también puede realizar un ciclo asexual por medio de la formación de mitosporangios (14a y 14b) con membranas más delgadas que los meiosporangios, pero las mitosporas (15a y 15b) son anatómicamente indistinguibles de las mitosporas al ser de tipo zoospórico. Finalmente como todas las zoosporas de los quitridios, las mitosporas buscan un lugar adecuado para enquistarse (16) y regenerar el micelio diploide (1).
10. Reproducción y ciclo de vida de los zigomicetos
10. Reproducción y ciclo de vida de los zigomicetos
En el mundo de los hongos, los zigomicetos son un grupo importante que se caracteriza por su capacidad de formar zigoesporas, estructuras resistentes que les permiten sobrevivir en condiciones desfavorables. La reproducción sexual en los zigomicetos involucra la fusión de dos hifas haploides para formar un cigoto que se desarrolla en una zigoespora. Este proceso de reproducción es crucial para la supervivencia y evolución de estos organismos, y su ciclo de vida presenta una serie de características únicas y fascinantes. En este artículo, exploraremos en detalle el ciclo de vida y la reproducción de los zigomicetos, brindando una visión general de la biología y ecología de estos hongos fascinantes.
10.1 La zigospora y el cuerpo fructificante de los zigomicetos
Las zigosporas estructura sexual de los hongos pertenecientes al grupo de los zigomicotas, aunque en el subgrupo de los phycomicetos se encuentran enterradas.

Figura 10.1. Formación de la zigospora.

Figura 10.2. Zigosporas maduras.
Las hifas que las producen son heterotálico de compatibilidad bipolar, los sexos se designan como (+) y (-). Las hifas de sexos opuestos se atraen por medio de la segregación de hormonas en el medio. Una vez que las dos hifas opuestas hacen contacto realizan la plasmogamia e inmediatamente la cariogamia, dando lugar a una zigospora. Las hifas (+) y (-) son haploides, mientras que la zigospora es diploide. En su interior se realiza de forma inmediata la meiosis.
La zigospora da lugar al cuerpo fructificante de loz zigomicetos llamado esporangio, en cual es una hifa que crece desde la zigospora, en cuya punta y encerradas en una membrana se encuentran almacenadas olas zigosporas, a la espera de su maduración. La mayoría se libera en el aire.
10.2 Ciclo de vida de los zigomicetos
El ciclo de vida de los hongos del linaje de los zigomicetos se explica por medio del modelo biológico de Rhizopus spp. mejor conocido como el humilde moho del pan.

Figura 10.3. Ciclo de vida.
El cuerpo somático que soporta la selección natural en los zigomicetos es haploide y se denomina micelio generador de gametos (1b), el micelio está compuesto por fibras individuales denominadas hifas (1a). A su vez el micelio cuando ha invadido fuertemente un tejido de alimento como un pan (1c) adquiere una apariencia aterciopelada y una coloración verde provocada por los esporangios (11, 7). Este micelio puede realizar un ciclo de vida sexual mayor y un solo epiciclo asexual en el cuerpo generador de gametos.
Iniciemos pues por el ciclo sexual mayor, las hifas del micelio parecen iguales, pero algunas son copatibles sexualmente, dado que son isogámicas, estas hifas gaméticas no reciben indicación como ovulo o esperma, además ni siquiera son células independientes para llamarlas de ese modo, por lo tanto los biólogos los han denominado como hifas (+) e hifas (-) (1d), hay que recordar que el micelio ya es de por si haploide por lo que no hay meiosis en este punto, simplemente el micelio ya es potencialmente sexual, pero dicha sexualidad solo se activa en caso de que las condiciones ambientales se hagan hostiles.
Una vez que dos fibras sexuales se han reconocido mutuamente se fusionan sus dos células apicales experimentando una fecundación cigámica (2) pero sin experimentar cariogamia. Esto da lugar a una célula dicarionte denominada como la zigospora inmadura (3). Posteriormente dicha zigospora inmadura experimenta la cariogamia (4) dando lugar a una zigospora madura dicarionte (5). Hay que destacar que la única célula que pasa por los estados de dicarionte y diploide es la zigospora, el resto del micelio que las rodea, incluyendo las células supensoras siguen siendo igual de haploides que siempre. En la propia zigospora ocurre la meiosis y se transforma en zigosporangio (6) en es igualito a una zigospora, solo que de él emerge un micelio haploide que formará un esporangio asexual (7) del cual emergen esporas asexuales estándar (8). Las esporas son liberadas del esporangio y estas por procesos mitóticos regeneran (9) el micelio haploide somático (1a, b, c, d).
El micelio puede a su vez generar un epiciclo asexual, en este caso las mismas hifas con capacidad de generar la zigospora, también pueden generar esporangios que no germinan de una zigospora por medio del proceso de esporulación (10) que al igual que el esporangio que sale de la zigospora posee la misma forma (11, 7) y genera las mismas esporas (12, 8). La única diferencia entre los dos esporangios es la carga genética, la zigospora emerge para recombinar los cromosomas de modo tal que el micelio de la siguiente generación tenga nuevas propiedades. En cualquier caso, estas esporas también pueden regenerar (9) el micelio haploide somático (1).
11. Reproducción y ciclo de vida de los ascomicetos
11. Reproducción y ciclo de vida de los ascomicetos
Los ascomicetos son un grupo de hongos que se caracterizan por producir estructuras reproductivas llamadas ascos, que contienen las esporas sexuales. Su ciclo de vida es complejo e implica tanto la reproducción sexual como asexual. La reproducción asexual se produce mediante la formación de esporangios o conidios, mientras que la reproducción sexual implica la fusión de gametos haploides y la formación de un cigoto diploide, que luego se divide por meiosis para producir ascos y esporas sexuales haploides. En este artículo se explorará en detalle el ciclo de vida y la reproducción de los ascomicetos, uno de los grupos de hongos más diversos y ampliamente distribuidos en la naturaleza.
11.1 La ascospora y el cuerpo fructificante de los ascomicetos
La espora sexual de los hongos ascomicetos se denomina ascospora, el cuerpo fructificante microscopio se denomina asco, y el cuerpo fructificante macroscópico se denomina ascocarpo. Un asco típico es como un estuche con ocho ascosporas formadas en secuencia.

Figura 11.1. Asco de una levadura.

Figura 11.2. Ascos de un micelio.
Los ascos generalmente se generan en masa como si fueran el tapiz atercipelado de la superficie del correspondiente ascocarpo. Los ascocarpos son estructuras de forma muy variable que crecen en el sustrato, algunos tienen forma de orejas, otros forman de cocos, algunos son muy coloridos y otros extremadamente venenosos llamados ascocarpos. Los ascocarpos en su interior poseen una enorme cantidad de estuches denominados ascos, los cuales contienen ocho gametos meióticos, y cada uno de esos gametos es una ascospora.
11.2 Fertilización en los ascomicetos
La parte sexual del ciclo de vida de los ascomicetos inicia cuando dos hifas haploides se unen realizando la plasmogamia "proceso llamado apareamiento o mating". Existen especies homotálicas y especies heterotálicas. La mayoría de las especies poseen compatibilidad bipolar, pero algunas especies pueden llegar a tener más sexos. Las hifas forman estructuras complejas, un anteridio que permite a los núcleos migrar hacia la hifa opuesta que ha formado un ascogonio, permitiendo una plasmogamia.

Figura 11.3. Ascocarpos.

Figura 11.4. Arriba, anteridio y ascogonio después de haber realizado el emparejamiento. Abajo, el proceso anterior llamado emparejamiento. En el modelo tenemos: A tenemos el emparejamiento, en B la plasmogamia, en C el micelio dicarionte que puede iniciar directamente la formación del asco como en las levaduras o puede crecer para formar estructuras enormes o ascocarpos, de los cuales algunas hifas iniciarán la formación del asco. La formación del asco se muestra desde D-G con la cariogamia y desde G-I con la meiosis. La maduración del asco se alcanza en J con una mitosis adicional.
A diferencia los animales y las plantas, la plasmogamia no es seguida por la cariogamia, en su lugar se genera una generación completa de células dicariontes que construyen micelios completos.
11.3 Formación del asco y del cuerpo ascocarpo
En las especies que generan un cuerpo fructificante macroscópico, el micelio dicarionte se agrupa en masas semejantes a tejidos para formar los ascocarpos. Al interior de una sección del ascocarpo se llevada a cabo la cariogamia. En las especies en las que no se forma el ascocarpo como en las levaduras, la cariogamia se da inmediatemte formando el asco. Una vez formado el asco cigótioco que posee solo una célula y un núcleo se lleva a cabo la meiosis, que conlleva a la formación de cuatro células encerrradas en un estuche. En la mayoría de las especies, al finalizar la meiosis prosigue una mitosis de las cuatro células generando ocho células a las cuales se las deniomina ascosporas, las cuales son haploides.
Figura 11.5. El apotecio, está compuesto por micelio dicarionte, en medio pueden verse los ascos maduros con sus ocho ascosporas.
11.4 El ciclo de vida de los hongos ascomicota
El ciclo de vida de los ascomicetos representa una alternancia de generaciones entre una generación haploide n (b) y una generación dicarionte n + n (c), mientras que el diploide 2n (a) se reduce a una etapa momentánea en la estructura reproductiva sexual llamada asco diploide (9).

Figura 11.6. Ciclo de vida de los ascomicetos.
Aunque en ocasiones la etapa dicarionte puede ser macroscópica y conspicua, en realidad la generación somática que afronta la mayor parte de la selección natural es la generación haploide. En consecuencia, iniciamos nuestra narración con el micelio haploide (1) el cual es una colección de fibras individuales denominadas hifas (1a), las cuales solo pueden ser vistas al microscopio (1b), aunque si se acumulan las suficientes se puede apreciar el micelio como si fuera una estructura aterciopelada (1c). Hay que destacar que el ciclo de vida ascomicota que mostramos en la figura anterior es el de los ascomicota formadores de micelio, existen otros ascomicetos como las levaduras que no forman micelios. El micelio crecerá de manera vegetativa por regeneración mitótica (20) a menos que las condiciones ambientales se hagan hostiles, momento en el cual se activa su potencialidad sexual.
Algunas hifas son compatibles con otras, aunque tal compatibilidad no es distinguible “isogamia” de allí que no se emplee el concepto de macho y hembra, sino de hifa compatible “+” e hifa compatible “-“(1d). Cuando las hifas compatibles experimentan la fecundación plasmogámica (2) emerge una nueva estructura. En lugar de ser una mera espora con un dicarionte transitorio como en los zigomicetos, lo que ocurre aquí es que no se da la cariogamia, y en su lugar crece un nuevo micelio vegetativo de tipo dicarionte (3). La función del micelio dicarionte es la formación de estructuras reproductivas sexuales mediante mitosis regenerativa (4), que pueden ser macroscópicas (5) o microscópicas (6) pero en general las denominamos como ascocarpos. Al interior de los ascocarpos se encuentran estructuras reproductivas llamadas ascos (7) los cuales inician en un estado de dicarionte. Cuando el ascocarpo alcanza la madurez el asco atraviesa la cariogamia (8) lo cual hace que los dos núcleos del asco se fusionen para generar un asco diploide (9). El asco diploide realiza meiosis inmediatamente (10) generando cuatro núcleos haploides que inmediatamente se independizan en cuatro proto-ascoesporas al interior de la membrana del asco haploide inmaduro (11). Posteriormente, el asco haploide inmaduro realiza una mitosis (12) generando cuatro ascosporas, y en este punto tenemos un asco maduro con ocho ascosporas (13a, 13b). Cuando es asco se rompe libera las ascosporas (14), las cuales por regeneración mitótica (15) regeneran el micelio haploide (1).
El ciclo sexual mayor puede ser acompañado por un ciclo asexual especializado, que se da para que el micelio invada más rápido una fuente de alimento en un ambiente favorable. En este caso el micelio haploide inicia la esporulación (16) generando estructuras especializadas denominados conidioforos (17a, 17b) los cuales liberan las conidiosporas (18). Las conidiosporas pueden germinar un nuevo micelio haploide por regeneración mitótica (19). Adicionalmente, el propio micelio puede regenerarse a sí mismo (20) para crecer o para dividirse por fragmentación.
12. Reproducción y ciclo de vida de los basidiomicetos
12. Reproducción y ciclo de vida de los basidiomicetos
Los basidiomicetos son un grupo diverso de hongos que incluye a las setas, los champiñones y los hongos portobello, entre otros. La reproducción de los basidiomicetos es única en comparación con otros grupos de hongos, ya que producen esporas en estructuras especiales llamadas basidios. El ciclo de vida de los basidiomicetos incluye tanto una fase haploide como una fase diploide, y puede variar dependiendo de la especie. En este artículo, exploraremos en detalle la reproducción y ciclo de vida de los basidiomicetos, incluyendo los diferentes tipos de basidios, la formación de esporas y cómo estos hongos se reproducen asexualmente. Además, discutiremos las aplicaciones prácticas de la investigación sobre los basidiomicetos en campos como la biotecnología y la medicina.
12.1 La basidospora y el cuerpo fructificante de los basidiomicetos
La espora sexual de los hongos basidiomicetos se denomina basidiospora, el cuerpo fructificante microscópica se denomina basidio, y el cuerpo fructificante macroscópico se denomina basidiocarpo.
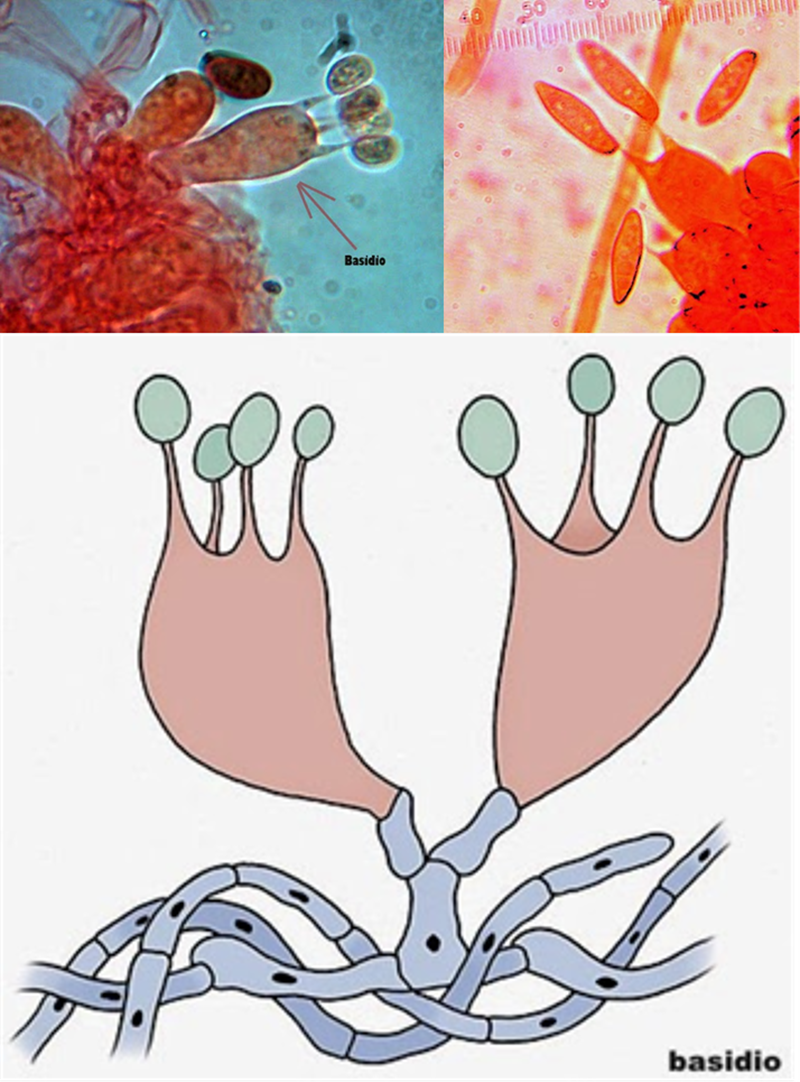
Figura 12.1. Microfotografías y modelos del basidio y sus cuatro basidiosporas. Los basidios producen cuatro basidiosporas, pero debido a que estas se liberan pueden encontrarse especímenes con menos.
Los basidiocarpos son estructuras macroscópicas con forma de sombrillas bastante conspicuas, poseen coloraciones muy marcadas y muchos producen metabolitos secundarios que pueden ser alucinógenas o incluso mortales. Los basidios en un basidiocarpo se encuentran por debajo de la umbrela “copa de la sombrilla”. El basidio es una estructura compleja, cuando está maduro lo forman cinco células, el basidio en sí y las cuatro esporas sexuales producto de la meiosis, llamadas basidiosporas. Las cuatro basidiosporas cuelgan del basidio y se desprende por gravedad, siendo transportadas por el viento donde formaran nuevas colonias.
12.2 Fertilización en los basidiomicetos
La fertilización en los basidiomicetos es similar a la de los ascomicetos, con la diferencia de que el micelio dicarionte posee una mayor importancia como cuerpo vegetativo que el micelio haploide. Los micelios sexuales o haploides no son tan importantes como estructura vegetativa en los basidiomicotas, el micelio vegetativo persistente es el dicarionte (n+n) el cual tiene acceso a la compensación de dosis, lo cual lo hace más vigoroso ante las enfermedades genéticas y los parásitos.
Esto se debe a que, al poseer dos núcleos, el micelio dicarionte tiene acceso a la compensación de dosis al poseer cromosomas homólogos, enmascarando los alelos recesivos letales. El micelio dicarionte no solo va a formar el cuerpo fructificante, de hecho, toma control sobre el sustrato. Los dicariontes tienen una expectativa de vida larga, algunos pueden llegar a vivir hasta cientos de años, además son persistentes en extremo, por ejemplo, un espécimen del parque Malheur de Oregon mide unos 8.9 kilómetros cuadrados y su edad se estima en unos 2.400 años. La especie del hongo es Armillaria solidipes, cuyos basidiocarpos “Figura 12.2” pueden emerger en cualquier lugar.

Figura 12.2. Basidiocarpo.
12.3 Formación del basidio y del basidiocarpo
Estos micelios dicariontes pueden realizar la meiosis de forma cíclica y estacional. Cuando esto sucede algunas secciones del micelio se agrupan para formar e basidiocarpo o cuerpo fructificante macroscópico en forma de sombrilla. El tejido que forma los basidios se llama himenio y se encuentra debajo de la humbrela o sombrilla del basidiocarpo.
Una vez formado el ascocarpo, en algunas secciones de este, el micelio especializa algunas células realizando la cariogamia para formar una célula diploide, la cual inmediatamente realiza una meiosis nuclear. En la imagen siguiente podemos apreciar un corte fino de un himenio, en azul está el micelio dicarionte de soporte y en rosado los basidios con las basidiosporas.
Los cuatro núcleos formados migran a la punta apical de la célula y se separan por gemación formando las cuatro células o basidiosporas.

Figura 12.3. Estructura interna del basiodiocarpo.
12.4 Ciclo de vida de los hongos basidiomicota
El ciclo de vida de los basidimicetos representa una alternancia de generaciones entre una generación haploide n (b) y una generación dicarionte n + n (c), mientras que el diploide 2n (a) se reduce a una etapa momentánea en la estructura reproductiva sexual llamada asco diploide (9). De hecho, es bastante semejante al de los ascomicetos, aunque las estructuras de los conodioforos pueden experimentar variaciones.

Figura 12.4. Ciclo de vida de los basidiomicota.
Aunque en ocasiones la etapa dicarionte puede ser macroscópica y conspicua, en realidad la generación somática que afronta la mayor parte de la selección natural es la generación haploide. En consecuencia, iniciamos nuestra narración con el micelio haploide (1) el cual es una colección de fibras individuales denominadas hifas (1a), las cuales solo pueden ser vistas al microscopio (1b), aunque si se acumulan las suficientes se puede apreciar el micelio como si fuera una estructura aterciopelada (1c). El micelio crecerá de manera vegetativa por regeneración mitótica (20) a menos que las condiciones ambientales se hagan hostiles, momento en el cual se activa su potencialidad sexual.
Algunas hifas son compatibles con otras, aunque tal compatibilidad no es distinguible “isogamia” de allí que no se emplee el concepto de macho y hembra, sino de hifa compatible “+” e hifa compatible “-“(1d). Cuando las hifas compatibles experimentan la fecundación plasmogámica (2) emerge una nueva estructura. En lugar de ser una mera espora con un dicarionte transitorio como en los zigomicetos, lo que ocurre aquí es que no se da la cariogamia, y en su lugar crece un nuevo micelio vegetativo de tipo dicarionte (3). La función del micelio dicarionte es la formación de estructuras reproductivas sexuales mediante mitosis regenerativa (4), que pueden ser macroscópicas (5) o microscópicas (6) pero en general las denominamos basidiocarpos. Al interior de los basidiocarpos se encuentran estructuras reproductivas llamadas basidios (7) los cuales inician en un estado de dicarionte. Cuando el basidiocarpo alcanza la madurez el basidio atraviesa la cariogamia (8) lo cual hace que los dos núcleos del basidio se fusionen para generar un basidio diploide (9). El basidio diploide realiza meiosis inmediatamente (10) generando cuatro núcleos haploides (11), pero hasta aquí llegan las similitudes con los ascomicetos. En lugar de atravesar una nueva mitosis, lo que sucede es que el basidio madura generando por gemación cuatro esporas a las cuales son enviados los cuatro núcleos mitóticos formando, este proceso de maduración (12) genera el basidio maduro con sus cuatro esporas típicas (13a, 13b). Cuando es basidio se rompe libera las basidiosporas (14), las cuales por regeneración mitótica (15) regeneran el micelio haploide (1).
El ciclo sexual mayor puede ser acompañado por un ciclo asexual especializado, que se da para que el micelio invada más rápido una fuente de alimento en un ambiente favorable. En este caso el micelio haploide inicia la esporulación (16) generando estructuras especializadas denominados conidioforos (17a, 17b) los cuales liberan las conidiosporas (18). Las conidiosporas pueden germinar un nuevo micelio haploide por regeneración mitótica (19). Adicionalmente, el propio micelio puede regenerarse a sí mismo (20) para crecer o para dividirse por fragmentación.
13. Reproducción y ciclo de vida de protistas de vida libre
13. Reproducción y ciclo de vida de protistas de vida libre
Al estudiar a los protistas hay que tener en cuenta su extraordinaria diversidad morfológica y de estrategias de vida, la cual se traduce casi que primordialmente en sus estrategias reproductivas. La mayoría de los protistas, al igual que los hongos, son capaces de tomar ventaja del ciclo sexual completo, incluso con los epiciclos asexuales, lo cual garantiza ciclos de vida de gran complejidad. Los ciclos de vida de mayor complejidad son los de los parásitos, que involucran alternancia de generaciones de cuerpos somáticos. En algunos casos solo uno de los dos cuerpos somáticos en el ciclo de vida de alternancia de generaciones es parásito, pero existen casos como en los plasmodios en los que los dos cuerpos somáticos parasitan a huéspedes diferentes.
En cuanto a la reproducción asexual, hay que tener en cuenta que el cuerpo somático puede ser multicelular o unicelular, así mismo las células pueden experimentar mitosis simple o mitosis asimétrica “también llamada gemación”. También se puede experimentar mitosis de policaronte, en la cual una célula experimenta divisiones nucleares sucesivas sin experimentar citocinesis, hasta que llega a tener un tamaño critico de núcleos, momento en el cual se experimenta una citocinesis masiva que fragmenta la célula madre en muchas células hijas. Esto es común en los protistas parasíticos como los apicomplejos.
El proceso mismo de la meiosis no parece ser tan estandarizado como lo hemos creído aquellos que nunca hemos estudiado a los protistas, ya que en ellos experimenta ciertas variaciones. De hecho, es bueno pensar como la espermatogénesis y ovogénesis de los mamíferos es diferente del proceso de meiosis estándar que viene en la sección de genética de la mayoría de los libros de texto. En consecuencia, es más conveniente hablar de los diversos procesos meióticos. Lo que si deben tener en común los procesos meióticos es la generación de células haploides. Y lo mismo sucede con la mitosis.
13.1 La mitosis en los protistas
Existen tipos diferentes de mitosis en los protistas, los cuales están definidos en base al huso mitótico y a la membrana nuclear, que podemos dividir en dos, ortomitosis y pleuromitosis.
Ortomitosis
El concepto de ortomitosis hace referencia a que el huso mitótico es simétrico y se puede observar un plano ecuatorial fácilmente definible. La ortomitosis puede ser abierta, semi-abierta o cerrada intranuclear. Es abierta si el núcleo de diluye y el huso atraviesa libremente, es semi-abierta si el núcleo se abre y permite la entrada del uso; y es cerrada si el núcleo no se diluye, en este caso, el huso se forma al interior del núcleo.
Pleuromitosis
La pleuromitosis hace referencia a un uso mitótico que no es simétrico con respecto al plano celular, por lo que no se puede referenciar un plano ecuatorial. La pleuromitosis puede semiabierta, cerrada intracelular o cerrada extracelular. Las dos primeras son semejantes a las que suceden en la ortomitosis, sin embargo, aún una diferente, la extra-nuclear cerrada. En este caso el huso mitótico se forma afuera y luego interactúa con el núcleo para formar un segundo huso al interior del núcleo.

Figura 13.1. Los dos tipos de mitosis.
13.2 Reproducción y ciclo de vida de los ciliados
El organismo tipo en este caso es el paramecio. Estas células son binucleadas pero con un estado cromosómico extraño. Un núcleo es más grande y es denominado macronucleo, es haploide y se encarga de las funciones somáticas, mientras que el segundo núcleo es más pequeño, es diploide y posee funciones reproductivas (1). Normalmente la reproducción asexual se da por mitosis simétrica (13), aunque también pueden existir divisiones repetitivas al interior de propágulos de resistencia conocidos como quistes.

Figura 13.2. Ciclo de vida de los ciliados
De lo anterior podemos decir que no es una generación sexual estándar, sino que es un diploide más haploide 2n + n (a).
La reproducción sexual no es estandarizada, por lo que merece la pena una discusión. La sexualidad de los ciliados recibe el nombre de conjugación. La conjugación se logra mediante la recombinación genética del micronúcleo, para lograrlo un paramecio debe identificar a otro individuo genéticamente compatible, debido a que no existe diferenciación entre las células no podemos emplear los conceptos de macho y hembra, sino el de conjugantes (2). Los conjugantes se orientan lado a lado en sus áreas bucales (3). En cada conjugante el micronúcleo lleva a cabo dos divisiones sucesivas que son equivalentes a las divisiones meióticas con un resultado de cuatro micronúcleos haploides más un macronucleo haploide (5), es decir un pentacarionte (b). Tres de los micronúcleos producidos se desintegran (6) con lo que nos deca un dicarionte (c), mientras que el micronúcleo restante se divide una vez más (7) creando un tricarionte (d). Al quedar dos micronúcleos haploides uno se queda estacionario y se llama sincarionte, mientras que el otro se mueve a través de un punto de unión que se ha formado (8). Las células intercambian el núcleo y luego este se une al otro micronúcleo restituyendo el estado diploide del micronúcleo (9).
Después del intercambio de material genético del micronúcleo, las células se separan y se denominan ahora exconjugantes. El proceso solo impacta el comportamiento celular hasta que el micronúcleo genera un nuevo macronúcleo. La generación del macronúcleo (10) tampoco es un evento sencillo, en el paramecio en sinarionte diploide se divide tres veces, produciendo ocho núcleos, todos recombinantes. Tres de ellos se degradan, uno permanece micronúcleo y los cuatro restantes se agrandan. El micronúcleo restante se divide dos veces a medida que el paramecio realiza la mitosis simétrica para producir cuatro células hijas, cada una de las cuales recibiendo un macro núcleo y un micronúcleo. Los ciliados presentan variaciones de estos eventos que involucran el número de divisiones nucleares y la cantidad de núcleos presentes.
En caso de no encontrar conjugantes, la célula puede igual recombinar consigo misma para aumentar la tasa de mutación, proceso denominado autogamia (11). La autogamia procede semejante a la conjugación en el sentido de que el micronucleo se divide de manera semejante a la meiosis para generar un pentacarionte, pero en lugar de que uno de los micronúcleos haploides sea transferido a otra célula, simplemente se vuelve a fusionar con el sincarionte que no se mueve. Una vez completado eso, se vuelve a dar un reemplazo del macronucleo.
13.3 Reproducción y ciclo de vida de la euglena no parasítica
Los euglénidos poseen formas de vida libre como la euglena común pero también hay formas parasíticas muy famosas como los tripanosomas. Examinaremos primero el ciclo de vida de la forma de vida libre. Los euglenoides poseen un cuerpo somático unicelular, haploide que solo lleva a cabo reproducción asexual (1). La mitosis (2) se realiza por pleuromitosis untranuclear cerrada.

Figura 13.3. Ciclo de vida de la euglena no parasítica.
13.4 Reproducción y ciclo de vida de los estramenopilos
La mitosis en la mayoría de los estramenopilos es caracterizada por una pleuromitosis abierta sin centriolos. Durante la división, los cuerpos basales de los dos flagelos se separan y se forma un huso mitótico de manera adyacente a los dos flagelos. En las formas que poseen escamas, la armadura escamosa se adiciona a medida que la superficie de la célula hija aparece. En las diatomeas cada célula obtiene una válvula de silicio y sintetizan la otra para completar el frustalo.

Figura 13.4. Ciclo de vida 1.

Figura 13.5. Ciclo de vida 2.
La reproducción sexual está pobremente estudiada en la mayoría de los linajes, aunque en las observaciones realizadas hasta el momento aparentemente se forman gametos haploides que luego se fusionan para generar un zigoto diploide. La mayoría de los gametos son isogámicos, aunque en las varias diatomeas se puede observar el gameto masculino con flagelos. De hecho, la variabilidad es tan grande, que se pueden observar dos tipos de ciclo de vida diferente.
En el primero la célula somática es haploide (1a) la cual puede realizar mitosis para aumentar su población (2). La misma célula haploide tiene capacidad gamética (1b), en caso de que sea anisogámico simplemente debe encontrar otra célula compatible, experimentando la fecundación y la cariogamia (2) para producir un cigoto diploide (3) que inmediatamente ingresa en meiosis (4) liberando nuevamente células haploides vegetativas.
El segundo ciclo de vida se caracteriza por una dominancia de la célula diploide (1) siendo esta la célula vegetativa que puede reproducirse por mitosis (2). La célula diploide puede ingresar en meiosis (3) que dependiendo del tipo genético puede ser espermatogénesis (3a) u ovogénesis (3b). Del proceso meiótico salen gametos anisogámicos, siendo espermatozoides masculinos (4) u óvulos femeninos (5). Los espermatozoides son atraídos por hormonas liberadas por los óvulos hasta que se unen en la fecundación (6) generando un cigoto diploide (7) que se transforma en una célula vegetativa (1).
13.5 Reproducción en actinipodios
La reproducción asexual es realizada por fisión binaria, fisión múltiple y gemación, con una división del núcleo por peluromitosis intranuclear cerrada. En los heliozoos la fisión binaria se da a lo largo del plano a través del cuerpo; en los polixistinios y varias otras formas acorazadas, sin embargo, la división ocurre a lo largo de planos predeterminados por la simetría corporal y el arreglo esquelético. La fisión múltiple tiende a ser la misma en todos los grupos. El núcleo poliploide surge a través de múltiples inicios mitóticos sin división nuclear, luego ese núcleo se fragmenta junto con el individuo, produciendo células hijas biflageladas que nadan a nuevos ambientes hasta que pierden sus flagelos y desarrollan la célula adulta acorazada. En los policistinios, la forma flagelada posee un cristal de tetraoxidosulfato de estroncio en sus citoplasmas. La fisión múltiple es el único modo de reproducción asexual de muchas especies, sino es que la única.
La reproducción sexual es rara en los actinopodios, aunque para iniciar, sus células vegetativas son diploides. Unos cuantos géneros de helizoos realizan autogamia que es una especie de autorecombinación y autofertilización como en los ciliados. En este caso la autofertilización inicia por cambios en el ambiente que indican a las poblaciones celulares que es necesario producir una nueva generación con variación genética para adaptarse a las nuevas condiciones, lo primero es que la célula se enquista para evitar morir en las nuevas condiciones. Posteriormente realiza una división mitótica para producir células sexualizadas llamadas gamontes. Cada núcleo del gamonte se divide meioticamente sin realiza citocinesis, por lo que ahora tenemos células dicarionticas. Cada uno de los gamontes pierde uno de sus núcleos haploides, volviéndose una célula gamética haploide, y posteriormente se vuelven a fusionar célula y núcleo, reformando una célula diploide autofertilizada.
13.6 Reproducción de los radiolarios
Poseen ciclos de vida complejos que no se han descrito completamente. Estos ciclos de vida involucran alternancia de generaciones, sin embargo, algunas especies pequeñas se reproducen únicamente de manera asexual mediante gemación y fisión múltiple. La división del núcleo se realiza por pleuromitosis intranuclear.
En las especies que se reproducen sexualmente no es raro encontrar individuos de la misma especie que se diferencian por tamaño y forma en diferentes etapas de su ciclo de vida. Las diferencias de tamaño generalmente están determinadas por el tamaño de la pared inicial o proloculum siguiendo una etapa de un ciclo de vida en particular. Generalmente el proloculum que se forma siguiendo procesos asexuales es significativamente más grande que el que se forma para el proceso sexual. Los individuos con proloculos grandes son denominados la generación megasférica, y los individuos sexuales son la generación microsférica. Durante la etapa sexual del ciclo de vida los individuos sexuales o gamontes realizan divisiones repetitivas para producir y liberar isogametos triflagelares, los cuales se fusionan para formar el individuo asexual de la generación diploide. Los individuos asexuales o agamontes realizan la meiosis para producir los gamontes sexuales.
En muchos foraminíferos los gametos flagelados son producidos y liberados, la fertilización ocurre en el océano para producir el agamonte joven. En otros linajes como los glabratellios, dos o más gamontes se unen de manera temporal sin fusionarse. Estos gametos que pueden ser flagelados o amebianos se unen en cámaras. Las cámaras eventualmente se separan, liberando los recién formados agamontes.
14. Reproducción y ciclo de vida los falsos hongos
14. Reproducción y ciclo de vida los falsos hongos
A parte de los cuatro filos de hongos verdaderos: quitridiomicetos, zigomicetos, ascomicetos y basidiomicetos, los micólogos también estudian tres linajes de protistas de manera común, los oomicetos, los mixomicetos y los mohos mucilaginosos.
14.1 Reproducción y ciclo de vida de los oomicetos
El grupo de organismos informalmente conocidos como oomicetos son ahora reconocidos como un filo independiente al de los hongos conocido como el reino Chromista. Son un producto notable de la evolución paralela, se asemejan tanto a los hongos verdaderos en la estructura y el estilo de vida que han sido estudiados siempre por los micólogos y fueron considerados hasta hace muy poco tiempo como hongos. Se conocen unas 700 especies.
Su fase sexual tiene una clara diferenciación entre estructuras femeninas grandes y estructuras masculinas pequeñas, denominadas oogonios y anteridios, respectivamente. Dentro de un oogonio se produce la meiosis y, dependiendo de la especie, se producen una o unas pocas oferasas (óvulos no fecundados). Cada uno contiene, cuando madura, un solo núcleo haploide. La meiosis también ocurre en los anteridios, a medida que estos crecen hacia un oogonio. Desde allí los tubos de fertilización del anteridio penetran la oogonio, y un solo tubo de fertilización entra en cada oosfera. Un solo núcleo haploide pasa del anteridio a través del tubo de fertilización, y se funde con el núcleo haploide en la oosfera. La oosfera entonces se convierte en la oospora (óvulo fertilizado) característico del grupo. Cada oospora tiene un solo núcleo diploide. Cuando la oospora germina da lugar a un micelio que es diploide, en contraste con el micelio haploide/dicarionte de la mayoría de los otros hongos. Otra característica de los oomicetos que los separa de los hongos verdaderos es la presencia de una zoospora biflagelada.

Figura 14.1. Ciclo de vida de los oomicetos.
La etapa dominante en el ciclo de vida de los oomicetos es un micelio diploide 2n (1, 1a), el cual está compuesto por fibras individuales denominadas hifas (1b), las cuales se manifiestan marcoscópicamente solo cuando el micelio es lo bastante grande como para manifestarse macroscópicamente (1c) en forma de una estructura aterciopelada. Cuando los nutrientes se agotan inicia el ciclo sexual. En este caso algunas hifas realizan una meiosos ovogénica mientras que otras realizan una meiosis espermatogénica (1d). El producto son estructuras diferenciadas sexualmebnte por estructura, por lo que se emplean los conceptos de un ateridio masculino (2) y una oosfera femenina (3). El anteriodio crece en dirección de la oosfera para realizar la plasmogamia (4) y posteriormente la fecundación de todos los óvulos presentes (5). Únicamente los núcleos contenidos al interior de las estructuras sexuales son haploides n (b), mientras que el micelio que los soporta es diploide 2n (a). La fecundación da lugar una oosfera madura o oosporangio llena de oosporas (6).
Cuando son liberadas las oosporas (7) germinan (8) para producir zoosporas (9) las cuales producen el micelio diploide por regeneración mitótica (10). La etapa asexual del ciclo se emplea para acelerar la colonización y en consecuencia ocurre cuando hay fuentes de alimento. En este caso el micelio diploide produce una esporulación (11) produciendo un zoosporangio (12), el cual al germinar (13) genera nuevamente las zoosporas (9) que prosiguen con el ciclo.
14.2 Reproducción y ciclo de vida de los mixomicetos
La característica especial del ciclo de vida de los mixomicetos es una estructura llamada plasmodio. Un protoplasma policarionte que funciona como una supercélula sin individualidad. Es una etapa pasajera del ciclo de vida, y es más común observar en la naturaleza los cuerpos fructificantes. Dependiendo de la especie el plasmodio puede ser muy pequeño, pero en otras, el plasmodio puede crecer en una entidad macroscópica del tamaño de un plato de cocina. Dichos plasmodios pueden observarse en ocasiones en la naturaleza en forma de una masa amarilla y babosa cerca de la madera en descomposición.
(YouTube) No son precisamente seres bonitos.
Los plasmodios poseen también una etapa de ameba flagelada y natatoria. Tanto la etapa de ameba y plasmodio son somáticas, en el sentido de que se alimentan y afrontan la mayor parte de la selección natural. La plasmogamia se da entre las amebas, las cuales no se distinguen entre sí, además genéticamente solo se diferencias por genes y marcadores de membrana que las hacen compatibles. El cigoto desarrollará generalmente el plasmodio. Algunas especies son apomiticas, es decir, son algo semejante a la partenogénesis, en la cual una ameba puede engañar su maquinaria genética para iniciar una metamorfosis en un plasmodio haploide. Otras especies son capaces de autofertilización.
La formación del plasmodio se da a partir de un cigoto que experimenta mitosis nuclear sin citocinesis, estas divisiones recuerdan a los clivajes de los animales en el sentido de que son sincronizadas, pasando por las etapas de 2, 4, 8, 16 y más núcleos. Pronto se obtiene una masa de núcleos incontables de millones de unidades. La mitosis es cerrada, evitando que los núcleos intercambien cromosomas de manera caótica. Los plasmodios no tienen sentido de individualidad, por lo que pueden fusionarse con otros plasmodios en crecimiento. A medida que la masa crece, se inicia una necesidad por llegar a regiones donde hay mejores condiciones, por lo que empieza a observarse una corriente protoplásmica hasta desarrollar venas de movimiento características de un plasmodio maduro.
Al igual que con otros microorganismos, la etapa somática no cambia a menos que existan limitaciones nutricionales. La falta de alimentos inicia cambios en el plasmodio, que dependerán de la presencia o ausencia de luz solar. En la oscuridad el plasmodio se transforma en un esclerotio, que consiste en múltiples esférulas, cada una encerrada en una pared gruesa que contiene muchos núcleos con citoplasma. Los esclerotios funcionan como un quiste de resistencia que puede soportar por varios años, pero en presencia de nutrientes, las esférulas germinan y reemerge el plasmodio. En presencia de luz se genera un cuerpo fructificante que esporula por medio de la meiosis. Las células que conforman el talo y la cubierta se sacrifican, mientras que las del interior del esporangio son meióticas, y se liberan al ambiente. Las esporas liberadas inicialmente son estructuras de resistencia. Si llegan a un ambiente favorable desarrollaran la etapa de ameba flagelada.
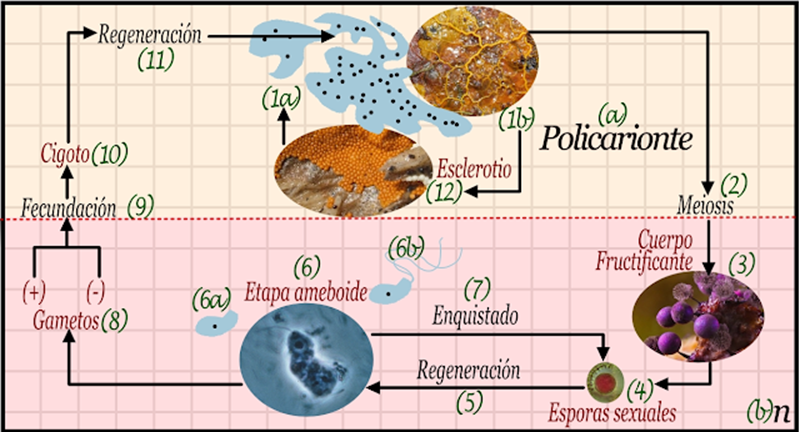
Figura 14.2. Ciclo de vida de los mixomicetos.
El ciclo de vida de los mixomicetos puede clasificarse como de alternancia de generaciones, ya que hay dos cuerpos que pueden alimentarse en cada una de las etapas policarionte (a) o haploide n (b). Iniciamos el ciclo de vida en la etapa policarionte, que forma el plasmodio (1a y 1b). Cuando las condiciones se ponen malas en presencia de luz formará a través de la meiosis (2) un cuerpo fructificante (3) que liberará esporas sexuales cuando madura (4). De las esporas emerge la etapa ameboidea del ciclo de vida, dichas amebas pueden alimentarse, y pueden oscilar en dos tipos, la ameba no flagelada (6a) característica de ambientes más secos y la ameba flagelada (6b) que nada en ambientes más húmedos. Si el ambiente se torna muy adverso ambas amebas pueden re-enquistarse (7), el quiste formado funciona igual que una espora sexual creando un epiciclo de resistencia.
Ambos tipos de ameba poseen compatibilidades sexuales, pero las amebas son isogámicas, por lo que no podemos emplear el concepto de macho y hembra, por lo que empleamos los conceptos de compatibilidad (+) y (-). Las amebas gaméticas (8) realizan la plasmogamia y la cariogamia en la fecundación (9), formando un cigoto diploide (10) el cual por medio de mitosis nuclear sin citocinesis regenera (11) la etapa de plasmodio.
14.3 Reproducción y ciclo de vida de los mohos mucilaginosos
Los mohos mucilaginosos se diferencian de los mixomicetos en que el plasmodio es celularizado, en otras palabras, las células mantienen su identidad a medida que se forma una estructura macroscópica. El grupo más famoso de este linaje es Dictyostelium.
En este ciclo de vida domina la generación haploide n (b) mientras que la etapa diploide 2n (a) se reduce al interior del macroquiste (4). Iniciamos la discusión de este ciclo de vida con una población de amebas que viven en suspensión (1a y 1b). Cuando las condiciones ambientales se tornan adversas se pueden tomar dos caminos, uno sexual y otro asexual, discutiremos inicialmente el ciclo sexual. En ambos ciclos el proceso comienza con una agregación (2) de las amebas, pero en el ciclo sexual existe un proceso de canibalización y maduración (3) en el cual las amebas muertas producen o contribuyen a la formación de una pared externa para formar un macroquiste de resistencia (4).
Las células al interior del macroquiste experimentan la fecundación (5) e inmediatamente la meiosis (6) para producir las esporas sexuales (7) que regeneran mitóticamente (8) a la población de amebas (1). Las amebas pueden aumentar su población por regeneración mitótica (9) o ingresar en el ciclo de esporulación asexual (10). En este caso la agregación produce una especie de plánula o babosa (12) que repta hasta llegar a un lugar adecuado, donde se desarrolla en un cuerpo fructificante (13). Cabe destacar que el ciclo del cuerpo fructificante (11) es la etapa más famosa del ciclo de vida de Dictyostelium. El cuerpo fructificante libera esporas asexuales (14) que regeneran la etapa de amebas somáticas (1).
15. Reproducción y ciclo de vida de algunos protistas parásitos
15. Reproducción y ciclo de vida de algunos protistas parásitos
A continuación, discutiremos los ciclos de vida de algunos protistas de importancia médica, aunque también discutiremos algunos otros linajes importantes.
15.1 Reproducción y ciclo de vida de las amebas
La fisión binaria simple es el medio más común para que las amebas realicen la reproducción asexual, cambiando solo en detalles menores al interior de cada linaje. En las amebas desnudas la división nuclear inicia primera y posteriormente la división del citoplasma la sigue. Durante la división citoplasmática las dos células hijas forman pseudópodos locomotores para ayudarse en la citocinesis. En las especies con una pared, esta se puede dividir más o menos de manera igualitaria en conjunción con la formación de las células hijas, aunque lo más común es que la pared sea retenida por una de las células “madre” y la célula hija salga inicialmente desnuda, pero al poco tiempo pueda desarrollar su propia pared.

Figura 15.1. Ciclo de vida de las amebas.
La fisión múltiple y también es común en algunos linajes. En Entamoeba hystolítica se producen propágulos llamados quistes, al interior de los cuales se produce fisión múltiple. La formación de los quistes se da cuando las condiciones ambientales se hacen desfavorables. La mitosis de la mayoría es una ortomitosis abierta sin centriolos; aunque el miembro más relevante del grupo que es E. hystolitica posee ortomitosis intranuclear cerrada. No se ha descrito ciclo sexual, por lo que los quistes se generan directamente del individuo vegetativo (Weedall & Hall, 2011).
El ciclo de vida se separa en dos etapas celulares, el quiste de cuatro núcleos (1) y el trofozoito amebiano de un núcleo (3). La ploidia de cada uno de ellos aún sigue siendo materia de debate (Weedall & Hall, 2011), además no se conoce etapa de recombinación genética (Tibayrenc, Kjellberg, & Ayala, 1990).
Los quistes se encuentran en la materia fecal y cuando esta contamina el agua lo la comida, o cuando se realizan prácticas sexuales como el ana-lingus (Burnham, Reeve, & Finch, 1980) los quistes ingresan junto con algunos trofozoitos al sistema gastrointestinal. Los trofozoitos mueren en el estómago, pero los quistes al ser propágulos de resistencia logran llegar al intestino donde reconocen la presencia de un ambiente favorable, y en consecuencia inician el proceso de exquistación (2).

Figura 15.2. Quiste.
Una vez que el trofozoito (3, Figura 60) se ha exquistado inicia el ciclo de regeneración asexual por medio de la mitosis (4). Dependiendo de la especie la etapa de crecimiento intestinal puede contenerse allí mismo, o en el caso de E. hystolitica, romper la pared intestinal e iniciar la etapa invasiva, las células del trofozoito son iguales, pero atacan otros órganos del cuerpo.

Figura 15.3. Trofozoito.
La infección histolitica es muy peligrosa y frecuentemente fatal. Algunos trofozoitos intestinales se enquistan (5) y pasan a través de las heces junto con trofozoitos activos para reiniciar el ciclo de vida.
15.2 Las euglenas parasíticas
Las euglenas son un grupo diverso de organismos unicelulares que habitan en ambientes acuáticos, incluyendo agua dulce, salada y de aguas estancadas. Aunque la mayoría de las euglenas son fotosintéticas y se alimentan de manera autótrofa, algunas especies son parásitas obligatorias de otros organismos. Estas euglenas parasíticas han evolucionado una amplia gama de adaptaciones para invadir y alimentarse de sus huéspedes, incluyendo la capacidad de perforar y penetrar las células de los hospedadores y extraer nutrientes de ellos. En este artículo, exploraremos las diferentes especies de euglenas parasíticas, su ciclo de vida y su impacto en los ecosistemas acuáticos.
Reproducción y ciclo de vida de Tripanosoma brucei
La especie parasita dos huéspedes, un vector insecto (la mosca tse-tse) y un huésped mamífero. Debido a la gran diferencia entre estos dos huéspedes, la célula experimenta cambios complejos para facilitar su supervivencia en el intestino de los insectos y en la sangre de los mamíferos. Asimismo, cuenta con una única y notable cubierta de glicoproteína variante de superficie (VSG) con el fin de evitar al sistema inmunitario del huésped.

Figura 15.4. Ciclo de vida de Tripanosoma brucei.
El ciclo de vida inicia cuando una mosca tse-tse pica a un ser humano infectado con tripanosomiasis (1), en ese momento los tripomastigotos del torrente sanguíneo infectan el tracto intestinal de la mosca transformándose en tripomastigotos procíclicos (2).
Figura 15.5. La mosca tse-tsé Glossina spp.
Los tripomastigotos procíclicos proceden a multiplicarse por fisión binaria mitótica (3). Posteriormente los tripomastigotos prcíclicos dejan los intestinos a medida que se transforman en epimastogotos (4). Los epimastigotos migran a la glándula salivar de la mosca y se transforman en tripomnastigotos metacíclicos (5) por medio de fisión binaria mitótica (6). En este momento la mosca se encuentra infectada y en condición de infectar a otros anfitriones.
Tradicionalmente se ha dicho que el ciclo de vida de T. brucei es asexual, sin embargo, Peacock y colaboradores han sugerido que al interior de la población de promastigotos metacíclicos existe una subpoblación de promastigotos haploides gaméticos (Peacock, Bailey, Carrington, & Gibson, 2014) que poseen marcadores genéticos para la meiosis y la plasmogamia (7), lo cual permitiría la variabilidad genética requerida para cualquier parásito. Sea cual sea el asunto, en este punto la mosca pica a un nuevo anfitrión (8) infectándolo con promastigotos metacíclicos recombinantes o normales, los cuales al ingresar al torrente sanguíneo se transforman en tripomastigotos del torrente sanguíneo (9) los cuales se alimentan y reproducen (10) a expensas de los nutrientes de la sangre reiniciando el ciclo a la espera de una nueva mosca no infectada.
Reproducción y ciclo de vida de Tripanosoma cruzi
Al igual que con otros tripanosomas el ciclo de vida de T. cruzi está dominado por la generación diploide, sin embargo, en esta especie aún no ha sido reportado el promastigoto gamético sexual haploide, por lo que asumiremos de manera estándar un ciclo de vida asexual.

Figura 15.6. Ciclo de vida de Toxoplasma cruzi.
Iniciaremos nuestra discusión del ciclo de vida con la picadura de un triatomino sano a un individuo infectado (1), en el intestino del insecto, se transforman en epimastigotos (2) los cuales constituyen una segunda etapa reproductiva por medio de la fisión binaria mitótica (3). Después de la reproducción a través de mitosis, los epimastigotos pasan al recto. Allí se convierten en tripomastigotos metacíclicos (4) y se evacúan a través de las heces (5). Las heces pueden infectar a un nuevo anfitrión.
En la sangre los tripomastigotos metacíclcios buscan algunas células de tejidos específicos para atacar intracelularmente, y allí se convierten en amastigotos (6), los cuales se reproducen intracelularmente por mitosis (8). Después de la reproducción, una gran cantidad de amastigotos se encuentran en la célula infectada, formándose pseudoquistos. Los amastigotos pueden romper la célula y convertirse en tripomastigotos (9) que buscan nuevas células para convertirse en amastigotos, creando un ciclo asexual en el anfitrión. El ciclo se reinicia cuando un triatomino sano pica absorbiendo los tripomastigotos libres en el torrente sanguíneo junto con la sangre de su alimento.
15.3 Reproducción y ciclo de vida de los apicomplejos
Los apicomplejos contienen varias especies parasíticas importantes, dentro de las cuales cabe destacar a los plasmodios que causan la malaria y los toxoplasmas que causan la toxoplasmosis. Debido a que poseen ciclos de vida notablemente diferentes, existen ciertas diferencias, por lo que nos enfocaremos en las generalidades. Los aplicomplejos poseen reproducción asexual por medio de fisión binaria, fisión múltiple o endopoligenia. El ciclo de vida sexual puede ser dividido en tres etapas generales: (1) la fase sexual o gamontogonia; (2) la fase formadora de esporas o esporogonia y (3) la fase de crecimiento. Adicionalmente casi siempre se involucra a dos anfitriones parasitados, uno por la etapa haploide y el otro por la etapa diploide. Esto hace que la morfología de ambas etapas y sus correspondientes propágulos estén especializados para alcanzar a sus anfitriones y sobrevivir hasta que lo logran.
Ciclo de vida de Toxoplasma gondii
El ciclo de vida de los toxoplasmas aprovecha la relación depredador-presa. En este caso el depredador es siempre un felino y las persas son evidentemente presas comunes de felinos. En el caso de la especie que más comúnmente afecta al ser humano Toxoplasma gondii el felino es el gato doméstico (b) y la presa normal son ratones y aves, sin embargo, el ser humano puede infectarse como una presa. Iniciamos nuestra discusión con el consumo por parte del felino de carne contaminada con quistes (1), estos quistes soportan la digestión hasta llegar al intestino delgado, donde se abren y liberan las células infectantes llamados trofozoitos (2).

Figura 15.7. Ciclo de vida de Toxoplasma gondii (b)
Los trofozoitos atacan las células del epitelio intestinal por el lumen, y allí se alimentan intracelularmente, regenerando sus números por medio de la mitosis (3). Las células producidas en esta etapa se denominan esquizontes (4) y cuando estos caturan la célula, la rompen liberando los esquizontes en el lumen intestinal donde infectan a más células del epitelio intestinal (4). Una vez allí los esquizontes inician la etapa haploide n (c) la meiosis (6) por microgpenesis (6a) y macrogénesis (6b). La microgénesis da lugar a los microgametos masculinos (7) que son liberados de sus células y nadan hasta donde están los macrogametos (8), dándose la fecundación y la generación de un ovoquiste inmaduro (9). La infección causa una diarrea severa que causa la caída del epitelio, por lo que los ovoquistes inmaduros salen a través de las heces (10) al ambiente (d).
Los ovoquistes ya maduros (11) son extremadamente volátiles y contaminan el ambiente, la tierra y los vegetales con los que se alimentan las presas (12), por lo que estas se infectan fácilmente. Una vez en el interior de la presa el ovoquiste viaja por el sistema digestivo hasta que alcabnza el intestino delgado (13), allí el ovoquiste se abre y libera los esporozoitos (14). Los esporozoitos atacan e infectan células del epitelio de la presa y allí se multiplican por mitosis (15). Los esporozoitos saturan la célula del epitelio (16) y lo rompen hacia el interior del organismo, liberando los peligrosos taquizoitos (17) al torrente sanguíneo. Los taquizoitos atacan monocitos y otras células del cuerpo a gran velocidad (18) lo cual activa al sistema inmune, generando síntomas de gripa. Una vez que los taquizoitos detectan que están siendo destruidos (19) se transforman en bradizoitos (20) que igual se reproducen muy lentamente en la célula que afectan (21) y la transforman en una estructura de resistencia tisular llamada quiste (22). El quiste posee mecanismos de resistencia que inhabilitan localmente el sistema inmue, especialmente de los linfocitos T cd8 citotóxicos, el problema es que dicha protección puede difundirse al tejido cercano, protegiendo no solo al quiste, sino también a células cancerígenas cercanas, por lo que los individuos afectados por toxoplasmosis crónica tienen una tendencia estadística a desarrollar cualquier tipo de cáncer donde el parásito pueda enquistarse (Cong et al., 2015; Vittecoq et al., 2012; Yuan et al., 2007).
Los quistes permanecen casi en dormancia por años, y a veces pueden retornar a la etapa de taquizoito (23) para renovar los quistes sin generar síntomas, pero si el sistema inmune falla entonces la etapa de taquzoito se activa de manera indetenible, destruyendo todos los tejidos importantes del cuerpo. El ciclo de vida reinicia cuando el felino caza a su presa. Algunos autores sitienen que los quistes también liberan hormonas que afectan la personalidad de la presa (Corrêa, Chieffi, Lescano, & Santos, 2014; Moore, 1995; Vyas, Kim, Giacomini, Boothroyd, & Sapolsky, 2007; Vyas, Kim, & Sapolsky, 2007; Webster, 2007), haciéndola tomar riesgos innecesarios, lo cual facilita su caída ante un depredador felino.
Ciclo de vida de Plasmodium falciparum
En el ciclo de vida de los plasmodios domina la etapa diploide 2n tanto en el humano (a) como en el mosquito (c) mientras que la haploide n (b) se reduce a dos células que en algún momento están encerradas en un eritrocito y en otro no lo están.
Cuando un mosquito sano pica a un individuo infectado con plasmodio (1) los gametocitos masculinos y femeninos pasan al mosquito donde se diferencian en gametos (2a, 2b), 4-8 microgametos (3) por cada gametocito masculino y un macrogameto (4) por cada gametocito femenino y al fusionarse (5) ambos gametos, se producen los cigotos (6). Los cigotos, a su vez, se convierten en oocinetos móviles y alargados (7), que invaden la pared intestinal del mosquito (8), donde se desarrollan en ooquistes (9). Los ooquistes crecen, se rompen (10) y liberan una nueva generación de esporozoitos (11), que hacen su camino a las glándulas salivares del mosquito. Es en esta fase en la que el Plasmodium puede volver a ser inyectado en el huésped a través de una picadura (12).

Figura 15.8. Ciclo de vida de Plasmodium falciparum.
Tras la picadura del mosquito, éste inocula el parásito existente en su saliva en la sangre o en el sistema linfático del huésped, que se encuentra en la etapa de esporozoito. Los esporozoitos pasan al torrente sanguíneo hasta que llegan a los hepatocitos del hígado. Allí se multiplican por esquizogénesis (disgregación) formando el esquizonte hepático (13), tras lo cual se rompe el hepatocito, apareciendo un nuevo estadio del Plasmodium, el merozoito (14). Aquí hay un primer ciclo asexual, en el que los merozoitos pueden o bien reinfectar hepatocitos (15) o bien volver al torrente sanguíneo, donde penetran en los eritrocitos (16). Alhunas especies generan una estructura de latencia en el hepatocito que es riduculamrente pequeña, llamada hipnozoito, este desarrolla más lento y puede generar reinicios de la etapa aguda de la malaria (17) (Battle et al., 2014; Imwong et al., 2007; Wells, Burrows, & Baird, 2010).
En los eritrocitos, los merozoitos comienzan a alimentarse de la parte proteíca de la hemoglobina contenida en éstos, apareciendo entonces el trofozoíto (18). Nuevamente por esquizogénesis (19) se multiplica en el interior de dichas células, formándose el esquizonte hemático (20). También se rompe la célula, en este caso el eritrocito, liberando nuevos merozoitos (21). La mayoría de los merozoitos continúan con este ciclo replicativo (16) infectando nuevos eritrocitos, pero algunos inician la meiosis (22) se convierten en microgametocitos (23) y macrogametocitos (24) a la espera de que un mosquito sano los capture reiniciando el ciclo.
15.4 Reprodicción y ciclo de vida de las diplomonas
Las diplomonas se reproducen únicamente por reproducción asexual por medio de ortomitosis semiabierta con los dos núcleos en sincronia. Los cuerpos basales de los flagelos actúan como los centros organizadores del hiso mitótico. La mayoría de las diplomonas simbiótivcas forman quistes de protección en algún punto de su ciclo de vida. La célula somática que se alimenta se denomina trofozoito, mientras que la forma de resistencia se denomina quiste. Las formas de vida libre jamás enquistan De todas las diplomonas el género Giardia es el más relevante debido a que posee muchos simbiontes parasíticos de importancia médica pero solo examinaremos a G. lamblia.

Figura 15.9. Ciclo de vida de Giardia lamblia.
El ciclo de vida es semejante al de las amebas con solo dos etapas celulares, el trofozoito activo en los intestinos (1) el cual puede iniciar el enquistado (2) dependiendo de las condiciones, saliendo con las heces. El trofozoito activo posee una ploidicidad no estándar para un eucariota siendo un dicarionte tetraploide, por lo que tiene ocho copias por cada cromosoma (a). El quiste (3) es una estructura de resistencia, aunque no para el amiente. La materia fecal debe contaminar la comida, y generalmente en esta etapa llegan vivos trofozoitos y quistes, pero al pasar por el estómago los trofozoitos se mueren por el ácido, pero los quistes sobreviven y al llegar al intestino inician el proceso de exquistado (4) retornando a la etapa de trofozoito. El trofozoito a demás es la etapa somática que se alimenta y se reproduce mitóticamente (5) para aumentar la población. La ploidicidad del quiste también es rara, siendo un tetracarionte y tetraploide, por lo que cada quiste posee 16 copias de cada cromosoma (Bernander, Palm, & Svärd, 2001).
15.5 Reproducción y ciclo de vida de parabasilidos
La reproducción asexual se realiza por fisión binaria longitudinal mediante pleuromitosis extranuclear cerrada. Se desconoce si poseen un ciclo sexual, pero se conoce en algunos hipermastigotos. En estos organismos la reproducción sexual está bien entendida e incluye gametogamia, gametongamia y autogamia.

Figura 15.10. Ciclo de vida de Pentatrichomonas hominis.
Adiocionalmente la meiosis puede ocurrir en una o dos divisiones dependiendo de la especie. Los hipermastigotos pasan la mayoría de sus vidas como células vegetativas haploides en el tracto gastrointestinal de insectos que se alimentan de madera. En términos médicos, sin embargo, el linaje que nos interesa es el de tricomonas. Tricomonas solo posee una etapa celular llamada trofozoito (1), el cual es una célula diploide (Yuh, Liu, & Shaio, 1997), el cual se reproduce por mitosis (2).
R. Referencias bibliográficas
R. Referencias bibliográficas
Arato, R. (2010). Protists: Algae, Amoebas, Plankton, and Other Protists (1st ed.). Crabtree Publishing Company.
Bannert, N., & Kurth, R. (2004). Retroelements and the human genome: new perspectives on an old relation. Proceedings of the National Academy of Sciences, 101(suppl 2), 14572–14579.
Battin, T. J., Sloan, W. T., Kjelleberg, S., Daims, H., Head, I. M., Curtis, T. P., & Eberl, L. (2007). Microbial landscapes: new paths to biofilm research. Nature Reviews Microbiology, 5(1), 76–81.
Battle, K. E., Karhunen, M. S., Bhatt, S., Gething, P. W., Howes, R. E., Golding, N., … Smith, D. L. (2014). Geographical variation in Plasmodium vivax relapse. Malaria Journal, 13(1), 144.
Belk, C., & Maier, V. B. (2013). Biology Science for Life with physiology. (Pearson, Ed.) (4th ed.).
Benton, E. (1974). Vitalism in nineteenth-century scientific thought: A typology and reassessment. Studies in History and Philosophy of Science Part A, 5(1), 17–48.
Bernander, R., Palm, J. E., & Svärd, S. G. (2001). Genome ploidy in different stages of the Giardia lamblia life cycle. Cellular Microbiology, 3(1), 55–62.
Beukeboom, L. W., & Perrin, N. (2014). The evolution of sex determination. Oxford University Press, USA.
Black, J., & Black, L. (2012). Microbiology Principles and Explorations (8th ed.). Jefferson City.: Wiley & Sons.
Borlee, B. R., Goldman, A. D., Murakami, K., Samudrala, R., Wozniak, D. J., & Parsek, M. R. (2010). Pseudomonas aeruginosa uses a cyclic‐di‐GMP‐regulated adhesin to reinforce the biofilm extracellular matrix. Molecular Microbiology, 75(4), 827–842.
Bowen, W. H., & Koo, H. (2011). Biology of Streptococcus mutans-derived glucosyltransferases: role in extracellular matrix formation of cariogenic biofilms. Caries Research, 45(1), 69–86.
Brockhurst, M. A., Chapman, T., King, K. C., Mank, J. E., Paterson, S., & Hurst, G. D. D. (2014). Running with the Red Queen: the role of biotic conflicts in evolution. Proceedings of the Royal Society of London B: Biological Sciences, 281(1797), 20141382.
Brusca, R., Brusca, G. J., & Haver, N. J. (2003). Invertebrates (2nd ed.). Sinauer Associates.
Burnham, W. R., Reeve, R. S., & Finch, R. G. (1980). Entamoeba histolytica infection in male homosexuals. Gut, 21(12), 1097.
Carlile, M., Watkinson, S., & Gooday, G. (2001). The fungi (2nd ed.). San Diego: Academic Press.
Cavalier-Smith, T. (1995). Cell cycles, diplokaryosis and the archezoan origin of sex. Archiv Für Protistenkunde, 145(3-4), 189–207.
Chalopin, D., Fan, S., Simakov, O., Meyer, A., Schartl, M., & Volff, J. (2014). Evolutionary active transposable elements in the genome of the coelacanth. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 322(6), 322–333.
Christie, P. J. (2016). Classic Spotlight: the Awesome Power of Conjugation. Journal of Bacteriology, 198(3), 372.
Claessen, D., Rozen, D. E., Kuipers, O. P., Søgaard-Andersen, L., & Van Wezel, G. P. (2014). Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies. Nature Reviews Microbiology, 12(2), 115–124.
Clewell, D. B. (2013). Bacterial conjugation. Springer Science & Business Media.
Cloutier, M. J. (1995). Antibiotics: Mechanisms of Action and Acquisition of Resistance--When Magic Bullets Lose Their Magic. American Journal of Pharmaceutical Education, 59(2), 167–172.
Cong, W., Liu, G.-H., Meng, Q.-F., Dong, W., Qin, S.-Y., Zhang, F.-K., … Zhu, X.-Q. (2015). Toxoplasma gondii infection in cancer patients: prevalence, risk factors, genotypes and association with clinical diagnosis. Cancer Letters, 359(2), 307–313.
Corrêa, F. M., Chieffi, P. P., Lescano, S. A. Z., & Santos, S. V. dos. (2014). Behavioral and memory changes in Mus musculus coinfected by Toxocara canis and Toxoplasma gondii. Revista Do Instituto de Medicina Tropical de São Paulo, 56(4), 353–356.
Cox, F. E. G. (1993). Modern Parasitology (2nd ed.). Blackwell Science Oxford.
De Duve, C., & Pizano, M. (1995). Polvo vital: origen y evolución de la vida en la tierra. Norma, Bogotá.
Drews, G. (2000). The roots of microbiology and the influence of Ferdinand Cohn on microbiology of the 19th century. FEMS Microbiology Reviews, 24(3), 225–249.
Eldredge, N. (2014). Time frames: The evolution of punctuated equilibria. Princeton University Press.
Gibson, A. K. (2015). The Red Queen hypothesis. Education, 2010, 2014.
Gorelick, R. (2012). Mitosis circumscribes individuals; sex creates new individuals. Biology & Philosophy, 27(6), 871–890.
Gould, N. E.-S. J. (2014). Punctuated equilibria: an alternative to phyletic gradualism. Essential Readings in Evolutionary Biology, 239.
Gould, S. J., & Eldredge, N. (1977). Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology, 115–151.
Greenspoon, P. B., & Mideo, N. (2017). Parasite transmission among relatives halts Red Queen dynamics. Evolution.
Hawser, S. P., Baillie, G. S., & Douglas, L. J. (1998). Production of extracellular matrix by Candida albicans biofilms. Journal of Medical Microbiology, 47(3), 253–256.
Henderson, J. (2012). Vitalism and the Darwin Debate. Science & Education, 1–11.
Hoefnagels, M. (2015). Biology: concepts and investigations (3rd ed.). McGraw-Hill New York.
Imwong, M., Snounou, G., Pukrittayakamee, S., Tanomsing, N., Kim, J. R., Nandy, A., … Looareesuwan, S. (2007). Relapses of Plasmodium vivax infection usually result from activation of heterologous hypnozoites. Journal of Infectious Diseases, 195(7), 927–933.
Johnston, C., Martin, B., Fichant, G., Polard, P., & Claverys, J.-P. (2014). Bacterial transformation: distribution, shared mechanisms and divergent control. Nature Reviews Microbiology, 12(3), 181–196.
Karp, G. C. (2013). Cell and Molecular Biology, Concepts and Experiments (7th ed.). USA: Wiley Online Library.
Kavanagh, K. (2011). Fungi, Biology and Applications (2nd ed.). Oxford: Wiley & Sons.
Kutschera, U., & Niklas, K. J. (2004). The modern theory of biological evolution: an expanded synthesis. Naturwissenschaften, 91(6), 255–276.
Lee, S. C., Ni, M., Li, W., Shertz, C., & Heitman, J. (2010). The evolution of sex: a perspective from the fungal kingdom. Microbiology and Molecular Biology Reviews, 74(2), 298–340.
Mackean, D. G., & Hayward, D. (2014). Biology (3rd ed.). IGCSE Cambridge.
Mader, S. S. (2010). Biology (10th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2015). Biology (12th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2018). Essentials of biology (5th ed.). McGraw-Hill Education.
Mason, K. A., Losos, J. B., Singer, S. R., & Raven, P. H. (2014). Biology (7th ed.). McGraw-Hill New York.
Mehlhorn, H. (2016). Animal Parasites, Diagnosis, Treatment, Prevention (7th ed.). Springer.
Moore, J. (1995). The behavior of parasitized animals. Bioscience, 45(2), 89–96.
Naville, M., Warren, I. A., Haftek-Terreau, Z., Chalopin, D., Brunet, F., Levin, P., … Volff, J.-N. (2016). Not so bad after all: retroviruses and long terminal repeat retrotransposons as a source of new genes in vertebrates. Clinical Microbiology and Infection, 22(4), 312–323.
Peacock, L., Bailey, M., Carrington, M., & Gibson, W. (2014). Meiosis and haploid gametes in the pathogen Trypanosoma brucei. Current Biology, 24(2), 181–186.
Pollard, T. D., Earnshaw, W. C., Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell Biology (3rd ed.). Elsevier.
Reece, J. B., Urry, L. A., Wasserman, S. A., Cain, M. L., Minorsky, P. V, & Jackson, R. B. (2014). Campbell Biology (10th ed.). Pearson Higher Ed.
Roberts, L. S., & Javony; J Jr. (2009). Fundations of Parasitology (8th ed.). McGraw-Hill Education.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of Biology (10th ed.). Sinauer & MacMillian.
Simon, E., Reece, J., & Dickey, J. (2013). Essential biology with physiology (4th ed.). Pearson.
Solomon, E., Martin, C., Martin, D. W., & Berg, L. R. (2014). Biology (10th ed.). Cengage Learning.
Speijer, D., Lukeš, J., & Eliáš, M. (2015). Sex is a ubiquitous, ancient, and inherent attribute of eukaryotic life. Proceedings of the National Academy of Sciences, 112(29), 8827–8834.
Speziale, P., & Geoghegan, J. A. (2015). Biofilm formation by staphylococci and streptococci: structural, functional, and regulatory aspects and implications for pathogenesis. Frontiers in Cellular and Infection Microbiology, 5.
Starr, C., Evers, C., & Starr, L. (2013). Biology: Today and Tomorrow With Physiology (4th ed.). Brooks/Cole.
Strick, J. E. (2009a). Darwin and the origin of life: public versus private science. Endeavour, 33(4), 148–151.
Strick, J. E. (2009b). Sparks of life: Darwinism and the Victorian debates over spontaneous generation. Harvard University Press.
Temin, H. M. (1985). Reverse transcription in the eukaryotic genome: retroviruses, pararetroviruses, retrotransposons, and retrotranscripts. Molecular Biology and Evolution, 2(6), 455–468.
Tibayrenc, M., Kjellberg, F., & Ayala, F. J. (1990). A clonal theory of parasitic protozoa: the population structures of Entamoeba, Giardia, Leishmania, Naegleria, Plasmodium, Trichomonas, and Trypanosoma and their medical and taxonomical consequences. Proceedings of the National Academy of Sciences, 87(7), 2414–2418.
Tortora, G., Funke, B., & Case, C. (2010). Microbiology an introduction (10th ed.). San Francisco: Pearson, Benjamin Cummings.
Vergara, D., Jokela, J., & Lively, C. M. (2014). Infection dynamics in coexisting sexual and asexual host populations: support for the Red Queen hypothesis. The American Naturalist, 184(S1), S22–S30.
Vittecoq, M., Elguero, E., Lafferty, K. D., Roche, B., Brodeur, J., Gauthier-Clerc, M., … Thomas, F. (2012). Brain cancer mortality rates increase with Toxoplasma gondii seroprevalence in France. Infection, Genetics and Evolution, 12(2), 496–498.
Volff, J. (2006). Turning junk into gold: domestication of transposable elements and the creation of new genes in eukaryotes. Bioessays, 28(9), 913–922.
Vyas, A., Kim, S.-K., Giacomini, N., Boothroyd, J. C., & Sapolsky, R. M. (2007). Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proceedings of the National Academy of Sciences, 104(15), 6442–6447.
Vyas, A., Kim, S.-K., & Sapolsky, R. M. (2007). The effects of toxoplasma infection on rodent behavior are dependent on dose of the stimulus. Neuroscience, 148(2), 342–348.
Webster, J. P. (2007). The effect of Toxoplasma gondii on animal behavior: playing cat and mouse. Schizophrenia Bulletin, 33(3), 752–756.
Weedall, G. D., & Hall, N. (2011). Evolutionary genomics of Entamoeba. Research in Microbiology, 162(6), 637–645.
Wells, T. N. C., Burrows, J. N., & Baird, J. K. (2010). Targeting the hypnozoite reservoir of Plasmodium vivax: the hidden obstacle to malaria elimination. Trends in Parasitology, 26(3), 145–151.
Yuan, Z., Gao, S., Liu, Q., Xia, X., Liu, X., Liu, B., & Hu, R. (2007). Toxoplasma gondii antibodies in cancer patients. Cancer Letters, 254(1), 71–74.
Yuh, Y.-S., Liu, J.-Y., & Shaio, M.-F. (1997). Chromosome number of Trichomonas vaginalis. The Journal of Parasitology, 551–553.
Zuccala, E. (2015). Evolutionary genetics: Red Queen dynamics in fruit fly sex. Nature Reviews Genetics.
Liu, & Shaio, 1997), el cual se reproduce por mitosis (2).
No hay comentarios:
Publicar un comentario