
Índice
Portada

1. Introducción
|| Regresar al Índice ||
A medida que los seres vivos evolucionaron
a estructuras multicelulares con funciones especializadas, se hizo necesario la
aparición de un tejido, órgano o sistema encargado de coordinar los estímulos y
de emitir respuestas al medio externo o interno. Los dos principales sistemas de
control que presiden las actividades de los cuerpos de los seres vivos son el
sistema nervioso y el sistema endocrino. Estos sistemas trabajan de forma
coordinada cuando ambos están presentes, sin embargo, claramente uno desciende
del otro. Esto se debe a que el sistema endocrino es un sistema mucho más
distribuido filogenéticamente y ha evolucionado de forma independiente en los
seres vivos eucariotas de tipo unicelular.
Por el contrario,
el sistema nervioso real se encuentra ubicado filogenéticamente en los animales
con algún tipo de simetría, excluyendo a los poríferos. La función endocrina es
glandular y depende de la secreción de compuestos químicos que generan efectos
en grupos de células distantes. La función nerviosa es eléctrica y glandular
con efectos endocrinos, pero más comúnmente paracrinos “afecta a células
vecinas”.
Debido a que el
sistema nervioso real presenta una restricción filogenética en los animales,
solo realizaremos la descripción para este linaje de seres vivos, enfocándonos
inicialmente en el sistema nervioso humano por dos motivos (1) nos interesa
médicamente y (2) es la base histórica para la nomenclatura del sistema
nervioso en los demás seres vivos. Al interior de la anatomía humana
discutiremos el fundamento bioquímico de la adicción y la dependencia a
sustancias bioquímicas. En cuanto a la anatomía comparada realizaremos una
descripción superficial sin mayores pretensiones filogenéticas debido a que el
sistema nervioso no deja fósiles con facilidad.
1.1 Santiago
Ramón y Cajal
Santiago Ramón y
Cajal nació el 1 de mayo de 1852 en Petilla de Aragón, España. De niño fue
aprendiz primero de barbero y luego de zapatero. Él mismo deseaba ser artista:
su don para el dibujo es evidente en sus obras publicadas. Sin embargo, su
padre, que era catedrático de Anatomía Aplicada en la Universidad de Zaragoza,
lo convenció de que estudiara medicina, lo que hizo, principalmente bajo la
dirección de su padre. (Más tarde, realizó dibujos para un atlas de anatomía
que estaba preparando su padre, pero que nunca se publicó).
En 1873 se
licenció en Medicina en Zaragoza y se desempeñó, tras un concurso, como médico
militar. Participó en una expedición a Cuba en 1874-75, donde contrajo malaria
y tuberculosis. A su regreso pasó a ser ayudante de la Escuela de Anatomía de
la Facultad de Medicina de Zaragoza (1875) y luego, a petición propia, Director
del Museo de Zaragoza (1879). En 1877 obtuvo el grado de Doctor en Medicina en
Madrid y en 1883 fue nombrado Catedrático de Anatomía Descriptiva y General en
Valencia. En 1887 fue nombrado catedrático de Histología y Anatomía Patológica
en Barcelona y en 1892 fue nombrado catedrático de la misma en Madrid. En
1900-1901 fue nombrado Director del «Instituto Nacional de Higiene» y de las
«Investigaciones Biológicas».
En 1880 comienza
a publicar trabajos científicos, de los que destacan los siguientes: Manual de
Histología normal y Técnica micrográfica, 1889 (2ª ed., 1893). Un resumen de
este manual refundido con adiciones, apareció bajo el título Elementos de
Histología, etc. (Elementos de histología, etc.), 1897; Manual de Anatomía
patológica general (Manual de anatomía patológica general), 1890 (3a ed.,
1900). Además, pueden citarse: Les
nouvelles idées sur la fine anatomie des centres nerux (Nuevas ideas sobre
la fina anatomía de los centros nerviosos), 1894; Textura del sistema nervioso
del hombre y de los vertebrados, 1897-1899; Die Retina der Wirbelthiere (La
retina de los vertebrados), 1894.
Aparte de estos
trabajos, Cajal ha publicado más de 100 artículos en revistas científicas
francesas y españolas, especialmente sobre la estructura fina del sistema
nervioso y especialmente del cerebro y la médula espinal, pero que incluye
también la de los músculos y otros tejidos, y diversos temas en el campo de la
patología general. Estos artículos se encuentran dispersos en numerosas
revistas españolas y diversas revistas especializadas de otros países
(especialmente franceses). Algunos artículos en castellano de Cajal y sus
alumnos aparecen en la Revista Trimestral de Histología normal y patológica
(Revista trimestral de histología normal y patológica) (1888 en adelante),
continuación de ellos aparece bajo el título Trabajos del Laboratorio de
Investigaciones biológicas de la Universidad de Madrid.
Los estudios de
Cajal sobre la estructura de la corteza cerebral han sido parcialmente
agrupados y traducidos al alemán por J. Bresler, 1900-1901. Cajal es también
autor de Reglas y Consejos sobre Investigación Científica, que apareció en seis
ediciones en español y fue traducido al alemán (1933). Entre las distinciones
obtenidas por Cajal se encuentran las siguientes: Miembro de la Real Academia
de Ciencias de Madrid (1895); de la Real Academia de Medicina de Madrid (1897);
de la Sociedad Española de Historia Natural y de la Academia de Ciencias de
Lisboa (1897); Miembro de Honor de la Academia Española de Medicina y Cirugía y
también de varias otras sociedades españolas.

También fue
nombrado Doctor honoris causa en
Medicina de las Universidades de Cambridge (1894) y Würzburg (1896) y Doctor en
Filosofía de la Universidad de Clark (Worcester, EE.UU., 1899). Cajal fue
miembro correspondiente de varias sociedades: la Sociedad Médico-Física de
Würzburg (1895); la Sociedad Médica de Berlín (1895); la Sociedad de Ciencias
Médicas de Lisboa (1896); la Sociedad de Viena de Psiquiatría y Neurología
(1896); la Sociedad de Biología de París (1887); la Academia Nacional de
Medicina de Lima (1897); Conimbricensis
Instituti Societas (Coimbra, 1898); y Miembro de Honor de la Sociedad
Italiana de Psiquiatría (1896) así como de la Sociedad Médica de Gante
(Bélgica, 1900). En 1906 fue elegido miembro asociado de la Academia de
Medicina de París; en 1916 se convirtió en miembro de la Academia Sueca de
Ciencias. Cajàl ha sido galardonado con varios premios, por ejemplo el Premio
Rubio de 1.000 pesetas por sus Elementos de Histología antes mencionados, etc.,
el Premio Fauvelle de 1.500 francos de la Sociedad de Biología de París (1896);
El Premio Moscú de 5.000 francos, establecido por el Congreso de Moscú (1897)
para premiar las obras médicas que, publicadas durante los últimos tres años,
han prestado los mayores servicios a la ciencia y la humanidad, fue otorgado a
Ramon y Cajàl por el Congreso Internacional de Medicina. en París (1900). En
1905, la Real Academia de Ciencias de Berlín le otorgó la Medalla Helmholtz.
Compartió el Premio Nobel de 1906 con Camillo Golgi por su trabajo sobre la estructura del sistema nervioso.
Cajal fue
convocado a Londres para impartir allí, en marzo de 1904, la Conferencia Croonian de la Royal Society y a la
Universidad Clark (Worcester, Mass., EE. UU.) En 1899 para impartir allí tres
conferencias sobre la estructura del cerebro humano y sobre la últimas
investigaciones sobre este tema. En 1952 se publicó un volumen de 651 páginas
«En honor a S. Ramón y Cajal en el centenario de su nacimiento 1852 por
miembros de un grupo de investigación en neurofisiología» en el Instituto
Caroline (Acta Physiol. Scand., Vol. 29, Supl. 106).
En 1879 Cajal se
casó con Doña Silvería Fañanás García. Tuvieron cuatro hijas y tres hijos. De
Conferencias Nobel, Fisiología o Medicina 1901-1921, Elsevier Publishing
Company, Amsterdam, 1967 Esta autobiografía / biografía fue escrita en el
momento de la concesión y publicada por primera vez en la serie de libros Les
Prix Nobel. Posteriormente se editó y se volvió a publicar en Nobel Lectures.
Para citar este documento, indique siempre la fuente como se muestra arriba.
Santiago Ramón y
Cajal murió en Madrid el 18 de octubre de 1934.
2. El sistema nervioso a nivel celular
|| Regresar al Índice ||
2.1 La célula como unidad funcional del
sistema nervioso
La célula es la
unidad estructural y funcional de los seres vivos, las funciones de los órganos
y sistemas no son más que la acción colectiva e interrelacionada de miles de
millones de células. Por tal razón resulta relevante recordar algunas funciones
básicas de cualquier célula. Las células de la mayoría de los órganos basan sus
funciones en los mecanismos de transporte a través de membrana y en los cambios
conformacionales de las proteínas integrales de la membrana.
La secreción
glandular de la neurona y su potencial eléctrico ocurren gracias a los
mecanismos de transporte a través de membrana, mientras que sus mecanismos de
percepción se dan gracias a los cambios conformacionales de las proteínas
integrales de la membrana y a un tipo de transporte entre membranas.
2.2 Transporte de sustancias a través de
membranas
Al igual que pasa
con el transporte de gases, la digestión o la excreción, el sistema nervioso
muchas veces requiere del paso de sustancias a través de membrana, lo cual
involucra procesos de transporte pasivo y de transporte mediado por proteínas.
👉 Transporte
pasivo: Cuando una sustancia en estado acuoso o
disuelto realmente está separada en dos cantidades por una membrana
semipermeable a dicha sustancia, esta fluirá libremente de forma tal que las
concentraciones a ambos lados tiendan a igualarse. La velocidad de flujo es
mayor a medida que la diferencia de concentración –también llamada gradiente
–es más alta, y la velocidad aparenta ser cero cuando las concentraciones son
las mismas.
👉 Transporte a través de membrana mediado por
proteínas: El transporte de gases también involucra
trasladar gases a las malas, ya sea porque se requiere un flujo muy rápido ya
que los tejidos demandan un tipo de gas con gran impaciencia; o a que hay un
gas acumulándose rápidamente que resulta tóxico. El transporte activo emplea
proteínas que gastan energía celular, lo que les permite transportar nutrientes
en contra del gradiente de concentración, mientras que el transporte pasivo
facilitado emplea proteínas que aceleran el paso de nutrientes a favor del
gradiente de concentración.
Algunas proteínas
empleadas en el transporte de nutrientes no están ancladas a la membrana, pero
si colaboran a que las concentraciones en un lado de la membrana sea lo
suficientemente alto como para que el proceso osmótico se acelere. A diferencia
del intercambio de gases, la absorción de nutrientes durante los procesos
digestivos requiere de proteínas, y principalmente de transporte activo, por
tal razón se dice que la digestión es un proceso que gasta energía, y que
algunos alimentos agotan más que otros a la hora de digerirlos.
2.3 Transporte vesicular y la digestión
intracelular
La digestión
puede llevarse a cabo segregando las enzimas al medio íntimo extracelular o el
lumen de un órgano, en cuyo caso se la clasifica como extracelular. En este
tipo de digestión se obtienen nutrientes moleculares que se absorben por los
mecanismos de transporte a través de la membrana externa. No todos los seres
vivos digieren extracelularmente, y por lo tanto deben emplear métodos que les
permita absorber macronutrientes del ambiente íntimo de la membrana y
envolverlos en vesículas digestivas para que en su interior se dé la digestión
molecular final. Una vez procesado, los nutrientes se absorben por los
mecanismos de transporte a través de la membrana vesicular y el material no
digerido se elimina de la célula.
Existen varios
mecanismos especializados basados en el proceso anterior. La digestión
intracelular tiene otros propósitos más allá de los de nutrición, ya que el
sistema inmune hace un uso extensivo e intensivo de la digestión vesicular para
sus mecanismos de presentación de antígeno.
👉 Endocitosis: La endocitosis es el proceso por el cual una célula cuya membrana externa
es flexible rodea a otra partícula, puede ser otra célula u sustancia química,
cuando termina de rodearla genera una vesícula interna llamada endosoma. La
endocitosis puede dividirse en dos categorías principales, la endocitosis no
específica y la endocitosis específica. La endocitosis no específica también denominada
pinocitosis engloba fluidos y todo material que se encuentre diluido en este.
La endocitosis
específica también se denomina endocitosis mediada por receptores. En este caso
la célula o partícula debe tener una serie de proteínas en si membrana que
sirven como marcadores de reconocimiento. La célula que va a realizar la
endocitosis también tiene otras proteínas que sirven como reconocedores.
Cuando ambos marcadores entran en contacto íntimo activan a la célula que
realiza la endocitosis para iniciar el proceso.
Independientemente
del tipo de endocitosis, el resultado final es un endosoma, es decir una
vesícula del sistema de membranas internas de la célula eucariota. Esta
vesícula y su contenido pueden sufrir diferentes destinos dependiendo del
contexto, ya sea en el sistema inmune, en la digestión celular o como una
manipulación de endoparásitos para atacar a la célula.
👉 Fagocitosis: La fagocitosis se puede definir literalmente como un tipo de digestión
interna por parte de una célula, y probablemente sea la primera digestión
interna que existió. La fagocitosis es un proceso más complejo que la
endocitosis, aunque muchas veces empleamos estos dos términos de manera
indiferenciada. Sin embargo, la diferencia está en el tratamiento que recibe el
material endocitado, en la fagocitosis en endosoma “llamado en este caso
fagosoma” recibe un tratamiento más complejo, poco después de que el fagosoma
ingresa a la célula este se fusiona con otra vesícula llamada lisosoma. Los
lisosomas son vesículas que contienen las enzimas digestivas y otras sustancias
químicas que degradan a la víctima.
Cuando el
fagosoma se fusiona con el lisosoma se forma un fagolisosoma, las enzimas
digestivas y las otras sustancias químicas oxidantes empiezan a degradar a la
víctima transformándola en sus componentes básicos. Una vez se han degradado,
los componentes básicos que se extraen de allí serán absorbidos por los
mecanismos de transporte a través de membrana dependiendo de la naturaleza del
nutriente liberado. Este modo de alimentación es muy común en los eucariortes
de vida libre unicelulares como las amebas. En los eucariotes multicelulares,
especialmente en los animales la fagocitosis cumple una función inmune más que
de nutrición.
La eliminación es
el proceso en el cual se desechan compuestos que no fueron absorbidos y que
nunca fueron parte de las rutas metabólicas del ser vivo. Otro proceso parecido
es el de excreción en el cual se desechan compuestos químicos que hicieron
parte de las redes metabólicas y que potencialmente podrían detenerlas de
mantenerse en el cuerpo.
2.4 Cambio estructural de las proteínas
La mayoría de las
proteínas de membrana funcionan mediante las siguientes capacidades: Como
receptores que se unen a sustancias especificas; como canales de transporte de
sustancias especificas; o como agentes que transfieren electrones con niveles
de energía elevados durante los procesos de transferencia de energía. Como los
fosfolípidos de la bicapa, las proteínas integrales de la membrana también son
anfipáticas, poseyendo una porción hidrófoba que se inserta en la membrana
celular y porciones hidrófilas que se exponen hacia el medio interno o externo
de la célula.
Como resultado,
la membrana sigue poseyendo una naturaleza constante, con un interior hidrófobo
y un exterior a ambos lados hidrófilo. Como resultado de esta constancia en la
naturaleza de la membrana, sus propiedades de permeabilidad a las diferentes
sustancias, permanece constante. Es importante destacar que los lípidos de
membrana que se encuentran asociados de manera íntima con la proteína pueden
afectar su funcionamiento, aunque el modo y el impacto de estas modificaciones
aún permanecen como algo poco claro. La
porción de la proteína integral que se proyecta hacia el citoplasma y hacia el
medio tiende a ser como las proteínas globulares es decir de naturaleza soluble
en agua o hidrófilas.
Una gran cantidad
de proteínas forman literalmente conductos de paso, canales para el flujo de
sustancias a través de la bicapa de lípidos. Bajo el modelo del mosaico fluido,
las proteínas integrales no se encuentran fijadas de manera rígida en una zona
de la membrana, si no que “flotan” (si es que podemos usar esa idea) a la
deriva en la matriz de los fosfolípidos. Por lo general una proteína integral
de membrana tiene tres dominios. Un dominio extramembranal de cara al exterior
que funciona como receptor de ligandos específicos, un dominio intramembranal
que puede cambiar de forma al activarse el receptor ya sea para transdusir una
señal o como un canal iónico. Finalmente, un dominio intramembranal que
puede funcionar como una cisterna interna del canal iónico o como un dominio
efector metabólico que dispara señales químicas al interior de la célula.
Los canales no
requieren el dominio receptor y funcionan por otros estímulos como los
gradientes de concentración pasivos.
3. El citoesqueleto en el sistema nervioso
|| Regresar al Índice ||
El sistema nervioso evoluciona del sistema endocrino y ambos emplean el citoesqueleto como su principal sistema de coordinación, así como el sistema que le da la forma a las células neuronales.

Figura 3.1. Transporte vesicular en una neurona. Las
vesículas que transportan los neurotransmisores, neuro-hormonas y hormonas
emplean el citoesqueleto de la neurona como autopista para llegar a la punta
terminal del axón, donde se mantienen en espera de una señal.
3.1 Alargamiento axonal
En 1907 Ross
Harrison de la universidad de Yale demostró dos cosas extrayendo tejido
neuronal de un embrión de rana y poniéndolo a cultivar con fluido linfático, la
primera es que los tejidos extraídos de un ser vivo podían mantenerse vivos
fuera de su contexto original, y la segunda que los axones neuronales podían
crecer por un sistema de desarrollo muy activo. En este caso los axones jóvenes
son muy delgados y crecen gracias al desarrollo del chasis de microfilamentos,
los cuales indagan el medio en busca de hormonas que les indiquen la dirección
a seguir, una vez que encuentran un rastro siguen creciendo, mientras que en la
base de la proyección se empiezan a importar microtúbulos que engrosan la
proyección para formar el cuerpo de un axón nuevo más firme. De este modo los
axones van creciendo de forma coordinada para encontrar las hormonas liberadas
por otras neuronas y así poder conectarse en sinapsis íntimas.
3.2 Filamentos intermedios
Estos recorren la
superficie del polo del lumen de la célula sirviendo como el marco que define
la célula y que permite identificar la parte del citiplasma que se encuentra en
los cilios y la parte del citoplasma celular general, su función es la de darle
su forma a la célula epitelial, por lo que estos filamentos se proyectan no
solo en el polo del unen sino que discurren por toda la superficie de la
célula, siendo algo semejante al marco que le da su forma a la carpa de un
circo.
3.3 Microtúbulos
Recorren la
célula y conectan a los diferentes organelos, funcionan como rieles de carga
entre un organelo y otro sobre el cual se movilizan proteínas de transporte
especializadas que jalan las vesículas desde un sistema de membranas a otro de
forma rápida y no estocástica. En este sentido los microtúbulos permiten la
organización, transporte, y coordinación de las funciones celulares.
3.4 Quinesinas
Son las proteínas
activas del citoesqueleto que caminan a través de los microtúbulos “literalmente”
estas son las grúas moleculares que transportan las vesículas y que permiten el
tráfico de membranas en cualquier célula eucariota. Estos elementos se
encuentran en todas las células eucariotas con niveles de especialización
variables, por lo que el modelo de la célula epitelial polarizada se emplea más
con objetivos didácticos.
3.5 Célula nerviosa
En las células
nerviosas la red de rieles de microtúbulos están asociados a los filamentos
intermedios, aunque la estructura y función de los componentes es análoga a los
de la célula epitelial polarizada. En este caso la polarización se da hacia el
núcleo neuronal del cual parte los diferentes rieles por cada una de las
dendritas.
4. Anatomía de la neurona
|| Regresar al Índice ||
A diferencia del músculo o el hueso con el sistema nervioso si tenemos cierta diferencia semántica entre la neurona como célula, el nervio como tejido y el sistema nervioso como sistema. En la presente sección discutiremos la estructura de la neurona aislada a nivel anatómico y a nivel fisiológico poniendo énfasis en los mecanismos que conllevan a la transmisión del impulso eléctrico o potencial de acción.

Figura 4.1. Anatomía de la neurona. Se
trata de una célula animal modificada con extensiones de citoplasma muy largas,
las cuales aumentan su área de superficie membranal, por lo que requieren
grandes cantidades de lípidos para poder ser producidas.
También
analizaremos los fundamentos moleculares que dan funcionamiento a la célula
neuronal, los cuales se pueden distinguir en dos tipos: la transmisión de unos
potenciales eléctricos a través de la membrana celular y la acción glandular
paracrina sobre tejidos adyacentes. Cabe destacar que ninguno de estos dos
procesos emplea estructuras absolutamente novedosas, todo gira a los fundamentos
de cualquier célula, transporte a través de membranas y movimiento de
vesículas, acción de proteínas mediante acoplamiento a ligandos específicos y
cambios conformacionales.
Lo que sucede con
todas estas piezas en la neurona es su enfoque, la neurona se encuentra
altamente especializada para que todos estos elementos celulares estén
dispuestos para transmitir señales electroquímicas a grandes velocidades. Finalmente cabe destacar que fenómenos como
el transporte con vesículas, la exocitosis o los canales iónicos han sido
estudiados con más profundidad en la neurona, y estos principios son los que se
han generalizado para el estudio de otras células como para la instrucción
nivel escolar.
En esta discusión
emplearemos la neurona humana como la neurona tipo, aunque evidentemente esto
es antropocentrista resulta más fácil hablar de las diferencias, si es que las
hay, con respecto a la neurona humana cuando se discute en la filogenia. Las neuronas son el elemento primordial del sistema
nervioso. La estructura y funciones de las neuronas permiten la transmisión
rápida de estímulos de una célula a otra mediante una combinación de pulsos
eléctricos y endocrinos. La neurona es una célula extremadamente especializada
enfocada en dos funciones primordiales, la secreción endocrina de sustancias
químicas y la transmisión de un impulso eléctrico a grandes distancias.
El cuerpo de la
neurona se denomina soma o pericarión, allí se almacena el núcleo y la mayoría
de los organelos de síntesis. Las proyecciones de la neurona que se dispersan
como raíces o tentáculos son denominadas dendritas. La membrana del soma posee
pocas proteínas generadoras del impulso eléctrico llamado potencial de acción,
aunque son capaces de realizar impulsos más débiles y lentos llamados potenciales
de membrana. En cualquier caso, una señal enviada al soma se hace más lenta.

Figura 4.2. Arriba: podemos
ver al cuerpo celular de la neurona o soma coloreado de morado y el axón en
rojo. En la microfotografía de abajo: las neuronas se encuentran en una etapa
de síntesis casi permanente, sus núcleos no muestran cromosomas ya que estos
solo se manifiestan cuando la célula va a reproducirse. En su lugar se
encuentra el nucleolo que es la fábrica de ribosomas, organelos empleados en la
fabricación de proteínas.
4.1 La dendrita, el axón, y la célula glial
Las dendritas y los axones se ven iguales en los modelos, siendo elongaciones de citoplasma que se alejan del núcleo, la diferencia radica en la función, específicamente en la dirección de transmisión de la señal. Las dendritas reciben impulsos electroquímicos de otras neuronas, y las llevan hacia adentro y hacia el soma, mientras que los axones llevan los impulsos lejos del soma. ... Generalmente, las dendritas reciben señales neuronales y los axones las transmiten. La mayoría de las neuronas tienen muchas dendritas (receptores de información) y solo tienen un axón (emisor de información).

Figura 4.3. La célula de Schuann. Son
células planas que se enroscan en varias capas alrededor de una dendrita, estas
capas se llaman mielina, aunque no son las únicas células capaces de formar
mielina.
Las dendritas/axones se encuentran aisladas del medio externo median células gliales. Como consecuencia no podemos hablar de las neuronas como un tejido aislado y homogéneo, la unidad funcional neuronal involucra de por si la conjunción de varios tipos de célula. Las células gliales de hecho son un grupo heterogéneo de distintos tipos de células con funciones altamente especializadas.

Figura 4.4. Oligodendrocitos. Los
oligodendrocitos son capaces de formar varias capas de mielina para dendritas
diferentes.
Al conjunto de dendrita más células gliales lo llamamos axón, este transmite la información a través del cuerpo de la célula a altas velocidades mediante señales eléctricas denominados potenciales de acción. Los axones pueden llegar a ser muy largos y por lo tanto se especializan en la transmisión de señales a largas distancias en poco tiempo, lo cual contrasta con la función glandular que transmite señales a velocidades más limitadas. Dos linajes de células gliales se asocian a los axones dependiendo del lugar donde se encuentra la dendrita, en el Sistema Nervioso Central (SNC) las dendritas están rodeadas por oligodendrocitos, mientras que los nervios que se ramifican hacia el cuerpo como los músculos y los demás órganos “Sistema Nervioso Periférico SNP” están rodeados por células gliales llamadas células de Schwann.

Figura 4.5. Otras células gliales. No
todas las células gliales forman mielina, otras se encargan de nutrir a las
neuronas, reciclar neurotransmisores, mantener el medio estable y protegerlo
contra infecciones.
Las células
gliales proveen un apoyo estructural y bioquímico a las funciones neuronales.
Algunos axones están envueltos en más de una capa de la membrana de la célula
glial produciendo la denominada capa de mielina, la cual actúa como aislante al
medio externo de la neurona. A parte de las células gliales formadoras de
mielina existen otras células de tipo glial encargadas de otras funciones de
apoyo, entre estas cabe destacar a las microgliales y astrocitos involucrados
en la función inmune. Se ha reportado que el VIH es capaz de infectarlas.
Las células
gliales no se extienden indefinidamente como las dendritas, varias células
gliales envuelven la neurona como los vagones de una locomotora, lo cual
implica la presencia de articulaciones o puntos donde las células gliales no
cubren a la dendrita llamados nódulos de Ranver. La combinación de la mielina y
los nódulos permiten que los potenciales de acción puedan ser conducidos mayor
velocidad.
4.2 La sinapsis neuronal
La región
terminal de un axón almacena vacuolas cargadas con compuestos químicos de
actividad glandular paracrino o endocrino, estas sustancias reciben el nombre
específico de neurotransmisores. Originalmente se los clasificó de forma
diferente a otros productos glandulares llamados hormonas, pero en la
actualidad se los percibe como un mismo tipo de sustancias debido a que sus
mecanismos de liberación y de acción son bastante similares.
Cuando un potencial de acción llega a la punta de un axón las vacuolas almacenadas son estimuladas ara realizar el proceso de exocitosis, liberando sus neurotransmisores al medio intersticial de las neuronas. La membrana de las neuronas o del tejido afectado posee proteínas con sitios activos específicos para atrapar los neurotransmisores y realizar el proceso de cambio estructural de una proteína membranal. Los neurotransmisores son un grupo heterogéneo de moléculas, cada tipo de señal emite un neurotransmisor diferente, de hecho, mucho del estudio de la función holística del cerebro depende de conocer qué tipo de neurotransmisores media alguna sensación, esto es importante especialmente para el estudio de las adicciones.

Figura 4.6. La sinapsis. Las
sinapsis químicas se comunican por medio de moléculas solubles llamadas
neurotransmisores. estas viajan por difusión simple en el medio acuoso entre la
sinapsis efectora y la sinapsis receptora. Existe un segundo tipo llamado
sinapsis eléctrica, pero no es tan común, donde la unión entre las dos neuronas
es mucho más íntima.
4.3 Organelos neuronales
Las neuronas
poseen requerimientos metabólicos especiales. Los axones de algunas neuronas
pueden extenderse hasta 1 metro de longitud y considerando los del soma o de
una célula eucariota promedio, la extensión de los axones implica la existencia
de una membrana celular muy larga, así como un contenido citoplasmático
bastante grande en comparación con el minúsculo y alejado núcleo. El soma de la neurona
agrupa las fábricas moleculares de la célula, el núcleo y especialmente los
retículos endoplasmáticos y el aparato de Golgi, las dendritas carecen de
dichos organelos. El núcleo de una
neurona es grande, y una cantidad considerable de su material genético se
encuentra en proceso de expresión de forma constante, lo anterior se debe a que
un solo núcleo debe mantener funcionando a una dendrita muy larga.
Basados en
estudios de hibridación se ha estimado que cerca de un tercio del genoma de las
células cerebrales se encuentra en su forma activa para la síntesis de
proteínas, básicamente las neuronas sintetizan más ARN mensajeros que cualquier
otra célula del cuerpo. Debido a las
altas tasas de transcripción, la cromatina del núcleo se encuentra dispersa, lo
cual permite que las proteínas de transcripción se acoplen y generen los ARN
mensajeros.
La mayoría de las
proteínas formadas por los ribosomas libres permanecen en el cuerpo de la
neurona, mientras que las proteínas sintetizadas en los ribosomas acoplados al
retículo endoplasmático rugoso son exportadas hacia las dendritas por medio de
vesículas. Las vesículas pueden ser enviadas directo a la dendrita o ser
enviadas al aparato de Golgi, esto último es lo más común. El retículo
endoplasmático rugoso en la neurona es bastante conspicuo, aun en micrografías
bastante simples se lo puede apreciar rodeando al núcleo como si fuera una
serie de capas muy densas.

Figura 4.7. El soma neuronal. Las
neuronas están en etapa de síntesis de proteínas permanente, por ende, su
sistema de membranas internas se encuentra hinchado por su sobreproducción y son
fácilmente visibles al microscopio óptico.
Al igual que
otras células, el aparato de Golgi se encarga de adicionar azúcares complejos a
las proteínas para que estas puedan realizar sus funciones. Las proteínas
activas son liberadas al citoplasma envueltas en partes de la endomembrana del
aparto de Golgi, es decir en vesículas que viajan hacia la punta de las
dendritas. Las dendritras no poseen ribosomas, retículos endoplasmáticos
rugosos ni aparatos de Golgi, por lo que dependen exclusivamente de los paquetes
o vesículas llenas de materiales enviadas desde el soma.
4.4 El citoesqueleto de una neurona
La forma
altamente especializada de la neurona y la habilidad de esta célula para
transportar proteínas y otros componentes estructurales depende de la presencia
de un citoesqueleto externo e interno. Especialmente el citoesqueleto interno
sirve no solo para dar forma, también proporciona señales químicas de dirección
para que las vesículas sintetizadas en el soma de la neurona puedan ser
transportadas en la dirección correcta hacia las dendritas.
👉 Microfilamentos: Están compuestos por actina, una proteína estructuralmente semejante a la actina muscular, al igual que la actina muscular forma filamentos cortos con funciones estructurales formando coos dendríticos donde es necesario o apoyando la movilidad y el crecimiento de una dendrita en formación. Son filamentos dispersos en el citoplasma, empleados en donde son requeridos.

Figura 4.8. El citoesqueleto de la neurona. A
parte de las funciones ordinarias del citoesqueleto, en la neurona este cumple
la función de riel de transporte para las vesículas emitidas por el aparato de
Gilgi, esto se logra mediante grúas moleculares con base en quinesina o dineina
(YouTube)(YouTube), las cuales funcionan de forma
semejante a la miosina.
👉 Neurofilamento: Son la columna vertebral de una dendrita,
proporcionan la mayor parte de la rigidez de la dendrita, adicionalmente
desprendiéndose del cilindro principal el neurofilamento posee ramificaciones
laterales que proveen la forma a la dendrita en su diámetro.
👉 Los microtúbulos: Proveen el marco de referencia de señalización para el transporte de
materiales entre la sinapsis y el soma de la neurona, no son fibras, sino
proteínas que emplean las fibras del citoesqueleto para transportar material.
👉 Quinesinas: Las grúas moleculares emplean dos cabezas
que actúan como "pies", pero su movimiento requiere
energía, por tal razón las dendritas si poseen una población bastante grande de
mitocondrias, lo cual hace del tejido neuronal uno de los que
requieren mayor energía para existir.
4.5 Mecanismos de transporte neuronales
Las proteínas,
organelos y otros materiales celulares deben ser transportados a través de la
neurona para mantener la integridad estructural y las funciones celulares
basales. Esto se complica por varios factores, la maquinaria de síntesis de
proteínas se encuentra en el soma, mientras que las dendritas con centímetros
de longitud solo reciben los materiales creados en el soma. Para compensar, la
neurona posee mecanismos de transporte para la movilización en direcciones
sinapsis-soma y viceversa soma-sinapsis. Los microtúbulos poseen proteínas
asociadas encargadas de transportar las sustancias como la quinesina y la
dineina.
El transporte
Soma-Sinápsis se denomina transporte anterógrado y posee dos tipos, rápido y
lento. El transporte lento se emplea para transportar la mayoría de las
sustancias pequeñas o suborganélicas como las proteínas estructurales, la
actina, los neurofilamentos y los microtúbulos. Estos materiales se emplean
para el mantenimiento y reconstrucción de las dendritas. El transporte rápido
mueve sustancias doscientas veces más raído y se emplea para movilizar
organelos, vesículas y glicoproteínas membranales empleadas en las sinapsis.
El transporte
Sinapsis-Soma es extremadamente lento, cinco veces más lento que el transporte
lento del soma a la sinapsis y mil veces más lento que el transporte rápido del
soma a la sinapsis. El transporte retrogrado de la sinapsis al soma es
importante, ya que retroalimenta los estímulos membranales al soma y recicla
materiales como como membranas y partes de neurotransmisores degradados. Estas
señales de retroalimentación activa al núcleo para sintetizar más sustancias y
hacer crecer sus dendritas. Defectos en el transporte ultralento de la sinápsis
al soma han sido vinculados a algunas enfermedades neurodegenerativas.
5. Propiedades eléctricas de la membrana
|| Regresar al Índice ||
Los estímulos
para la marcha lenta de una señal en un potencial de membrana dependen de
algunas propiedades eléctricas. A continuación, mencionaremos brevemente
algunas de estas propiedades. Este cambio localizado en el potencial de
membrana posee componentes tanto de tiempo como de espacio. Las propiedades
eléctricas de las membranas biológicas gobiernan como las perturbaciones en el
estado de reposo se transfieren a través de la superficie de la neurona.
5.1 Conductancia
membranal
Es la facilidad
con la que los iones cruzan la membrana, a mayor conductancia más iones fluyen
a través de la membrana. La conductancia para un nervio es la suma de las
conductancias individuales de los iones importantes en el establecimiento de la
diferencia de potencial. Evidentemente esto depende de la cantidad de canales
iónicos presentes en la membrana.
5.2 Resistencia
membranal
Opuesto a la
conductancia y se mide en siemens. Para una proteína de trasporte iónico
individual en una solución iónica dada la conductancia es un valor constante
determinado parcialmente por el tamaño del ión con respecto al canal y la
distribución de cargas de las paredes del canal proteínico.
5.3 Dispersión
eléctrica
Una perturbación
en el potencial de membrana se dispersa a lo largo de la superficie
disminuyendo proporcionalmente en magnitud a medida que se aleja del punto de
origen. El cambio de voltaje en cualquier punto a lo largo de la membrana
puede modelarse matemáticamente como una función de la corriente y la resistencia
mediante la ley de Ohm. La razón para la disminución del cambio de voltaje con
el área de dispersión es precisamente que el área aumenta en una función
cuadrática, mientras que la cantidad de iones es relativamente la misma,
dejando menos iones para perturbar en distancias más largas. Este tipo de
propagación es denominada conductividad eléctrica o conductividad pasiva.
5.4 Capacitancia
membranal
Es la capacidad
para almacenar diferencias de carga. Un capacitor consiste en dos conductores
separados por un aislante, uno de los conductores almacena cargas positivas y
otras cargas negativas. En el contexto biológico la membrana biológica es el
aislante, y las matrices acuosas de la matriz intercelular y del citoplasma son
los dos conductores. La capacitancia se mide en faradáis. El producto de la
capacitancia y la resistencia de la membrana determinan el tiempo que tarda un
estímulo en causar un cambio en el estado de reposo de la membrana.
5.5 Potenciales eléctricos neuronales
Seamos claros, en la membrana de la neurona lo que no corre son los electrones, pero si la definición de corriente eléctrica es la de un flujo de electrones, como le hace una neurona para ser, bueno ya saben, ¡eléctrica! Esto está ligado con el asunto del potencial, debido a que la corriente eléctrica se encuentra comúnmente ligada a la de un flujo de electrones a través de un medio, el potencial eléctrico medido en voltios se define análogamente a la presión de electrones en dicho medio, pero nuevamente no hay flujo de electrones en la membrana de la célula, por lo menos no de electrones desnudos.

Figura 5.1. Potencial de acción. La
expresión potencial hace referencia de forma más correcta a una energía
potencial lista para ser disparada, una neurona cargada posee potencial, una
neurona descargada debe construir su potencial aplicando energía y separando
los iones por sus cargas positivas y negativas.
El punto es que
las analogías clásicas sobre la electricidad no aplican a la neurona, a menos
que uno sepa cómo funciona una batería, pero eso es poco común, pero eso
simplemente implica que la electricidad es un fenómeno mucho más complejo que
el que permite visualizar la analogía clásica de la corriente de agua, por lo
que para entender la fisiología neuronal hay que borrar dicha analogía de
nuestras cabezas. En la neurona no hay una corriente de electrones libres, pero si existe
flujo de cargas. Por lo general quien fluye son las cargas positivas a través
de portadores en forma de iones con cargas positivas como el ion sodio(1+) y el
ion potasio(1+) con respecto a portadores estáticos de cargas negativas como
las proteínas y el ion cloro(1-).
En este sentido
definiremos como corriente eléctrica al flujo de portadores de carga de un lado
al otro de la membrana, en otras palabras, al flujo de iones. En este contexto
el potencial eléctrico se define no como la presión a través de un tubo sino
como la diferencia de cargas a ambos lados de la membrana, y nuevamente se mide
en voltios.
Por lo general
una neurona en descanso presenta un potencial ya formado, lo cual implica dos
cosas: (1) la existencia de una corriente pasiva que ha formado el potencial y
(2) la posibilidad de generar una corriente retrograda fuerte que restaure el
equilibrio de cargas. Las leyes de las cargas nos dicen que entre mayor es el
potencial, es decir a mayor diferencia de cargas se genera una fuerza creciente
que induce a que las cargas se igualen, esa fuerza es precisamente el potencial
o energía eléctrica.
6. Potencial de membrana
|| Regresar al Índice ||
(YouTube) Normalmente representamos la transmisión de información en una
neurona como si se tratara de un cable eléctrico por el cual fluyen electrones,
pero eso es un mito. Lo que fluyen son iones, sustancias químicas simples con
carga a través de canas iónicos en la membrana.
Los potenciales
eléctricos a través de membrana son la base de la función neuronal. Uno puede
visualizar a una neurona como una célula glandular muy larga. Recibe
información por hormonas en un lado, trasmite la información a su región
interna, y luego emite una respuesta glandular en otro lado. El problema es que
las neuronas son ¡Muy LARGAS! La transmisión química de una señal a través de
reacciones en el citoplasma es útil para las glándulas y otros tejidos donde
las señales deben viajar cortas distancias sin que exista un control detallado,
pero las neuronas son otra historia. La longitud de su citoplasma hace que la
transmisión metabólica de una señal sea inefectiva y allí es donde ingresan los
pulsos eléctricos de las neuronas.
El pulso
eléctrico avanza más rápido que una molécula vibrando estocásticamente en un
fluido, y tiene la ventaja de no tener que transmitir su información al aparato
de Golgi por medio de vesículas. Las neuronas emplean el pulso eléctrico para
acelerar el transporte de una señal entre dos puntos comunicados
glandularmente. El asunto es ¿Cuál es la naturaleza eléctrica de la neurona?
Por lo general cuando hablamos de pulso eléctrico pensamos en corriente
eléctrica, la cual de forma estándar se define como una corriente de
electrones. Sin embargo, esta definición es solo práctica en metales donde los
electrones pueden fluir fácilmente, pero en la célula no hay metales puros para
un flujo de electrones, por lo menos no para la neurona.
Lo más parecido a
una corriente eléctrica es la cadena de transporte de electrones que finaliza
el ciclo de Krebs, sin embargo, el funcionamiento eléctrico de la membrana
neuronal no emplea los principios de transporte de electrones, de hecho, la
palabra electrón es lo que menos escucharemos cuando describamos la fisiología
de la membrana neuronal. Por lo anterior antes de involucrarnos con la anatomía
ajustaremos un poco las analogías empleadas a la hora de hablar de fuerza
eléctrica y potencial en el contexto de la neurona.
Las neuronas
poseen un potencial eléctrico generado por la membrana externa y en
consecuencia se la referencia como el potencial de membrana. El potencial de
membrana puede medirse mediante la inserción de un microelectrodo en una célula
nerviosa. El voltaje al interior de la célula es comparado con el valor detectado
mediante un electrodo localizado en la matriz extracelular. No existe un marco
de referencia absoluto, el valor que arroja la matriz se asume arbitrariamente
como cero. Los valores promedio para una neurona arrojan generalmente valores
dentro del intervalo de -60mV a -90mV y es denominado como el potencial de
membrana en reposo. Cabe destacar que las neuronas no son las únicas células capaces de formal
potenciales eléctricos, los miocitos o células musculares también poseen dichos
mecanismos y al igual que en las neuronas, el pulso eléctrico tiene como objeto
primordial la liberación de iones calcio (2+). Los iones calcio(2+) son
empleados para generar respuestas al interior de las células, en las neuronas
activa su función neuronal, en los miocitos provoca la contracción del
sarcómero. Para el caso de la neurona, los potenciales de membrana deben
generarse mediante canales iónicos, proteínas especializadas que permiten el
flujo de iones a través de la membrana, por ejemplo, la bomba de sodio y
potasio.
6.1 La bomba de sodio y potasio

Figura 6.1. Bomba de sodio y potasio. La
bomba de sodio y potacio funciona como un cargador de baterías, aumentando el
potencial eléctrico en dos contenedores, en este caso, los dos lados de una
membrana, la energía no se extrae de un enchufe, sino del ATP que se obtiene a
partir de la glucólisis y la fosforilación oxidativa de la cadena de transporte
de electrones. La bomba de sodio y
potasio tiene tres dominios de acople principal: El primer dominio acopla tres
unidades del ion sodio (1+) en el interior de la célula, el segundo dominio
acopla dos unidades del ion potasio(1+) en el exterior de la célula, y el
tercer dominio acopla ATP, la unidad portadora de energía de la célula (YouTube).
Uno de los
principales mecanismos empleados por la célula neuronal para crear una
diferencia de cargas a ambos lados de la membrana es mediante la bomba de sodio
y potasio. Junto con otras proteínas de transporte transmembranal, la bomba de
sodio y potasio contribuye a que la célula se encuentre en un estado de
desequilibrio eléctrico y de concentraciones con respecto al medio circundante,
esto genera fuerzas que pueden ser empleadas para diversos propósitos.
Un ejemplo
relacionado es el uso que hace la célula de gradientes electroquímicos en base
al ion protio(1+) con el cual es capaz de mover mecánicamente proteínas tipo F
para la síntesis de energía. En este caso, la célula mantiene un estado de
desequilibrio estable, o lo que llamaríamos en física clásica, un estado de
equilibrio muy inestable, el sistema en apariencia se encuentra en un estado
inactivo, pero a cualquier estímulo la inestabilidad se manifiesta con una energía
mucho mayor de la empleada para haber realizado el estímulo. A esto lo
denominaremos estado de estabilidad.
Normalmente las
concentraciones intracelulares del ion sodio(1+) “10 milimol por litro en una
célula muscular” es mucho más baja que la concentración extracelular
“140milimol por litro”, en consecuencia hay una presión osmótica sobre la
célula para que ingrese el ion sodio(1+) pero existe un mecanismo que lo extrae
con mayor eficacia. La entrada del ion sodio(1+) es permitida por canales iónicos
sin cisternas, lo cual implica que siempre se encuentran abiertos. La salida se
encuentra mediada por una proteína de transporte transmembranario conocida
comúnmente como la bomba de sodio y potasio.
La bomba de sodio
y potasio se emplea para transportar sodio al exterior y potasio al interior de
forma selectiva creando un gradiente de concentraciones químico y eléctrico.
Químico porque cada ion genera una presión osmótica contra la membrana para
intentar igualar sus concentraciones individuales. El funcionamiento de la
bomba de sodio y potasio es cíclico.
Aunque la carga
de ambos iones es la misma, la cantidad de iones que se mueve es desigual,
hacia un lado de la membrana se transportan más cargas positivas, lo cual con
el tiempo contribuye a una diferencia de cargas. Como mencionamos antes, la
generación de un diferencial de cargas “potencial eléctrico” puede definirse
como una corriente eléctrica a través de la membrana, y el aumento del
diferencial de cargas incrementa el voltaje o potencial eléctrico de la
membrana.
6.2 Creando el potencial de membrana
El potencial de
membrana en reposo también llamado potencial de membrana o potencial en reposo
refleja un equilibrio inestable entre los iones a ambos lados de la membrana,
así como la diferencia de cargas, un lado de la membrana es más positivo que el
otro. Esta diferencia iónica es denominada gradiente de concentración o
diferencia de concentración iónica y es mediada por proteínas de transporte
activo. En una célula excitable como la neurona o el miocito, las
concentraciones de potasio(1+) al interior de la célula es mucho mayor que al
exterior de la célula, mientras que las cantidades de sodio(1+) al interior son
menores que al exterior de la célula. Esta diferencia de concentraciones se
debe a la acción de una proteína de transporte activo denominada bomba de sodio
y potasio, cuyo mecanismo de acción ya describimos en el artículo anterior.
El movimiento desigual de los iones potasio(1+) “dos unidades” y sodio(1+) “tres unidades” crea una diferencia osmótica y eléctrica, sin embargo la diferencia eléctrica generada por la bomba de sodio y potasio es insuficiente para explicar el alto voltaje “potencial eléctrico” que se mide al interior de la célula. Para establecer la diferencia de cargas o potencial eléctrico la membrana en estado de reposo posee canales pasivos que permiten que el ion potasio(1+) fluya al exterior de la célula. Estos canales se denominan rectificadores del ingreso de potasio, son canales de transporte pasivo que transfieren al potasio hacia el exterior de la célula en favor de su gradiente de concentración, pero incrementando el desequilibrio voltaico. Cuando el desequilibrio voltaico alcanza su límite la fuerza eléctrica impide que los iones potasio(1+) fluyan facilmente y los comienza a enviar devuelta al interior de la neurona.

Figura 6.2. Creando el potencial de membrana. En
el modelo anterior podemos apreciar como el potencial de membrana se mantiene
principalmente por las bombas de sodio y potasio "izquierda" y por
los canales de rectificación de ingreso de potasio "derecha", aunque
también existen canales de sodio, estos en su mayoría mantienen sus cisternas
cerradas de forma tal que ingresa muy poco sodio en comparación del que la
bomba expulsa fuera de la neurona.
Este punto de
equilibrio donde la cantidad de potasio(1+) que sale por los canales
rectificadores por la fuerza química es igual a los iones de potasio(1+)
que ingresan por la fuerza eléctrica se denomina potencial
de membrana. El desequilibrio voltaico se genera debido a que los portadores de carga
negativa permanecen inmóviles, ya sean los iones cloro(1-) al exterior de la
neurona o las proteínas al interior de la neurona.
En resumen,
llegados a este punto la membrana posee una diferencia de concentraciones iones
sodio(1+) y de cargas “interior negativo y exterior positivo”, lo cual trae una
tensión sobre la membrana a nivel osmótico y eléctrico también conocido como
potencial electroquímico.
6.3 Función del potencial de membrana
Bueno, una corriente de iones crea un diferencial voltaico en la membrana celular, ¿y para qué sirve eso? Los potenciales de membrana son estados de equilibrio inestable con dos cambios posibles. Haciendo la analogía con un automóvil, es como si tuviéramos un coche de dos tiempos y neutro. Neutro es el potencial de membrana listo para arrancar. En el segundo tiempo o marcha lenta ocurre un estímulo que afecta el flujo de iones a través de la membrana, desviando las concentraciones por fuera del punto de equilibrio quimioeléctrico. Esta primera marcha posee una serie de propiedades, por ejemplo, dispersa su desequilibrio desde un punto de origen de forma circular.

Figura 6.3. Potencial de membrana en reposo. El
potencial de membrana en reposo funciona como un estado neutro en el cual la
neurona reconoce que no hay estímulo mediante la conformación de proteínas al
interior de la célula. Cuando hay un estímulo que depolariza la membrana
parcialmente, las proteínas cambian de forma y transfieren información al
metabolismo neuronal para que esta reaccione de alguna forma. Este es un modo
de transferir información, sin embargo, no es el que se encarga de enviar
mensajes a largas distancias.
El modo en que
esta señal se dispersa está gobernado por las propiedades de la membrana con
respecto a la conductividad de los iones, lo cual en últimas está gobernado por
canales iónicos. Los fisiólogos han logrado determinar las propiedades
eléctricas de una membrana bajo los mismos términos de un dispositivo eléctrico
y medirlos bajo el sistema internacional de unidades.
Independientemente
de dichas definiciones, unidades y ecuaciones, dos propiedades emergen de estos
estímulos en marcha lenta: (1) la señal viaja lentamente a través de la
membrana, (2) la fuerza del estímulo se debilita a medida que el desequilibrio
se aleja del punto de estímulo. En otras palabras, un estímulo en potencial de
membrana viaja lento y viaja poco, lo cual lo hace poco efectivo para
transmitir una señal a largas distancias a través de una neurona, aunque
nuevamente, solo es la primera velocidad en nuestra analogía de un auto.
7. Potenciales de acción
|| Regresar al Índice ||
(YouTube) La neurona puede describirse como un coche de
tres marchas, neutro, lento y muy rápido. Neutro hace referencia al potencial
de membrana no estimulado generado por la bomba de sodio y potasio, así como
por los canales de rectificación del potasio, los cuales crean una diferencia
de iones sodio “más afuera de la membrana” y una diferencia neta de cargas o
potencial eléctrico “negativo al interior de la membrana”. La segunda marcha es
cuando un estímulo afecta el estado de equilibrio del potencial de membrana
también conocido como conductividad pasiva, transmite señales lentamente y a
distancias limitadas.
El potencial de
acción por el contrario es la marcha rápida, transmite señales a gran velocidad
y a distancias muy largas, por lo que es el mecanismo que transmite información
a través de las larguísimas dendritas de una neurona. El potencial de acción se
causa cuando ocurre una fuerte despolarización de la membrana, lo cual implica
un rápido flujo de cargas a través de la membrana para que la diferencia de
cargas se anule, nuevamente este flujo de cargas es lo que en este contexto
definiremos como corriente eléctrica. Como tal el potencial de acción se define
entre dos momentos, el inicio de la despolarización fuerte de la membrana y la
restauración del potencial de acción.
La propiedad
principal de los potenciales de acción es que se genera a sí mismo a medida que
se propaga a través de la superficie de la membrana. De esta manera, una señal
generada en el soma de la célula puede viajar a través de toda la célula hasta
la punta de un axón localizado centímetros de distancia, una longitud enorme
desde la perspectiva de una célula.
7.1 Generando el potencial de acción
Generar un
potencial de acción en una neurona requiere la presencia de canales activados
por voltaje para el sodio(1+). Cuando se abren estos canales la diferencia
de concentración disminuye, así como la diferencia de cargas “despolarización”.
La despolarización crea una corriente localizada lejos del sitio de despolarización.
Estos tipos de canales de sodio(1+) solo están presentes en el axón. La membrana del soma neuronal solo puede trasmitir cambios de potencial de forma pasiva, sin los canales de sodio(1+) no son capaces de generar los potenciales de acción. En la mayoría de los tipos de neuronas la región del axón más cercano al soma es el sitio de inicio del potencial de acción y se denomina segmento de inicio o zona gatillo. Esta zona del axón posee una alta densidad de canales de sodio activados por voltaje.

Figura 7.1. Iniciando el potencial de acción. La
despolarización de la membrana es solo una parte de la historia, de hecho en
realidad ingresa tanto sodio que la membrana invierte su polarización, desde
-70milivoltios que es el potencial de membrana en reposo y llegando casi a los
+30 milivoltios en el pico del potencial de acción. La segunda parte del
potencial de acción lo describiremos en el siguiente numeral y tiene que ver
con la recuperación de la polaridad estándar de la membrana. , (0) Potencial de
membrana; (línea roja) umbral, el espacio en medio es la despolarización por
conductancia pasiva; (1) las cisternas rápidas de los canales de sodio se
abren; (2) sobrecarga, la polaridad interna de la membrana se invierte; (3) pico
de despolarización, se activan las cisternas lentas de los canales de sodio y
estos se cierran; (4) Las bombas de sodio y potasio y los canales de
rectificación de ingreso de potasio restauran la polaridad de la neurona; (5)
Sobrecarga por demasiada efectividad de las bombas de sodio y potasio, las
fuerzas voltaicas restringen la actividad de los canales de potasio hasta
volver a (0) potencial de membrana.
El potencial de
acción no inicia fácilmente, se requiere que cierta cantidad de sodio(1+) halla
cruzado el canal activado por voltaje, o que una gran cantidad de estímulos de
conductividad pasiva desde el soma se sumen en la zona de gatillo, proceso llamado
suma de estímulos. Una vez se alcanza una cantidad límite de despolarización
conocida como umbral de despolarización, los canales de sodio se abren
despolarizando totalmente la sección de la membrana. El gatillo afecta las secciones de la
membrana que se encuentra a su alrededor despolarizándolas, lo cual crea un
efecto dominó, más canales de sodio se abren y estos a su vez afectan a la
sección de la membrana cercana que aún no se ha despolarizado.
7.2 Apertura de los canales de sodio
(YouTube) Los potenciales de acción son creados por la
apertura y cierre cronometrado de los canales del ion sodio (1+) dependientes
de voltaje. Un potencial se acción ocurre debido a los cambios en la
conductancia de una membrana al sodio(1+) y al potasio(1+). Estos canales poseen
dominios que sirven como exclusas y compuertas que permiten acceder o bloquear
el paso de los iones de manera diferencial a través del canal. Los canales del
sodio poseen exclusas de activación y desactivación que son movidos debido a
las fuerzas que ejerce la despolarización en la membrana.
Cuando una
neurona no se encuentra excitada solo los canales para el ion potasio(1+) se
encuentran abiertos, estableciendo el potencial de reposo. Existen muchos tipos
de canales para potasio(1+) en las neuronas y otros tejidos excitables como los
músculos. La forma más simple del canal de sodio(1+) tiene como respuesta
abrirse cuando la membrana está despolarizada y cerrarse cuando se ha repolarizado
hasta recuperar el potencial de membrana en reposo. Los potenciales de acción no se
desencadenan hasta que la membrana ha alcanzado cierto nivel límite de
despolarización o umbral. Un nervio puede ser llevado al umbral de
despolarización por medio de la conducción pasiva solo si esta se emite de
forma constante. Esto le permite al soma transmitir señales a cortas distancias
de una dendrita a otra o bloquear la señal si es el caso.
7.3 Pico de conductancia y sobrecarga neuronal
Cuando la
membrana es despolarizada más allá del umbral, los canales de sodio(1+) se
abren para un transporte pasivo facilitado, permitiendo el flujo de iones
sodio(1+) equilibrando el gradiente de concentración del sodio(1+) e
invirtiendo la polaridad interna de la neurona hasta casi los 30milivoltios.
Esta etapa se conoce como sobrecarga neuronal positiva "overshoot". Después de este pico
empieza la repolarización de la membrana, momento en que las proteínas de
transporte como las bombas de sodio y potasio actúan a su máximo, consumiendo
gran cantidad de energía. Son tan eficientes que hacer que la carga negativa al
interior de la membrana sea superior al del potencial de membrana normal, a
esta etapa se la conoce como sobrecarga neuronal negativa
"undershoot".
La cantidad de iones
que se mueven a través de la membrana durante la sobrecarga es bastante
pequeña, por ejemplo, en cuanto al sodio, solo sale de la célula menos del
0,01%. Lo
anterior implica un mecanismo de control para evitar a perdida extrema de
sodio. Esto se logra gracias a cisternas ubicadas en el canal del sodio.
Existen dos cisternas, una de abertura y otra de cierre que operan a
velocidades diferentes. Ambas se encienden al tiempo durante la
despolarización, pero la de abertura es rápida y permite el escape de sodio por
transporte pasivo, lo cual permite el ingreso de sodio al interior de la célula
invirtiendo la polaridad hasta llegar al pico.
Cuando se alcanza
el pico de despolarización se activan las cisternas de cierre evitando que
ingrese demasiado sodio a la célula. El momento de mayor ingreso de sodio
también se conoce como punto de máxima conductancia o etapa de pico, y una vez
se pasa las cisternas de cierre empiezan a bloquear paulatinamente los canales
de sodio en la membrana. En ese momento la bomba de sodio y potasio comienza a
restaurar la polaridad del potencial de membrana.
7.4 Propagación de un potencial de acción
La velocidad con
la que se propaga una señal neuronal “potencial de acción” depende del diámetro
del axón y del nivel de mielinización. A medida que un axón aumenta su
diámetro el potencial de acción aumenta su velocidad (YouTube)(YouTube)(YouTube).
En los axones sin
mielina los canales de sodio y potasio se encuentran distribuidos de forma
uniforme a lo largo de la membrana axonal. La entrada de iones sodio(1+) en la
región de gatillo causa que las zonas adyacentes se despolaricen, activando
subsecuentemente sus canales iónicos. Esta dispersión continua es semejante a
una onda. Note que no existe un flujo neto de electrones a través de la
membrana, lo que fluye son iones de cargas positivas. Aunque parte de la señal llega al soma, el
potencial de acción se bloquea en este punto debido a la ausencia de los
cabales de sodio. Por tal razón la dirección efectiva del potencial de acción es
hacia la punta de la dendrita.
La conductividad
de la membrana a un potencial de acción también mejora por medio de la
mielinización del axón. Las células gliales, ya sea las células de
Schwann en el sistema nervioso periférico o los oligodendrocitos en el sistema
nervioso central rodean los axones con múltiples capas membranales como si
fueran las hojas de un tamal conocido como capa de mielina. La mielina atenúa
el paso de los iones que se alejan de la membrana axonal, en otras palabras,
asegura la presencia de una alta concentraciones de iones cercanos a la
membrana. Las capas de mielina no son indefinidas, cada cierta distancia hay región
sin mielina en la que la dendrita queda desnuda. Estas regiones son denominadas
como los nódulos de Ranvier.
Por lo general
los axones más largos que 1 mm se encuentran mielinizados y entre más largo la
capa de mielina aumenta. La conductividad de la señal se altera con la
presencia de la mielina. La mielina impide que los iones de sodio(1+) se salgan
ya que aíslan químicamente la membrana, estos se transfieren a lo
largo del axón hasta encontrar un nódulo de Ranvier.
Allí despolarizan la región interna de la membrana iniciando un
potencial de acción.
Allí reinicia el
proceso, ingresa sodio que se dispersa por el axón, al ingresar a una región
con mielina el sodio no puede escapar y se transmite por difusión acusa
impulsada adicionalmente por la dispersión de cargas "iones positivos se
repelen entre sí", los pocos iones que llegan al siguiente nódulo de
Ranvier reinician nuevamente un potencial de acción.
El resultado neto es el de
fuertes pulsaciones voltaicas en los nódulos de Ranvier que amplifican débiles
señales transmitidas a altas velocidades por el nódulo mielinizado.
8. La sinapsis
|| Regresar al Índice ||
Los potenciales de acción permiten comunicar a una dendrita con el soma o viceversa. Los potenciales de membrana permiten comunicar señales entre dendritas a través del espacio del soma de la neurona. Esto le permite a una señal viajar rápidamente por toda la superficie de una célula neuronal individual. El asunto es que una señal debe viajar por más de una neurona y eso implica que estas deben comunicarse entre sí. Las neuronas se comunican en las sinapsis. Existen dos tipos de sinapsis, las sinapsis eléctricas y las sinapsis químicas.

Figura 8.1. Sinapsis eléctrica. El
contacto se logra mediante proteínas que actúan como puentes salinos para que
el ion sodio(1+) se difunda por los canales y sirva para iniciar el potencial
de acción en la otra célula.
En las sinapsis
eléctricas poseen canales de comunicación proteínicos que permiten el paso del
ion sodio(1+), cuando este difunde por el canal a la otra neurona la depolariza
iniciando un nuevo potencial de acción. Este tipo de canales es común en
la etapa embrionaria humana y en algunos tejidos como el muscular, donde dichos
canales penetran la célula muscular para comunicar un potencial de acción al
interior del miocito. En los humanos y en general, la mayoría de los mamíferos
la sinapsis más común es la sinapsis química mediada por mensajeros químicos
con efecto paracrino llamado neurotransmisores.
8.1 La neurona como glándula
En la sinapsis
química no existen puentes que comuniquen físicamente las neuronas, el
potencial de acción originado en una neurona muere efectivamente en la neurona
del cual nació. En consecuencia, la comunicación no se realiza por métodos
voltaicos. Aquí es donde ingresa la función glandular de la neurona. En una
sinapsis química tenemos una dendrita efectora y una dendrita receptora. La
dendrita efectora posee una alta densidad de vesículas de trafico
intermembranario que contienen químicos mensajeros. Aunque clásicamente los
químicos del sistema nervioso se los denomina como neurotransmisores su
similitud con las hormonas es evidente al tratarse de moléculas orgánicas
relativamente pequeñas en comparación con una proteína que generan respuestas
en otros tejidos. En la actualidad se las considera más cercanas ya que muchos
neurotransmisores poseen efectos glandulares de tipo endocrino como la
neurotensina.
Informalmente podemos decir que la mayoría de los neurotransmisores son hormonas paracrinas ya que sus efectos están restringidos a regiones cercanas a las células producidas. Estas distancias son de hecho muy cercanas, y en promedio alcanzan unos 50 nanómetros entre la dendrita efectora y la dendrita receptora. Las vesículas son estimuladas por una despolarización de la membrana, que incrementa la disponibilidad de iones calcio(2+) que activan los rieles de actina sobre los cuales las grúas de dienina o quinesina transportan a las vesículas. Cuando estas grúas acercan lo suficiente las vesículas a la membrana de la dendrita se produce la exocitosis liberando los neurotransmisores al medio de la sinapsis.

Figura 8.2. La sinapsis química. En
este caso una neurona efectora por medio de un axón emite sustancias químicas
llamadas neurtransmisores que entran en contacto con receptores específicos de
una dendrita (YouTube)(YouTube).
8.2 Los Neurotransmisores
Un
neurotransmisor es una molécula química sintetizada por el propio organismo,
acumulado en vesículas y liberado en la sinapsis. La cantidad de
neurotransmisores en una vesícula de exocitosis se denomina cuanto, y el total
de cuantos liberados en una sinapsis se denomina contenido cuántico “no
relacionado con la física cuántica”. Los neurotransmisores viajan por el fluido
de la sinapsis hasta acoplarse con receptores específico. Existen diferentes
tipos de neurotransmisores, cada uno con su receptor específico. Los
receptores son muy variables, pero estos a su vez están acoplados a una
proteína efectora, la cual se activa cuando el receptor se encuentra en su
forma de dímero con el neurotransmisor acoplado. El primero de este tipo de
proteínas efectoras acopladas a receptores son las ionotróficas, no es otra
cosa que un canal de iones activado por neurotransmisores, al unirse al
neurotransmisor la cisterna pasa de cerrada a abierta. Los canales son
diversos, algunos son depolarizadores y otros hiperpolarizadores dependiendo
del ion que ingresa a la célula receptora.
El segundo tipo de receptores se encuentran acoplados a proteínas integrales de membrana del tipo G. Las proteínas G poseen siete dominios transmembranales y un dominio al interior de la membrana, el dominio externo con los siete pliegues es el receptor y el interno es el efector. Cuando una proteína G es activada esta efectúa una transducción de señal al interior de la célula por medio de una cascada metabólica que involucra señalizadores metabólicos de la célula como el AMP cíclico el GMP cíclico y la fosfolipasa C.

Figura 8.3. Receptores acoplados a canales iónicos.
Cuando ciertas neuronas reciben
8.3 Potencial postsináptico
Si el canal activado por una sinapsis ionotrófica es despolarizante, la membrana inicia una señal pasiva denominada potencial postsináptico exitatorio, es semejante en propiedades a la conducción pasiva por medio de un potencial de membrana, viaja a cortas distancias y se debilita a medida que se aleja del punto de activación. Los potenciales postsinápticos por si solos no son capaces de crear un potencial de acción a menos que se sumen varias señales sinápticas. El aumento de estímulos permite llegar al umbral en el que inicial el potencial de acción. Por el contrario, si los canales sinápticos activados son hiperpolarizantes se genera un potencial postsináptico de inhibición que bloque la activación de la neurona, también llamado potencial postsináptico inhibitorio.

Figura 8.4. Tipos de inervación. Las neuronas efectoras receptoras pueden conectarse: (Arriba) de manera axón a dendrita; o (Abajo) axón a soma. Recuerde que un axón y una dendrita son semejantes estructuralmente, y se diferencian solo por la dirección del mensaje, el axón emite y la dendrita recibe.
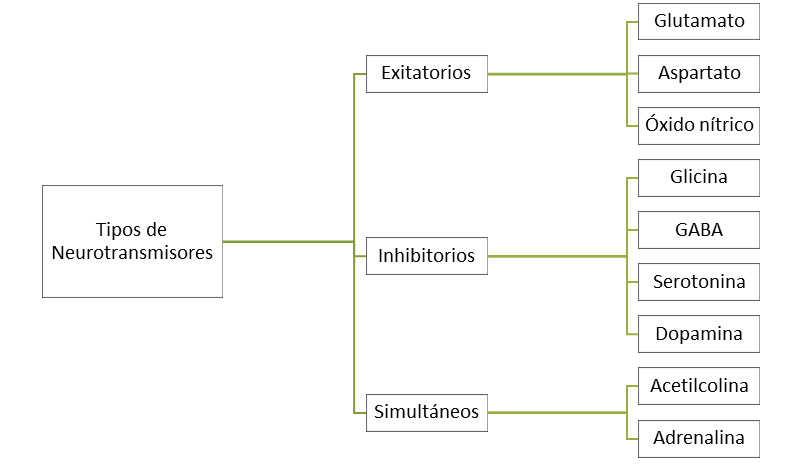
Figura 8.5. Tipos de neurotrasmisores Los
neurotransmisores pueden ser estimulantes/exitatorios de potenciales de acción
o inhibidores; unos pocos pueden tener ambos efectos dependiendo del receptor
al que se acoplan.
Los potenciales
postsinápticos de acción pueden sumarse a lo largo de una dendrita si en su
ruta hay varias sinapsis activadas de forma simultánea o casi simultánea
iniciando así el potencial de acción. Sin embargo, esto depende mucho de la
cercanía a la región del gatillo neuronal. Recordemos que los gatillos
neuronales se encuentran en la coyuntura entre el soma y la dendrita, por lo
que las sinapsis cercanas al soma o que se encuentran en el soma
“sinapsis axosomática” puede activar un potencial de acción con mayor
facilidad que las sinapsis ubicadas de forma muy lejana al soma en una dendrita
“sinapsis axodendrítica” (Figura 8.4).
8.4 Diversidad de neurotransmisores
Los
neurotransmisores tienden a ser moléculas de un peso molecular bajo cuando se
las compara con una proteína completa. Por lo general son compuestos orgánicos
de tipo aromático o con anillos orgánicos, por lo que su estructura puede ser
enmascarada por sustancias imitadoras. Históricamente los primeros
neurotransmisores en ser descritos fueron la acetilcolina y la norepinefrina.
Estos fueron aislados en las sinapsis de los sistemas nerviosos periféricos.
Muchos otros
neurotransmisores han sido identificados desde entonces, sin embargo, aun
contando los péptidos de neurotransmisión, estas cuentan unas 50 moléculas. Lo
que proporciona la especificidad en la señalización es el modo en que las
neuronas se encuentran conectadas unas con otras, y la distribución de los
receptores de los diferentes tipos de neurotransmisores.
El problema surge
en que los receptores de los neurotransmisores no son tan específicos como
cabría de esperarse de un receptor proteínico. En muchas ocasiones existen
reacciones cruzadas con moléculas del ambiente que imitan los
neurotransmisores. Estas sustancias en consecuencia son capaces de efectuar
estímulos neuronales muy potentes. Su estudio es importante en el caso de las
adicciones debido a la formación de dependencia por destrucción de los
receptores. Los detalles concretos de esto lo veremos en artículos futuros.
9. Tipos entre los neurotransmisores
|| Regresar al Índice ||
Los
neurotransmisores se han clasificado tradicionalmente en tres categorías
principales: aminoácidos, monoaminas y polipéptidos. Ejemplos de aminoácidos
que actúan como neurotransmisores son el glutamato, la asparangina, la glicina
y el pacido aminobutírico G. Las monoaminas se definan como tal debido que
se sintetizan a partir de un único aminoácido precursor. Ejemplos de esta clase
incluye la acetilcolina derivada de la colina y los transmisores de la
cetocolamina como la dopamina, la norepinefrina y epinefrina, derivado del
aminoácodo tirosina y la indolamina, serotonina o la 5-hidroxitriptamina
derivadas del triptófano.
Los neurotransmisores
con base en polipeptidos llamados neuropeptidos consisten en una cadena de
aminoácidos que van desde tres hasta varias docenas de aminoácidos. Ejemplos de
los polipeptidos que actúan como neurotransmisores son los opiodes. También se los puede
clasificar de acuerdo a su función, de allí los denominamos exitatorios,
inhibitorios o simultáneos. El efecto de un neurotransmisor no depende de la
molécula en si, sino del receptor y la ruta de tramisión de información al
interior de la célula.
Algunos como el
glutamato o el aspartato se unen a receptores que inician potenciales de accíon
excitando las neuronas. Otros se unen a receptores que bloquean los gatillos
impidiendo que las neuronas se activen aun cuando son estimuladas, como en el
caso del GABA o la dopamina, esto es típico de las sensaciones de anestesia.
Los simultáneos se unen a receptores que pueden ser uno o lo otro dependiendo
del tejido neuronal que sirve de blanco.
9.1 Acetilcolina

Figura 9.1. Acetilcolina (YouTube) etanoato de 2-(N,N,N-trimetil)-etanoamonio
Las neuronas que usan la acetilcolina como
su neurotransmisor se denominan colinérgicos. La acetilcolina se sintetiza a
partir de colina y acetato bajo la enzima colina acetiltransferasa. La
acetilcolina se sintetiza y se almacena en las vesículas en la parte final del
axón. La acetilcolina se sintetiza en el soma donde se encuentra los organelos
de síntesis como el aparato de Golgi y luego se transporta por medio de
vesículas a la punta de la dendrita.
El paso limitante
para la síntesis de acetilcolina es la disponibilidad de loa molécula de
colina, la cual debe ser reciclada constantemente desde el exterior de la
neurona. Los receptores de la acetilcolina se conocen como receptores
colinérgicos, y se puede dividir en dos categorías basadas en las drogas que
son capaces de imitar o antagoniza las acciones de la acetilcolina en las
células blanco.
En los estudios clásicos
durante el siglo XX, las drogas de muscarina aislada de los hongos venenosos y
la nicotina aislada del tabaco fueron empleados para distinguir los dos tipos
de receptores. La muscarina estimulaba algunos receptores, mientras que la
nicotina estimula los demás, de allí emerge la clasificación de receptores
muscarinicos y receptores nicotínicos.
9.2 Receptores de acetilcolina
9.3 Receptores
de nicotina
Los receptores
nicotínicos están compuestos por cinco subunidades, dos alfas, una beta, una
gamma y una delta, los cuales están vinculados directamente a canales iónicos
que afectan directamente al potencial postsináptico de acción. El canal iónico
se encuentra en el centro de la macromolécula. Existen múltiples formas
de las subunidades lo cual da lugar a una familia de receptores nicotínicos. Las
dos subunidades se emplean para acoplarse a la acetilcolina. Cuando la
acetilcolina se acopla a las dos subunidades ocurre un típico cambio
conformacional abriendo el canal iónico iniciando el potencial de acción.
9.4 Los
receptores de muscarina
Los receptores de muscarina poseen siete dominios transmembranales típicos de todas las proteínas G acopladas a receptores extramembranales. Se han reportado hasta el momento cinco tipos de receptores de muscarina. La señal transmitida por los receptores de muscarina tiene múltiples rutas de acción, pudiendo activar cascadas de reacciones hacia (1) la despolarización de la membrana iniciando un potencial de acción o (2) iniciando una transducción de información por medio del metabolismo celular hacia el núcleo.

Figura 9.2.
Funcionamiento de la acetil-colina. Los músculos en la placa neuromuscular poseen
receptores nicotínicos para la acetilcolina (A); Los receptores de muscarina
son proteínas G transmembranarias de siete dominios "el modelo no los
muestra" acopladas a receptores. El efecto metabólico tiende a bloquear
los potenciales postsináticos de acción impidiendo la depolarzación
"izquierda" o hiperpolarzando la membrana extrayendo potasio(1+) del
interior de la célula haciéndola aún más negativa (B).
9.5 Desactivación de la acetilcolina
La acetilcolina
es un material demasiado potente y escaso al interior de las neuronas, por lo
que su liberación debe estar altamente regulada para evitar (1) la sobreactivación
de más neuronas “o músculos” de lo estrictamente necesario y (2) para reciclar
la colina, molécula que sirve como mecanismo de control de la acción neuronal.
Los receptores de acetilcolina responden a la molécula completa, por lo que
retirar el grupo acetilo inactiva su acción como neurotransmisor, para lograrlo
las neuronas segregan rápidamente la enzima acetilcolinesterasa que hidroliza
la acetilcolina.
9.6 Reciclaje de la acetilcolina
La colina
liberada al medio intermembranario es luego capturada por receptores de
membrana y endocitada en vesículas para ser transportada nuevamente al soma de
la neurona para fabricar nuevamente acetilcolina. Otro tejido con receptores
para acetilcolina es el tejido muscular, la estimulación con acetilcolina es quien
da inicio a la cascada de eventos que permite el movimiento del sarcómero.
9.7 Catecolaminas
Las catecolaminas
reciben su nombre debido a que consisten en un grupo catecol. El grupo
catecol es un nombre tradicional para el benceno-1,2-diol. El grupo catecol
está unido a una cadena lateral de dos o tres carbonos que puede ser o un ácido
carboxílico o una amina. Las catecolaminas no solo están relacionadas
estructuralmente, también biosintéticamente ya que una de produce de otra
mediante una modificación enzimática. Las principales catecolaminas en su orden
de biosíntesis son las siguientes: L-tirosina, L-dihidroxifenilalanina,
hidroxifeniletilanina, L-nerepinefrina y L-epinefrina.
Dentro del grupo anterior cabe destacar el origen de la expresión “dopar”. La L-dihidroxifenilalanina es un nombre muy pesado para usarlo de forma común, por lo que se emplea generalmente un acrónimo de sus principales prefijos en inglés: DihydrOxyPhenylAlanina; o L-DOPA. Por otra parte, la molécula sintetizada a partir de L-DOPA se caracteriza por la adición de un grupo amino en la cadena lateral, por lo que el nombre acrónimo se modifica con la terminación amina, quedando L-dopamina.
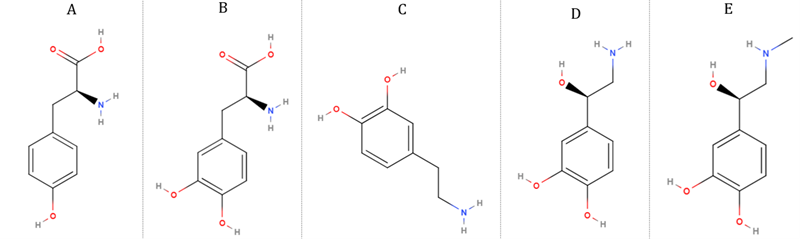
Figura 9.3.
Las catecolaminas. Las catecolaminas son neurotransmisores
famosos: tirosina (A); DOPA (B); dopamina (C), Noradrenalina (D); y adrenalina
(E). Tienen efectos profundos en el sistema nervioso y endocrino, por lo que se
las denomina neuro-hormonas (YouTube)(YouTube)(YouTube)(YouTube)(YouTube).
9.8 Receptores catecolinergicos
Tres de las
enzimas involucradas: hidroxilasa de tirosina, betahidroxilasa de dopamina y
N-metiltransferasa de feniletanolamina; son expresadas de forma exclusiva por
las células secretoras de cetocolaminas. Las neuronas dopaminicas expresan
únicamente hidroxilasa de tirosina, mientras que las neuronasnoradrenergicas
segregan hidroxilasa de tirosina y betahidroxilasa de dopamina. Las células
adrenérgicas expresan las tres enzimas.
Las células
adrenérgicas incluyen pequeñas poblaciones de neuronas del Sistema Nervioso Central,
así como células de chromafin en la médula adrenal, la cual segrega epinefrina
durante la activación generalizada del componente simpático del sistema nervios
autónomo durante la respuesta de combate o huida. Los receptores de las
catecolaminas son típicas proteínas G con siete dominios transmembranales con
receptores especializados para las catecolaminas. Dichos receptores pueden
categorizarse en varios subtipos englobados en dos categorías principales. Los
receptores D1 y D5 que estimulan la producción de adenilil ciclasa y los
receptores D2, D3 y D4 que bloquean la producción de denilil ciclasa. Los
receptores de adrenérgicos se encuentran estimulados por la norepinefrina y la
epinefrina y se localizan en células a través de todo el cuerpo, incluyendo el
Sistema Nervioso central y órganos periféricos del sistema nervioso simpático.
9.9 Receptores adrenérgicos
Los receptores
adrenérgicos de denominan así debido a que la epinefrina
y norepinefrina también son conocidos por nombres aún más famosos, la
adrenalina y noradrenalina respectivamente. Los receptores se clasifican como
alfa o beta dependiendo de la potencia de la catecolamina y sus análogos en
estimular cada tipo de receptor. Los análogos que se emplearon originalmente
para distinguir los tipos de receptores fueron la norepinefrina, la epinefrina como
sustancias naturales, el isoproterenol y la fenilefrina.
En 1948 se
identificó que los receptores alfa tenían una afinidad decreciente en la que la
epinefrina tenía la mayor afinidad seguida de la norepinefrina, mientras que el
isoprotenerol tenía una afinidad radicalmente inferior. Los receptores beta
exhibían una afinidad completamente diferente, respondiendo con mayor afinidad
es el isoprotenerol, mientras que la epinefrina tenía un comportamiento
semejante al de la norepinefrina. Los estudios con la fenilefrina también
distinguieron los dos tipos de receptores, los tipo alfa respondían a esta
molécula, mientras que los tipo beta no lo hacían.
Los receptores
tipo alfa fueron posteriormente divididos en los alfa uno y los alfa dos. Los
receptores alfa uno incrementan la liberación del ion calcio(2+), lo cual a su
vez activa las proteínas dependientes de la quinesina y la actina en el
citoesqueleto, lo cual acelera el desplazamiento de vesículas y organelos, así
como la secreción de la proteína quinasa C. La activación de los receptores
alfa dos inhiben la adenilil ciclasa y reduce los niveles de AMP cíclico en la
célula, lo cual tiende a inactivarla, el AMP cíclico es un señalizador
energético, altos niveles de este empujan a la célula a adquirir nutrientes
encendiendo varias rutas metabólicas. Los receptores tipo beta inhiben la
actividad de la adenilil ciclasa aumentando los niveles de AMP cíclico.
9.10 Inactivación y reciclado de las
catecolaminas
Después de su
liberación en la sinapsis, receptores de transporte en la dendrita efectora
remueve las catecolaminas del medio y las transfiere al interior por
endocitosis. Los receptores de membrana son dependientes de concentración, al
disminuir la cantidad de catecolaminas en el medio, los receptores de catecolaminas
pierden afinidad y las liberan al ambiente sináptico, donde la dendrita
efectora continua su labor de reciclaje.
La endocitosis de
las catecolaminas culmina en su empaquetado en vesículas de transporte a las
terminales vesiculares presinápticas donde son reempacadas en vesículas de
transporte hacia la membrana. Las catecolaminas pueden ser inactivadas en caso
de haberse producido demasiadas por dos enzimas, la oxidasa de omonaminas y la
cetocol-o-metiltransferasa. Una es producida en el interior de las neuronas y/o
sus mitocondrias, mientras que la segunda es segregada por los astrocitos.
9.11 Serotonina

Figura 9.4. Serotonina. 3-(2-aminoetil)indol-5-ol. La serotonina es uno de los neurotransmisores
del placer.
La serotonina es
formalmente conocida como 5-hidroxitriptamina es el mensajero de las neuronas
serotoninérgicas. La enzima responsable para la producción de la
serotonina es la hidroxilasa de triptófano, la cual convierte el aminoácido
triptófano obtenido de la dieta en el intermediario 5-hidroxitriptofano, el
cual es convertido en serotonina por descarboxilación. La serotonina es
almacenada en vesículas de transporte neuronal y segregadas por exocitosis por
el mecanismo típico de liberación de ion calcio(2+). Se han descrito dos tipos de receptores
para la serotonina. En la primera categoría se localizan las serotoninas
de tipo uno, tipo dos y tipo cuatro, estos receptores son proteínas G unidas al
receptor de serotonina. S efecto intracelular es la inhibición de la adenilil
ciclasa. Este tipo de receptores se encuentran en el Sistema Nervioso Central y los vasos sanguíneos craneales.
Los receptores de
serotonina tipo dos estimulan la producción de fosfolipasa C la cual a su
vez induce la producción de pirofosfato “1, 4, 5-trifosfato” y diacilglicerol.
Los receptores de este tipo se encuentran en el Sistema Nervioso Central, el
musculo liso, las estructuras vesiculares, el tracto gastrointestinal y en las
plaquetas. Los receptores tipo cuatro estimulan la producción de adenilil
ciclasa y se encuentran en el Sistema Nervioso
Central y el tracto gastrointestinal. La segunda gran categoría de receptores de
serotonina agrupa a los receptores del tipo tres. Estos no son proteínas G,
sino canales iónicos activados por receptores de serotonina, los cuales
permiten la entrada de sodio y la formación de un potencial de acción
postsináptico. Las neuronas con receptores de dopamina tipo tres inervan la corteza
cerebral asociada con las sensaciones de nausea. Al igual que las
catecolaminas, la serotonina es capturada por receptores y endocitada en
vesículas.
9.12 Glutamato

Figura 9.5. Ácido glutámico/glutamato. Acido
2-aminopentanodioico. También llamado glutamao, es relevante en la adicción
al alcohol y los inhalantes depresores (YouTube).
El glutamato es
el mensajero principal para el estímulo de las neuronas del Sistema Nervioso Central. Las neuronas con receptores de glutamato
son la población predominante en la transmisión de información sensorial
en el Sistema Nervioso
Central. El glutamato es un aminoácido que sirve como materia prima para varias
rutas metabólicas en el metabolismo basal de las células, pero las neuronas
también lo han especializado en la neurotransmisión.
La fuente
principal de glutamato neuronal se encuentra en la mitocondria a través de la
conversión del cetoglutarato derivado del ciclo de Krebs al cual se adiciona un
grupo amino para formar el glutamato. Como todos los neurotransmisores, el
glutamato empleado para la neurotransmisión se almacena en vesículas que liberan
su contenido por medio de exocitosis.
9.13 Receptores iónicos de glutamato
Después de su liberación, el glutamato interactúa con receptores específicos en la neurona receptora. Los receptores de glutamato se denominan receptores GLU y pueden ser del tipo canal iónico ligado a receptor o una proteína G ligada a un receptor de siete dominios transmembranales. Los receptores ionotróficos se clasifican en base a análogos que pueden realizar reacciones cruzadas que las activan: kainato, ácido α-amino-3-hidróxi-5-metil-4-isoxazolpriónico (AMPA) y N-metil-d-aspartato (NMDA).

Figura 9.6. Reciclaje de glutamato. La
mayoría de los modelos representan al receptor del astrocito como el principal
mecanismo de reciclaje del glutamato en la sinapsis.
Los receptores de
kainato y AMPA producen potenciales de acción postsinápticos mediante la
apertura de canales específicos para el ingreso del ion sodio(1+) y en menor
medida para el calcio(2+), lo cual despolariza la región interna de la membrana
neuronal. La activación del receptor NMDA es único debido a que debe ser activado por
dos neurotransmisores derivados de aminoácidos o sus homólogos cruzados, en
este caso se trata del glutamato y la glicina. Los canales iónicos del NMDA
permiten el ingreso de los iones sodio(1+) y calcio(2+). El canal iónico tiende
a ser afín también al ion magnesio(2+) pero este ion lo bloquea justo al inicio
de la primera fase de despolarización.
A medida que la
membrana se hace más despolarizada, el bloqueo con los iones magnesio(2+) se
distiende, probablemente debido a que el interior de la membrana se hace más
positivo, y en consecuencia las cargas empiezan a repeler los iones positivos
de magnesio(2+). La transmisión de glutamato a través de los receptores NMDA es importante
en el desarrollo de conexiones neurales especiales relacionadas con el
aprendizaje y la memoria.
9.14 Receptores de glutamato acoplados a
proteínas G
Las proteínas G
que se unen al glutamato se conocen como receptores metabotrópicos de
glutamato, es un nombre bastante largo, así que las llamaremos simplemente
proteínas G para el glutamato. Como ya es bastante repetitivo, los receptores de
glutamato de tipo proteína G dependen de un receptor de membrana con siete
dominios transmembranales, en los cuales la superficie extramembranaria se
genera una estructura afín al neurotransmisor, en este caso el glutamato.
Cuando el
receptor forma su binomio con el glutamato hay un cambio conformacional que
inicia la reacción de la proteína G al interior de la membrana, la cual genera
una cascada de reacciones metabólicas, las cuales en este caso redirigen de
forma indirecta a la activación de canales iónicos. La activación de todos
estos tipos de receptores inhiben los canales de calcio(2+). La función de
otros canales iónicos por el contario puede estimular su apertura.
9.15 Reciclaje del neurotransmisor glutamato
Al igual que los
demás neurotransmisores, estos duran muy poco tiempo liberados en el pasaje
sináptico, lo cual implica que el tiempo en que los receptores de la neurona
preceptora solo se estimulan por un periodo de tiempo muy breve. En el caso del
glutamato existen dos mecanismos de transporte activo muy eficientes. El
control del glutamato es muy importante debido a que posee un efecto
neurotóxico debido a la inducción del ingreso de iones calcio(2+). La
razón para este es bastante simple, cuando la cantidad de iones calcio(2+)
aumenta en el citoplasma, no solo se activan todos los rieles del
citoesqueleto, también se activan las rutas metabólicas del suicido
celular o apoptosis. A parte de esto que ya es muy malo, existen otros efectos secundarios que
impiden el ingreso de oxígeno a la neurona.
Los dos
mecanismos dependen del linaje neuronal, uno se encuentra en la neurona
efectora y otro en los astrocitos, células gliales encargadas de la protección
inmune del sistema nervioso. En la neurona el glutamato regresa al aparato de Golgi,
mientras que en el astrocito el glutamato se convierte en glutamina que es
enviada a la neurona para reconvertirla en glutamato, almacenado en vesículas
en el aparato de Golgi y de allí almacenado en vesículas para la exocitosis.
9.16 Ácido ϒ-aminobutírico GABA

Figura 9.7. GABA. Acido
4-aminobutirico. (YouTube)
Otro aminoácido
empleado como neurotransmisor él es ácido gama-aminobutírico o GABA por sus
siglas en inglés. El GABA es un neurotransmisor con una función
predominantemente inhibitoria. Las neuronas GABAnérgicas se encuentran
distribuidas por el Sistema Nervioso Central. La síntesis de GABA en la neurona
posee características semejantes al glutamato, ya que se encuentra vinculado al
metabolismo de los ácidos tricarboxílicos o ciclo de Krebs. De hecho, se
produce mediante la descarboxilación del glutamato mediante la enzima glutamato
descarboxilasa, un marcador genético que permite identificar las neuronas
GABAnérgicos y mediante la transaminación del grupo amino.
Como todos los
neurotransmisores el GABA se almacena en vesículas para la exocitosis. Existen dos tipos de
receptores de GABA llamadas GABAA y GABAB. Los
receptores GABAA son canales iónicos ligados a receptores de
GABA. Estos canales iónicos permiten la entrada de iones cloro(1-) en la
neurona hiperpolarizándola generando un potencial postsináptico de inhibición,
noqueando la neurona para posibles estímulos. Los receptores GABAB son
proteínas G acopladas a receptores transmembranarios. El estímulo de estos
segundos receptores aumenta el ingreso de iones potasio(1+) al interior de la
célula o de forma alternativa inhibiendo los canales de calcio(2+) también al
interior de la célula, ambos pasos impidiendo que se genere la despolarización.
GABA se recicla
igual que el glutamato por receptores de alta afinidad en la membrana de la
neurona efectora o en los astrocitos. En el interior de la célula, las
vesículas con GABA endocitado es enviado a la mitocondria donde es convertido
mediante una modificación del ciclo de Krebs en glutamato.
Los receptores GABAA aumentan
su efecto mediante el uso de las benzodiacepinas, las cuales son empleadas para
tratar la ansiedad y en algunos anestésicos generales. Las drogas que inhiben
el GABA pueden causar eventos epilépticos, indicando que el rol inhibitorio es
una función primordial para el control del sistema nervioso central.
9.17 Glicina

Figura 9.8. Glicina. Acido
2-aminoetanoico.
La glicina es
otro aminoácido con funciones de neurotransmisor, y que al igual que el GABA se
caracteriza por una función inhibitoria. Las neuronas que responden a este
neurotransmisor se denominan glicinérgicas y se encuentran principalmente en la
medula espinal y el tallo cerebral. La glicina se produce a partir de la serina
mediante la enzima hidroximetiltransferasa de serina. La glicina se almacena en
vesículas para exocitosis y liberada cuando se presenta una despolarización de
la membrana terminal del axón.

Figura 9.9. Sinapsis. En
la imagen se observan los receptores A del GABA y los receptores de glicina, su
efecto neto es el mismo, ser un canal para el ion cloruro(1-).
Los receptores de
la glicina son semejantes a los GABAA, se trata de canales iónicos
que permiten la entrada de iones cloro(1-) que hiperpolarizan la neurona y la
bloquean para estímulos de otros neurotransmisores. Su efecto termina mediante
receptores de alta afinidad en la membrana de la neurona efectora.
9.18 Neuropéptidos
Los péptidos
pueden funcionar como neurotransmisores. Muchas de estas sustanciad fueron
descubiertas originalmente en la pituitaria y el sistema gastrointestinal, pero
posteriormente fueron aislados en las neuronas.
Los péptidos con
funciones de neurotransmisión se generan a partir de precursores mucho más
largos, los cuales son empaquetados en el aparato de Golgi en vesículas de
transporte. Al interior de la vesícula en movimiento, algunas proteasas cortan
los precursores en sus formas definitivas. Dichas vesículas generalmente se
almacenan junto a neurotransmisores más clásicos y son exocitadas cuando el
potencial de acción llega a límites muy altos.
Al ser moléculas
compuestas por varios aminoácidos “que van de cinco hasta más de veinte” existe
la posibilidad de una variabilidad considerable. Lo anterior implica que existe
la posibilidad de existencia de neurotransmisores aún no reportados, de hecho,
en la actualidad existe una lista de péptidos de los cuales se sospecha tienen
una probable función de neurotransmisión. Por lo general los receptores de los
péptidos son proteínas G acopladas a un receptor con siete dominios
transmembranarios. El reciclaje de los péptidos depende del clivaje enzimático
en el espacio sináptico.
9.19 Óxido Nítrico
El ácido nítrico
presenta algunas diferencias con otros neurotransmisores. En primera instancia
se trata de un gas difusible de forma pasiva a través de las membranas
biológicas, por lo que una vez producido en una neurona, este se difunde a
través de la sinapsis a las neuronas y células gliales cercanas generando reacciones
metabólicas de tipo secundario. A estas rutas metabólicas complejas se las
conoce como mensajeros secundarios. La actividad del óxido nítrico se encuentra
altamente regulada por los iones de calcio(2+), por lo que su liberación
depende del ingreso a la célula de dichos iones.
El óxido nítrico (NO) al ser producido por
su enzima sintetizadora (NOS) se difunde a través de la membrana sin necesidad
de proteínas de transporte activo o facilitado. Básicamente se trata de un neurotransmisor que no emplea la exocitosis para
salir de la neurona efectora ni de receptores de membrana para introducir su
señal en la neurona receptora. El óxido nítrico fue originalmente identificado como un factor relajante
del endotelio de los vasos sanguíneos, posteriormente también fue señalada su
función como un neurotranmisor, especialmente en lo referente al sistema
nervioso relacionado con el sistema reproductor durante la erección.
9.20 Neurolípidos
La
principal molécula con función de neurotransmisión de naturaleza lipídica
es el ácido araquidónico, que en últimas es un ácido carboxílico. Este es un
ácido graso que se libera desde los fosfolípidos de membrana mediante la
activación de la fosfolipasa A2. A diferencia de otros neurotransmisores, la
liberación del ácido araquidónico no depende de un potencial de acción, por el
contrario depende de la señal efectuada desde una proteína G. Al ser de
naturaleza lipídica el ácido araquidónico puede difundirse a las células
cercanas y a la misma neurona efectora activando segundos mensajeros
metabólicos de forma semejante al óxido nítrico.
Los
endocanabinoides son otras sustancias de tipo lipídico que exhiben propiedades
de señalización sináptica. Tampoco requieren de transporte vesicular peri si de
receptores de tipo proteína G en la membrana. Debido a un vincula con drogas de
abuso como la marihuana la investigación de los endocanabinoides aún se
encuentra en una etapa muy temprana, lo cual es paradójico tomando en cuenta
que el receptor para canabinoides del tipo 1 es la proteína G ligada a
receptor más abundante en el cerebro humano, regulando una abundante serie de
respuestas fisiológicas. Lo que sí es claro es que su síntesis aumenta cuando
la cantidad de calcio(2+) en la célula aumenta, por lo que pueden estar
involucradas en procesos como el suicidio celular o apoptosis.
10. Sistema nervioso de los cnidarios y gusanos
|| Regresar al Índice ||
Todas las células vivas responden a los estímulos y transmiten señales químicas desde el exterior hasta el interior de sus células mediante receptores específicos de membrana. Incluso los organismos unicelulares que viven en colonias son capaces de transmitir una señal a sus parientes, aunque sea por cortas distancias. Esto se debe en parte a la función del sistema endocrino, sin embargo, a medida que la función del movimiento se hace cada vez más importante, y los cuerpos de los animales se hacen más grandes, surge la necesidad de un sistema especializado en integrar la información y emitir respuestas a grandes velocidades, este es el sistema nervioso, derivado del sistema endocrino.
Figura 10.1. Sistemas nerviosos de los vertebrados.
Existen varios tipos del sistema nervioso, pero exclusivos de los animales, en
todos los casos el tejido funciona igual que en el ser humano, mediante nervios
que transmiten información mediante potenciales de acción y neurotransmisores (YouTube).
La unidad
funcional y estructural del sistema nervioso en los invertebrados, al igual que
en los vertebrados como el ser humano es la neurona. Aunque los vertebrados son
famosos por poseer un ganglio anterior altamente centralizado y protuberante al
que llamamos cerebro, todos los animales se caracterizan por poseer diferentes
regiones de integración nerviosa o ganglios menores. En algunos invertebrados
la centralización del sistema nervioso en único gran ganglio cerebral parece no
ser una norma necesaria, por lo que es común encontrarse animales con cerebros
descentralizados, y en consecuencia, que son fáciles de regenerar. Esto contrasta con
los vertebrados en los que un daño en el cerebro es usualmente algo
catastrófico. Cabe anotar que los vertebrados aun poseemos algún grado de
descentralización del cerebro en ganglios menores ubicados en paralelo a la
médula espinal, los cuales se enraízan a un grupo de nervios unificando la
información en ambos sentidos.
Lo anterior nos
lleva a una distinción común, todos los cerebros son ganglios, pero no todos
los ganglios son cerebros. Existe una amplia diversidad en los ganglios que
presentan los invertebrados, y aun en los casos en los que se presenta un gran
ganglio anterior no se lo denomina necesariamente como cerebro. Algunas
anatomías comparadas lo denominan ganglio cerebral. Los vertebrados como el ser
humano también poseen ganglios, regiones donde varios nervios se trenzan y
generalmente donde descansan los cuerpos de las neuronas del sistema nervioso
periférico.
Lo que cambia es
la inervación, es decir el modo en que los nervios forman redes, así como la
complejidad de las trenzas nerviosas o ganglios. En las Cnidarios (Figura 10.1 a) hay una red no centralizada, en los
equinodermos (Figura 10.1 b) hay una red organizada pero también
descentralizada, algunos filos presentan el sistema nervioso en escalera, con
al menos dos hileras longitudinales unidas por nervios perpendiculares (Figura 10.1 c, f, g) mientras que otros linajes han
desarrollado la centralización, con un solo nervio longitudinal principal, del
cual emergen bifurcaciones periféricas (Figura 10.1 d, e, h). Comenzaremos
omitiendo a los poríferos, ya que las esponjas no poseen un sistema nervioso,
su coordinación celular obedece a hormonas, es decir al su sistema endocrino.
10.1 El sistema nervioso en los cnidarios
Consistentemente con su plan corporal radial, los cnidarios poseen un sistema nervioso difuso y descentralizado. Usualmente las neuronas se organizan en dos redes reticulares denominadas redes nerviosas, una entre la epidermis y el mesénquima y otra entre la gastrodermis y el mesénquima. Generalmente la red nerviosa del gastrodermo se encuentra menos desarrollada y en algunas especies puede haber desaparecido completamente.

Figura 10.2. El sistema nervioso de los cnidarios. En los modelos anteriores podemos apreciar el sistema nervioso de algunos cnidarios, los pólipos "izquierda" presentan las redes más complejas, mientras que las hidras "derecha" presentan las redes más simples.

Figura 10.3. Sistema nervioso en platelmintos de
vida libre. Algunos turbellarios no presentan un sistema
nervioso centralizado, en su lugar se asemeja más a una red "izquierda en
el modelo anterior", pero a medida que el animal crece aparecen cordones
longitudinales centralizados "derecha en el modelo anterior" que
terminan con el desarrollo del sistema nervioso en escalera.
Las células
nerviosas generalmente no se encuentran polarizadas, permitiendo un flujo de la
información de modo bidireccional, aunque existen algunas pocas sinapsis
polarizadas. De hecho, no existe una polarización lineal tampoco, lo cual
implica que una señal generada en un punto, se dispersa radialmente a todo el
cuerpo, en lugar de dirigirse a una región central. En otras palabras, el
sistema nervioso de los cnidarios no posee distinción entre un sistema nervioso
central y un sistema nervioso periférico.
10.2 El sistema nervioso en los platelmintos
👉 El
sistema nervioso en los turbellarios: Los gusanos turbellarios poseen sistemas nerviosos que varían desde ser una
mera red con ganglios muy pequeños cerca de la cabeza a tener un sistema
bilateral, en donde los nervios principales se proyectan paralelamente a cada
lado con un ganglio cerebral bien organizado en la cabeza, como si se tratara
de una escalera.
Entre más grande
es hace el gusano la cantidad de cordones nerviosos centrales disminuye, lo
cual centraliza los impulsos nerviosos con mayor eficacia. Adicionalmente
se observa una mayor cefalización, con un ganglio cerebral más grande, órganos
sensoriales agrupados cerca del gran ganglio. Por otra parte, los nervios
parecen especializarse en rutas motoras y rutas sensoriales, lo que plantea una
mayor organización.
👉 Tremátodos
y cestodos: Los nervios principales de los gusanos en
cinta se distinguen en tres grupos de parejas paralelas, la pareja lateral, la
pareja ventral y la pareja dorsal con un ganglio cerebral cerca de la ventosa
bucal. Esta estructura se mantiene en las formas con segmentos repetitivos,
incluso los ganglios se repiten en cada segmento.
10.3 El sistema nervioso en los nematodos
Con algunas variaciones la estructura del sistema nervioso de los nematodos es similar a lo largo de todo el clado. Existe un ganglio cerebral compuesto por un anillo nervioso y varios ganglios menores asociados que contienen generalmente los cuerpos de las neuronas. Las ramificaciones de los nervios motores se extienden de forma anterior desde el anillo nervioso conectándose con los órganos de los sentidos. Mediante una serie de ganglios asociados, los nervios longitudinales se extienden posteriormente a través de médulas epidérmicas. El mayor de estos nervios troncales es ventral, conteniendo fibras motoras y sensoriales. Está formado por la unión de tractos nerviosos pareados que emergen ventralmente en el anillo nervioso y se fusionan posteriormente, donde el tronco principal porta los ganglios.

Figura 10.4. Ganglios cerebrales de los nematodos. Si
hay una palabra con la que podemos describir al sistema nervioso de los
nemátodos es "el cilindro", se trata de una red compleja de nervios
longitudinales unidos por nervios perpendiculares que forman anillos. Los
ganglios son algo común a lo largo del cuerpo, aunque hay un proceso se
cefalización importante.
A parte del que
se origina ventralmente hay otro cordón dorsal que es motor, y otros dos
cordones laterales que son predominantemente sensoriales. Existen comisuras
laterales que unen todos los tractos nerviosos principales en una red unificada
previa a los ganglios cerebrales.
10.4 El sistema nervioso en los anélidos
El sistema
nervioso de los anélidos aparece como una simplificación del sistema nervioso
de otros gusanos, con una menor cantidad de nervios longitudinales principales,
una mayor cefalización, así como un énfasis en ganglios en cada uno de los
segmentos del animal.
En los poliquetos
se extienden dos cordones nerviosos principales que terminan en la región
anterior principal en una pareja de ganglios conectados por comisuras
nerviosas, lo cual puede ser interpretado como un cerebro bilobular. Algunos
poliquetos pueden presentar cuatro ganglios en lugar de dos, mientras que, en
otros, los dos cordones ventrales han tendido a juntarse hasta formar una única
medula o cordón nervioso.
Una diferencia entre los oligoquetos y los poliquetos es que en los primeros el cerebro se encuentra lejos de la punta anterior, tan lejos como el tercer segmento, mientras que en los poliquetos el ganglio cerebral se ubica generalmente en el primer segmento. Del mismo modo en que en los nematodos, el ganglio cefálico extiende nervios de forma anterior que se conectan con los órganos de los sentidos. El ganglio cerebral de los poliquetos posee una estructura especializada en tres regiones denominadas: cerebro superior, cerebro medio y cerebro inferior.

Figura 10.5. Sistema nervioso de un oligoqueto. Cada
segmento posee ganglios propios que regulan el movimiento del segmento y los
quetos, mientras que el ganglio cerebral controla los movimientos mediante
efectos inhibitorios.
El cerebro
superior se encuentra conectado a los palpos bucales, el cerebro medio regula
los ojos, las antenas y los tentáculos y el cerebro inferior los órganos quimiosensoriales.
El cerebro medio genera nervios que controlan el sistema gastrointestinal, la
boca y los órganos asociados a esta como la probóscide. La unificación de la
información se da en un ganglio subentérico que se conecta a los cordones
nerviosos longitudinales.
11. Sistema nervioso de los moluscos
|| Regresar al Índice ||
El sistema nervioso de los moluscos se deriva del plan de protostomo básico de una disposición circumental anterior de ganglios y cordones nerviosos ventrales emparejados. En los moluscos, el más ventral y medial de los dos pares de cables nerviosos se llaman cordón del pedal (o cordones ventrales); inervan los músculos del pie. Los pares de nervios más laterales son los cordones viscerales (o cordones laterales); Sirven el manto y las vísceras. Las comisuras transversales interconectan estos pares de nervios longitudinales, que creando un sistema nervioso en forma de escalera. Este plan básico se ve con mayor facilidad en los moluscos primitivos, como los aplacóforos y los poliplacóforos. Sin embargo, el sistema nervioso de los moluscos difiere del de los anélidos y los artrópodos, en los que el cordón nervioso ventral lleva ganglios dispuestos en segmentos.

Figura 11.1. Sistema nervioso en escalera. Sistema nervioso ancestral de los moluscos, sin torsión corporal típico de (A) aplacoforos, (B) polipacoforos.

Figura 11.2. Sistema nervioso torsionado o en
estructura de 8.
11.1 Aplacóforos, monoplacóforos y poloplacóforos
En los moluscos más simples, como los
aplacoforos, monoplacoforos y poliplacoforos, los ganglios están poco
desarrollados. Un simple anillo nervioso rodea el esófago, a menudo con
pequeños ganglios cerebrales a cada lado. Cada ganglio cerebral, o el propio
anillo nervioso, emite pequeños nervios a la región bucal y da lugar al pedal y
a los cordones nerviosos viscerales.
La mayoría de los otros moluscos tienen
ganglios más bien definidos. Sus sistemas nerviosos están construidos alrededor
de tres pares de ganglios grandes que se interconectan para formar un anillo
nervioso parcial o completo alrededor del intestino (Figura 11.2).
Dos pares, los ganglios cerebral y pleural, se encuentran dorsal o lateral al
esófago, y un par, los ganglios de pedal, se encuentran ventral al intestino,
en la parte anterior del pie.
11.2 Cefalópodos, bivalvos y gasterópodos
En los cefalópodos, bivalvos y
gasterópodos avanzados, los ganglios cerebrales y pleurales suelen estar
fusionados. Desde los ganglios cerebrales, los nervios periféricos inervan los
tentáculos, los ojos, los estatocistos y la superficie general de la cabeza,
así como los ganglios bucales, con centros especiales de control para la región
bucal, el aparato radular y el esófago.
Los ganglios pleurales dan lugar a los
cordones viscerales, que se extienden posteriormente y suministran nervios
periféricos a las vísceras y al manto. Los cordones viscerales eventualmente se
unen a un par de ganglios parietales (= intestinales, = paliales) y desde allí
pasan para terminar en ganglios viscerales emparejados. Los ganglios parietales
inervan las branquias y el osfradio, y los ganglios viscerales sirven o se
dilatan en la masa visceral. Los ganglios del pedal también dan lugar a un par
de cordones nerviosos del pedal que se extienden posteriormente y proporcionan
nervios a los músculos del pie.
👉 Gastropodos: Como resultado de la torsión, la
porción posterior del sistema nervioso de gasterópodos se tuerce en una figura
ocho, una condición conocida como estreptoneuria (Figura 11.2).
Además de torcer el sistema nervioso, la torsión lleva los ganglios posteriores
hacia adelante. En muchos gasterópodos avanzados, esta concentración anterior
del sistema nervioso se acompaña de un acortamiento de ciertos cordones
nerviosos y la fusión de ciertos ganglios. En la mayoría de los gasterópodos
deformados, el sistema nervioso muestra una simetría bilateral derivada secundariamente
y cordones nerviosos más o menos rectos, paralelos y viscerales, una condición
conocida como eutineuria.
👉 Bivalvos: En los bivalvos, el sistema nervioso es claramente bilateral y la
fusión lo ha reducido a tres ganglios grandes y distintos. Los ganglios
cerebropleurales anteriores dan lugar a dos pares de cordones nerviosos, uno
que se extiende posterodoralmente a los ganglios viscerales, y el otro que
conduce ventralmente a los ganglios de pedal (Figura 11.3).
Los dos ganglios cerebropleurales están unidos por una comisura dorsal sobre el
esófago. Los ganglios cerebropleurales envían nervios a los palpos, el músculo
aductor anterior y el manto. Los ganglios viscerales emiten nervios al
intestino, corazón, branquias, manto, sifón y músculo aductor posterior.

Figura 11.3. Sistema nervioso de los bivalvos.
👉 Cefalópodos: El grado de desarrollo del sistema nervioso dentro de los
cefalópodos es inigualable entre los invertebrados. Aunque los ganglios
apareados que se ven en otros moluscos también son reconocibles en los
cefalópodos, se ha producido una cefalización extrema. La mayoría de los
ganglios se han desplazado hacia adelante y se concentran como lóbulos de un
cerebro grande que rodea el intestino anterior (Figura 11.4).

Figura 11.4. Sistema nervioso de los cefalópodos.
Además de los nervios de la cabeza
habituales que se originan en el ganglio cerebral, un gran nervio óptico se
extiende a cada ojo. En la mayoría de los cefalópodos, gran parte del cerebro
está encerrado en un cráneo cartilaginoso. Los lóbulos de pedal suministran
nervios al embudo, y las divisiones anteriores de los ganglios de pedal,
llamados lóbulos braquiales, envían nervios a cada uno de los brazos y
tentáculos, una disposición que sugiere que el embudo y los tentáculos se
derivan del pie del molusco. Los pulpos pueden ser los invertebrados "más
inteligentes", ya que se les puede enseñar algunas tareas dependientes de
la memoria con bastante rapidez.
Muchos cefalópodos muestran un
comportamiento de escape rápido que depende de un sistema de fibras motoras
gigantes que controlan las contracciones potentes y sincrónicas de los músculos
del manto. El centro de comando de este sistema es un par de neuronas gigantes
de primer orden muy grandes en el lóbulo de los ganglios viscerales fusionados.
Aquí, se realizan conexiones a neuronas gigantes de segundo orden que se
extienden a un par de ganglios estrellados grandes. En los ganglios
estrellados, se realizan conexiones con neuronas gigantes de tercer orden que
inervan las fibras musculares circulares del manto. Otros nervios se extienden
posteriormente desde el cerebro y terminan en varios ganglios que inervan las
vísceras y las estructuras en la cavidad del manto.
Durante varias décadas, los neurobiólogos
han utilizado los axones gigantes de Loligo
como un sistema experimental para el estudio de la fisiología y la mecánica de
los nervios, y gran parte de nuestro conocimiento fundamental sobre cómo
funcionan las células nerviosas se basa en la neurología de los calamares. La
liebre marina Aplysia, un
opistobranquio, se ha utilizado de la misma manera y, aunque carece de axones
gigantes, posee neuronas y ganglios excepcionalmente grandes que pueden
empalarse fácilmente con microelectrodos.
12. Sistema nervioso de los artrópodos
|| Regresar al Índice ||
El plan general del sistema nervioso de artrópodos es casi idéntico al de los anélidos, y existen muchas homologías obvias.

Figura 12.1. Sistema nervioso de los artrópodos y
algunos órganos sensoriales. (A) El sistema nervioso central
de un cangrejo de río, que muestra el plan básico anélido (vista dorsal). (B)
El cerebro de un crustáceo. (C) El cerebro de un quelicerado.
El "cerebro" de artrópodos
(ganglios cerebrales) en realidad comprende dos o tres regiones, cada una de
las cuales es un conjunto separado de ganglios segmentarios unidos. Los
ganglios anteriores forman el protocerebro dorsal (supraesofágico) y, cuando
está presente, el deutocerebro. El ganglio más posterior, el tritocerebro,
generalmente forma conectivos circumentericos alrededor del esófago a un
ganglio subesofágico ventral (subentérico). Este último está formado por la
fusión de varios otros ganglios de la cabeza, generalmente los asociados con
las mandíbulas y los maxilares. Un cordón nervioso ventral ganglionado doble o
simple se extiende a través de algunos o todos los segmentos del cuerpo. Los
crustáceos, los hexapodos y los miriápodos poseen un cerebro tripartito
compuesto por los dos ganglios anteriores (protocerebrum y deutocerebrum) y el
ganglio posterior (tritocerebrum). En los queliceriformes, el deutocerebrum
está ausente. Cada una de estas regiones da lugar a un par importante de
nervios a los apéndices particulares de la cabeza (Figura 12.1 B,
C).
Los ganglios segmentarios del cordón
nervioso ventral muestran varios grados de fusión lineal entre sí en diferentes
grupos de artrópodos. Por lo tanto, así como la tagmosis se refleja en la unión
externa de los segmentos del cuerpo, también es evidente en la unión de grupos
de ganglios a lo largo del cordón nervioso ventral.
12.1 Crustáceos
El sistema nervioso central de los crustáceos se construye en concierto con el bauplan segmentado, en la misma línea que hemos visto en los anélidos y otros artrópodos (Figura 12.2). En la condición más primitiva es como una escalera (Figura 12.2 A), los ganglios segmentarios están en gran parte separados y unidos por comisuras transversales y conectivos longitudinales. El cerebro de los crustáceos está compuesto por tres ganglios fusionados, siendo los dos anteriores el protocerebro dorsal (supraesofágico) y el deutocerebruo, que se cree que son de origen preoral. Desde el protocerebro, los nervios ópticos inervan los ojos. Desde el deutocerebro, los nervios antenulares pasan a las antenulas, mientras que los nervios más pequeños inervan la musculatura de los tallos oculares. El tercer ganglio del cerebro es el tritocerebro posterior, que presumiblemente representa el primer ganglio somático postoral. El tritocerebro forma un par de conectivos circumentericos que se extienden alrededor del esófago a un ganglio subesofágico o subenterérico y conectan el cerebro con el cordón nervioso ventral que lleva los ganglios del cuerpo segmentario. Del tritocerebro también surgen los nervios antenarios, así como ciertos nervios sensoriales de la región anterior de la cabeza.

Figura 12.2.
Sistema nervioso central de cuatro crustáceos. (A)
El sistema en forma de escalera de un anostracan. Obsérvese la ausencia de
ganglios bien desarrollados en la porción posterior, apodous, del tronco. (B)
Sistema metamérico alargado de un cangrejo de río. (C) Sistema altamente
compactado de un cangrejo brachyuran, en donde todos los ganglios torácicos se
han fusionado y los ganglios abdominales se reducen. (D) Sistema nervioso de un
anfípodo hiperiido. Tenga en cuenta la pérdida de los ganglios urosómicos
típicos de todos los anfípodos.
La naturaleza del cordón nervioso ventral
a menudo refleja claramente la influencia de la tagmosis corporal. En los
crustáceos primitivos con cuerpos relativamente homónimos (p. Ej., Remiidos,
cefalocaridos y muchos ramiópodos), los ganglios asociados con cada segmento se
mantienen separados a lo largo del cordón nervioso ventral. Sin embargo, en las
formas avanzadas, se forma una masa ganglionar subenterica única por la fusión
de los ganglios asociados con los segmentos cefálicos postorales (p. Ej., Los de
las mandíbulas, los maxilares etc).
Los ganglios del tórax y el abdomen
también pueden fusionarse de manera variable, dependiendo de la fusión del
segmento y la compactación del cuerpo. Por ejemplo, en la mayoría de los
decápodos de cuerpo largo (langostas y cangrejos de río), los ganglios
torácicos y abdominales se fusionan principalmente en la línea media del
cuerpo, pero permanecen separados uno del otro longitudinalmente (Figura 12.2 B).
Sin embargo, en los decápodos de cuerpo corto (p. Ej., Cangrejos), todos los
ganglios torácicos segmentarios se fusionan para formar una placa nerviosa
ventral grande, y los ganglios abdominales se reducen mucho (Figura 12.2 C).
Incluso en la langosta espinosa (Panulirus), los ganglios se fusionan en
gran medida y todo el sistema se concentra en la región anterior del cuerpo.
12.2 Insectos
El sistema nervioso hexápodo se ajusta al plan básico de artrópodos descrito en la introducción. Las dos hebras del cordón nervioso ventral, así como los ganglios segmentarios, a menudo se fusionan en gran medida.

Figura 12.3. Sistema nervioso de los insectos. (A)
El insecto sistema nervioso central. (B) Vista frontolateral del cerebro de una
langosta (Orthoptera).
Los indipteranos, por ejemplo, incluso los
tres ganglios torácicos se fusionan en una sola masa. La mayor cantidad de
ganglios libres se produce en los insectos primitivos sin alas, que tienen
hasta ocho ganglios abdominales no fusionados. También se han reportado fibras
gigantes de varias órdenes de insectos.

Figura 12.4. Sistema nervioso de los insectos 2.
Órganos endocrinos y sistema nervioso central en la cabeza y el tórax de un
insecto generalizado. Todos estos órganos juegan un papel en el control de la
muda y la metamorfosis.
Al igual que los "cerebros" de otros artrópodos, el ganglio cerebral de los insectos comprende tres regiones distintas: el protocerebro, el deutocerbro y el tritocerebro. El ganglio subesofágico está compuesto por los ganglios fusionados del tercer, cuarto y quizás el quinto segmento de la cabeza y controla las piezas bucales, las glándulas salivales y alguna otra musculatura local.

Figura 12.5. Sistema nervioso de los miriápodos. El
cerebro y los ganglios anteriores de un ciempiés, Lithobius forficatus
(vista dorsal).
Los insectos poseen un ganglio
hipocerebral entre el ganglio cerebral y el intestino anterior. Asociados con
este ganglio hay dos pares de cuerpos glandulares llamados los cuerpos
cardiacos y los cuerpos allata (Figura 12.4).
Estos dos órganos trabajan en concierto con las glándulas protorácicas y
ciertas células neurosecretoras en el protocerebro. Todo el complejo es un
importante centro endocrino que regula el crecimiento, la metamorfosis y otras
funciones.
12.3 Miriapodos
El
sistema nervioso miriápodo se ajusta al plan básico de artrópodos. Se produce
muy poca fusión secundaria de ganglios, y el cordón nervioso ventral retiene
gran parte de su doble naturaleza primitiva, con un par de ganglios fusionados
en cada segmento. Los milpiés poseen dos pares de ganglios fusionados en cada segemento
(Figura 12.5). Al
igual que los "cerebros" de otros artrópodos, el ganglio cerebral de
los miriápodos comprende tres regiones distintas: el protocerebro (asociado con
los ojos), el deutocerebro (asociado con las antenas) y el tritocerebro. El
ganglio subesofágico está compuesto por ganglios fusionados del tercer, cuarto
y quizás el quinto segmento de la cabeza y controla las piezas bucales, las
glándulas salivales y alguna otra musculatura local.
12.4 Quelicerados
Como en todos los artrópodos, la forma del cuerpo externo de los queliceratos generalmente se refleja en la estructura del sistema nervioso central. Estos animales muestran diversos grados de compactación y fusión de las somitas del cuerpo y los componentes del sistema nervioso asociados, mientras que aún se ajustan al plan básico anélido-artrópodo. Los ganglios cerebrales, o cerebro, incluyen el protocerebro y el tritocerebro; el deutocerebrum está ausente. El tritocerebro generalmente contribuye a los conectivos circulatorios, que se unen ventralmente con una gran masa ganglionar formada en parte por la fusión de los ganglios anteriores emparejados del nervio ventral.

Figura 12.6. Sistema nervioso de los quelicerados. El
sistema nervioso central de (A) un escorpión y (B) una araña.
En los
xifosuranos y los escorpiones, esta masa neuronal subenterica incluye todos los
ganglios prosomales, mientras que en las arañas incluso los ganglios
optostomales se fusionan anteriormente. Por lo tanto, en la mayoría de las
arañas, el sistema nervioso adulto ya no está obviamente segmentado (excepto en
algunos miembros del suborden Mesothelae), aunque una cadena de ganglios
ventrales es evidente durante el desarrollo. El cordón nervioso ventral
persiste en el opistoesoma de los xifosuranos y tiene cinco ganglios
segmentarios; en los escorpiones tiene siete ganglios (Figura 12.6A).
El protocerebro y
el tritocerebro producen nervios en los ojos y los quelíceros, especialmente.
En las arañas, los nervios quelicerales emergen realmente del protocerebro,
pero se pueden rastrear histológicamente hasta su origen en la parte posterior
de los ganglios cerebrales debajo del intestino. La masa ganglionar ventral
(subenterica), que incluye los ganglios prosomales segmentarios fusionados,
origina tractos nerviosos en las piernas y, en las arañas, lleva un par de
ganglios abdominales de los cuales surgen nervios ramificados hacia el
opistosoma. Los ganglios segmentarios en el cordón nervioso ventral en los
xifosuranos y los escorpiones sirven a los apéndices optostomales, músculos y
órganos sensoriales.
13. Sistema nervioso de algunos deuterostomos
|| Regresar al Índice ||
13.1 Equinodermos
El bauplan radial derivado de forma secundaria de los equinodermos se refleja claramente en la anatomía de sus sistemas nerviosos y la distribución de sus órganos sensoriales. El sistema nervioso está descentralizado, algo difuso y sin ganglio cerebral. Hay tres redes neuronales principales, integradas entre sí y desarrolladas en varios grados entre las clases. Estas redes son el sistema ectoneural (oral), el sistema hiponeural (oral profundo) y el sistema entoneural (aboral). El sistema ectoneural es principalmente sensorial, aunque se producen fibras motoras; El sistema hiponeural es en gran parte motor en función. El sistema entoneural está ausente de los holoturianos y se reduce en diferentes grados en otros grupos, excepto los crinoides, en los que es el componente nervioso primario y cumple funciones motoras y sensoriales.

Figura 13.1. Sistema nervioso de los equinodermos. El
sistema nervioso de cinco radios de los erizos puede verse con claridad en el
modelo anterior coloreado de negro que recorren el cuerpo en paralelo con el
sistema vascular de ampullas. En las
estrellas de mar los esquemas visualizan el sistema nervioso de cinco radios de
forma más simplificada.
Los tres "sistemas" nerviosos
están interconectados por una red nerviosa derivada principalmente de los
componentes ectoneural y entoneural. La red nerviosa a menudo se describe como
un plexo subepidérmico, pero da lugar a neuronas intraepidérmicas y claramente
tiene una asociación íntima con el epitelio. A excepción de los crinoides, en los que
domina el componente entoneural, los nervios más obvios en los equinodermos se
derivan del sistema ectoneural. Un anillo circular o pentagonal del nervio
circumoral se encuentra justo debajo del epitelio oral y rodea el esófago. De
este anillo surgen nervios radiales que se extienden a lo largo de cada
ambulatorio. En las estrellas de mar, por ejemplo, estos nervios radiales
aparecen como un engrosamiento distintivo en forma de V en la epidermis de cada
surco ambulacral.
En algunos casos, los componentes
entoneurales del plexo nervioso también se producen como cordones radiales,
como los que se encuentran a lo largo de los márgenes laterales de los brazos
de los asteroideos. El sistema hiponeural generalmente es paralelo a los
nervios del sistema ectoneural. Las neuronas hiponeurales son subepidérmicas y
se encuentran cerca del seno hiponeural de cada área ambulacral. Estas neuronas
dan lugar a fibras motoras y ganglios en los pies del tubo.
13.2 Cefalocordados
Un cordón nervioso dorsal se extiende la mayor parte del cuerpo y generalmente se expande ligeramente como una vesícula cerebral en la base de la capucha oral. Los nervios dispuestos en segmentos surgen del cordón a lo largo del cuerpo en el patrón típico de vertebrados con raíces dorsales y ventrales (Figura 13.2).

Figura 13.2. Sistema nervioso de los cefalocordados. Una
vista lateral muy esquemática de una lanceta (Brachiostoma sp.) Con 63
miómeros (m1 a m63), que muestra la organización general de los sistemas
nerviosos central y periférico en relación con las estructuras periféricas. Los
primeros cuatro y últimos tres miembros de la serie de miómeros están
resaltados (sombreado claro) para aclarar la relación entre el número de
miómero y el número de nervio. Las regiones ocupadas por bloques principales de
músculo no miomérico están indicadas por un sombreado más oscuro.
13.3 Tunicados
El sistema nervioso de los tunicados está muy reducido y refleja su estilo de vida planctónico sésil y flotante relativamente inactivo.

Figura 13.3. Sistema nervioso de los tunicados. Es
relevante, y visible en los modelos, solo en su etapa de larva (c), pero este
se reduce a nervios poco prominentes que no alcanzan a ser representativos en
el adulto (b).
Un pequeño ganglio cerebral se encuentra
justo dorsal al extremo anterior de la faringe y da lugar a algunos nervios en
varias partes del cuerpo, especialmente los músculos y las áreas sifonales. Un
cordón nervioso dorsal bien desarrollado está presente en las colas de las
larvas de tunicado, pero esta estructura se pierde durante la metamorfosis,
excepto en los apendicularianos. La mayoría de los tunicados poseen una
glándula neural, o subneural, ubicada entre el ganglio cerebral y la porción anterodorsal
de la faringe (Figura 13.3).
Esta glándula se abre a la faringe a través de un pequeño conducto, pero su
función es desconocida. Algunos autores han sugerido que puede ser el precursor
de la glándula pituitaria de los vertebrados. Los receptores sensoriales están
poco desarrollados en los tunicados, aunque las neuronas sensibles al tacto
prevalecen alrededor de los sifones.
14. Sistema nervioso periférico de los vertebrados
|| Regresar al Índice ||
El sistema
nervioso de los seres humanos es un buen ejemplo de un sistema nervioso
vertebrado, por lo que podríamos afirmar que ya hemos trabajado a grandes
rasgos varios de los aspectos fundamentales: neuronas, potenciales de acción,
neurotransmisión entre otros. Cuestiones que analizamos en detalle ya. En la
presente sección analizaremos brevemente la anatomía comparada de los
vertebrados en cuanto al sistema nervioso. Para ello asumiremos la
diferenciación normal, primero analizaremos el sistema nervioso periférico y
luego terminaremos con el sistema nervioso central.

Figura 14.1. SNP. El sistema
nervioso periférico de los vertebrados refleja el patrón cefalocordado, tenemos
varios ganglios cerebrales protegidos por un cerebro “región central” del cual
emerge una médula en el dorso del cual salen los nervios “periféricos”
Los términos
empleados para describir los componentes del sistema nervioso periférico hacen
referencia a estructuras anatómicas o a sus propiedades funcionales. De allí se
ha genera una nomenclatura relativamente compleja para describirlos. Nervios
somáticos, son aquellos que atraviesan los tejidos somáticos, especialmente el
músculo voluntario, pero también la piel y el esqueleto. Nervios viscerales,
atraviesan los músculos involuntarios, las glándulas y las vísceras. Los
nervios que transportan la información desde los tejidos al sistema nervioso
central se denominan aferentes o sensoriales. Los nervios que envían la
información desde el sistema nervioso central a los tejidos blancos se
denominan eferentes o motores.

Figura 14.2. Nervios periféricos. Los
principales nervios periféricos se nombran en base a las vértebras de las que
emergen.
Dado que los dos sistemas de categoría no son excluyentes, sino complementarios, esto nos crea cuatro categorías en pareja: Los nervios somáticos eferentes o nervioso somáticos motores, que envían información del movimiento del músculo esquelético. Los nervios somáticos aferentes o nervios somáticos sensoriales que transportan la información desde los órganos de los sentidos, especialmente desde la piel al sistema nervioso central. Los nervios viscerales eferentes se encargan de hacer mover los órganos internos por medio de las peristalsis y del movimiento cardíaco, mientras que los nervios viscerales aferentes o sensoriales se encargan de recolectar la información sensorial de las vísceras. Los componentes del sistema nervioso periférico que inerva las vísceras por lo general no se controlan voluntariamente, de allí que también se lo denomina, sistema nervioso involuntario.

Figura 14.3. Los nervios espinales. Las
inervaciones se hacen más complejas en la medida que algunos miotomos se
fusionan o combina para dar lugar a las diferentes estructuras de los
vertebrados.
Los nervios poseen dos propiedades
adicionales, basados en su distribución. Las neuronas se pueden clasificar como
generales si inervan tejidos de distribución general a lo largo del cuerpo, o
pueden ser clasificadas como especiales si inervan órganos sensoriales especializados
u órganos muy especializados, donde dichas neuronas se encuentran de manera
endémica. Desde el punto de vista anatómico, los nervios periféricos pueden
dividirse en nervios espinales que emanan de la médula espinal y nervios
craneales que emanan directamente desde el cerebro.
14.1 Nervios espinales
Son aquellos que emergen de la médula a lo largo del dorso “espalda”. Los nervios espinales se encuentran organizados de manera secuencial y en consecuencia se los nombra de manera sistemática (C-1, T-1, L-1, S-1) de acuerdo a su asociación con las regiones de la columna vertebral y correspondientes vértebras cervicales, torácicas, lumbares y sacras. Los primeros anatomistas reconocieron las raíces dorsales y ventrales de cada uno de los nervios espinales.

Figura 14.4. Nervios cerebrales. Los
principales nervios craneales se nombran por un código numérico o por medio de
las estructuras que inervan.
Las
fibras aferentes ingresan a la médula espinal por vía de las raíces dorsales, y
los nervios eferentes emergen por medio de las raíces ventrales.
Adicionalmente, cerca de la raíz dorsal los nervios se inervan en regiones que
denominamos ganglios, donde se ubican los cuerpos neuronales contribuyendo a
los nervios espinales. Paralelo a la médiala espinal y unidos a cada nervio
espinal a través del ramus communicans se encuentra la cadena simpática
de ganglios o ganglios paraventrales. Los ganglios paraventrales son una serie
de parejas de ganglios unidas que se encuentran adyacentes a la columna
vertebral o notocordio.
Otros ganglios
periféricos forman los ganglios colaterales o ganglios prevertebrales. Ejemplos
de ganglios prevertebrales son los ganglios cervicales, coeliacos y
mesentéricos. Los ganglios viscerales se encuentran al interior de las paredes
de los órganos viscerales efectores. En base a lo anterior, existen tres tipos
de ganglios, somáticos, colaterales y viscerales. Las fibras de cada nervio
espinal inervan estructuras restringidas a nivel de la médula. Esto es
especialmente pronunciado en la inervación de los músculos esqueléticos. Cada
nervio espinal tiende a acompañar a su miotomo embrionario y estructuras
seccionales adjuntas, como hueso y piel.
14.2 Nervios craneales
Los nervios craneanos poseen raíces
encerradas en la cavidad craneana. La mayoría se encuentran nombrados y
numerados mediante numerales romanos desde la región anterior a la región
posterior. Debido a la compleja tradición histórica, la numeración de estos
nervios es en ocasiones inconsistente. Por ejemplo, la mayoría de las anatomías
establecen una numeración con base en 10 nervios craneanos numerados y seis
pares no numerados.
Algunos anamniotas y todos los amniotas
poseen hasta 12 nervios numerados. De hecho, existe un nervio terminal
adicional al principio de esta serie, generalmente se omite, pero si se lo toma
en cuenta lo que se hace es numerarlo como 0 para evitar renombrar los demás
nervios y crear confusiones. El segundo nervio craneano no es un nervio real,
sino la extensión del cerebro en sí mismo, encargado de procesar la información
visual, pero por tradición y convención se lo denomina simplemente como el
nervio óptico. Otros nervios son complejos, pues son fusiones, el XI nervio
craneano representa la fusión de una rama del nervio craneano X y elementos de
los primeros nervios cervicales espinales denominados C-1 y C-2. A pesar de su
naturaleza compuesta, es denominado el nervio espinal sensorial y designado con
el numeral romano XI.
En adición a estos nervios numerados,
cerca de seis parejas de nervios no numerados que se ubican lateralmente se
encuentran presentes en los peses mandibulados y muchos anfibios.
Filogenéticamente, la hipótesis más aceptada es que los nervios craneanos que
han evolucionado desde los nervios dorsales y ventrales de unos cuantos nervios
espinales que fueron incorporados al cráneo. Los nervios dorsales y ventrales
se fusionan en el tronco, pero no en la cabeza produciendo dos series: Los
dorsales V, VII, IX y X; y los ventrales III, IV, VI y XII. Al igual que los
nervios espinales, los nervios craneanos suplementan los tejidos somáticos y
viscerales y transportan información sensorial y motriz general. Algunos
nervios craneanos consisten únicamente en nervios sensoriales o nervios motores
de forma exclusiva, mientras que otros son mezclas, conteniendo ambos tipos
aferentes y eferentes. Los nervios craneanos relacionados con los órganos de
los sentidos se denominan nervios craneanos especiales.
Originalmente,
todos los nervios que servían a las hendiduras branquiales formaban tres ramas
por hendidura: la rama pretrematica, la rama posttrematica y la rama faríngea.
En los amniotas esta tendencia se complejiza debido a la pérdida de las
hendiduras branquiales que servían para organizar cada trio de nervios. De allí
que las homologías de los 17 nervios sobrevivientes se hagan incierta con
respecto a los tríos correspondientes a cada hendidura branquial. Algunos de
estos 17 nervios son reclutados hacia el cráneo.
En
los ciclóstomos estos nervios permanecen fuera del cráneo y se denominan
nervios accipitospinales. En otros peces y anfibios algunos nervios son
reclutados en el cráneo. Los celacantos y muchos anfibios “y puede esperarse
que todo lo que estaba en medio” poseen 17 nervios craneanos, algunos de los
cuales se perdieron hasta llegar a 12 nervios craneanos en el linaje de los
amniotas.
14.3 Nervio
terminal, 0 par
Se encuentra presente en todos los
vertebrados con mandíbula, excepto en las aves, emergiendo como un complejo de nervios
de las placas olfatorias. Viaja hasta los vasos sanguíneos del epitelio
olfativo en el saco olfatorio y transporta nervios viscerales sensoriales y
algunas fibras motoras. En los mamíferos inferiores está muy desarrollado y es
importante en la recepción de sustancias químicas no odoríficas (feromonas). La
adecuada recepción de estas sustancias químicas garantiza la supervivencia de
estos animales, ya que por medio de ellas se localiza a la madre, a la futura
pareja, a un posible depredador o a un posible alimento.
En el humano se ha demostrado la
existencia de este nervio durante la vida prenatal y durante los primeros meses
de vida extrauterina, involucionando hasta desaparecer casi por completo en el
adolescente. Sin embargo, algunos autores aseguran que persiste en la vida
adulta, percibiendo feromonas sexuales y explicando de esta manera las
emociones intensas que se pueden manifestar con la presencia de otra persona
desconocida, ya sea que nos agrade o nos disguste, sin tener explicación racional
alguna
14.4 Nervio
olfatorio, I par
Especializado con el sentido del olfato y
en consecuencia sus nervios se denominan como especiales. El nervio olfatorio
es un nervio únicamente sensorial, conduciendo impulsos olfatorios de la nariz
al sistema nervioso central, siendo clasificado como fibras aferentes
viscerales especializadas. Es posible encontrar células olfatorias
especializadas ante todo en la mucosa pituitaria, con una prolongación
periférica y una prolongación central, siendo verdaderas células nerviosas,
homólogas de las células del Ganglio de Corti y del Ganglio de Scarpa de la vía
auditiva y homólogas, por consiguiente, de las células constituidas de los
ganglios espinales. Anexo al nervio olfativo va el nervio terminal cuya función
es hasta ahora incierta; se presume de un nervio atrófico

Figura 14.5. I par. El nervio
olfatorio no solo es importante para el olfato, sino también para casi el 80%
de lo que denominamos como el sentido del gusto.
14.5 Nervio
óptico, II par
No un nervio, sino una evaginación del
cerebro en sí mismo relacionado con el procesamiento de la información visual.
Mide aproximadamente 4 cm de longitud, y se orienta en sentido anteroposterior.
Se describen en él cuatro segmentos:
👉 Primer segmento: intraocular. Los axones de las células ganglionares de la retina convergen en la papila óptica, desde allí el nervio perfora las capas superficiales del ojo (esclerótica y coroides) en un sitio llamado zona cribosa.

Figura 14.6. II par. Los nervios
ópticos están conectados directamente a las regiones primitivas e instintivas.
👉 Segundo
segmento: intraorbitario. El nervio queda
comprendido en un cono formado por los músculos rectos del ojo y se sumerge en
la grasa retroocular. En el vértice de la órbita por el anillo fibroso que
presta inserción a los músculos rectos (anillo de Zinn). En este segmento el
nervio se relaciona por arriba con la arteria oftálmica y por fuera con el
ganglio oftálmico.
👉 Tercer
segmento: intracanalicular. El nervio atraviesa el
foramen óptico acompañado por la arteria oftálmica. En este sitio, es frecuente
la lesión del nervio por fracturas que comprometan la base del cráneo y el
vértice de la órbita.
👉 Cuarto
segmento: intracraneal. Mide 1 cm y está
comprendido entre el foramen óptico y el quiasma óptico. En este segmento el
nervio está situado sobre la tienda de la hipófisis y sobre el conducto óptico
del esfenoides. En el quiasma óptico, todas las fibras que llevan información
de la hemirretina nasal pasan al lado contralateral, y las fibras de la
hemirretina temporal pasan ipsilaterales. A través del tracto óptico, las fibras
proyectan al núcleo geniculado lateral del tálamo, localizado en la profundidad
del cuerpo geniculado lateral. Las neuronas talámicas proyectan, a través de
las radiaciones ópticas, a la corteza visual primaria, o estriada, finalizando
fundamentalmente en la capa IV.
14.6 Oculomotor,
III par
Nervio motor que controla los músculos oculares, incluyendo aquellos que controlan la apertura del iris. El nervio oculomotor tiene dos núcleos motores: El núcleo motor principal; El núcleo parasimpático accesorio. El núcleo oculomotor principal se ubica en la parte anterior de la sustancia gris que rodea al acueducto cerebral del mesencéfalo, se ubica a nivel del colículo superior. El núcleo consiste en grupos de células nerviosas que inervan todos los músculos extrínsecos del ojo excepto el oblicuo superior y el recto lateral. Las fibras nerviosas eferentes se dirigen anteriormente a través del núcleo rojo y salen sobre la superficie anterior del mesencéfalo en la fosa interpeduncular. El núcleo principal del nervio oculomotor recibe fibras corticonucleares desde ambos hemisferios cerebrales.

Figura 14.7.
III par, IV par, VI par. El oculomotor
(III) como su nombre indica, junto con el troclear (IV) y el abducens (VI),
sirve para mover los ojos.
Recibe fibras tectobulbares del colículo
superior y a través de esta vía le llega información de la corteza visual.
También recibe fibras desde el fascículo longitudinal medial, por el cual está
conectado con los núcleos de los nervios craneanos cuarto, sexto y octavo. El
núcleo parasimpático accesorio (núcleo de Edinger - Westphal u oculomotor
accesorio) está ubicado por detrás del núcleo motor principal. Los axones de
las células nerviosas, que son preganglionares, acompañan a otras fibras
oculomotoras hasta la órbita. Aquí hacen sinapsis en el ganglio ciliar y las
fibras post ganglionares pasan a través de los nervios ciliares cortos hasta el
esfínter de la pupila del iris y los músculos ciliares. El núcleo parasimpático
accesorio recibe fibras corticonucleares para el reflejo de acomodación y
fibras del núcleo pretectal para los reflejos foto motor directo y consensual.
14.7 Troclear,
IV par
Se relaciona con el movimiento de otro
músculo ocular denominado el músculo superior oblicuo del ojo. Con el fin de
entender las acciones del músculo oblicuo superior, es útil imaginar el globo
ocular como una esfera que se ve limitada -como la rueda de desplazamiento de
un ratón de ordenador- de tal manera que de los movimientos de rotación sólo
algunos son posibles. Los movimientos permitidos para el oblicuo superior son:
(1) la rotación en un plano vertical -mirando hacia abajo y hacia arriba (la
depresión y elevación del globo ocular)-; y (2) la rotación en el plano de la
cara (intorsión y extorsión del globo ocular).
El cuerpo del músculo oblicuo superior se
encuentra detrás del globo ocular, pero el tendón (que es redirigido por la
tróclea) se acerca al globo ocular desde el frente. El tendón se adhiere a la
parte superior (cara superior) del globo ocular en un ángulo de 51 grados con
respecto a la posición primaria de los ojos (mirando recta hacia adelante). La
fuerza de tracción del tendón por lo tanto tiene dos componentes: un componente
que tiende hacia adelante para tirar del globo ocular hacia abajo (depresión),
y un componente interno que tiende a girar el globo ocular hacia fuera (torsión
externa).
La fuerza relativa de estas dos fuerzas
depende de la forma en que el ojo esté mirando. Cuando el ojo es abducido
(mirando hacia afuera), la fuerza de depresión aumenta. Cuando el ojo está
aducido (mirando hacia la nariz), la fuerza de intorsión aumenta, mientras que
la fuerza de depresión disminuye. Cuando el ojo está en la posición primaria
(mirando de frente), la contracción del músculo oblicuo superior produce
depresión y torsión externa en cantidades casi iguales.
14.8 Trigémino,
V par
Formado por tres ramas denominadas oftálmica V1, Maxilar V2 y Mandibular V3 en los amniotas.

Figura 14.8. V par. El trigémino es
un nervio famoso, debido a que está involucrado en la jaqueca, dolores de la
mitad o todo el rostro.
La función sensitiva del trigémino se
conforma por fibras somáticas que transportan impulsos exteroceptivos, como
sensaciones táctiles, de propiocepción y dolor de los 2/3 anteriores de la
lengua, contribuye a la sensorialidad de los 2/3 anteriores de la lengua
(fibras gustativas proporcionadas por el nervio cuerda del tímpano del VII par),
piezas dentarias, la conjuntiva del ojo, duramadre, la mucosa bucal, nariz, y
los senos paranasales, además de aproximadamente la mitad de la piel anterior
de la cabeza.
El nervio oftálmico conduce información
sensitiva del cuero cabelludo y frente, párpado superior, la córnea, la nariz,
la mucosa nasal, los senos frontales y partes de las meninges. El nervio
maxilar: conduce información sensitiva sobre el párpado inferior y la mejilla,
dorso y punta de la nariz, el labio superior, los dientes superiores, la mucosa
nasal, el paladar y el techo de la faringe, los senos maxilar etmoidal y
esfenoidal. El nervio mandibular: conduce información sensitiva del labio
inferior, dientes inferiores, alas de la nariz, barbilla, de dolor y
temperatura de la boca. La sensibilidad de los 2/3 anteriores de la lengua,
acompaña a una rama de este nervio, la lingual, aunque estos tipos de fibras
nerviosas luego se desvían para formar parte del par VII.
El trigémino es uno de los nervios más
famosos debido a la neuralgia del trigémino (NT o NTG), también conocida como
prosopalgia, tic doloroso, enfermedad suicida o enfermedad de Fothergill,
migraña crónica, es un trastorno neuropático del nervio trigémino que provoca
episodios de intenso dolor en los ojos, orejas, labios, nariz, cuero cabelludo,
frente, mejillas, boca, mandíbula y el lado de la cara.
14.9 Nervio abducens,
VI par
Tercero de los nervios craneanos que
controla el movimiento de los ojos. también conocido como nervio motor ocular
externo, nervio abducente o VI par craneal, es el nervio que se genera al lado
del bulbo raquídeo y posee como función el movimiento del músculo recto lateral
por lo que permite la abducción del ojo. El núcleo de este nervio está situado
en el fondo de la base del cuarto ventrículo, se encuentra rodeado por las
fibras emergentes del núcleo facial, con las cuales forma el colículo facial.
Tiene conexiones tanto a las vías voluntaria y sensitiva general, como al resto
de núcleos oculomotores y a las vías coclear y óptica estas por medio del
colículo superior.
Este nervio emerge en el borde inferior
del puente por arriba de la pirámide bulbar y pasa hacia adelante y arriba de
un lado a otro en la cisterna póntica por detrás, a veces por delante, de la
arteria cerebelosa anteroinferior. El nervio atraviesa la duramadre del
compartimiento craneal posterior y llega hasta el vértice del peñasco del hueso
temporal; dispuesto bajo una delgada banda ligamentosa, se dobla hacia adelante
en el ángulo que forman los senos. El motor ocular externo atraviesa el seno
por fuera, y después por fuera y afuera de la carótida interna. En seguida se
introduce en la órbita por la hendidura esfenoidal; dentro del anillo tendinoso
común de los músculos rectos (de zinn), está colocado por debajo del motor
ocular común, y, pasándose al recto, termina en su cara interna.
14.10 Nervio facial, VII par
Es un nervio craneal mixto, es decir, contiene fibras sensitivas como motoras, presente en mamíferos incluyendo humanos en el cual forma el séptimo par craneal o VII par. Por ser un par craneal, emite dos fibras, una que corre por el lado derecho de la cara y el contralateral por la izquierda. Parte del tallo cerebral, justo entre el puente troncoencefálico y el bulbo raquídeo y controla los músculos de expresión facial, así como el sabor en los dos tercios anteriores de la lengua. También suple inervación preganglionar parasimpática a varios ganglios nerviosos de la cabeza y el cuello.

Figura 14.9. VII par. El
nervio facial inerva casi todo el rostro.
14.11 Nervio auditivo,
VIII par
Es responsable del equilibrio y la función
auditiva. Es un nervio aferente de tipo sensorial. Consta de dos orígenes, uno
real y otro aparente. Origen Real: dos de sus partes tienen origen en los
ganglios periféricos, análogos a los nodos espinales. Origen Aparente: penetra
en el puente encefálico por el extremo lateral del surco bulbopontino. En este
punto emerge de la caja craneal transmite los impulsos auditivos del oído
externo. Está compuesto por el nervio coclear, que transporta la información
sobre el sonido y el nervio vestibular, que transporta la información sobre el
equilibrio.
El equilibrio se evalúa por la marcha y la estabilidad general del cuerpo que conecta el oído al tallo cerebral, transmitiendo los impulsos nerviosos desde las células pilosas en el oído interno hasta el cerebro, donde se realiza realmente la percepción auditiva. En rigor, su origen se encuentra en ganglios periféricos. Así, el nervio coclear comienza en el ganglio de Corti, y el vestibular, en el de Scarpa. La función auditiva se evalúa haciendo sonar los dedos bilateralmente en los oídos y preguntando al paciente si escucha los sonidos bilateralmente y si son parejos en intensidad. Las tumoraciones (neuromas acústicos) son frecuentemente responsables de disturbios (desequilibrio y la audición) en este nervio.

Figura 14.10. VIII par. El
nervio auditivo transmite la señal que ha sido convertida de mecánica a química
por los receptores auditivos, directamente a regiones primitivas del cerebro.
14.12 Nervio
glosofaríngeo, IX par
Es un nervio mixto que emerge del bulbo raquídeo del encéfalo (del latín encefalus: cabeza humana) y pasa anterolateralmente para abandonar el cráneo a través de la fosa yugular junto a los X y XI. En esta fosa están los ganglios inferiores (ganglio de Andersch) y superior (ganglio de Ehrenritter) de este nervio, que contienen los cuerpos celulares para los componentes aferentes del nervio. El nervio craneal IX sigue al estilofaríngeo y pasa entre los músculos constrictores superior y medio de la faringe para alcanzar la orofaringe y la lengua. Contribuye al plexo nervioso faríngeo. El nervio glosofaríngeo es aferente de la lengua y la faringe, de ahí su nombre, y eferente para el estilofaríngeo y la glándula parótida.

Figura 14.11. IX par. El nervio
glosofaríngeo controla las estructuras derivadas de los arcos branquiales, lo
cual incluye regiones del oído, la lengua, la boca, y la faringe.
Origen aparente: surco lateral del bulbo.
Origen real: Fibras Sensitivas: nacen de ángulo externo de la sustancia
reticulada gris, descienden en sentido vertical al fascículo solitario, sigue
longitudinalmente. Su volumen aumenta de abajo a arriba por ramas descendentes
del Intermediario, neumogástrico al núcleo solitario, termina en la parte media
y superior de núcleo (Gustativo de Nageotte). Fibras motoras: nacen del núcleo
ambiguo comienza en la parte superior del entrecruzamiento sensitivo y termina
en la oliva bulbar, representa la cabeza de la asta anterior. Límites: Sale por
la fosa yugular, pasa acompañado de X y XI, se dirige a la base de la lengua
(va en sentido del pabellón auricular hasta la lengua) pasa entre la vena
yugular interna y la arteria carótida interna, rodea la arteria sigue la pared
lateral de la faringe y se distribuye por la mucosa de la lengua.
El nervio glosofaríngeo es: (1) Sensitivo
general de la mucosa de la faringe, la amígdala palatina, el tercio posterior
de la lengua, la trompa auditiva y el oído medio. (2) Sensitivo para la presión
arterial y la química del seno carotídeo y el cuerpo carotídeo. (3) Motor y
propioceptivo para el estilofaríngeo. (4) Parasimpático (secretomotor) para la
glándula parótida.
14.13 Nervio
vago, X par
Nace del bulbo raquídeo e inerva la faringe, el esófago, la laringe, la tráquea, los bronquios, el corazón, el estómago, el páncreas, el hígado y vísceras que irriga la arteria mesentérica superior por plexos periarteriales que llegan a los plexos mientéricos o de Auerbach donde se localizan las fibras postsinápticas parasimpáticas.

Figura 14.12. X par. El nervio vago
se encuentra distribuido por todo el cuerpo, pero inerva de manera
independiente a los nervios espinales, este controla órganos vitales, y explica
la razón por la cual los órganos vitales pueden seguir funcionando, mientras
que los nervios del movimiento pueden bloquearse en lesiones espinales graves.
Su origen real se encuentra en las células
del ganglio petroso, que terminan a nivel del tracto solitario del bulbo. Su
origen aparente está entre los nervios craneales accesorio (XI) y glosofaríngeo
(IX), en el surco colateral posterior del bulbo raquideo o surco retroolivar.
Su
trayecto comienza en las células del ganglio petroso, luego atraviesa el
agujero yugular (rasgado posterior) de la base del cráneo y llega al espacio
retroestíleo. En dicho espacio se une a la arteria carótida interna y a la vena
yugular interna formando con ellas el paquete vasculonervioso principal del
cuello. Así, desciende por el cuello englobado en este paquete, posteromedial
al músculo esternocleidomastoideo (ECM). En su descenso emite el nervio laríngeo
superior, y también da ramos para la faringe. Una vez dentro del tórax, los
nervios vagos derecho e izquierdo se comportan de manera diferente:
Nervio
vago izquierdo. Entra en el tórax entre las arterias carótida izquierda y
subclavia izquierda, y a la altura del arco aórtico emite el nervio laríngeo
recurrente izquierdo. Después se va hacia abajo y hacia adelante (se hace
anterior) y pasa por detrás del pedículo pulmonar antes de alcanzar el esófago,
donde contribuye a formar el plexo esofágico. Nervio vago derecho. Cruza por
delante de la arteria subclavia derecha, y a esta altura emite el nervio
laríngeo recurrente derecho. Después se dirige hacia abajo y hacia atrás (se
hace posterior) y pasa por detrás del pedículo pulmonar derecho antes de alcanzar
el esófago, donde también contribuye a formar el plexo esofágico, al igual que
su homólogo izquierdo.
Dentro
del tórax, los nervios vagos dan ramas para el plexo cardíaco y el plexo
pulmonar. Ambos nervios vagos realizan la última parte de su trayecto por el
tórax junto con el esófago, y junto a él se introducen la cavidad abdominal,
atravesando el diafragma a través del hiato esofágico. Una vez en la cavidad
abdominal, el nervio vago izquierdo se distribuye por el estómago, mientras que
el nervio vago derecho finaliza en el plexo solar desde donde da ramas para las
vísceras abdominales (estómago, intestinos, riñones e hígado).
14.14 Nervio
espinal accesorio, XI par
Principalmente
es un nervio motor. Además, facilita la inervación propioceptiva de los músculos
por él movilizados y de esta forma desempeña un importante papel en relación
con el tono de los músculos del cuello.

Figura 14.13. XI par. Nervio espinal
accesorio.
Es el
responsable del movimiento cefalogiro por su inervación del musculo
esternocleidomastoideo y produce de esta forma la rotación de la cabeza hacia
el lado opuesto, al mismo tiempo que la inclina hacia su lado. La inserción de
este músculo en la mastoides, un poco por detrás del eje transverso de los
cóndilos hace de él un extensor de la cabeza más bien que un flexor, a menos
que la cabeza se halle previamente flexionada. Esta función extensora del
espinal está acentuada todavía más por el hecho de que inerva un músculo
posterior, el trapecio, que proyecta la cabeza hacia atrás.
El
espinal es ante todo el nervio que permite realizar la extensión de la cabeza
hacia atrás. Los músculos por el inervados actúan mediante su inserción
inferior sobre el orificio superior del tórax, el ECM lo eleva durante la
inspiración, el trapecio tira y lleva la clavícula y el hombro hacia arriba y
hacia atrás, dirige los hombros hacia dentro y de esta forma el espinal entra a
formar parte de los mecanismos de inhalación forzada. Mediante sus raíces
bulbares, antes de confundirse con el neumogástrico, el nervio participa en la
inervación de la laringe interviniendo funcionalmente en el estrechamiento de
la glotis y en la función fonatoria.
14.15 Nervio
hipogloso, XII par
Es un nervio craneano que se encarga especialmente de coordinar los movimientos de la lengua. Se encuentra en el cráneo y se extiende hasta llegar al borde lateral de la lengua.

Figura 14.14. XII par. El
nervio hipogloso también está relacionado con la lengua, especialmente en su
control más fino.
Tiene 7 ramos colaterales: Ramo meníngeo o
recurrente; Ramos vasculares; Ramo anastomótico para el ganglio plexiforme del
neumogástrico; Nervio del tirohioideo; Nervio del hiogloso y del estilogloso;
Ramo anastomótico para el nervio lingual; Nervio del genihioideo. El nervio
hipogloso es motor para los músculos intrínsecos y extrínsecos de la lengua
(estilogloso, hiogloso, geniogloso). También transmite fibras motoras desde los
nervios espinales C1 y C2 a los músculos infra hioideos menos el tirohioideo
(Inervado por el glosofaríngeo), fibras propioceptivas para estos músculos y
sensitivas generales para la duramadre de la fosa craneal posterior.
14.16 Nervios
de la línea lateral
En adición a los nervios numerados, los
peces poseen unos nervios de línea lateral preoticos y postoticos. Sirve para
detectar el movimiento y las vibraciones del agua circundante, lo que ayuda al
pez a evitar colisiones, a orientarse en relación a las corrientes de agua, y
localizar la presa. Las líneas laterales comúnmente son visibles como tenues
líneas que recorren cada lado desde las cercanías del opérculo (estructura que
cubre las branquias) hasta la base de la cola. Algunas veces partes de este
órgano son modificadas como electrorreceptores, los cuales son órganos que
detectan impulsos eléctricos. Es posible que vertebrados como los tiburones
usen también las líneas laterales para detectar campos magnéticos.

Figura 14.15. Sexto sentido. Los
nervios de la línea lateral son exclusivos de los peces. La forma más avanzada
de este órgano la tienen los tiburones y parientes cercanos, especialmente el
pez martillo, pues le permite detectar sutiles campos eléctricos de sus presas.
Los receptores que componen la línea
lateral son los neuromastos, cada uno compuesto de un grupo de células
ciliadas. Los cilios están rodeados por una cúpula sobresaliente, típicamente
1/20 a 1/5 mm de largo. Las células ciliadas y las cúpulas de los neuromastos
están comúnmente bajo el agujero visible en los peces. Las células ciliadas de
la línea lateral son similares a las células ciliadas que se encuentran en el
oído interno, indicando que la línea lateral y el oído interno comparten un
origen común.
Los teleósteos y eslasmobranquios tienen
comúnmente canales de la línea lateral, en los cuales los neuromastos no están
directamente expuestos al ambiente, pero se encuentran comunicados mediante
poros del canal, sin embargo, dicha línea lateral se halla directamente expuesta
al ambiente en algunos elasmobranquios como en Clamydoselachus (ALLIS, 1923), Notorynchus
(Daniel, 1922) y en
Echinorhinchus (Garrick, 1960).
Pueden aparecer neuromastos adicionales en varios lugares de la superficie
corporal, el desarrollo del sistema de la línea lateral depende del modo de
vida del pez. Por ejemplo, los peces que son nadadores activos tienden a tener
más neuromastos en los canales, y la línea se ubicará más lejos de las aletas
pectorales, lo que probablemente reduce la cantidad de "ruido" que se
genera por el movimiento de las aletas. La mayoría de las larvas de anfibios o
renacuajos y algunos anfibios que son acuáticos toda la vida tienen receptores
sensoriales similares a la línea lateral.
14.17 Los reflejos
Los reflejos espinales exhiben el nivel más simple de control al interior del sistema nervioso. A pesar de que los reflejos pueden dispersar información a niveles de control más altos, todos sus componentes funcionales se encuentran enraizados en la médula espinal.

Figura 14.16. Los reflejos. Los
nervios reflejos son nervios que se saltan la burocracia del cerebro, y permite
realizar movimientos sin tener que pensarlos.
El reflejo espinal es un circuito de neuronas sensoriales y motoras que puede regular una respuesta sin que esta tenga que llegar el cerebro, en cierto sentido el cuerpo se mueve sin que el cerebro sepa la razón. La mayoría de los reflejos somáticos están compuestos por tres neuronas, una sensorial somática, una sensorial motora y una neurona de regulación que une a las dos anteriores. Sin embargo, existen otros tipos de arcos reflejos, denominados arcos reflejos viscerales, que son estructuralmente más complejos involucrando ganglios y una mayor cantidad de neuronas. En lugar de una neurona motora el arco visceral emplea dos neuronas motoras.
14.18 Sistema
nervioso autónomo
Los primeros anatomistas se dieron cuenta
que la actividad visceral no se encontraba bajo el control voluntario. Los
nervios periféricos y ganglios asociados con las vísceras aparentemente eran
autónomos a la conciencia. El sistema nervioso autónomo incluye fibras motoras
y fibras sensoriales. Las fibras
sensoriales se encargan de monitorear las condiciones internas del cuerpo como
la presión sanguínea, la cantidad de oxígeno y dióxido de carbono, la
temperatura de los órganos y de la piel y la actividad de las vísceras. Las
fibras motoras por el contrario se especializan en la peristalsis de las
vísceras y del impulso cardíaco, así como de la contracción de las glándulas,
regulando al músculo liso y al músculo cardíaco.
Aun cuando son autónomos, algunos centros
de control consiente pueden tomar el control de algunas regiones viscerales,
como el ritmo respiratorio, e incluso el ritmo cardíaco, aunque algunos son más
fáciles de controlar que otros y se requiere de entrenamiento especial. Los
reflejos controlan las actividades manteniendo la homeóstasis. En su forma más
simple, un circuito neuronal del sistema nervioso autónomo incluye cuatro
neuronas unidas en un circuito cerrado. Sensorial → asociación → pregangliónica
→ motora postgangliónica. Ambas, motora y sensorial se encuentran unidas en el
tejido blanco, de esta forma cuando una manda el impulso motor, la otra lo
registra completando el circuito.
En los mamíferos el sistema nervioso
autónomo se divide en dos sistemas antagonistas, el simpático y el
parasimpático. El simpático se encarga de preparar el cuerpo para la lucha o el
escape mediante la emisión de hormonas como la adrenalina, los órganos son
sometidos a mucho estrés, pero esto permite realizar proezas físicas para
situaciones de vida o muerte. El parasimpático regula una homeóstasis de
relajación para actividades pasivas, su función es mantener el equilibrio del
cuerpo para que los órganos estén siempre en un estado de funcionamiento óptimo
y que no sufran daños, el principal neurotransmisor en este caso es la
acetilcolina.
En los mamíferos cada órgano posee una
inervación independiente del simpático y del parasimpático, excepciones a esta
doble conexión son la glándula adrenal, los vasos sanguíneos periféricos, las
glándulas sudoríparas, las cuales solo reciben inervación del simpático. La
división de simpático y parasimpático es razonable para los mamíferos, sin
embargo, en otros vertebrados la diferencia no es tan clara. Aunque
anatómicamente están dos tipos de inervación, funcionalmente no hay una
especialización. En consecuencia, la descripción del sistema nervioso
periférico de los no mamíferos se basa únicamente en criterios anatómicos, sin
mayor posibilidad de inferencia funcional.
15. El sistema nervioso central de los vertebrados
|| Regresar al Índice ||
La estructura general de la médula y de componentes accesorios los describiremos posteriormente cuando hablemos del sistema nervioso humano, ya que estructuralmente son semejantes, sin embargo, el cerebro es otra historia, comparativamente los cerebros de los vertebrados poseen diferencias que valen la pena resaltar. El sistema nervioso central es la principal zona de control de la información de los vertebrados, regulando el comportamiento a todos los niveles, ya sea para alimentarse, reproducirse, pelear o escapar, por lo que se encuentra conectado con todo, desde los sentidos hasta los músculos. La principal estructura de control es el cerebro.

Figura 15.1. El cerebro. El
cerebro humano y de otros mamíferos esta hecho por capas, las cuales son más
recientes mientras más hacia arriba están.
El cerebro se forma embrionariamente a
partir del tubo neuronal anterior “punta frontal de la proto médula”. Incluye tres regiones anatómicas, la primera es
el cerebro posterior o rombocéfalo. Morfogenéticamente, la especialización del
rombencéfalo durante la vida embrionaria da lugar a la formación de diversas
estructuras del sistema nervioso central en vertebrados. También se lo llama
cerebro primitivo posterior, siendo uno de los tres esbozos de cerebros
primitivos, junto al prosencéfalo o cerebro primitivo anterior y al mesencéfalo
o cerebro primitivo medio. Se encuentra localizado en la parte inmediatamente
superior de la médula espinal y está formado por tres estructuras: el bulbo, la
protuberancia anular o puente de Varolio, y el cerebelo. En él se encuentra,
también, el cuarto ventrículo.
Posteriormente se encuentra el cerebro medio o mesencéfalo que une el puente troncoencefálico o puente de Varolio y el cerebelo con el diencéfalo. Su eje longitudinal se inclina hacia atrás y los flagelos se alejan de la línea media en su ascenso por el foramen de Pacchioni para penetrar en el hemisferio cerebral correspondiente. Conduce impulsos motores desde la corteza cerebral hasta el puente troncoencefálico y conduce impulsos sensitivos desde la médula espinal hasta el tálamo. Los tubérculos cuadrigéminos superiores coordinan los movimientos de los globos oculares en respuesta a estímulos visuales y a otros estímulos, y los tubérculos cuadrigéminos inferiores coordinan los movimientos de la cabeza y tronco en respuesta a estímulos auditivos. Contiene el núcleo de origen para los nervios craneales III (nervio oculomotor) y IV (nervio patético). El mesencéfalo también se relaciona con algunos aspectos de la visión, la audición, el sueño y no vigilia.

Figura 15.2. Cerebro ancestral. Los
cerebros de los vertebrados primitivos son más lineales, en este caso el “Cerebrum”
es la región que en los mamíferos se hace masiva y llena de circunvoluciones
que le da su textura arrugada, pero en los peces es pequeña.
Finalmente encontramos en cerebro frontal,
cerebro superior o prosencéfalo. Esta región incluirá al telencéfalo y el
cerebrum así como el diencéfalo así como al tálamo. El prosencéfalo tiende a
hacerse muy grande en varios grupos de vertebrados, que son de hecho los
vertebrados tipos: mixinos, tiburones, peces óseos y tetrápodos. Mucho de este
crecimiento se correlaciona con el desarrollo evolutivo de órganos de los
sentidos más potentes, como el olfato. Acompañando el crecimiento de las
regiones sensoriales, también hay un aumento de tamaño de las regiones de
control, lo que permite un comportamiento más complejo. A medida que los
vertebrados evolucionaron sistemas de movimiento más complejo, otras zonas del
prosencéfalo aumentaron a su vez de tamaño.
15.1 Anatomía comparada de los cerebros
Con los cerebros sucede algo semejante que,
con los corazones, y es que tendemos a ponerlos en una secuencia lineal desde
el más simple hasta el más complejo, que sería el nuestro, sin embargo esta
representación no concuerda con la teoría sintética de la evolución moderna y
es más propia de los esquemas de la evolución lamarckiana o platónica, pues en
ambos modelos si existía el concepto de perfección teleológica (Figura 15.3).
Sin embargo, la historia es muchísimo más compleja, y debería esquematizarse en una metáfora relacionada al árbol de ramificación, pues es de este modo en que se enciente la evolución desde el siglo XIX, el hecho de que las representaciones lineales aun sigan siendo populares demuestra una vez más el modo en que la cultura popular confunde crónicamente dos interpretaciones opuestas de la naturaleza, la darwiniana y la lamarckiana.

Figura 15.3. Organización lineal. En
el proceso de cefalización, varias regiones se han hecho más grande en el curso
de la evolución de algunos grupos de mamíferos, especialmente el cerebro / forebrain y el cerebelo /hindbrain, pero hay que recordar que
especies concretas tendrán variaciones fuertes.
Evidentemente es un modelo simplificado con solo algunos grupos de vertebrados tipo que describimos clásicamente en los cursos de introducción a la anatomía de los vertebrados, pero igual es, más útil que el esquema lamarckiano lineal. En verde encontramos el neocórtex que es la estructura más reciente en especializarse a partir del prosencéfalo. En los mamíferos, y especialmente en la rama que da lugar a los primates el neocórtex es enorme y cubre las demás estructuras del cerebro, y en este modelo preciso el más grande de los cerebros representados es el del chimpancé, el nuestro es aún más grande, pero es la misma idea. En naranja encontramos al cerebelo, en verde oscuro el bulbo olfatorio que es muy pronunciado en varios grupos de vertebrados como los reptiles. Finalmente, en amarillo las estructuras del mesencéfalo que son el puente entre la médula y el cerebro.

Figura 15.4. Organización ramificada. En
la figura anterior tenemos la vista dorsal “de espalda” de los cerebros de
varios grupos de vertebrados representados en un modelo de ramificación
darwiniano.
15.2 La evolución de los cerebros
Al pensar en la evolución del cerebro de
los vertebrados, o en la evolución de cualquier estructura es útil considerar
que preguntas pueden ser realizadas a cerca de un problema concreto. En el caso
de la evolución de los vertebrados hay cuatro preguntas principales: que
cambios ocurrieron en el cerebro, cuando ocurrieron los cambios, como
ocurrieron los cambios y como se dieron estos cambios (Northcutt, 2001). Por cuestiones de extensión, solo nos enfocaremos en la primera
pregunta, cuáles fueron los cambios que ocurrieron en el cerebro, ya que es la
más directamente relacionable a la anatomía de manera introductoria, para una
discusión más extensa recomiendo leer el siguiente revisión de Northcutt (2002).
El problema con el cerebro es que es un
tejido blando que no se fosiliza con facilidad, sin embargo la carcasa de los
cráneos intactos permite realizar tomografías en las cuales se puede inferir
con cierto nivel de confianza la silueta tridimensional de un cerebro
desaparecido, aunque evidentemente esta técnica requiere una calibración de
cerebros modernos para asegurarse que los modelos que empleamos para crear los
cerebros desaparecidos se hagan sobre terreno firme de los ya conocidos (Falk et al., 2000; Marino, Uhen,
Pyenson, & Frohlich, 2003; Rogers, 1999).
Después más de 200 años de descripciones
anatómicas y más de 60 años de anatomía experimental y estudios fisiológicos,
existe una cantidad de datos considerables a cerca de los principales cambios
que han ocurrido al interior del cerebro de los vertebrados. Adicionalmente la
filogenética cladística ha emergido como una herramienta poderosa para analizar
la variabilidad de los caracteres cerebrales (Butler & Hodos, 2005; Kaas,
2016; Northcutt, 1984; Ubeda-Bañon et al., 2014).
Dado este enorme cuerpo conceptual se hace
necesario poder delinear tendencias a cerca de los cambios, los cuales se
enmarcan en tres grandes conclusiones: (1) todos los vertebrados exceptuando
los peces sin mandibular agnatos poseen la misma cantidad de subdivisiones
cerebrales, en los agnatos aparentemente el cerebelo se perdió o no había
evolucionado. (2) El tamaño cerebral, especialmente del cerebrum en su cortex y
mucho más en el neocortex, ha experimentado un aumento de tamaño independiente
en varios linajes de vertebrados. (3) El aumento de tamaño del cerebrum
conllevó al desarrollo de crestas neurales o circunvoluciones que aumentan la
superficie del tejido cerebral, lo cual ha conllevado a un aumento en la
complejidad comportamental, especialmente relacionado a los hábitos
depredatorios (Northcutt, 2002).
15.3 Especialización cerebral
Incluso un análisis superficial de la anatomía del cerebro de cada linaje principal de vertebrados revela que los vertebrados comparten el mismo número de subdivisiones del cerebro. La ausencia de un cerebelo en los mixinos y lampreas es la única excepción. Ambos grupos de peces que denominamos ciclóstomos “que puede ser un nombre monofilético o no, ha discusión en eso” poseen un cerebro fuertemente lineal, con un cerebrum pequeño (Northcutt, 2002).

Figura 15.5. Pez-gusano.
Reconstrucción del sistema nervioso central de Haikouella., un
fósil de la transición pez-gusano.
El hecho de que los principales grupos de
vertebrados posean una arquitectura conservada se puede explicar por dos
hipótesis, la primera es que esta arquitectura hubiera evolucionado
independientemente en cada linaje, y la segunda es que el ancestro común de los
vertebrados la hubiera fijado y heredado a sus descendientes. Los métodos de
máxima parsimonia favorecen la segunda interpretación (Northcutt, 2002), sin
embargo determinar la forma del cerebro del ancestro común de todos los
vertebrados puede ser complejo dado que el cerebro no se fosiliza. Sin embargo
el cráneo si puede fosilizarse, además se poseen fósiles de algunos fósiles
importantes como Yunnanozzon y Haikouella que ya describimos
generalmente en el capítulo del sistema esquelético (Donoghue & Keating, 2014).
De Haikouella se conocen cientos de
especímenes, lo cual ha facilitado la reconstrucción de su cuerpo, incluyendo
su sistema nervioso central por medio de tomografía computarizada (Northcutt, 2002). Haikouella
fue un pequeño animal semejante a un cefalocordado primitivo, aunque con menos
hendiduras branquiales. Su cerebro era grande para su tamaño u se encontraba
dividido en dos o tres secciones lineales. Existe evidencia de que poseía ojos
y en consecuencia el lóbulo visual debía estar presente, pero no hay evidencia
de otras estructuras sensoriales especializadas como las del olfato. Existe
controversia acerca de si Haikouella poseía hemisferios cerebrales, dado
que no hay rastros de bulbos olfatorios se pensaría que su cerebro era sólido,
pues la hipótesis imperante en estos momentos es que los hemisferios cerebrales
emergieron a partir de los bulbos olfatorios, siendo posteriormente rodeados
por el cerebrum “cortex + neocortex” en crecimiento.
Adicionalmente otros carneados primitivos,
han sido encontrados, como Haikouichthys y Myllokunmingia, en los
cuales las cabezas mejor preservadas han revelado la presencia de las capsulas
nasales y ócticas, revelando la presencia de órganos especializados del olfato
y del oído. Si estas observaciones se confirman es posible indicar que las
principales partes del cerebro de los vertebrados estaban presentes en
craneados verdaderamente primitivos (Hall & Gillis, 2013;
Rosslenbroich, 2014).
15.4 Tamaño
Visto a través de los principales linajes
de vertebrados, el tamaño del cerebro varía aproximadamente 30 veces o 3000%
con respecto al tamaño corporal, por lo que podemos tener cerebros grandes con
respecto al tamaño del cuerpo y cerebros pequeños con respecto al tamaño del cuerpo.
Esta relación entre el tamaño del cerebro y el tamaño del cuerpo es altamente
organizada. Los agnatos que son los peces más primitivos encontramos los
cerebros más pequeños. Los peces óseos poseen en general tamaños cerebrales más
grandes con algunas excepciones. Los condrictios poseen cerebros un poco más
grandes, con lóbulos olfatorios prominentes. En cada principal ramificación de
los linajes de vertebrados, los tamaños cerebrales aumentan paulatinamente, en
sucesión los peces de aleta lobulada, los tetrápodos, los anfibios primitivos y
los amniotas (Northcutt, 2002). Tanto las aves como los
mamíferos poseen cerebros que son entre 6 y 10 veces más grandes que los de un
reptil de tamaño semejante, mientras que los dinosaurios caen en medio con
mucha variación (Bhullar et al.,
2016; Burish, Kueh, & Wang, 2004; Hurlburt, Ridgely, & Witmer, 2013;
Jerison, 1969; Larsson, Sereno, & Wilson, 2000). Al
interior de los mamíferos, los primates, los cetáceos y los carnívoros poseen
los cerebros más desarrollados (Northcutt, 2002). La relación tamaño cerebral-tamaño corporal se encuentra
relacionado con un incremento en la complejidad del comportamiento, la
capacidad de elaborar planes y de cooperar de maneras complejas no instintivas.
16. Anatomía comparada del cerebro de los vertebrados
|| Regresar al Índice ||
16.1 Ciclostomos y grupos externos
Los agnatos que son los peces más primitivos
encontramos los cerebros más pequeños. En el modelo de la izquierda podemos
analizar tres tipos de cerebros, en (A) tenemos el crebero larvario de un
mixino, en (B) el cerebro larvario de una tunicado y en (C) el cerebro de una
lamprea adulta. De estos animales solo A y B son peces en toda regla además de
vertebrados, mientras que el tunicado es un grupo externo, si bien cercano a
los vertebrados. Si bien los tunicados son sésiles en sus formas adultas y se
asemejan más a una copa, sus larvas motiles se asemejan mucho a lo que debió
haber sido una forma general de los primeros pre-vertebrados, con una notocorda
una médula y un cerebro especializado en el procesamiento de información
motora. De todos ellos solo la lamprea representa una estructura tripartita del
cerebro de los vertebrados de forma muy clara, aunque algunos estudios revelan
que los mixinos poseen estructuras homólogas a los demás vertebrados. El
cerebro en cualquier caso es altamente lineal con un bulbo olfatorio claramente
diferenciado en la punta.
16.2 Los peces con mandíbula
Los peces óseos poseen en general tamaños
cerebrales más grandes que los ciclostomos con algunas excepciones. Los
condrictios poseen cerebros un poco más grandes especialmente en el cerebrum
"telencefalo", con lóbulos olfatorios prominentes, además de la
presencia de un cerebelo. En los modelos siguientes tenemos la comparación del
cerebro de un tiburón con respecto al de una lamprea de un tamaño corporal
semejante. Otro aspecto significativo es
que el telencefalo y el cerebrum poseen tamaños semejantes en estos animales,
lo cual revela que son principalmente instintivos, siendo que el telencéfalo al
tener un tamaño limitado solo sirve para procesar información motriz y
sensorial a cortos plazos de memoria (Figura 15.3).
16.3 Anfibios
El cerebro anfibio conserva la estructura heredada de los peces, pero con un telencéfalo o cerebrum más grande, lo cual se caracteriza por un mecanismo de desplazamiento más complejo, que no solo involucra el movimiento caudal, sino también el movimiento de los apéndices anteriores y posteriores.

Figura 16.1. Cerebro de rana.
16.4 Cerebro reptil
Aunque el telencéfalo "cerebrum"
sigue haciéndose más grande, la estructura general del cerebro sigue siendo la
misma, aunque la característica más predominante es la presencia de un enorme
bulbo olfatorio cuya estructura varía de especie en especie, aunque este tamaño
revela que los reptiles se especializan mucho en la recolección de información
olfatoria.

Figura 16.2. Cerebro de cocodrilo bebe.
16.5 Aves y dinosaurios
El cerebro de las aves se distancia un poco de la tendencia que hemos visto con un enorme telencéfalo, el cual evolucionó en paralelo con el de los mamíferos. Por muchos años los anatomistas estuvieron viciados con la visión lamarckiana-ortogenesista de la evolución, siendo incapaces de aceptar que este telencéfalo al igual que el de los mamíferos estaba enfocado en el análisis de información compleja comportamental, y por el contrario la asociaron a un comportamiento instintivo. En la actualidad tales vicios cognitivos se han dejado de lado por una tendencia más congruente con una visión de ramificación de la evolución, las aves no se colocan como un escalón inferior a los mamíferos, sino como un linaje propio con una historia evolutiva compleja en la cual se experimentó en paralelo el incremento del telencéfalo para el aumento de la inteligencia.

Figura 16.3. Cerebro de pollo o de chorlito. Los
cerebros de las aves son semejantes al de los mamíferos por un proceso de
evolución paralela, cuando el cerebrum o telencéfalo se agranda, se abulta
dorsalmente, alejándolo de una apariencia lineal.
En el caso de los dinosaurios, la tomografía computarizada ha dado acceso a la silueta de sus cerebros, que es de hecho lo que hemos analizado hasta el momento con todos los demás. Estos cerebros se aproximan mucho más al de las aves que al de los reptiles con una prominente región dedicada al telencéfalo, pero en algunas especies conservan rasgos asociables a los reptiles como prominentes bulbos olfatorios en la parte más frontal. De todos los dinosaurios los que presentan el mayor telencéfalo son los dinonicos, terópodos relacionados con las aves especializados en la caza.

Figura 16.4. Cerebro de los dinosaurios. La
tomografía computarizada ha permitido reconstruir la superficie de los cerebros
de animales extintos como los dinosaurios.
Contrario al mito popular, los dinosaurios
no poseían dos cerebros, lo que pasa es que en la región del sacro la médula
espinal tiende a expandirse en los grandes vertebrados para controlar los
apéndices, aunque si es cierto que en los saurópodos esta expansión es
particularmente grande, y también hay que resaltar que estos son los
vertebrados terrestres más grandes que han existido y sus descomunales músculos
necesitaban muchas neuronas motoras.
En cada principal ramificación de los
linajes de vertebrados, los tamaños cerebrales aumentan paulatinamente, en
sucesión los peces de aleta lobulada, los tetrápodos, los anfibios primitivos y
los amniotas (Northcutt, 2002). Tanto
las aves como los mamíferos poseen cerebros que son entre 6 y 10 veces más
grandes que los de un reptil de tamaño semejante, mientras que los dinosaurios
caen en medio con mucha variación (Bhullar et al., 2016; Burish, Kueh, &
Wang, 2004; Hurlburt, Ridgely, & Witmer, 2013; Jerison, 1969; Larsson,
Sereno, & Wilson, 2000). Al interior de los mamíferos, los primates, los
cetáceos y los carnívoros poseen los cerebros más desarrollados (Northcutt,
2002).
16.6 Mamíferos
La evolución del cerebro de los mamíferos se ha estudiado con demasiada frecuencia con la suposición implícita de que todos los cerebros de los mamíferos se hicieron de acuerdo con las mismas reglas de escala. La relación entre el tamaño de las estructuras cerebrales y su número de neuronas varía tanto entre los órdenes y las estructuras de los mamíferos.

Figura 16.5. El cerebro de los mamíferos vivos.
El gran tamaño del cerebro ha evolucionado
varias veces independientemente de un antepasado común que tenía un cerebro
pequeño. Por lo tanto, no hay ninguna razón a priori
para suponer que todos los cerebros que se han vuelto grandes lo hicieron a
través de los mismos mecanismos en la evolución (Herculano-Houzel, 2012).
16.7 Consecuencias del aumento de tamaño
La relación tamaño cerebral-tamaño
corporal se encuentra relacionado con un incremento en la complejidad del
comportamiento, la capacidad de elaborar planes y de cooperar de maneras
complejas, sin embargo, no debejos dejarnos llevar por el orgullo. Los neurocientíficos
se han acostumbrado a una serie de "hechos" sobre el cerebro humano:
tiene 100 mil millones de neuronas y 10 a 50 veces más células gliales; es el
más grande de lo esperado para su cuerpo entre primates y mamíferos en general,
y por lo tanto, el más capaz cognitivamente; consume un destacado 20% del
presupuesto total de energía corporal a pesar de representar solo el 2% de la
masa corporal debido a una mayor necesidad metabólica de sus neuronas; y está
dotado de una corteza cerebral sobredesarrollada, la más grande en comparación
con el tamaño del cerebro (Herculano-Houzel, 2012).
Estos hechos llevaron a la noción
generalizada de que el cerebro humano es literalmente extraordinario: un ente
atípico entre cerebros de mamíferos, desafiando las reglas evolutivas que se
aplican a otras especies, con una singularidad aparentemente necesaria para
justificar las capacidades cognitivas superiores de los humanos sobre los
mamíferos con cerebros aún más grandes. Sin embargo, estos hechos, con
profundas implicaciones para la neurofisiología y la biología evolutiva, no se
basan en pruebas sólidas o suposiciones sólidas. Herculano-Houzel (2012) desarrolló de un método que permite una cuantificación rápida y
confiable del número de células que componen todo el cerebro, que ha
proporcionado un medio para verificar estos hechos.
A pesar de nuestros continuos esfuerzos
por comprender la biología a la luz de la evolución, a menudo hemos recurrido a
considerar el cerebro humano como un ente atípico para justificar nuestras
habilidades cognitivas, como si la evolución se aplicara a todas las especies,
excepto a los humanos. Sorprendentemente, todas las características que
parecían distinguir al cerebro humano como extraordinario, un punto fuera de la
curva, ahora se pueden entender, en retrospectiva, como resultado de las
comparaciones contra el tamaño del cuerpo con las suposiciones subyacentes de
que todos los cerebros están uniformemente ampliados o reducidos entre sí y que
el tamaño del cerebro (y, por lo tanto, el número de neuronas) está
estrechamente relacionado con el tamaño del cuerpo (Herculano-Houzel, 2012).
Herculano-Houzel
(2012) muestra datos cuantitativos recientemente
adquiridos sobre la composición celular del cerebro humano y su comparación con
otros cerebros, tanto primates como no primarios, los cuales indican
fuertemente que necesitamos repensar el lugar que ocupa el cerebro humano en la
naturaleza y la evolución, y reescribir algunos conceptos básicos. que se
enseñan en los libros de texto.
El cerebro humano tiene la cantidad de
neuronas y células no neuronales que se esperaría para un cerebro de primates
de su tamaño, con la misma distribución de neuronas entre su corteza cerebral y
cerebelo que en otras especies, a pesar del aumento relativo de las primeras;
cuesta tanta energía como se esperaría de su número de neuronas; y puede haber
sido un cambio de una dieta cruda a una dieta cocinada que nos proporcionó su
notable número de neuronas, posiblemente responsables de sus notables
habilidades cognitivas.
17. Introducción al sistema nervioso central
|| Regresar al Índice ||
Para este punto
ya hemos examinado la neurona como célula y tejido, así como sus especializaciones
moleculares, las cuales le permiten realizar funciones muy precisas de
comunicación y transferencia de estímulos a grandes velocidades. Al observar el
sistema nervioso de forma separada al resto del cuerpo se observa una densa
masa de tejido nervioso donde debería estar la cabeza, con un denso cordón
neural que se extiende a través del dorso junto donde la línea de la espalda
debería estar. Estas estructuras son el cerebro y la médula espinal
respectivamente, y constituyen el así llamado Sistema Nervioso Central, el cual integra y coordina la mayoría, si
no es que todas, las funciones voluntarias e involuntarias del sistema
nervioso.

Figura 17.1. Sistema nervioso humano.
incluye no solo los sistemas central y periférico, también incluyen los órganos
de los sentidos, los cuales se encuentran directamente vinculados a los nervios
y en algunos casos como los ojos, emplean las mismas estructuras bioquímicas
como las proteínas G ligadas a receptores transmembranales de siete dominios.
Conectados al cerebro y a médula espinal existe una
amplia cantidad de bifurcaciones que se desprenden como cables, los
denominaremos nervios. Los nervios transmiten mensajes desde y hacia el Sistema
Nervioso Central. Los nervios forman una vasta red, en la cual las
diferentes subramificaciones crean un sistema de micronervios que se incrusta
en los tejidos del cuerpo, estos micronervios generalmente no son representados
en los modelos “igual que los capilares en el sistema sanguíneo” pero están
allí y son la parte más importante del sistema nervioso ya que están en
contacto literal con las superficies a ser controladas o de las que se registra
información.
En ocasiones
los nerviosos se unen para formar masas llamadas ganglios. Los nervios y
ganglios se encuentran localizados fuera del Sistema Nervioso Central se
denominan el Sistema Nervioso Periférico.
El Sistema Nervioso Periférico puede ser dividido en
el Sistema Nervioso Somático y el Sistema Nervioso Autónomo.
El Sistema Nervioso Somático consiste en nervios que
transportan información desde el Sistema Nervioso Central creando
la sensación de movimientos voluntarios. El Sistema Nervioso Autónomo
como su nombre lo dice envía información que mantiene bajo control la
peristalsis autónoma de diferentes órganos como el canal gastrointestinal, las
venas gruesas o el corazón. El Sistema Nervioso Autónomo a
su vez posee dos partes que generalmente producen efectos diferentes en los
músculos o glándulas que controlan. Uno es el Sistema Nervioso Simpático que
se encarga de las situaciones estresantes o de emergencia. El otro es el Sistema
Nervioso Parasimpático que ajusta las funciones corporales de
forma tal que la energía se reserve lo más posible en condiciones no
estresantes. Recuerde sin embargo que la clasificación obedece a un modelo
humano se compartimentalizar la información para poder entenderla, sin embargo,
todos los sistemas nerviosos funcionan como una unidad anatómica y fisiológica.

Figura 17.2. Las meninges. Las
meninges son capas con textura de cuero (A). En la figura superior central
podemos apreciar de derecha a izquierda, cuero cabelludo, periostio, hueso
craneal, dura madre, aracnoides, piamadre y tejido nervioso cerebral (B). En la
figura inferior derecha podemos ver las capas de tejido que aíslan al cerebro
del ambiente externo (D).
El SNC “Sistema
Nervioso Central” incluye el cerebro y la médula espinal, los cuales están
hechos de tejido neuronal densamente trenzado. Las neuronas en si son bastante
frágiles, y en la práctica se encuentra en una fase S “síntesis” cuasiperpetua
por lo que es muy raro que ingresen en fase M “mitosis” y las pocas que lo
hacen no son capaces de regenerar tejido perdido, en un sentido práctico el
tejido neuronal no se regenera. El cerebro y la médula espinal se encuentran
protegidos por capas de hueso “el neurocráneo y la columna vertebral”, un
sistema especializado de membranas llamadas meninges y un fluido elástico
denominado fluido cerebroespinal.
17.1 Las meninges
Las meninges son
tres cubiertas protectoras del cerebro y la médula espinal que reciben los
nombres de: duramadre, aracnoides y piamadre.
La capa más
externa es la dura madre “Figura
17.2”, es una capa dura y de una textura
semejante al cuero. Más allá de la dura madre está la aracnoides “imagen
siguiente”, se la denomina así porque es semejante a una seda de telaraña, la
cual consiste en extensiones delgadas semejantes a una seda de araña. La capa
más interna es la piamadre, esta se encuentra moldeada alrededor del cerebro
como si se tratara de un leotardo ajustado que adquiere la forma de la
superficie de todas las circunvoluciones del cerebro.
La inflamación de
las meninges se denomina meningitis. La meningitis puede llegar a ser una
condición patológica bastante peligrosa, ya que puede presionar el cerebro o
transmitir la causa subyacente “generalmente una infección bacteriana o viral”
al tejido nervioso conllevando a la encefalitis. La presión sumada de ambos
tejidos puede provocar derrames sanguíneos, condición generalmente fatal.
Muchos tipos de bacterias y algunos virus pueden causar meningitis. Si las
bacterias son la causa la persona debe ser tratada con antibióticos. Si la causa
es un virus la cosa es más compleja, no existen medicamentos que realmente
afecten a los virus más allá de las vacunas o los costosos interferones, por lo
que básicamente se depende del sistema inmune mientras se tratan los síntomas
tratando de evitar una mayor inflamación por causa de la fiebre.
17.2 El fluido cerebroespinal
El fluido cerebro
espinal "cefalorraquídeo" llena los espacios entre las meninges y el
tejido nervioso llamados ventrículos en el cerebro y canal central en la
columna. El fluido es segregado en los ventrículos y fluye hacia el canal
central para eventualmente se reabsorbido hacia la sangre. Las funciones del
fluido cerebroespinal son:
👉 Absorción de impactos: se trata de un fluido amortiguante que absorbe la
energía de un impacto, impidiendo que los traumas neuronales sean mayores.
👉 Soporte: al
igual que un esqueleto hidrostático el fluido le permite al cerebro mantener su
forma sin aplastarse en su propio peso gracias a una fuerza opuesta a la
gravedad llamada empuje hidrostático, esto provoca que el peso efectivo del
cerebro sea menor aun cuando posea la misma masa.
👉 Remoción de los desechos de neurotransmisores y alimentación de iones salinos: El fluido proporciona fluidos con nutrientes y mensajeros químicos, y remueve los desechados cuando no son degradados completamente. Aunque el cerebro es protegido por el cráneo, las meninges y el fluido cerebroespinal las heridas cerebrales son uno de los traumas más comunes con 7 millones de caso solo en los Estados Unidos de América.

Figura 17.3. Punción lumbar. La
punción lumbar es uno de los procesos más dolorosos que existen, por el riesgo
de lesiones en el sistema nervioso central.
El fluido
cefalorraquídeo es normalmente estéril, y su obtención es un método de
diagnóstico extremo debido a que no puede emplearse anestesia. Si en un cultivo
de fluido cefalorraquídeo se detectan bacterias, implica una peligrosa
condición, pues las bacterias que crucen la barrera hematoencefálica pueden
ocasionar graves lesiones o la muerte rápidamente.
17.3 La barrera hematoencefálica
La barrera hematoencefálica es la razón por la que el líquido cefalorraquídeo es estéril normalmente. Se trata de un mecanismo que selecciona sustancias para que puedan ingresar al fluido cefalorraquídeo desde la sangre. Esta barrera es formada por la unión íntima entre las células de los capilares sanguíneos que soportan el cerebro y la médula espinal.

Figura 17.4. La barrera hematoencefálica.
Compuesta por células gliales que aíslan los vasos sanguíneos de la matriz
extracelular neuronal, media el flujo de sustancias, e impide el paso de muchos
tipos de células, como varios tipos de bacterias y a la mayoría del sistema
inmune, debido a su capacidad inflamatoria, si el cerebro se inflama, es el
fin.
Estos capilares
especializados generan una estructura diferente de los capilares sanguíneos que
los capilares normales, las sustancias son forzadas a pasar por varias capas de
células que se encargan de realizar un filtro. En este sentido la barrera hematoencefálica
no se la debe imaginar como un muro, sino como una capa que recubre todo el
sistema circulatorio que penetra en el sistema nervioso.
La barrera
hematoencefálica es una microestructura formada por la relación entre los
astrocitos y los capilares sanguíneos del cerebro. La idea es análoga a las
capas de mielina, una célula glial envuelve con proyecciones al capilar como si
fuera un nervio y en lugar de llamarlo mielina lo llamamos barrera
hematoencefálica. Sin embargo, la membrana hematoencefálica está hecha con
membranas biológicas y en consecuencia se encuentra sujeta a las mismas
limitaciones, por ejemplo, es incapaz de impedir el paso a sustancias solubles
en lípidos como los gases metabólicos como el oxígeno y el dióxido de carbono,
así como a moléculas liposolubles como los canabinoides o la cafeína. Al mismo
tiempo, la barrera hematoencefálica puede llegar a ser bastante frustrante ya
que impide la llegada al cerebro de medicamentos contra infecciones o el cáncer
que no sean solubles en lípidos.
18. El cerebro humano
|| Regresar al Índice ||
El cerebro es una estructura compleja, de hecho, es el órgano más complejo en el ser humano, y tal vez el único cuya biología aún guarda secretos serios para su comprensión completa. El cerebro humano es el producto de millones de años de evolución.

Figura 18.1. Principales regiones del cerebro.
Disección del encéfalo; (YouTube)
(YouTube) (YouTube) (YouTube) (YouTube).
Al igual que un
servidor masivo, el proceso evolutivo del cerebro se ha realizado en paralelo a
su funcionamiento, por lo que las partes más viejas y los “software” más viejos
jamás fueron borrados o reemplazados realmente, simplemente han sido tapados
con parches estructurales y de programación más complejos, para funciones más
concretas. A pesar de esto, las partes más antiguas del cerebro son con
frecuencia más relevantes a la hora de imponer un curso de acción, ya que allí
se encuentran respuestas instintivas a estímulos de supervivencia como el
miedo, el deseo sexual o el hambre. En general podemos dividir al cerebro en
tres partes, que en su orden evolutivo son: el tallo cerebral, el cerebelo y el
encéfalo.
18.1 Encéfalo
En encéfalo es la
parte más grande de todo el cerebro, y también es la más reciente en la
historia evolutiva, representando el 83% de la masa cerebral. El encéfalo agrupa
las regiones responsables del pensamiento complejo, siendo responsable de la
generación de la mayor parte de las características que describimos como
humanas.
Las muchas
circunvoluciones de la superficie del encéfalo lo hacen ver arrugado. Algunos
pliegues son más profundos que otros. La más profunda se encuentra en el centro
y se proyecta de forma ventral separando al encéfalo en dos hemisferios. Estructuralmente
el encéfalo resguarda lo que comúnmente se llama materia gris y materia blanca.
La materia gris es denominada formalmente como córtex, y también es la que
posee mayor superficie, siendo el centro del pensamiento racional.
18.2 La materia gris y la materia blanca
La delgada membrana externa de cada hemisferio se denomina córtex cerebral.

Figura 18.2. Materia gris y materia blanca. El
color se debe a una diferencia en la cantidad de grasa, provocada por las capas
de mielina. Las capas de mielina se emplean para comunicar a largas distancias.
El córtex
consiste en billones de células gliales, neuronas y axones no mielinizados
descritos colectivamente como la materia gris. Aunque el córtex cerebral tiene
un espesor de unos 2.5 milímetros, se encuentra altamente plegado
sobre sí mismo, estos pliegues o circunvoluciones triplican el área de
superficie.
Más allá del córtex se encuentra la materia blanca, su color se debe a que los axones neuronales se encuentran altamente mielinizados. Los axones de la materia blanca les permiten a varias regiones del cerebro para comunicarse unas con otras y con la médula espinal. Una banda muy importante de la materia blanca se denomina cuerpo calloso. El cuerpo calloso conecta los dos hemisferios de forma tal que se pueden comunicar entre sí. Los pliegues más profundos permiten definir los cuatro lóbulos de cada hemisferio: frontal, parietal, temporal y occipital. Cada uno de estos lóbulos posee sus propias especializaciones, sin embargo, la asignación de una función única a cada lóbulo es bastante imprecisa. A pesar de esto parece ser que existen al menos tres áreas especializadas: sensorial, motora y de asociación.

Figura 18.3. El sentido del tacto no es uniforme. Las
regiones sensoriales se encuentra detrás de los lóbulos frontales, siendo una
especie de arco o cinta que se distribuyen de manera no homogénea la superficie
del cuerpo, algunas regiones tienen más sensores que otras (YouTube) (YouTube).
18.3 Áreas sensoriales
Nuestra
conciencia de las sensaciones depende de las áreas sensoriales del córtex
cerebral. Los receptores sensoriales ubicados en los órganos de los sentidos
envían pulsos por medio de potenciales de acción hacia el cerebro. Las áreas
para el oído, el olfato y parte del gusto se encuentran en el lóbulo temporal,
para la vista en el lóbulo occipital, otras partes del gusto son registradas en
el lóbulo parietal. El sentido más complejo es el tacto, ya que involucra
receptores de presión, dolor y temperatura, y esta información es enviada al
área primaria somatosensorial. Esta región forma una banda en los lóbulos
parietales que se extiende desde la posición de la oreja derecha hasta su
opuesta. Las sensaciones de diferentes partes del cuerpo son representadas por
diferentes regiones del área principal somatosensorial “verde en el modelo
anterior”. El grado de sensibilidad es directamente proporcional a la
superficie del córtex empleada para su análisis. En consecuencia, las regiones
más sensibles como la lengua, las manos, el rostro y los genitales poseen más
superficie del córtex dedicadas a ellas que otras áreas que en comparación son
más grandes como la espalda o los brazos.
18.4 Áreas
motoras
El movimiento se caracteriza por estar descentralizado en el cerebro, con áreas dedicadas tanto en la corteza del encéfalo como en el cerebelo. En este artículo solo hablaremos de las regiones del encéfalo dedicadas al movimiento. El área motora primaria se dedica a realizar movimientos altamente voluntarios, como emprender la caminata en una dirección determinada. El área motora primaria se encuentra organizada de forma similar al área somatosensorial, es como una cinta que va de un lado al otro del cerebro, y del mismo modo, el nivel de control del cuerpo depende de la superficie neuronal del córtex. Las regiones más predominantes son las dedicadas al rostro, el habla y las manos.

Figura 18.4.
La corteza motora está detrás de la sensorial.
Justo en frente
de la cotiza motora se encuentra el córtex premotor. Este coordina el
aprendizaje de habilidades que se entrenan en patrones repetitivos como la
ejecución de un instrumento musical, sin embargo, el cerebelo también se
encuentra involucrado en estos procesos. Los jugadores de futbol profesionales
se caracterizan por entrenar patrones de movimiento que les otorgan control
sobre el balón bajo situaciones de alta presión, esto les permite ejecutar
movimientos sin pensar en ellos de acuerdo a la información cognitiva
proveniente del cuerpo de aliados y oponentes en el campo de juego. A mayor
entrenamiento mayor fluidez en los movimientos.
18.5 Áreas de asociación
Posterior a cada una de las áreas somatosensoriales primarias se encuentra la región de asociación. Esta se comunica con las regiones sensoriales y motoras, así como con otras partes del cerebro, para analizar y actuar sobre la información sensorial. En particular, cada área de asociación se comunica con un área general de interpretación para reconocer cuales son los receptores sensoriales involucrados en una señal concreta. La región de interpretación general asigna un significado a la información sensorial mediante la integración de influjo desde las regiones sensoriales junto con información memorizada desde los sentidos por medio de experiencias previas.

Figura 18.5. Áreas de asociación. Las
regiones de asociación permiten vincular información que emerge de varias
regiones del cerebro. Son región es de análisis y control, de las cuales la más
famosa es la prefrontal, donde se almacena el yo personal de cada individuo.
Este sistema de análisis es holístico y permite al individuo estar alerta a señales hostiles del medioambiente, pero en el mundo moderno con frecuencia puede derivar en generalizaciones emocionales sobre fenómenos concretos. Una vez que la información sensorial ha sido interpretada por esta primera área conectada a las emociones, es enviada a la zona de asociación más compleja del cerebro, la corteza prefrontal. Esta es la región más anterior del cerebro “la frente”, es la base del pensamiento lógico y los razonamientos elaborados. Una de sus habilidades principales es la de detectar patrones en variables desligadas aun cuando estos no se encuentren allí realmente, tal como sucede en la pareidolia. La corteza prefrontal también involucra gran parte de lo que hace a una tal cual es ya que determina la toma de decisiones lógicas y gran parte de la personalidad.

Figura 18.6. Puro instinto. Esta
región límbica está compuesta por el hipotálamo, el tálamo, la amígdala, y el
hipocampo, todas regiones que controlan los instintos de supervivencia, como el
miedo, la ira, o el impulso sexual (YouTube)
(YouTube) (YouTube) (YouTube).
18.6 El tálamo
El tálamo era uno
de los nombres para las camas nupciales de los griegos, donde hombre y mujer se
ubicaban juntos para unirse. De consecuencia el tálamo cerebral es el
receptáculo que sostiene los dos hemisferios del encéfalo. Dejando las
analogías poéticas de lado el tálamo es un portal maestro de la información que
ingresa y sale de la corteza cerebral debido a que todos los mensajes enviados
a la corteza deben pasar obligatoriamente a través del tálamo.
El tálamo se
encuentra involucrado en la experiencia sensorial, la actividad motora, la
estimulación de la corteza cerebral y la memoria. La información proveniente de
los sentidos es enviada al tálamo. El tálamo organiza y filtra la información
de acuerdo a la función y la reenvía a las regiones apropiadas de la corteza
para su procesamiento. Algunas regiones del tálamo también integran la
información de diferentes fuentes tan solo con transmitirla. El tálamo procesa
información básica como las sensaciones placenteras y no placenteras como el
dolor, sin embargo, el cerebro no sabrá de donde provienen esas sensaciones
hasta que la información alcance la corteza.
18.7 Hipotálamo
Literalmente
significa “por debajo del tálamo”, se trata de una región pequeña del cerebro
que es responsable por el mantenimiento de la homeostasis corporal, como el
mantenimiento de los niveles de estabilidad de la matriz extracelular de
diferentes tejidos a lo largo del cuerpo. El hipotálamo es una especie de
director de orquesta, pequeño pero poderoso, sus efectos se ejercen sobre los
dos principales sistemas de control en el cuerpo, el sistema nervioso de forma
inmediata y el sistema endocrino de forma más profunda y permanente. De forma
más concreta, del hipotálamo dependen la presión sanguínea, el latido cardíaco,
la actividad digestiva, la tasa de respiración, el hambre, el miedo, la ira y
el desarrollo sexual.
18.8 El cerebelo
El cerebelo es una parte del cerebro encargada de procesar información sensorial y motora, siendo en tal caso redundante, o, mejor dicho, complementada por la corteza motora. Mientras que la corteza es consiente el cerebelo es inconsciente. El cerebelo actúa como un piloto automático que produce movimientos suaves, delicados, y bien coordinados que incluyen el equilibrio y la postura. En cierto sentido almacena el producto de entrenamientos sistemáticos, de forma tal que, aunque la corteza sensorial decide que hay que moverse, es el cerebelo quien determina como es que hay que moverse.

Figura 18.7. El cerebelo. El
cerebelo es importante en la memorización y experiencia de movimientos finos
como escribir o jugar bien al futbol, se dice que una habilidad se aprende
cuando se ejecuta sin pensarla, pues está inscrita en el cerebelo. La diferencia
radica tanto en la finura como en a velocidad y es lo que diferencia a un
profesional de un aficionado.
El cerebelo está
conectado a los sensores de presión “tacto”, dolor “tacto”, gravedad “en el
oído”, posición “ojos” de forma tal que puede determinar la postura del cuerpo
con respecto al ambiente en un momento determinado. El cerebelo se encarga de
dos funciones básicas, la corrección motora durante el entrenamiento y la
predicción motora. La primera función es básicamente recordar los errores de las
repeticiones de una destreza concreta para que el movimiento se almacene
permanentemente de forma las fluida e inconsciente. El cerebelo también posee
una función análoga a la de una computadora de análisis cinético, prediciendo
la posición de las partes del cuerpo durante el movimiento. Ambos aspectos del
cerebelo involucran la experiencia y el entrenamiento.
18.9 El tallo cerebral
El tallo cerebral
consiste en la médula oblongata, el mesencéfalo, y el puente troncoencefálico.
La médula oblongata es llamada simplemente médula, posee funciones semejantes a
las del hipotálamo como controlar el ritmo cardíaco así como la fuerza de su
contracción, el ritmo respiratorio así como su sincronización con el ritmo
cardiaco, y la presión sanguínea, todo conectado con las demandas metabólicas
de los músculos. La médula oblongata es el punto que conecta a la médula
espinal con el cerebro propiamente dicho, por lo tanto, toda la información
sensorial de ida y vuelta debe atravesarla en algún momento.
El mesencéfalo
procesa información acerca de reflejos simples a estímulos visuales y
auditivos. Muy probablemente representa las primeras unidades de procesamiento
de información sensorial conectada a respuestas instantáneas de combate o
escape. El troncoencéfalo representa un puente las porciones inferiores del
Sistema Nervioso Central con las estructuras cerebrales superiores. Más
específicamente conecta la médula espinal y el cerebelo con el encéfalo, el
tálamo y el hipotálamo. Adicionalmente el troncoencéfalo posee una región que
apoya la médula en regular la respiración.
18.10 El sistema límbico
El sistema
límbico es un término colectivo para estructuras que controlan las emociones y
la memoria. El sistema límbico se define en base a su funcionamiento y no en
base a su localización física, incluyendo muchas partes del cerebro
interconectadas. En resumen, el sistema límbico es el cerebro de las emociones,
mientras que los lóbulos frontales del encéfalo es el cerebro lógico. El
sistema límbico es la base para emociones como la ira, el dolor, el miedo, la
tristeza, la alegría y el placer sexual.
Las emociones son
importantes debido a que son un factor motivante que incrementa las
posibilidades de supervivencia cuando están conectadas a conductas que mejoran
la aptitud darwiniana. El miedo por ejemplo sirve para que el cuerpo se prepare
para el combate o el escape contra fenómenos ambientales potencialmente
fatales.
Las conexiones
entre el encéfalo y el sistema límbico permiten conectar emociones a los
pensamientos. Esto le permite al cerebro sentirse emocionado ante posibilidades
futuras y por lo tanto estimula la planeación lógica para recibir estímulos
placenteros. Pero esta conexión no es equitativa, sin el adecuado entrenamiento
el cerebro emocional puede fácilmente subyugar al cerebro lógico provocando
comportamientos que pueden ser poco pertinentes. El sistema límbico está
compuesto por el hipotálamo, la médula oblongata, neuronas del sistema nervioso
periférico del canal gastrointestinal entre otras. Conectado con las emociones
se encuentran las memorias, de hecho, las memorias generalmente requieren de
una emoción para ser recordadas con mayor fuerza, mientras que otros recuerdos
se pierden de forma rápida.
Esta sensación
intuitiva se corresponde con la clasificación de la memoria, ya sea a corto
plazo o a largo plazo. La memoria a corto plazo solo puede mantener pequeñas
cantidades de información por unos pocos segundos o minutos. La memoria a largo
plazo puede almacenar aparentemente cantidades ilimitadas de información por
horas, días o años. Aparentemente la memoria a largo plazo se relaciona con la
actividad social ya que se requieren emociones conectadas a los recuerdos para
que estos se consoliden con mayor fuerza.
No todos los
recuerdos a corto plazo son almacenados en la memoria a largo plazo, pero
cuando esto sucede el hipocampo juega un rol esencial, así como la amígdala. De
hecho, la amígdala también está conectada a diferentes órganos de los sentidos
como el olfato, de forma tal que los olores “o sabores complejos” pueden
desencadenar recuerdos vinculados a emociones fuertes.
18.11 Sistema de activación reticular
El sistema de activación reticular es una extensa red neuronal que corre desde la médula hasta la corteza cerebral, y funciona como un filtro de la información proveniente de los órganos de los sentidos.

Figura 18.8. Filtros cognitivos. El
sistema reticular filtra la información para no quedar bloqueados por tantas
señales sensoriales, algunos síntomas del espectro autista indican que el
sistema reticular a veces no funciona adecuadamente.
Nuestro cerebro
se encuentra constantemente inundado con información sensorial, esta
información puede sobrecargar al cerebro si no se hace un filtro de la
información relevante del ruido ambiental. Evidentemente al ser un sistema
holista tiende a pasar por alto muchas cosas que parecen ruido pero que son
importantes, haciendo al cerebro muy vulnerable a los juegos mentales. El
sistema de activación reticular es quien mantiene alerta a la corteza creando
el estado de vigilia, a menos que se encuentre inhibido por otras regiones
cerebrales. La vigilia o estado de conciencia estándar solo ocurre cuando el
sistema de activación reticular se encuentra activo, de lo contrario el estado
de conciencia está alterado como en el sueño. La conexión entre los estímulos
sensoriales y la actividad del sistema de activación reticular explica porque
es difícil conciliar el sueño en un ambiente con muchos estímulos sensoriales
como una terminal aérea con mucha luz, ruido y olores.
18.12 La médula espinal
El segundo componente de importancia para el sistema nervioso central y al mismo tiempo uno de los más antiguos es la médula espinal. La médula es un tubo de tejido neuronal trenzado que discurre de forma continua con la médula de las vértebras a lo largo de la columna vertebral. En los humanos en promedio se extiende por unos 45 centímetros. El diámetro de este tubo trenzado es en promedio del de un dedo meñique, aunque se hace un poco más gruesa en dos regiones: justo por debajo del cuello y al final debido a la aglomeración de una gran cantidad de bifurcaciones hacia las extremidades superiores e inferiores. Al interior de las vértebras la médula se encuentra protegida por las meninges, y por lo tanto se encuentra aislada del ambiente general del cuerpo como si se tratara de una bolsa hermética y resistente llena del fluido cerebroespinal.

Figura 18.9. La médula.
Estructura de un corte de médula espinal (YouTube)
(YouTube).
Los nervios
principales se bifurcan en pares, siendo la primera etapa del sistema nervioso
periférico. La médula espinal tiene dos funcionas primordiales: (1) transmisión
de señales entre el cerebro y los receptores neuronales en todo el cuerpo en
ambas direcciones y (2) centro reflejo que crea efectos somáticos de forma
independiente al cerebro. La transmisión de las señales es realizada
principalmente por la materia blanca de la médula que se encuentra hacia el
centro de la masa neuronal. Recordamos que su color blanco se debe a la mielina
que recubre los axones para que los potenciales de acción se transmitan a mayor
velocidad.
La médula también
posee materia gris, estas neuronas poco mielinizados son capaces de tomar
decisiones de forma independiente al cerebro recortando tiempo entre el
estímulo y la respuesta, este circuito independiente es importante cuando el
cuerpo debe responder a estímulos peligrosos que solo dejan fracciones de
segundo para tomar una decisión sensata.
19. El sistema nervioso periférico humano
|| Regresar al Índice ||
El sistema
nervioso periférico consiste en los nervios que se proyectan fuera de la médula
y sus subdicvisiones, así como los nervios craneales. El cuerpo posee 31 pares
de nervios espinales, cada uno de los cuales se origina en la médula espinal y
controla una región específica del cuerpo. Cada miembro de la pareja controla a
uno de los lados del cuerpo. Los lugares donde los nervios se fusionan
se denominan ganglios, son puntos sinápticos entre dendritas y cuerpos de
neuronas que unifican las señales actuando como microcerebros.

Figura 19.1. El sistema nervioso periférico espinal.
Nervios periféricos espinales conectados a la médula. Recuerde que hay una
segunda inervación al cuerpo mediada por el nervio vago que no pasa por la
médula.
Todos los nervios
espinales poseen fibras nerviosas motoras y sensoriales. Los nervios no están
compuestos únicamente por axones, poseen neuronas completas, pero su
organización tiene una serie de patrones. Los cuerpos celulares de las neuronas
sensoriales se ubican en regiones entretejidas del sistema nervioso denominadas
ganglios, al igual que ocurre en los artrópodos, cada segmento nervioso posee
una pareja de ganglios que se alimentan de nervios de la médula denominados
raíces dorsales, las raíces se trenzan en los ganglios y de estos emergen los
dos nervios principales a cada lado de un segmento.
Por otra parte,
los cuerpos de las neuronas de los nervios motores se ubican generalmente al
interior de la médula espinal al interior de la materia gris. Ambos nervios
emergen como una pareja en cada lado de la columna siendo denominados par
dorsal y par ventral. En ese sentido los nervios espinales salen como tétradas,
sin embargo, poco después se fusionan en el ganglio sensorial, y desde allí
vuelven a separarse, por lo que hay una clara diferenciación entre el sistema
nervioso periférico motor y el sistema nervioso periférico sensorial.
Existen 12 pares
de nervios craneales que emergen directamente del cerebro y controlan las
estructuras de la cabeza y ciertas partes del cuerpo incluyendo el corazón y el
diafragma. Algunos nervios craneales portan únicamente fibras sensoriales otros
llevan únicamente fibras motoras y otras portan ambas mezcladas.
El sistema nervioso
periférico se divide en el sistema nervioso autónomo y el sistema nervioso
somático. El sistema nervioso somático lleva mensajes sensoriales desde los
órganos de los sentidos distribuidos en todo el cuerpo, como la vista el sonido
o el tacto. El sistema nervioso somático también controla nuestros movimientos
voluntarios, que nos permite movernos e interactuar con nuestro medio ambiente.
El sistema nervioso autónomo se encarga de vigilar la homeostasis del cuerpo.
Estos nervios tienen control sobre las glándulas del sistema endocrino en caso
de que no exista un neurotransmisor con función endocrina liberado desde el
cerebro, también regulan la contracción de los músculos de tipo liso de los
órganos internos. De este modo el sistema nervioso autónomo ejerce control
sobre los órganos, ajustando su funcionamiento para mantener el equilibrio
dinámico interno.
El sistema
nervioso somático transmite la información sensorial al sistema nervioso
autónomo, lo cual crea arcos de decisión reflejos u autónomos al cerebro, esto
acelera la toma de decisiones corporales para mantener la homeostasis. El
sistema nervioso autónomo posee una subdivisión, el llamado simpático y el
parasimpático. El sistema nervioso simpático controla la respuesta de combate o
escape transmitida desde el cerebro, su control es potente ya que pone a los
órganos a trabajar al máximo por el bien de la supervivencia, pero sus efectos
a largo plazo puede dañarlos. En oposición el parasimpático regula las
actividades regulares del cuerpo, distribuyendo y ahorrando la energía para las
funciones cotidianas y repetitivas. Los nervios del sistema nervioso autónomo
simpático y el parasimpático envían sus fibras a todos los órganos del cuerpo,
con funciones antagonistas, mientras uno está activo el otro se inactiva. Esto
se logra mediante diferentes tipos de neurotransmisores, siendo los más famosos
la epinefrina y norepinefrina, estimulantes del sistema nervioso simpático.
El sistema
nervioso parasimpático emplea la acetilcolina “igual que el sistema nervioso
somático”. El sistema nervioso simpático actúa como un todo unificado, por lo
que sus efectos se manifiestan rápido y al mismo tiempo, justo lo que se
requiere para responder ante problemas de emergencia. Los estímulos permiten un
aumento del poder neuromuscular y del metabolismo anaeróbico esencial para una
mayor potencia muscular que proporciona picos de fuerza y picos de velocidad
propios de la respuesta de Combate o Escape (YouTube) (YouTube). En contraposición se inhiben los órganos con
efectos a largo plazo “que completan su funcionamiento en horas” como el
sistema gastrointestinal. El sistema parasimpático ocurre de forma
independiente. Esto se debe a los ganglios. Los ganglios funcionan como
microcerebros donde los somas neuronales parecen comunicarse entre sí,
sincronizando la información de las dendritas. En el sistema simpático existen
ganglios maestros centrales que unifican la respuesta, mientras que los
ganglios del parasimpático no están centralizados y se ubican de forma más
cercana a los órganos que controlan.
20. Fisiología del sistema nervioso humano
|| Regresar al Índice ||
Ya hemos descrito varias de las principales regiones del sistema nervioso central con algunas de sus funciones principales. Ahora analizaremos más a fondo algunos detalles relacionados con su funcionamiento. El cerebro se encuentra involucrado en actividades coordinadas del comportamiento humano, y por lo tanto se considera como un sistema de integración. En general se pueden describir tres conjuntos de funciones básicas: motivación; aprendizaje y memoria; y comunicación. Como sistema motivacional es responsable de crear un empuje a base de emociones para distintos comportamientos y actividades humanas, las cuales pueden ser socialmente aceptadas o no, el punto real es satisfacer necesidades básicas como conseguir energía, reproducirse o simplemente sobrevivir. Como sistema de memoria y aprendizaje permite que la información sensorial y las emociones puedan almacenarse para crear respuestas mucho más rápido.

Figura 20.1. Anatomía del cerebro. En
el modelo anterior tenemos un corte medio sagital del cerebro humano, es la
visión típica de los modelos dibujados en dos dimensiones para el cerebro
humano. Aunque la mayoría de las partes son bastante iguales al idioma español,
las referentes al tallo cerebral "brainstem"
reciben otros nombres: midbrain
"mesencéfalo" y pons
"tronco cefálico".
Como sistema de comunicación refiere a los
mecanismos de comunicación complejos como el habla o el lenguaje escrito.
Muchas de estas funciones dependen del tallo cerebral. El cerebro también hace parte del sistema endocrino, no
solo por las funciones paracrinas de la mayoría de los neurotransmisores, sino
porque algunas de sus partes segregan verdaderas hormonas, las cuales pueden
movilizarse por el sistema circulatorio a otras regiones del cerebro, regulando
entre otras el crecimiento, el desarrollo sexual, los comportamientos e
incluso, manifestaciones no cerebrales de las emociones, especialmente en el
tracto gastrointestinal o en el sistema cardíaco. Estas manifestaciones
extracerebrales de las emociones fueron las que en la antigua Grecia llevaron a
filósofos de la talla de Aristóteles a pensar que el corazón o el estómago se
encargaban del pensamiento.
20.1 Actividades funcionales del cerebro
Las actividades funcionales del cerebro dependen de las
regiones que lo componente y que ya hemos mencionado anteriormente. El cerebro
de organiza de forma estratificada, en las que las partes más antiguas poseen
mayor influencia que las más novedosas. Nuestro conocimiento acerca del
funcionamiento del cerebro aún es muy incompleto. Aún no sabemos cómo es que
las redes de neuronas transforman los impulsos sinápticos en emociones, ideas o
imágenes mentales, lo cual lo convierte en el órgano más complejo del cuerpo de
los vertebrados, la última frontera de la anatomía y la fisiología. La
neurobiología ha avanzado muchísimo en las últimas décadas, tal vez mucho más
rápido que la psicología, sin embargo, esta información es muy concreta y
rebasa los objetivos básicos de esta serie de escritos.
20.2 Función endocrina o función nerviosa
Nosotros los humanos tenemos la tendencia a querer separar estructuras que funcionan de forma integrativa, este es el caso de los sistemas nervioso y endocrino. Generalmente por sus descripciones puramente anatómicas se los podría considerar como cosas completamente diferentes, un órgano es eléctrico y el otro es químico, pero tales distinciones son meramente apariencias. Las neuronas son células con función glandular esencialmente, su alargamiento y procesos iónicos son adaptaciones secundarias de un tejido que originalmente debía ser paracrino-endocrino y aún lo es. Como evidencia de esto tenemos al hipotálamo, una región del cerebro que manifiesta bellamente la función glandular endocrina y la función nerviosa de potenciales de acción.

Figura 20.2. Nervioso y endocrino. El
sistema endocrino y nervioso están estrechamente vinculados y emplean las
mismas moléculas de señalización, las neuro-hormonas como las catecolaminas.
En la Figura 20.2 podemos apreciar la
pretensión algo simplista de separar las dos funciones de control en dos tipos
de sistemas diferentes, a la izquierda se encuentra la función endocrina en la
que una glándula descarga a la sangre, la cual transporta al tejido a ser
afectado. A la derecha tenemos una neurona que mediante un potencial de acción
"impulso nervioso" afecta al tejido al final. Por ningún lado se
menciona que la neurona también descarga compuestos químicos, ya sea de forma
cercana "paracrina" o a la sangre "endocrina".
La definición de un órgano endocrino es que
es un sistema de glándulas que descarga sus mensajeros químicos llamados
hormonas al torrente sanguíneo, ya que carecen de conductos propios
independientes. En este sentido cabe hacer notar la existencia del sistema de
vasos porta entre el hipotálamo y la hipófisis, el cual es un conducto
sanguíneo que permite una comunicación por sustancias que fungen
simultáneamente como hormonas o neurotransmisores dependiendo del punto de
vista.
Una vez que el hipotálamo activa a la hipófisis y la pituitaria, estas funcionan como órganos endocrinos liberando hormonas que reguilan procesos en el resto del cuerpo por medio de los vasos sanguíneos. A este nivel, los compuestos químicos son poco diferenciables y se clasifican como reflejos neuroendocrinos para representar una función íntimamente relacionada. Algunas hormonas-neurotransmisores empleados en este punto son la arginina, la vasopresina y la oxitocina.

Figura 20.3. Hormonas y neurotransmisores. Las
hormonas son moléculas que fluyen por la sangre a largas distancias con efectos
a largo plazo; mientras que los neurotransmisores afectan a cortas distancias rápidamente.
Algunas moléculas tienen ambos efectos
Todos los vertebrados poseen un hipotálamo,
haciéndolo uno de los órganos neuronales más antiguos, y en los seres humanos
es del tamaño aproximado de una almendra. A pesar de su pequeño tamaño domina
fuertemente las funciones “más avanzadas” del cerebro, coordinando señales
provenientes de los sentidos a través del tallo cerebral, así como procesos de
aprendizaje. La razón de que un órgano del tamaño de una almendra domine sobre
los lóbulos frontales del pensamiento analítico son simples, el hipotálamo está
allí para organizar funciones y reflejos vitales, regulando la homeostasis del
cuerpo y enviando señales al resto del cerebro para mantener la homeostasis, en
este sentido el hipotálamo presta atención a lo básico, todo lo demás son solo
lujos.
El hipotálamo debe estar en contacto directo
con la sangre para poder medir la cantidad de muchos compuestos químicos
relevantes para mantener la homeostasis, pero que regularmente se encuentran
excluidos del cerebro por medio de la barrera hematoencefálica. Para ello,
algunos nódulos hipotalámicos denominados órganos circunventriculares poseen
fenetras “poros” en su barrera hematoencefálica, permitiendo a este un contacto
libre con la sangre. Las funciones que describiremos a continuación representan
una combinación de la función nerviosa y endocrina actuando simultáneamente.
20.3 El cerebro y la conducta alimenticia
(YouTube) El balance energético es uno de los componentes basales de la
homeostasis, la disponibilidad de ATP y otros nucleósidos trifosfatados permite
que las células trabajen, es como el efectivo en una ciudad o una empresa, sin
dinero nada se mueve, en la célula sin nucleósidos trifosfatados y otros
portadores energéticos nada sucedería. El cuerpo no gasta toda la energía que
consume, de hecho, la energía consumida se divide en tres, la energía empleada
para realizar trabajo químico-mecánico, la energía almacenada en moléculas de grasa
y otros macropolímeros y la energía desperdiciada en forma de calor para
cumplir con los límites impuestos por la segunda ley de la termodinámica. La
energía calórica y de trabajo se conoce como gasto energético o de forma más
común como la Tasa metabólica Básica. La Tasa Metabólica Básica depende de la
cantidad de energía que demanda el cuerpo, y no todos los cuerpos poseen la
misma. Los individuos con un mayor nivel de entrenamiento y por ende con mayor
cantidad de tejidos de alta demanda energética, poseen niveles basales más
altos que aquellos cuerpos que se encuentran en un estado sedentario o de
estasis.
Esto se debe a que el sedentarismo es una
estrategia evolutiva para almacenar energía en periodos de escases, solo que
los biólogos la conocen con un nombre más “cool” en la naturaleza, hibernación.
Cuando un animal se prepara para hibernar se satura de comida y luego establece
una conducta sedentaria, su cuerpo está adaptado en consecuencia para acumular
grasa y mantenerla lo más posible mientras su cuerpo permanezca inactivo. La
demanda energética es fácilmente medible por el hipotálamo de muchas formas,
puede determinar desechos metabólicos producto de la actividad física de alta
demanda, como las necesarias durante la cacería o la recolección de frutos, lo
cual implica arduas caminatas y el encuentro con depredadores, todas
actividades aeróbicas de esfuerzo físico, las cuales bajan las reservas de
grasa del cuerpo al incrementar la tasa metabólica Basal. La baja en el tejido
graso se mede en forma de una disminución en la hormona leptina, la cual
es generada por el tejido graso. Cuando esto sucede el hipotálamo desencadena
la respuesta de alimentarse.
Esta regulación se logra mediante la
liberación de neurohormonas de tipo peptídico, estas se clasifican como
orexigénicas “estimulantes del apetito” o anorexigénicas “bloqueadores del
apetito”. Estas hormonas pueden afectar tanto los hábitos alimenticios a largo
plazo como la sensación de satisfacción al alimentarse, en efecto, el sentirse
lleno después de comer depende más de la liberación de hormonas por parte del
tracto gastrointestinal que del hecho mecánico de tener el estómago físicamente
lleno.
Los estados de ánimo pueden afectar mucho la
red hormonal que controla la alimentación, debido a que es dependiente de la
historia evolutiva. Puestos en el contexto, cuando un grupo de primates afronta
una temporada de poco alimento, se encuentran ante un periodo de tiempo en el
que no vale la pena desplazarse y gastar energía, por lo que presentarán una
actitud sedentaria de bajo gasto energético, adicionalmente el estrés libera
hormonas que favorecen la acumulación de grasa para aguantar el periodo de
escases. Por lo tanto, es lógico pensar que al juntar estos factores evolutivos
se herede una fisiología que conspira para acumular grasa cuando nos sentimos
preocupados, evitamos la ingesta de alimentos y no hacemos ejercicio, el cuerpo
simplemente cree que estamos en un invierno fuerte o una poderosa sequía, baja
la Tasa Metabólica Básica y se hace receptivo a acumular grasa.
¿Cómo lo resolvemos? Una opción sería
aumentar las cantidades de leptina y engañar al cuerpo haciéndole creer que
tiene más grasa, sin embargo, los medicamentos empleados para tal proeza tienen
el mismo efecto de las drogas de abuso, generan resistencia por disminución de
receptores de leptina, al final se generan efectos secundarios en el que el
cuerpo cree que tiene menos grasa de la que en verdad posee, y las
consecuencias son un aumento de peso mórbido. En la actualidad se está trabajando
con los receptores de endocanabinoides, los cuales reciben neurohormonas que
estimulan el apetito, pero evidentemente también se puede caer en el problema
de la resistencia. Parece ser que la única forma sana de bajar peso es hacerle
creer al cuerpo que se encuentra en un ambiente de plenitud, en el que el
cuerpo hace mucho esfuerzo para consumir alimento. Nota, inhibir la
alimentación es contraproducente ya que le hace pensar al cuerpo que está en
una estación de escases incrementando las hormonas de acumulación de grasa, por
eso las dietas de bloqueo de calorías simplemente no funcionan.
20.4 El desarrollo sexual y el cerebro
(YouTube) El hipotálamo controla el desarrollo sexual y la conducta sexual. El
desarrollo sexual es controlado por medio de una hormona maestra que inicia
todo lo demás denominada Hormona Liberadora de Gonadotrofinas mejor conocida
como GnRH por sus siglas en inglés. Esta hormona es tan potente que por sí sola
inicia el desarrollo sexual al inicio de la pubertad, y posee funciones tan
diversas que no se las conoce todas completamente aun en la actualidad. El
aumento de la GnRH es regulado mediante la alimentación retroactiva desde las
gónadas mediante la detección por segundos mensajeros “receptores unidos a
proteínas G” de la testosterona producida por las gónadas. Cuando esto se logra
se genera un ciclo de liberación hormonal que puede ser mensual “mujeres” o
diario “hombres” cuando la pubertad está a punto de terminar.
Los esteroides no son solo detectados por el
hipotálamo, otras regiones del cerebro poseen receptores para testosterona,
estradiol y otras hormonas masculinizantes o feminizantes, lo cual crea
diferencias anatómicas en el cerebro masculino y femenino, las cuales crean
consecuencias en su funcionamiento. El problema radica en que el cerebro posee
una gran complejidad, por lo que los efectos fisiológicos en el cerebro humano
no se han podido establecer de forma tan clara como las diferencias anatómicas.
20.5 Cerebelo y deporte
(YouTube) El éxito de un individuo en cualquier deporte
depende en última instancia de su cerebelo. La coordinación, los reflejos, los rápidos
tiempos de reacción y el aprendizaje quinestésico dependen de esta pequeña
estructura cerebral. Recibe información de la médula espinal y los sistemas
sensoriales, y utiliza esta información para regular y ejecutar movimientos
motores. Es el cerebelo el que ayuda a un tenista profesional a golpear con la
pelota con tanta precisión y una parada corta para atrapar un impulso de línea.
Los estudios realizados en atletas de élite han encontrado diferencias en las
conexiones neuronales de sus cerebelos en comparación con los no profesionales.
Esto probablemente se deba a años de entrenamiento y práctica que han
optimizado redes neuronales específicas en el cerebro de un atleta. Cada vez
que practicas deportes y actividad física, ejercitas tu cerebelo, reforzando
redes musculares que permiten ejecutar movimientos precisos sin pensar, lo cual
hace más corto el tiempo de reacción.
20.6 El reloj cerebral
El hipotálamo regula eventos repetitivos, que
pueden clasificarse en dos categorías principales, rítmicas y cíclicas. Un
evento rítmico es aquel que ocurre a diario, mientras que uno cíclico posee un
patrón de un mes o más. Un ejemplo de un evento rítmico son los ritmos
circadianos de vigilia y descanso, los cuales dependen de la estimulación
lumínica mediada por los ojos y otros órganos neuronales como la glándula
pineal. Sin embargo, el lóbulo prefrontal puede afectar estos ciclos e
imponerse parcialmente invirtiendo la vigilia y el descanso con respecto a la
luminosidad, pero nunca pueden desaparecer el impulso del sueño, ya que es un
estado que requiere el cuerpo para mantener la homeostasis mediante la
reparación de los tejidos.
Un ejemplo de un evento cíclico es el ciclo menstrual de las mujeres que en promedio dura 28 días, pero que puede ser afectado por factores ambientales como la convivencia con otras hembras sexualmente fértiles, lo cual genera sincronización del ciclo menstrual. El control se hace mediante la neurohormona melatonina segregada ya sea por la glándula pineal o por neuronas conectadas a la retina. Esta hormona llega a una región hipotalámica llamada centro neuroquiasmático, el cual funciona como un marcapaso. Normalmente en los humanos la melatonina se libera en la noche, sin embargo, el centro neuroquismático es capaz de mantener su propio ritmo en ausencia de melatonina o de estar conectado a un cuerpo “cultivos in vitro de sus tejidos mantienen sus propios ritmos en ausencia de estímulos externos”.

Figura 20.4. La melatonina y el reloj biológico. En
los vertebrados con un cerebrum pequeño esta recibe la luz directamente por
medio de un orificio pineal en el cráneo, pero en los que tienen un cerebro grande, la información lumínica
proviene de los ojos (YouTube).
Los trabajadores que experimentan cambios
horarios pueden sufrir de problemas cognitivos debido a la falta de sueño, esto
se debe a que el centro neuroquismático requiere de tiempo para reiniciar el
programa de recepción de melatonina, aunque algunos estudios han revelado que
una juiciosa administración de melatonina exógena puede ayudar a resetear más
rápido el ritmo circadiano.
21. El cerebro y el sueño
|| Regresar al Índice ||
(YouTube) La función del sueño es una de las más
relevantes, por lo que le dedicaremos un poco más de espacio. El sueño sigue los ritmos circadianos y es un
recurrente estado de conciencia alterada, en la cual la actividad sensorial y
motora se encuentra suspendida, así como la mayoría de los movimientos
voluntarios. Durante el sueño las rutas
metabólicas anabólicas, es decir, aquellas que se encargan de la síntesis de
macromoléculas para la reparación de daños fisiológicos, aumentan de forma
significativa reparando al cuerpo. El sueño se encuentra regulado por el reloj
biológico en el centro neuroquismático. El reloj biológico para el sueño
trabaja en tándem con la adenosina, un nucleósidos del ARN que puede
energizarse mediante la adición de grupos fosfato, pero que también actúa como
neurotransmisor, inhibiendo la mayoría de los procesos funcionales asociados
con los estados de conciencia de la vigilia.
La adenosina se eleva durante el curso del día,
y cuando se acumula se induce una sensación de agotamiento cognitivo. La
hormona melatonina se libera durante este estado de agotamiento induciendo el
sueño, causando simultáneamente una disminución de las actividades metabólicas
catabólicas que tiende a liberar calor. La actividad eléctrica asociada con el
sueño fue descubierta en 1937 empleando una nueva tecnología denominada
electroencefalogramas. En esta sección analizaremos algunos aspectos
fisiológicos del sueño.
21.1 El electroencefalograma
(YouTube) El electroencefalograma o EEG tiene una
historia bastante extensa que se rastrea hasta finales del siglo XIX cuando
Richard Caton publicó en 1875 sus resultados sobre la actividad eléctrica del
cerebro expuesto de conejos y monos. Sus trabajos fueron rápidamente retomados
por otros investigadores como Adolf Beck en 1890, Neminsky en 1912, Cybulski en
1914, hasta que finalmente en 1924 se publicó el primer EEG en humanos por
parte de Hans Berger. Berger también le dio nombre al procedimiento y al
instrumento tal como lo conocemos en la actualidad. El EEG es un instrumento
sensible de diagnóstico y seguimiento de la actividad eléctrica de la superficie
del cerebro mediante el uso de electrodos ubicados en posiciones específicas de
la cabeza. Aunque no es invasivo, en ocasiones si requiere un buen corte de
cabello y el uso de geles especiales que mejoran la conductividad, aunque otras
versiones emplean electrodos menos sensibles.
El EEG mide potenciales de acción realizados
por grupos completos de millones de neuronas en la superficie, lo cual se
representa en forma de picos y valles, las cuales fueron denominadas ondas
cerebrales. Con la investigación realizada en las décadas de 1920 y 1930 se
describió una nomenclatura relativamente simple para describir la actividad
cerebral empleando letras del alfabeto griego. A continuación, describiremos de
forma bastante resumida algunos de estos tipos de ondas cerebrales.
21.2 Las ondas cerebrales
(YouTube) Las ondas cerebrales medidas por un
electroencefalograma EEG cambian de forma correspondiente al estado de
conciencia que sufre el individuo. Un EEG mide la actividad eléctrica que se
manifiesta cerca de la superficie del cerebro y que es capaz de agitar
electrones cerca de la superficie de la cabeza. Esto genera valores de
frecuencia que va de 1 a 30 hertzios, con amplitudes de onda que van usualmente
de 20 a 100 milivoltios. Debido a que las ondas medidas son el producto de
potenciales de acción en redes muy complejas, las pulsaciones son altamente
variables y no están exentas de ruido de fondo constante.
A pesar de la variabilidad y el ruido,
diferentes estados de conciencia manifiestan diferentes patrones en los picos
de onda registrados en un EEG. Los mayores estados de conciencia se
caracterizan por ondas de picos altos o poca amplitud, mientras que los estados
de conciencia alterados en el descanso se caracterizan por picos bajos y
amplitudes de onda muy largas. Las ondas cerebrales se clasifican de acuerdo a
su frecuencia:
👉 Ondas alfa: Poseen ritmos de 8 a 13 hertzios, son las más
cortas y son características del estado de conciencia estándar o de alta alerta
cognitiva cuando una persona se encuentra despierta y consiente de las señales
ambientales, pero con los ojos cerrados.
👉 Ondas beta: Con ritmos de 13 a 30 hertzios, las ondas
extra son desencadenadas por el análisis de la información sensorial de uno de
los sentidos, la vista. Con solo abrir los ojos el estado de conciencia duplica
su actividad cerebral. Más allá de 30 hertzios se las denomina gamma, un estado
de percepción incrementado por medio de químicos exógenos.
👉 Las ondas teta y
delta: Las primeras tienen rangos de 4 a 7 hertzios
y las segundas 0.5 a 4 hertzios que se manifiestan durante el sueño inicial y
el sueño profundo respectivamente.
👉 Otros: Existen otros patrones no estandarizados que se manifiestan durante estados alterados de conciencia como la epilepsia. La epilepsia es un desorden neurológico caracterizado por descargas eléctricas espontáneas que generan lapsos de baja atención, pasando por la pérdida de la conciencia hasta espasmos musculares no controlables.

Figura 21.1. Las ondas cerebrales.
21.3 Las etapas del sueño
(YouTube) (YouTube) La actividad eléctrica asociada al sueño fue reportada inicialmente en
1937 mediante el uso del electroencefalograma EEG. Los resultados del EEG
durante el sueño revelan un cambio persistente en las amplitudes y
frecuencias. Durante milenios se pensó que el sueño era un estado similar al de
la muerte, de allí una enorme cantidad de metáforas, sin embargo, el EEG reveló
que el cerebro muerto y el cerebro dormido son cosas completamente diferentes.
Un cerebro muerto no presenta actividad cerebral, pero por el contrario el
cerebro dormido mantiene una actividad bastante alta, la cual pasa durante
varios tipos de etapas que pueden sucederse de forma cíclica.
El patrón EEG durante el sueño es cíclico,
repitiéndose en amplitudes de 90 minutos aproximadamente, durante un periodo
estándar de sueño de unas 7 u 8 horas. Adicionalmente algunas etapas del sueño
fueron vinculadas con movimientos involuntarios de los ojos, por lo que el
diseño experimental del análisis del sueño no solo involucra un EEG, también
requiere de del análisis de la actividad eléctrica de los músculos de los ojos
llamada electromiografía. El sueño clásicamente se divide en dos categorías
bautizadas por sus nombres en inglés, siendo REM y NREM. Cada etapa posee
características funcionales, neurológicas y fisiológicas distintivas.
Posteriormente la etapa NREM fue dividida en 4 etapas secundarias que se
designan con la letra N mayúscula, desde N1 hasta N4.
21.4 El sueño no REM
El sueño se distingue por el movimiento involuntario de los ojos, el cual está relacionado con las “visiones del sueño o imágenes oníricas”. En el idioma español no tenemos una palabra que distinga el acto de dormir “sleep en inglés” con las ilusiones oníricas del sueño “dreams” de forma clara, lo más cercano es referirnos al acto de dormir como “el sueño” y a las ilusiones oníricas como “los sueños”.

Figura 21.2.
Sucesión de las etapas de una noche de sueño saludable.
En el sueño no REM no hay un movimiento
rápido de los ojos, sin embargo, los neurobiólogos han sido capaces de
distinguir varias etapas en su desarrollo. La etapa N1 se denomina somnolencia,
las ondas cerebrales pasan de alfa a teta, lo cual disminuye el tono muscular y
el estado de alerta. Durante la etapa N2 la actividad muscular consiente se
detiene, así como el estado de conciencia, pero no hay sueños. Esta etapa es la
más común, ocupando entre el 45 y el 55% del tiempo que permanecemos dormidos,
esto explica porque el tiempo que recordamos sobre los sueños es tan corto en
comparación al tiempo real de descanso.
La etapa N3 se puede referir como un primer
momento del sueño profundo, caracterizado por ondas cerebrales delta. Esta
etapa es extraña ya que puede contener malos sueños sin existir un rápido
movimiento de ojos característicos de una pesadilla normal, algunas personas
pueden presentar incontinencia nocturna, sonambulismo “caminar dormidos” y
somnílocua “hablar dormidos”. La etapa N4 es una designación histórica que ya
no parece emplearse (Fuller, Gooley, & Saper, 2006), originalmente se
la designaba como una etapa de sueño aún más profunda que N3 pero no tanto como
la etapa REM, en la actualidad esta designación ha quedado en desuso,
considerándose la etapa N4 como una parte de la etapa N3. Las ondas cerebrales
tienen muy poca frecuencia y se lo denomina como sueño de ondas lentas. El
proceso no es como los cambios de un carro, para llegar a la etapa REM hay que
pasar por un ciclo: despierto → N1 → N2 → N3 → N2 y finalmente desde N2 se
llega a REM.
21.5 El sueño REM
(YouTube) (YouTube) El sueño REM se caracteriza por un
movimiento rápido de los ojos, de hecho REM es el acrónimo de “Rapid Eye
Movement”, representando entre el 20% al 25% del tiempo total del sueño. En
esta etapa ocurren las experiencias oníricas o sueños que podemos recordar. El
sueño REM presenta una serie de características paradójicas, por ejemplo, en
lugar de presentar una amplitud de onda más larga que las etapas del sueño no
REM, las ondas cerebrales acortan su amplitud, lo que implica una mayor
actividad cerebral, llegando al patrón beta, como si el cerebro estuviera
despierto, pero sin que exista conciencia.
El sistema nervioso autónomo se encuentra en
un alto grado de excitación, la presión arterial y el rítmo cardíaco aumentan,
mientras que la respiración se torna irregular. En los hombres el sueño REM
también puede involucrar erecciones del pene. Los hombres con impotencia
fisiológica generalmente no pueden tener una erección firme durante el sueño
REM. Cuando los sujetos son despertados de un sueño REM usualmente reportan
experiencias oníricas, que son las ilusiones mentales o sueños. Otra
característica del sueño REM es que la mayoría de los músculos voluntarios se
paralizan. En ocasiones esta parálisis puede no estar sincronizada con la
recuperación de la conciencia, por lo que muchos de nosotros hemos
experimentado la parálisis del cuerpo mientras nos despertamos de un mal sueño.
21.6 La deuda del sueño
¿Cuánto tiempo debemos dormir? De hecho
¿debemos dormir en absoluto? La actitud ante el sueño ha ido cambiando con el
curso de la historia y nuestra capacidad tecnológica de crear productividad
durante las horas en las que no podemos emplear la luz o el calor del Sol.
Para el siglo XIX las actitudes culturales
con el sueño se habían tornado hostiles, el sueño era, y es en algunos casos,
considerado como una enfermedad que debía ser combatida a base de medicamentos.
Durante la época comenzaron a emplearse los primeros medicamentos contra el
sueño, estimulantes como la cafeína y la cocaína permitían un estado de
conciencia y actividad continuas, sin que la sensación de cansancio disminuyera
la productividad.
A pesar de que desde la biología se ha ido descubriendo
que el sueño hace parte de la biología natural del cuerpo como una etapa
importante de regeneración celular, las actitudes culturales que hemos heredado
aún son hostiles. Primero comenzaremos determinando el tiempo que necesitan las
personas dependiendo de su rango de edad. Los recién nacidos deben dormir
aproximadamente 16 horas al día, la mitad de las cuales son sueño REM. Los
adultos normales deben dormir entre 7 y 8 horas al día, un cuarto de las cuales
son sueño REM. El procentaje de sueño REM declina con el aumento de la edad,
así como la habilidad de llegar al estado N3 del sueño NREM. Durante el sueño
nuestros tejidos se regeneran, además producimos antioxidantes que nos
rejuvenecen, nuestro músculo crece después del ejercicio, nuestros huesos se
regeneran después de las lesiones, nuestro tejido graso es consumido por el
metabolismo necesario para regenerar todos los demás tejidos (Egydio, Pires,
Tufik, & Andersen, 2012; Everson, Laatsch, & Hogg, 2005; Kahan,
Andersen, Tomimori, & Tufik, 2010). Por lo anterior, nuestros cuerpos son habidos de un descanso adecuado, y
por eso el sueño es un acto tan placentero.
La deuda del sueño se define como la
depravación de estados profundos del sueño como N3 y REM. De todos los sistemas
del cuerpo humano, el que presenta mayor desgasto normalmente es el nervioso, y
en consecuencia es el primero que empieza a manifestar problemas cuando la
deuda del sueño aumenta. Se puede experimentar dolor, alucinaciones, cambios de
personalidad, microsueños entre muchos otros. Una deuda de sueño que se
mantiene de forma crónica por años en forma de dormir menos de 7 horas diarias
puede provocar que los tejidos no puedan regenerarse adecuadamente, lo cual
provoca: envejecimiento prematuro, irritabilidad, limitación cognitiva, perdida
de la memoria, temblores musculares y pérdida en la masa muscular, degradación
del colágeno de la piel y aparición de arrugas de forma prematura. Evidentemente si los tejidos no se regeneran
la grasa no se consume, por lo que es un factor de riesgo para el aumento del
peso y todas las demás enfermedades asociadas como por ejemplo hipertensión,
arteriosclerosis, diabetes tipo 2 e incluso una disminución de la efectividad
del sistema inmune.
22. Generalidades de la adicción
|| Regresar al Índice ||
(YouTube) En los artículos previos hemos examinado la
anatomía y la fisiología de los circuitos del sistema nervioso. Ahora
examinaremos como es que algunas sustancias químicas capaces de realizar
reacciones cruzadas con los receptores de neurotransmisión son capaces de
alterar el estado homeostático de la mente. Analizaremos las razones
fisiológicas de la dependencia a estas sustancias y también las consecuencias
que tienen, no solo en el sistema nervioso, sino en otros sistemas de órganos.
Posteriormente describiremos los efectos de algunas de las sustancias de abuso
más comunes, para finalmente ofrecer una explicación de su existencia.
En términos
básicos, las sustancias de abuso psicoactivo afectan específicamente la
sinapsis neuronal, estimulando circuitos cerebrales en el tallo cerebral o el
sistema límbico, todos relacionados con el sistema de recompensas del cerebro.
Su efecto es poderoso y pueden alterar el estado de conciencia y emocional del
individuo con independencia a sus interacciones socioafectivas. La mayoría de
estas sustancias son liposolubles, por lo que pueden atravesar la barrera
hematoencefálica con gran facilidad, y a diferencia de un neurotransmisor
normal, también poseen un efecto endocrino, es decir, pueden transportarse por
la sangre a grandes distancias sin que el cuerpo sea capaz de degradarlas
rápidamente, a diferencia de los neurotransmisores que son degradados
efectivamente por enzimas digestivas.
Los efectos
postsinápticos de una droga de abuso son variados:
👉 estimulando la liberación de otros
neurotransmisores por medio de proteínas G acopladas a receptores para la
sustancia de abuso,
👉 Inhibiendo la estimulación de algún grupo de
neuronas por medio de la apertura de canales del ion cloro que hiperpolarzan la
neurona o
👉 inhibiendo las enzimas de reciclaje de los
neurotransmisores, y en consecuencia amplificando los efectos de un determinado
neurotransmisor. La degradación de los receptores es disparada por una
prolongación del tiempo en que el complejo efector-receptor se mantiene unido,
el receptor se endocita en una vesícula de tráfico celular, luego enviada al
cuerpo neuronal donde es degradada.
22.1 Origen de la dependencia a nivel molecular
(YouTube) La adicción se da cuando hay una dependencia a
un estímulo, esta tiene varios orígenes simultáneos, pero por el momento nos
enfocaremos en su naturaleza biológica a nivel molecular. En este orden de
ideas, la dependencia se origina por la degradación de los receptores de
membrana por exceso de estimulación de neurotransmisores. En el video vemos el
caso de los opioides, sin embargo, esta es una ruta de acción casi que
universal para todas las drogas de abuso que crean una dependencia fisiológica.
Cuando el ligando
"molécula de abuso" se conecta con el receptor de siete pliegues
membranales activa la proteína G. La sobre exposición o la prolongada unión con
el neurotransmisor activan mecanismos homeostáticos que endocitan el receptor
en una vesícula de transporte hacia el soma de la neurona donde es eliminado.
Es posible que el cuerpo interprete la sobreexitación del receptor como un daño
en sus mecanismos de afinidad y por eso lo degrade. Pero la consecuencia final
es una menor cantidad de receptores y la necesidad de consumir más sustancia
para obtener el estímulo al mismo nivel.
22.2 Síndrome de abstinencia y la dependencia
Las
sustancias de abuso se distinguen de otras sustancias psicoactivas debido a
tres factores clave:
👉 el cuerpo se hace tolerante o inmune a su efecto,
👉 y al mismo tiempo pierde la capacidad de generar
el efecto natural con sus neurotransmisores nativos, y
👉 los órganos generan efectos sobre las neuronas del
dolor cuando se retiran las sustancias de abuso.

Figura 22.1. Síndrome de abstinencia.
Aunque al principio parece solo un defecto de personalidad, la adicción es una
enfermedad del sistema nervioso a la que algunos individuos están predispuestos
molecularmente (YouTube) (YouTube) (YouTube).
La tolerancia es una disminución progresiva
en la efectividad de una sustancia de abuso a lo largo de una serie de
administraciones, la tolerancia puede ser permanente o restaurarse después de
un lapso de descanso. Cuando una persona cae en un ciclo de tolerancia, debe
consumir una mayor cantidad para lograr el mismo efecto inicial.
La tolerancia puede ser cruzada, moléculas
con estructuras similares pueden crear tolerancia entre sí, por ejemplo: el
abuso de la codeína puede crear tolerancia moléculas semejantes como la morfina
o la heroína.
La dependencia tiene una definición más
compleja, pero la podemos clasificar en dos tipos, la dependencia fisiológica y
la dependencia psicológica. En ambos casos se trata de fenómenos que provocan
que la persona consuma la misma sustancia cada vez en mayores cantidades. La
dependencia fisiológica también es conocida como el síndrome de abstinencia. El
cuerpo constantemente reajusta los receptores para mantener una normalidad
funcional, por lo que cuando una molécula de abuso es retirada del sistema se
crea un desequilibrio que puede bloquear o estimular rutas del sistema nervioso
que generalmente son interpretadas por el cerebro como dolor.
El asunto es que no es un dolor falso,
parafraseando a Matrix: “tu cerebro lo hace real”, los circuitos estimulados por el síndrome de abstinencia son recíprocos,
creando espasmos musculares que a su vez reenvían más señales de dolor, maros,
jaquecas, vómitos entre otros. En consecuencia, el síndrome de abstinencia es
extremadamente doloroso, e incluso puede llegar a ser mortal. Los adictos
graves a una sustancia la consumirán, no para sentir placer ya que serán muy
tolerantes, sino para evadir el síndrome de abstinencia y los terribles dolores
físicos que esto acarrea. Lo anterior es un caso común en drogas de abuso como
la nicotina y la heroína; pero no tan claro en otras sustancias como la
ayahuasca.
Más
problemático es la dependencia psicológica. El síndrome de abstinencia se puede
controlar médicamente, y a esto generalmente lo denominan limpieza. Sin embargo,
las drogas de abuso no solo estimulan los centros nerviosos del placer, esto se
debe a que los circuitos emocionales también se encuentran vinculados al
sistema límbico y al tallo cerebral, por lo que desarrollan una dependencia
psicológica que no puede ser borrada de forma efectiva por tratamientos
externos. El individuo mismo debe desarrollar conexiones psicológicas con su
medioambiente, de forma que su tejido emocional vuelva a estar unido con su
comunidad, en lugar de estar enfocado a lo que las moléculas de abuso pueden
proporcionar.
La
dependencia psicológica es mucho más ambigua y compleja, y es la razón por la
cual muchas personas recaen en el consumo. Otra razón es que los receptores del
cerebro y los circuitos nerviosos estimulados jamás recuperan totalmente su
homeostasis normal, por lo que estos individuos siempre tendrán una sensación
de carencia.
22.3 El placer y el comportamiento evolutivo
(YouTube) (YouTube) ¿Que
distingue un comportamiento adictivo, obsesivo de, por ejemplo, el
entrenamiento para alcanzar la maestría en una labor? En cualquiera de los
casos el cerebro recibe neurotransmisores que estimulan los circuitos de
recompensa y se estimula el sistema nervioso simpático “como en los deportes”. Una
respuesta a esto involucra la conexión que ocurre entre los comportamientos
adaptativos, y la recompensa. Si un individuo involucra un comportamiento
negativo para la supervivencia de su prole, y sus genes inducen la generación
de neurotransmisores placenteros, las probabilidades de supervivencia serán muy
bajas. Es como pensar en ratas que experimentan placer en observar el rostro de
una serpiente, simplemente no funcionaria.
Los
comportamientos adaptativos como: la ingesta de alimentos con alto contenido
calórico, la reproducción sexual, la victoria en combate contra competidores y
depredadores, o la mera expectativa durante la caza; se refuerzan si están
enlazados a neurotransmisores relacionados al placer y además aumentan la
probabilidad de supervivencia y reproducción de la población. De esta manera
aseguramos almacenar suficiente grasa para el invierno, tener descendientes en
gran número para afrontar la selección natural, desear la victoria ante los
problemas y las dificultades, o meramente desear un objetivo con ambición.
Un comportamiento
adictivo que mejora la integración del individuo con su sociedad, que mejora su
respuesta ante sus responsabilidades cotidianas no se lo considera adictivo
para nada. La adicción se señala en el momento en que la recompensa por
comportamientos convenientes es secuestrada por comportamientos poco
convenientes para la supervivencia y conexión social. En este sentido
comportamientos que son vistos como “buenos” como ser muy trabajador, se los
puede clasificar como adictivos si cortan las conexiones sociales, aislando al
individuo de sus familiares y amigos llevándolo a un ciclo de depresión. El
problema con las sustancias de abuso, es que la probabilidad de desconexión
social es mucho más alta.
22.4 No por ser natural es bueno
La mayoría de las
drogas de abuso se extraen o modifican a partir de sustancias generadas por
plantas u hongos. Las plantas generalmente no excretan sus metabolitos finales,
en lugar de ello se transforman en metabolitos secundarios y se almacenan en
las vacuolas de forma tan densa que estos se solidifican formando cristales(Stern, Bidlack,
& Jansky, 2008; Wayne, 2009). Estas moléculas son de gran importancia
pues son la fuente de las propiedades farmacéuticas o alucinógenas de muchos
vegetales, los cuales los emplean en sus relaciones de forrajeo (Hartmann &
Ober, 2000; Launchbaugh, Provenza, & Pfister, 2001; Levin, 1976; Mithöfer
& Boland, 2012).
En este sentido
la relación de forrajeo es clave, pues se trata de sustancias que las plantas y
los hongos emplean para defenderse de sus herbívoros respectivos,
desorientándolos, para que los depredadores puedan atacarlos y así disminuir la
cantidad de herbívoros que las lastiman. Los seres humanos a lo largo de los
milenios hemos aprendido a emplear ciertas cantidades de estas sustancias para
nuestros propósitos, sin embargo, jamás hay que olvidar que toda sustancia
natural generalmente evolucionó como un veneno, y por lo tanto al consumirla en
suficientes cantidades o mediante ciertas técnicas sus efectos peligrosos se hacen
patentes.
22.5 Anhedonia
Es la incapacidad para experimentar placer, la pérdida de interés o satisfacción en casi todas las actividades. Se considera una falta de reactividad a los estímulos habitualmente placenteros. Constituye uno de los síntomas o indicadores más claros de depresión, aunque puede estar presente en otros trastornos, como por ejemplo, en algunos casos de demencias (Alzheimer) y el trastorno esquizoide de la personalidad (Argyropoulos & Nutt, 2013; Olin et al., 2002).

Figura 22.2. Anhedonia. Uno
de los síntomas más inquietantes de la adicción es la anhedonia, pues no solo
es consecuencia de la adicción, sino también la refuerza, creando un círculo
vicioso que solo se rompe con mayores dosis o drogas más fuertes, culminando en
la muerte por sobredosis (YouTube).
Mientras que las primeras definiciones de
la anhedonia enfatizan experiencia placentera, modelos más recientes han puesto
de relieve la necesidad de considerar diferentes aspectos del comportamiento
divertido, como la motivación o deseo de participar en una actividad
(«anhedonia motivacional»), en comparación con el nivel de disfrute de la
actividad en sí misma («anhedonia consumatoria»). La cultura tiene una gran
influencia. Una conducta puede ser aceptada por un grupo cultural y no ser bien
vista por otro. Los factores biológicos son la causa principal donde se
encuentran las influencias perinatales y la salud física. Es el médico el que
determinará si es normal o anormal.
¿Por qué el drogodependiente, sea cual
fuere su adicción, es incapaz de experimentar satisfacciones de baja
estimulación? La contemplación, la intimidad, lo entrañable los percibe con
aburrimiento ¿Por qué solo se sienten a gusto buscando y notando sensaciones
intensas? El drogodependiente sobrelleva una vida extrema y vehemente,
instalado en el bipolo placer-displacer sin transiciones intermedias: o se
encuentra bajo los efectos agudos de la sustancia o sufre dramáticamente la
ausencia de la misma y su imperioso deseo, oscilando desde la intoxicación a la
insufrible abstinencia, desde el trance estimulativo o nirvánico al bajón
deprivativo.
En definitiva, apenas tiene épocas de
estabilidad sin protagonismo de la sustancia por presencia o ausencia
traumática de la misma. De hecho, cuando se rehabilita un adicto, una de sus
afirmaciones más frecuentes es que la vida sin drogas es agobiante y vacua. El
vacío emocional que deja como grave secuela la droga es uno de los handicaps
más importantes para la recuperación. El drogodependiente debe aprender a
encontrar satisfacción sin buscar la sensación (Baker, Piper,
McCarthy, Majeskie, & Fiore, 2004; Elman, Borsook, & Volkow, 2013;
Panksepp, Knutson, & Burgdorf, 2002). Esto es
causado precisamente por la degradación de los receptores de placer y/o de la
destrucción del tejido especializado en la sensación del placer (Puig & Gutstein, 2017; Wise
& Koob, 2014).
Estar inmerso en una depresión o en un
estado de ansiedad hace que las cosas que usualmente nos hacían disfrutar,
dejen de importarnos y de tener sentido para nosotros. Esto afecta, por
ejemplo, a nuestro apetito y a las relaciones sexuales, pero lo que
inmediatamente y de una manera especial se ven afectadas son las satisfacciones
de tipo social. La persona deja de disfrutar de las relaciones sociales, huye
de fiestas y de relacionarse con gente que no sea de su entorno más cercano, se
vuelve huidiza y huraña. En psicología se han establecido dos tipos de
cuestionarios para diagnosticar el grado de la anhedonia y sus causas, son las
llamadas escalas de Chapman. (Chapman, Chapman, & Raulin,
1976; Chapman, Edell, & Chapman, 1980).
El primer cuestionario basa sus preguntas
en la capacidad del paciente de obtener satisfacciones de tipo físico, el
segundo cuestionario interroga acerca de la capacidad para tener satisfacciones
de tipo social. El segundo cuestionario se ha revelado mucho más útil para el
diagnóstico de la anhedonia y sus causas, de tal manera que a través de éste se
han podido diagnosticar y observar el desarrollo de enfermedades como la
esquizofrenia, también el cuestionario sobre insatisfacción social ha sido muy
valioso para el estudio y tratamiento de trastornos como el autismo, que tiene
como una de sus principales características la anhedonia social.
23. Negación y negacionismo de la adicción
|| Regresar al Índice ||
“Yo lo controlo”
es una de las ideas más repetitivas que expresan los adictos cuando se los
confronta con las decisiones y el estilo de vida que han decidido llevar, la
negación de la adicción, o formalmente conocida como Sistema Delusional de la Adicción,
es un mecanismo psicológico que le permite al adicto convivir con su vida en
torno a la sustancia de abuso (Allan, 1997; Martínez-González,
Vilar López, Becoña Iglesias, & Verdejo-García, 2016; Pickard, 2016; Rinn,
Desai, Rosenblatt, & Gastfriend, 2002).
Este sistema
psicopatológico forma el núcleo de la enfermedad de la adicción en su parte
psicológica (MARTÍNEZ & AMAR, n.d.). Se compone de
una serie de creencias, mecanismos de defensa, negación, autoengaño y otras
distorsiones del pensamiento que en conjunto son conocidas como el Pensamiento
Adictivo y que combinadas con las causas moleculares de la anhedonia hacen de
la adicción una enfermedad tan difícil de combatir. Generalmente podemos
clasificar el autoengaño del adicto en tres categorías principales:
23.1 Negación o Autoengaño
Cuya función es la de separar al adicto de
la conciencia de las consecuencias que las adicciones tienen en su vida. De
esta manera se reduce la ansiedad y además se protege el sistema adictivo,
estableciéndose un equilibrio enfermo, del cual el adicto se mantiene atrapado,
y que además es la razón por la que el adicto insiste en volver a usar aún
luego de una crisis. A su vez la negación puede dividirse en varias
subcategorías que pueden ser concomitantes:
👉 Negación simple: La negación simple de
las situaciones negativas producto de la adicción.
👉 Minimización: Restarles importancia o
significado a los eventos relacionados con las consecuencias de la adicción.
👉 Racionalización: Asignar una razón
lógica a algo que no la tiene o que es por naturaleza irrazonable.
👉 Justificación: Justificar el uso en
virtud de algo que ocurrió en el pasado o por la forma en que lo tratan o por
cualquier otra condición existente en la vida del adicto.
👉 Proyección: Ver en los demás los problemas que el adicto está pasando en su
propia realidad, de modo que puede culpabilizar a otros de su problema. La
corporización de esta categoría es señalar que la sociedad da legalidad a otros
comportamientos adictivos, generalmente es una forma de falacia tu quoque,
pues, aunque es verdad que otras sustancias o comportamientos adictivos son
legales, eso no quita el hecho de que un comportamiento adictivo es
destructivo.
👉 Futurización: Salirse de la realidad
presente y vivir en el futuro como una manera de no ponerse en contacto con su
realidad presente. En este caso generalmente se corporiza en la expresión “lo
puedo dejar cuando yo quiera, pero hoy no”.
23.2 Distorsiones del Pensamiento
Propias de la adicción, son generadas por
las creencias adictivas. La función de estas distorsiones es producir
sufrimiento de modo que se justifique el uso de sustancias o conductas
adictivas para "aliviar" ese dolor de manera enferma. Sumadas a la
negación y el autoengaño, estas distorsiones tienden a facilitar el proceso
adictivo y despegan al adicto de la realidad de sus enfermedades.
👉 Catastrofización: Asignarle una
categoría de catástrofe aún al más leve inconveniente de la vida personal. Es
una manera de auto-sabotearse el crecimiento con el fin de justificar
finalmente el uso. El adicto desarrolla una visión catastrófica que lo coloca
en una actitud de víctima frente al mundo.
👉 Mortificación: La preocupación
constante ante cualquier problema hace de la vida cotidiana, una constante
ansiedad y sufrimiento que se usan como justificativo para continuar el uso.
👉 Rigidez: Dificultad para abrir la
mente y considerar otros puntos de vista. El adicto se aferra a sus
percepciones con una fuerza tal, que puede hacer la comunicación muy difícil,
sobre todo en el área del uso y el comportamiento adictivo.
👉 Control: El miedo es la emoción
fundamental en la personalidad del adicto y para manejar ese miedo, que muchas
veces es desproporcionado, el adicto desarrolla relaciones basadas en el
control interpersonal. Esto causa dolor en las relaciones y muchas veces
termina produciendo violencia.
👉 Pensamiento Blanco y Negro: La
incapacidad para ver los grises es muchas veces un fuerte componente de la
mentalidad adictiva, dificultando la comprension mutua y animando a los juicios
severos.
👉 Impaciencia e Intolerancia: El adicto
desarrolla una intolerancia marcada por situaciones que involucra incomodidad o
es y más bien actúa por impulsividad siguiendo la regla de "quiero lo que
quiero, cuando lo quiero y como lo quiero".
👉 Soberbia y grandiosidad: La
personalidad del adicto regresiona en cierta manera a la adolescencia,
aflorando muchas veces conflictos no resueltos con la autoridad y, conducta de
reto permanente. Esto se debe a una hipertrofia del ego como compensación al
deterioro en la autoimagen que se produce por el impacto de la adicción en la
vida de la persona.
23.3 Sistema de Creencias Adictivo
Todas estas distorsiones provienen del
sistema de creencias adictivo que conforman la raíz del desorden adictivo en el
plano mental. Para poder recuperarse de la adicción es necesario detectar y
cambiar dichas creencias adictivas.
23.4 Negacionismo industrializado, del tabaco
al calentamiento global y de regreso
Traduzco así lo que en inglés se llama DENIAL dado que la poca vitalidad del Castellano se extiende a la falta de palabras y la carencia de conceptos. A los Denialists los llamo Negacionistas, por lo tanto. El Negacionismo es el rechazo automático frente a pruebas científicas contundentes. Muchas veces se trata de gente a sueldo de organizaciones políticas, de empresas y monopolios, de Iglesias y religiones. Hay que diferenciar entre el escepticismo y el negacionismo. El escéptico toma un enfoque científico y analítico frente a ciertas afirmaciones. Un escéptico del cambio climático, por ejemplo, considera los hechos y las pruebas y está siempre dispuesto a cambiar su opinión admitiendo la evidencia. El escepticismo es una posición filosófica sana y de hecho nace con el Renacimiento Europeo y Descartes, por ejemplo) y se refuerza en la Ilustración, aunque hubo escépticos famosos entre griegos y romanos, por ejemplo, Epicuro. El negacionista ya tiene su idea de antemano y todo lo que lea lo acepta o rechaza según se adapte a lo que él ya cree antes de enterarse de los hechos -y eso suponiéndole honestidad intelectual, que rara vez la tiene porque la mala intención se le nota.

Figura 23.1. Drogas de combustión. Toda
sustancia consumida como vapor de combustión es cancerígena, independiente de
su legalidad o su naturalidad.
Ahora hay que
aclarar algo, todo puñetero adicto es un negacionista de que la droga lo esté
matando, sin embargo el negacionismo del tabaco es algo un poco más
industrializado, y recuerda más a las campañas anticientíficas que rodean a las
regulaciones de xenobióticos industriales como el plomo (Markowitz &
Rosner, 2013), a la ciencia del cambio climático (Jylhä, Cantal,
Akrami, & Milfont, 2016; Lewandowsky, Oreskes, Risbey, Newell, &
Smithson, 2015) o a teorías prestigiosas y bien
establecidas al interior de las comunidades científicas como la Teoría Sintética
de la Evolución (Allmon, 2011;
Sakar, 2009; Scott, 1997). Es decir, se trata de campañas
mediáticas financiadas por industrias y/o grupos religiosos por diversas
razones. De cierta manera se trata de una fabricación deliberada de ignorancia
en masa. Ahora la relación de otros tipos de negacionismo con la negación de
los efectos nocivos del tabaco es más que una analogía.
Naomi Oreskes y
Erik M. Conway dan algunas muy buenas e inquietantes respuestas en
su fascinante, detallado e ingeniosamente escrito nuevo
libro; Comerciantes de la duda (Oreskes &
Conway, 2011). En este libro muestran cómo un
pequeño grupo de estudiosos de la extrema derecha inmersa en la miopía de la
guerra fría, con una financiación sustancial de poderosos contaminadores
corporativos, lograron engañar a amplios sectores de la opinión pública
estadounidense y hacerles creer que la evidencia de origen humano calentamiento
global era incierta, irracional, políticamente manipulada y no debía
servir de base para cualquier tipo de acción política.
Su historia
comienza con lo que ellos llaman la “estrategia de tabaco,” la campaña lanzada
a mediados de la década de 1950 por los fabricantes de cigarrillos para refutar
y ridiculizar las pruebas de la vinculación de consumo de tabaco y mortalidad.
Parecería que la estrategia del tabaco está
conectada a la negación de calentamiento global puramente por analogía:
otra industria poderosa, tratando de evitar la legislación, pero Oreskes y
Conway demuestran que esta estrategia de negación y manipulación ha sido
el modelo seguido después en la negación del cambio climático (Oreskes &
Conway, 2011).
Frederick Seitz,
por ejemplo, un ex Presidente de la Academia Nacional de Ciencias y miembro
nato del Comité Asesor científico del Presidente, en 1979 fue contratado por r.
j. Reynolds Tobacco Company. Camel lo colocó a la cabeza de su Comisión de
investigación médica. En el l proyecto Manhattan, le asignó a Seitz 45
millones de dólares en subvenciones de investigación para reforzar el prestigio
de tabaco — subvenciones que, como él más tarde admitiría, iban dirigidas a
evitar cualquier información negativa sobre el tabaco. “No quisimos mirar
los efectos de fumar cigarrillos en la salud,” dijo en una entrevista de 2006.
Seitz cobró cientos de miles de dólares durante los seis años durante los
cuales sirvió en este proyecto. No fue mucho tiempo después que él y un
conjunto de colegas comenzaron a negar la realidad del cambio climático de
origen humano (Oreskes &
Conway, 2011).
En 1984 Seitz,
Robert Jastrow y William Nierenberg fundaron el Instituto de Marshall George,
que supuso para el cambio climático lo que el Instituto del tabaco supuso
durante mucho tiempo para los cigarrillos. Seitz y sus colegas afirmaron que el
calentamiento global fue causado por las variaciones naturales en el flujo
solar, tal como la lluvia ácida fue causada por las erupciones volcánicas.
Sostuvieron que cualquier calentamiento provocado por las emisiones de efecto
invernadero es inundado por las variaciones climáticas naturales. El Instituto
de Marshall en su sitio Web aún hoy afirma que no hay consenso sobre el cambio
climático global, y que realmente puede haber efectos “beneficiosos del
aumento de CO2 en la atmósfera” (por ejemplo, la fantasía del
incremento de la productividad agrícola). A Seitz y sus cómplices, se unió el
físico, Fred Singer, y ganaron la atención de enormes medios de
comunicación y de periodistas sensacionalistas. Sus reivindicaciones
también encontraron en las administraciones de Reagan y Bush (Oreskes &
Conway, 2011).
¿Por qué Seitz y
compañía han sido tan beligerantes en la negación de la realidad del
cambio climático mundial? Oreskes y Conway muestran que el cambio climático era
realmente una frontera que los protegía de males mayores en un Estado
regulador — un Estado considerado cada vez más dispuesto a restringir las
libertades de libre mercado en nombre de la protección del medio ambiente. Para
contrarrestar la amenaza imaginada de un misil soviético, Seitz y su camarilla
defendió la iniciativa de defensa estratégica (guerra de las galaxias) del
Presidente Reagan, en un esfuerzo de millones de dólares para militarizar el espacio.
De hecho, el propósito original del Instituto Marshall era defender la línea
dura de Reagan. Cuando el imperio soviético se derrumbó en 1989, estos físicos
de la “guerra fría” se trasladaron a atacar a un nuevo enemigo, el
ecologismo, que según ellos promovía el mismo programa antiestadounidense.
El ecologismo (y más en particular las ciencias del clima) han sido
invocados como el más reciente de una larga línea de amenazas a la
libertad: “un árbol verde con raíces rojas,” como dijo el periodista
conservador George Will (Oreskes &
Conway, 2011).
Todos esto ayuda
a explicar por qué estos fundamentalistas de libre mercado, inmersos en
las oposiciones de la guerra fría (economías de mercado frente a
las economías planificadas, el individuo contra el Estado, el mundo libre
frente a Big Brother), atacaron todos
los esfuerzos para realizar un seguimiento de las enfermedades ambientales
producidas por las empresas de productos químicos. Fluorocarbonos clorados Noerna,
según ellos, los responsables del deterioro en la capa de ozono y los sulfatos
se Belched de las plantas de carbón no causaban lluvia ácida
que perjudicaba de bosque; el humo de los cigarrillos Ino no causaba
ningún daño demostrable. Oreskes y Conway muestran como Singer, Seitz y
un número de otros negacionistas del cambio climático sirvieron como
asesores en la Advancement of Sound Science Coalition, una
plataforma de la Philip Morris destinada a impugnar las pruebas que
vinculaban a los fumadores pasivos con el riego de padecer
enfermedades asociadas (Oreskes &
Conway, 2011).
24. Mecanismos de consumo de sustancias psicoactivas
|| Regresar al Índice ||
Los peligros de cada sustancia de abuso no
solo obedecen a la sustancia en sí, sino a los modos o mecanismos con los
cuales las dosis de sustancia son ingresadas al cuerpo humano, de las cuales
podemos señalar las siguientes (Ansel, Popovich, & Allen, 1995;
Martínez Ruiz & Rubio Valladolid, 2002).
24.1 Ingesta por aparato digestivo de solidos o
líquidos
En este caso la sustancia de abuso es consumida en una dilución líquida o semisólida. Por ejemplo, la cafeína se puede diluir en diferentes líquidos, que dependiendo de la composición secundaria denominamos café, té o bebidas energizantes. Los componentes activos de la marihuana también se pueden diluir en agua u otras sustancias alimenticias como la harina para crear galletas o pasteles.

Figura 24.1. Ingesta diluida. Uno
de los mecanismos más seguros para el consumo de sustancias es mediante la
elaboración de bebidas diluidas, la mayoría consumimos diariamente alcaloides
estimulantes como la cafeína a través del café y el té.
Los efectos del
consumo de sustancias, en este mecanismo por lo general es lento y con una
intensidad baja debido a que las sustancias deben atravesar los diferentes
mecanismos de filtrado de toxinas del cuerpo, especialmente el hígado. Sin
embargo algunas sustancias son capaces de saturar rápidamente la capacidad de
filtro del hígado como en el caso del alcohol de bebida (Ferré & O’Brien, 2011; Pesta,
Angadi, Burtscher, & Roberts, 2013).
24.2 Consumo de vapores de combustión
El principal
problema con la combustión es el hecho de que al quemar materia orgánica
compleja no solo se genera dióxido de carbono, sino muchos otros compuestos
orgánicos altamente carcinógenos como el alquitrán, los toluenos y diversos
óxidos de nitrógeno. Los vapores sin embargo son capaces de atravesar con mayor
facilidad las mucosas olfativas y digestivas, lo cual hace que el impacto o
efecto de la sustancia sea mayor y más rápido. El problema fundamental con este
mecanismo de dosificación es el cáncer, ya que las sustancias producto de la
combustión provocan o facilitan la mutación de las mucosas (Benowitz, 2008; Cecinato, Balducci,
& Nervegna, 2009; Maertens et al., 2009; Marselos & Karamanakos, 1999;
Tashkin, 1990, 2001).
El cigarrillo de nicotina es el más culpado, pero cualquier sustancia consumida por combustión generará los efectos carcinógenos, y de hecho un cigarro ilegal de marihuana tiene muchos más problemas dado que no posee los filtros, que están allí precisamente para intentar eliminar parcialmente los vapores tóxicos producto de la combustión de cualquier tipo de materia orgánica.

Figura 24.2. Vapores de combustión.
Cualquier vapor de combustión genera alquitrán y otros carcinógenos, aun cuando
lo que se quema sea “anticancerígeno” al quemarlo se hace cancerígeno.
24.3 Esnifado
Consiste en aspirar o inhalar la sustancia por la mucosa nasal, ya sea en polvo o gas no generado por combustión, para aprovechar así la membrana de las paredes mucosas nasales en su absorción y paso al torrente sanguíneo. Inhalar una sustancia implica un riesgo relativamente menor a inyectarla debido a que la droga logra entrar en menor cantidad y con menor velocidad. En el caso de los estimulantes,1 esta vía de administración se asocia generalmente con la capacidad de desarrollar un patrón de consumo compulsivo debido a su rápida acción y aparente seguridad frente a las inyecciones.

Figura 24.3. Esnifado.
Compartir los
utensilios para inhalar (popote, billete, etc.) puede tener las mismas
implicaciones que compartir una jeringa, pues ambos objetos pueden transportar
bacterias o virus como el VIH y Hepatitis C y, debido a la irritación de las
paredes nasales en donde se generan microlasceraciones por las cuales pueden
penetrar los patógenos. En el caso de los gases (óxido nitroso o gas de la risa
y los productos opiáceos modernos como el Fentanyl en forma de gas) o los líquidos
(cemento, resistol, gasolina, ether, la mona, nitritos en los poppers) se
utilizan globos, bolsas, estopas, trapos o frascos en los cuales se puede
transportar la sustancia y desde los cuales se inhalan las emanaciones gaseosas
(Bonomo &
Proimos, 2005; Neumark, Delva, & Anthony, 1998; Schütz, Chilcoat, &
Anthony, 1994; Young, Longstaffe, & Tenenbein, 1999).
24.4 Transdérmica
También llamada
transcutánea, esta vía consiste en aprovechar la permeabilidad de la piel para
absorber la sustancia simplemente por frotarla en la superfcie de esta. El uso
del LSD de esta manera puede ser peligroso porque es fácilmente absorbido y es
altamente activo desde dosis muy pequeñas (30 microgramos). Esta ruta también
se utiliza para tratamientos hormonales con cremas con esteroides y otras hormonas
(Granger &
Simon, 1992; Prausnitz & Langer, 2008; Shah, Maibach, & Jenner, 1993;
Tiffany, Cox, & Elash, 2000).
24.5 Rectal o vaginal
Con esta ruta se aprovechan las membranas
y los vasos sanguíneos del interior de estas zonas. Quien consume la sustancia
introduce la dosis al interior del recto o de la vagina y espera alrededor de
45-60 minutos para que se absorba completamente. Esta vía de administración es
similar en tiempo a la ingesta, pero al no haber jugos gástricos presentes, la
sustancia no se descompone ni se reduce su concentración por lo que el efecto
puede ser más intenso. Sin embargo muchas sustancias provocan irritaciones y en
ciertos casos pueden llegar a presentarse infecciones vaginales o en las vías
urinarias (Richardson & Illum, 1992).
24.6 Sublinguial
Que consiste en colocar la sustancia
debajo de la lengua y dejar que se absorba. En este lugar existen vías que transportan directamente la sustancia
absorbida al torrente sanguíneo por lo que su utilización es común para
diversos medicamentos pero también para usos tradicionales como el de la hoja
de coca y el tabaco mascado (Mei, Zhang, & Xiao, 2010).
24.7 Inyecciones
Es la más
peligrosa ya que cada vez que se rompe la barrera protectora de la piel, se
abre la puerta a patógenos externos al organismo que pueden provocar
infecciones, abscesos, coágulos y otras complicaciones fisiológicas. Además, es
la ruta de consumo más vinculada con la muerte por sobredosis (Bourgois & Schonberg, 2007;
Vitellone, 2015). Es necesario siempre usar equipo estéril y
cuidar mucho la limpieza utilizando algodón con alcohol para limpiar la zona
donde se aplicará la inyección antes y después (Chaisson et al.,
1989; Kerr, Tyndall, Li, Montaner, & Wood, 2005; Watters, Estilo, Clark,
& Lorvick, 1994). Es posible utilizar diferentes tipos de agujas y jeringas para llevar la
sustancia a distintos tejidos y profundidades. A continuación, se presenta una
clasificación de acuerdo al lugar en el que se libera la dosis inyectada:
👉 Subcutánea: Es la menos común y
consiste en utilizar una aguja muy delgada para atravesar únicamente las capas
superiores de la piel y liberar la sustancia en la capa de tejido graso que se
encuentra debajo de la piel, pero por encima del músculo. No hay mucha
irrigación sanguínea así que se absorbe lentamente. Sin embargo, no se pueden
utilizar volúmenes grandes ni líquidos muy viscosos.
👉 Intramuscular: Se utiliza una aguja mucho más gruesa y larga para atravesar todas las
capas de la piel y el tejido graso y entrar en el músculo. También tiene una
absorción lenta, pero se puede introducir un mayor volumen, el líquido
inyectado puede ser más denso y el movimiento del músculo puede ayudar a que se
absorba más rápido.
👉 Intravenosa: Se trata del tipo de inyección más riesgoso tanto por el impacto de la sustancia como por la velocidad de su efecto. Para lograr una inyección intravenosa se utilizan agujas muy delgadas (generalmente de insulina) para perforar la pared de una vena e introducir la sustancia directamente en el torrente sanguíneo. Para ello, es necesario filtrar muy bien el líquido y tener mucho cuidado con la dosis ya que esta es la vía que logra llevar la sustancia al cerebro en el menor tiempo.

Figura 24.4. Adicto al
crocodile con lesiones por inyecciones insalubres.
25. Relevancia social de la adicción en Colombia
|| Regresar al Índice ||
La mayoría de las
personas no usa drogas ilícitas y entre quienes llegan a probarlas, sólo una
fracción desarrollará patrones de dependencia. Sin embargo, el consumo abusivo
de drogas constituye un serio problema de salud para muchas personas en las
Américas. Es particularmente importante destacar que el inicio temprano del
consumo está asociado con un mayor riesgo de dependencia y de otros problemas.
Uno de los principales desafíos es prevenir el uso y retrasar el inicio del
consumo entre los jóvenes (Buvinic, Morrison,
& Orlando, 2005; Concha, 2002; Hopenhayn, 1997; Peruaga, Rincón, &
Selin, 2002).
El consumo de drogas, incluyendo el alcohol, produce mayores niveles de mortalidad y discapacidad en el Hemisferio que en el resto del mundo. Es un factor de riesgo importante en el caso de sesenta enfermedades y lesiones asociadas a accidentes y violencia. El consumo de drogas en el Hemisferio es muy variable entre los países, tanto en términos de la magnitud del uso, como del tipo de sustancia. Esta variabilidad genera importantes interrogantes, tales como si el Hemisferio está frente a un solo problema de drogas o a diferentes problemas y, consecuentemente, cuál o cuáles deberían ser las mejores políticas para enfrentarlos (Buvinic et al., 2005; Concha, 2002; Hopenhayn, 1997; Peruaga et al., 2002).

Figura 25.1. Sin hogar. La
expresión políticamente correcta para llamarlos es ‘habitantes de calle’, pero
lo usual es que les digan indigentes y, cómo no, ‘desechables’. Desechables
para la mayoría, menos para mafias locales que tienen amarrado a ellos un
millonario mercado y, a la vez, mano de obra barata para sus negocios ilegales.
La marihuana es
la droga ilícita de mayor consumo en el mundo, y uno de cuatro usuarios se
encuentra en el continente americano. Si bien es una droga asociada con menor
mortalidad que muchas otras sustancias, la marihuana presenta riesgos para la
salud, especialmente para los usuarios adolescentes cuyos cerebros están aún en
desarrollo El consumo de la cocaína está en aumento en algunas regiones del
Hemisferio. Sus formas fumables se han constituido en un problema de salud
pública en varios países de América del Sur Argentina, Brasil, Chile y
Uruguay), particularmente cuando se trata de los grupos vulnerables marginados.
El consumo de heroína es mucho menos común en América Latina y el Caribe, y la
mayoría del consumo se concentra en Estados Unidos, Canadá y México. Sin
embargo, se está convirtiendo en tema de preocupación para otros países, como
Colombia y la República Dominicana, en los cuales debieran buscarse las
alternativas sanitarias que han probado ser más eficaces en países donde el
problema tiene una historia más larga (Buvinic et al.,
2005; Concha, 2002; Hopenhayn, 1997; Peruaga et al., 2002).
Drogas sintéticas como los estimulantes de tipo anfetamínico tienen una alta prevalencia de consumo en Canadá y Estados Unidos. El consumo de sustancias tipo éxtasis es cada vez más común en la población joven de muchos otros países del Hemisferio. El consumo de alcohol por estudiantes secundarios es un problema en todo el Hemisferio, y el consumo excesivo y compulsivo es de particular preocupación. Aun cuando el consumo de alcohol sea legal para los adultos, el uso de esta sustancia en menores corresponde a uso nocivo. Según estudios epidemiológicos realizados en el Caribe y América del Sur, la prevalencia de consumo en el último mes en estudiantes secundarios es superior al 50% en algunos países. En cuanto a la respuesta, se debe considerar que los programas de prevención escolares o de persuasión deben acompañarse con otras medidas que regulen la disponibilidad y que vinculen a las familias o establezcan lazos con políticas comunitarias. Todo lo anterior implica que los países deben priorizar esta estrategia destinando los recursos necesarios para su adecuada implementación y evaluación (Buvinic et al., 2005; Concha, 2002; Hopenhayn, 1997; Peruaga et al., 2002).

Figura 25.2. Sin hogar 2.
Ninguno de ellos pensó que terminaría así, no se trata de un defecto de la
personalidad que se pueda controlar, esto es una consecuencia de desligar al yo
de la química del cerebro que tiene nuestra sociedad occidental, nuestro yo es
la química de nuestro cerebro, alterarla, nos afecta profundamente.
Por otro lado, el
tratamiento de problemas relacionados con el uso de sustancias debe ser parte
de un proceso continuo, que involucre todos los niveles de la red asistencial,
con especial énfasis en la detección temprana e intervenciones breves en el
primer nivel de atención. Las intervenciones deben contar con base científica y
efectividad probada. Deben estar a cargo de personal calificado y cumplir con
estándares de calidad. La salud mental es un importante factor de riesgo para
el desarrollo de dependencia de las drogas. Sin embargo, muchos países carecen
de servicios adecuados o suficientes recursos humanos en este ámbito de la
salud. Es importante, por lo tanto, reconocer que estamos frente a una
enfermedad crónica que debe ser tratada como tal, con pleno compromiso de las
estructuras sanitarias y respeto por los derechos de los pacientes. Para la
implementación de políticas eficaces de salud pública y su correspondiente
evaluación, se requiere información actualizada, en cantidad necesaria y de
calidad. Sin embargo, son pocos los países que invierten en este ámbito. Es preciso
que se fortalezcan los sistemas de información que permitan hacer un debido
monitoreo del problema de las drogas y también que se financie la investigación
para abordar el problema de manera más eficaz; del mismo modo, se debe invertir
en capital humano y en infraestructura (Buvinic et al.,
2005; Concha, 2002; Hopenhayn, 1997; Peruaga et al., 2002).
En Estados Unidos
y Canadá, los derivados de opioides (empleados principalmente como
analgésicos), los tranquilizantes y sedantes (especialmente las
benzodiacepinas) y los estimulantes (como el metilfenidato o la
dextroanfetamina) son los fármacos de mayor uso indebido. En el resto de los
países, esta situación es menos clara, en parte por una ausencia de información
que describa el fenómeno correctamente (Buvinic et al.,
2005; Concha, 2002; Hopenhayn, 1997; Peruaga et al., 2002).
25.1 Impacto en Bogotá Colombia
Los colombianos nos los encontramos a
diario en cualquier calle, y pocos podrían decir sin mentir, que, cuando menos,
su presencia no los molesta. Su número se ha multiplicado en las últimas
décadas, amarrado al aumento del consumo interno de droga, pero los ojos del
país solo se vinieron a fijar en lo que representan en mayo pasado, cuando la
toma de las autoridades al sector del ‘Bronx’, en Bogotá, mostró el oscuro
mundo al que están sometidos. La expresión políticamente correcta para
llamarlos es ‘habitantes de calle’, pero lo usual es que les digan indigentes
y, cómo no, ‘desechables’. Desechables para la mayoría, menos para mafias
locales que tienen amarrado a ellos un millonario mercado y, a la vez, mano de
obra barata para sus negocios ilegales (Justicia, 2016).
Censos que las fuentes en todo el país
califican como precarios hablan de al menos 40.000 personas en condición de
indigencia extrema. Representan, en conjunto, una población superior a la del
casco urbano de Leticia, la capital del Amazonas, y más allá de las apariencias
gira a su alrededor una danza de millones de pesos. (Lea también: Fiscalía
confirma nuevo hallazgo de restos humanos en el 'Bronx'). La mayoría de ellos
puede caminar 7 kilómetros a diario –escarbando en la basura en busca de
reciclaje– y aguantar un día sin mayor bocado, pero no resiste más de tres
horas sin el humo de una ‘bicha’ o una ‘pipa’ de PVC con bazuco, la droga más
barata y más contaminada de las calles (Justicia, 2016).
Son 40.000 potenciales compradores de todo tipo de alucinógenos que mueven un mercado que se tasa por ‘bombas’ (10 dosis de bazuco) que se compran en las ollas a 10.000 pesos; bolsas de ‘sacol’ (pegante) a $ 500 o ‘diablos’ (mezcla de bazuco y marihuana) que se consiguen por 2.000 pesos. Una dosis se puede pagar con monedas, pero cuando se suman las que les venden a diario la cuenta se hace en cientos de millones de pesos. Habitantes de calle que hablaron con EL TIEMPO coincidieron en que consumen un promedio de 10 ‘bichas’ de bazuco al día, pero hay quienes llegan a las 25. Esa es la razón por la que en todas las ciudades las bandas se disputan a muerte ese mercado cautivo de la indigencia (Justicia, 2016).

Figura 25.3. Sin hogar 3. Se
ha demostrado que aquellos individuos más propensos en desarrollar la adicción
son aquellos con déficit de atención innatos, que comúnmente son personas que
no logran concentrarse en actividades normales, por lo que son excluidos de la
sociedad por sus comportamientos impulsivos, esta impulsividad es causada por
una búsqueda de lograr el equilibrio de dopamina en el cerebro, que en ellos es
naturalmente más baja, de allí que caigan más rápido en la adicción.
En Medellín, por ejemplo, los ‘combos’
ganan por lado y lado. Así, les cobran vacuna por dejarlos dormir en las aceras
(entre 500 y 800 pesos diarios), pero también les prohíben comprar a redes que
no sean las suyas. Si se tiene en cuenta que en esa capital hay unos 3.500
habitantes de calle y que, según las autoridades, el 95 por ciento tiene
problemas de adicción, ese negocio movería más de 30 millones de pesos al día.
Y en todo el país la cifra podría estar entre los $ 300 y $ 400 millones
diarios, tan solo en el target de los que, supuestamente, no tienen cómo pagar
por nada. La plata que puede recibir en un día normal alguien de la calle
sorprende, y eso hablando de actividades legales. Así, una noche de reciclaje
puede dejar ingresos por 60.000 pesos; cuidar carros, entre 20.000 y 30.000; y
pedir limosna, entre 70.000 y 150.000 pesos en las esquinas más concurridas de
las grandes ciudades (Justicia, 2016).
Los golpes de las autoridades contra las
ollas más grandes del país, empezando por el ‘Bronx’, tienen patas arriba el
mercado de los narcóticos. Así, de la venta abierta que se realizaba en algunas
calles dominadas por el hampa se ha pasado a situaciones en las que los dueños
de la droga, usualmente bandas locales, tienen que salir a buscar a sus
clientes. (Además: Lo que viene después de seis meses de la intervención al
'Bronx'). El secretario de Seguridad de Medellín, Gustavo Villegas, dice que
hoy en esa ciudad el 60 por ciento de los expendedores son ambulantes y que,
literalmente, persiguen a sus antiguos clientes para mantener su negocio boyante
(Justicia, 2016).
En Bogotá, la primera estrategia que
usaron los ‘ganchos’ –capos de las ollas– fue utilizarlos para generar una
crisis de seguridad en sectores céntricos y forzar a la Alcaldía Mayor y a la
Policía a desistir del plan de demoler el ‘Bronx’. No lo lograron, pero siguen
acechando. Algunos de los habitantes de calle que decidieron acogerse a los
programas de recuperación que el Distrito abrió simultáneamente con la
intervención aseguran que han visto cerca de los centros a jíbaros que incluso
les dan dosis gratis (Justicia, 2016).
Y no se trata solo de mantener la venta de
alucinógenos. En todo el país, ellos son usados para transportar todo tipo de
sustancias ilegales, empezando por la droga. “Les pagan por llevar cargamentos
de un sitio a otro, pero también les pagan con dosis por lo que han robado.
Parte de ese pago lo revenden en los barrios y ese es uno de los factores que
han disparado el microtráfico en el país”, dice una alta fuente de la Fiscalía.
También los están usando como ‘campaneros’ y hasta les dan celulares para que
avisen de cualquier movimiento inusual en los barrios. “Para no ‘boletearse’
tanto e intentar manejar un bajo perfil, las bandas les entregan pequeños
paquetes de droga que ocultan entre sus prendas y sacos para transportar a pie
por extensos recorridos, reciben diariamente dosis mínimas de droga que los
mantienen en el vicio”, insistió una fuente policial en Bucaramanga. Rubén
Darío, un hombre de 31 años que ha vivido casi toda su vida en la calle y que
se define como ‘lujero’ (así les dicen a los que se roban partes de carros), lo
explica así: “A uno de ‘ñero’ nadie se le arrima, ni la Policía” (Justicia, 2016).
El transporte de droga no deja de ser para
ellos una actividad riesgosa. No tanto por la posibilidad de que sean detenidos,
sino porque los dueños de la carga no aceptan excusas y si no la entregan
tienen que pagarla de alguna manera. El pago puede ser un asalto, llevar la
carta para una extorsión o incluso un asesinato. Fernando, un antiguo sicario
del sector de El Cartucho en Bogotá que terminó en la indigencia, dice que su
‘negocio’ se dañó en parte porque a los habitantes de calle los usan también
para atentados y ajustes de cuentas, y que les pagan entre 100.000 y 200.000
pesos por “hacer un trabajo por el que antes se cobraban 15 millones”: “Los
buscan porque vestidos así, mugrosos, pueden pasar cerca de la gente. Los
contratan para echarle pegante a alguien en el pelo o para pegarle una
‘cascada’ ” (Justicia, 2016).
Una encuesta del Distrito del 2015 les
preguntó a 4.000 habitantes de calle por situaciones de violencia en las que
hubieran estado involucrados. El 52 por ciento admitió haber cometido hurtos y
atracos; el 49,41 %, lesiones personales; el 28,54 %, heridas graves; el 25,08
%, estafas; el 22,56 %, amenazas; un 8,52 % homicidios; un 7,48 %, cobro de
vacunas; un 2,2, violaciones, y el 1,34 por ciento, secuestros. En Bucaramanga,
‘Muelón’, quien hasta hace tres meses deambulaba por las calles del norte de la
ciudad, le dijo a EL TIEMPO que perdió la cuenta del número de personas que
robó durante 25 años. Este hombre de 53 años de edad, de tez delgada, con la
dentadura incompleta y con numerosas cicatrices producto de heridas con navajas
y picos de botella, narró que con cuchillo en mano y hasta con revólver asaltó
a muchos transeúntes desprevenidos. En su haber registra 17 entradas a la
cárcel Modelo, 2 a la cárcel del mismo nombre en Bogotá y 2 más a un reclusorio
de Barranquilla. “Sé de muchos que como yo siguen en la calle y roban todos los
días para comprar droga, porque la comida se consigue fácil con la gente y
fundaciones que ayudan a los que como yo cayeron en el mundo de las calles”,
asegura (Justicia, 2016).
La delincuencia asociada al consumo de
drogas tiene en esta población índices mucho más altos que en otros grupos de
adictos. Este factor es clave para explicar la millonaria rentabilidad que deja
hoy el microtráfico, hasta niveles del 2.400 por ciento en algunas ciudades,
según un reciente estudio de Planeación Nacional. La razón es sencilla: las
bandas que venden la droga les reciben a los habitantes de calle, a precios
irrisorios, lo que traigan, desde material reciclado hasta objetos robados de
todo tipo. En Bogotá, aunque las ollas más famosas son las de El Cartucho, el
‘Bronx’ y San Bernardo, hay otras no menos activas. En el extremo occidente de
la ciudad, cerca del humedal de Jaboque, hay una que tiene años y a dónde
llegan los que tienen algo que canjear por droga. (También: A bala, mafias del
'Bronx' se iban a tomar San Bernardo) “Allí cambiábamos lo que lleváramos por
‘bichas’: desde unos zapatos ‘sanos’, un pantalón, herramientas, hasta el lujo
de un carro. Cualquier cosa que se pudiera vender servía para la ‘soplada’. Por
unos ‘pisos’ completos –así le llama a las zapatillas– me daban 5.000 pesos”,
le dijo a este diario un habitante de calle. En ese lugar, a dónde aún no
entran las autoridades, hay dos grandes ‘ganchos’. Uno lo lideran dos mujeres y
el otro es llamado ‘los Paisas’. Allá, dice la fuente, sigue habiendo casas
donde “dan fuego” (ofrecen la droga y el sitio para consumirla) (Justicia, 2016).
La historia se repite en todas las
ciudades, y tiene temporadas. En medio de su adicción, los habitantes de calle
terminan convertidos en una especie de zombis del delito. “En su mayoría son
personas enfermas, consumidoras. Eso los hace proclives a que se aprovechen de
ellos”, dice el general Nelson Ramírez, comandante de la Policía Metropolitana
de Cali. En esa capital las bandas organizadas que se dedican al robo de
cableado y tapas de alcantarilla los subcontratan para que arriesguen su vida
por esos elementos, a cambio de unos pocos pesos o de algunas papeletas. Eso le
pasó a un reciclador que murió electrocutado cuando intentaba hurtar cable de
una red pública en noviembre pasado. Como las autoridades de todo el país han
tomado medidas para proteger ese material y han terminado por cambiar las tapas
de alcantarilla por otras no metálica, ahora la modalidad que usan es robarse
las rejas de antejardines (Justicia, 2016).
En Cali, la Policía ha detenido en lo que
va de este 2016 por robo de cableado a 56 personas. El año pasado fueron 64. De
ese número, más del 90 por ciento corresponde a miembros de bandas
delincuenciales. Los habitantes de calle figuran en una pequeña proporción y lo
hacen, sobre todo, para tener dinero para el consumo. Esas bandas, según la
Policía, los contactan para identificar los sitios para posibles hurtos y en
otros casos, para ir sacando pedazos de material, como ocurrió el mes pasado
con la destrucción del monumento de Héctor Lombana, hecho en cuarzo y pizarra,
en plena avenida sexta. En la capital del Valle el robo de tapas de
alcantarillas se viene frenando porque se están reemplazando por las de corcho
y quitar alguna requiere herramientas específicas. Los mayores robos ocurrieron
en el 2011, con pérdidas de 2.000 unidades por valor de 500 millones de pesos.
El negocio resultaba atractivo porque el kilo costaba en el mercado clandestino
alrededor de $ 6.500; cuando lo compran las fundidoras supera los 30.000 y
40.000 pesos. Para Emcali, la reposición de una sola tapa costaba hasta 330.000
(Justicia, 2016).
Desarticular esas organizaciones que se
aprovechan de su adicción es una de las claves para empezar a controlar el
creciente problema de la indigencia en el país. Pero, agrega la secretaria de Integración
Social de Bogotá, María Consuelo Araújo, esa es la mitad de la tarea. Asegura
que hay que persistir en los programas tanto para sacarlos de la droga como
para ofrecerles verdaderas oportunidades para recuperar sus familias, su
trabajo y la dignidad. (También: Capturan a integrantes de mafia del 'Bronx'
con 10.000 dosis de bazuco) De 2.050 personas que salieron del ‘Bronx’ y fueron
atendidos en el operativo conjunto de la Alcaldía Mayor, Bienestar Familiar,
Fiscalía, Policía y la Secretaría de Seguridad, hay 550 que se mantienen en el
proceso de rehabilitación. “En la medida que no nos fortalezcamos en la
prevención en el consumo de drogas no vamos a hacer nada”, advierte (Justicia, 2016).
25.2 Los colegios colombianos
El 72 por ciento de los estudiantes de
seis colegios públicos de Bogotá consultados en medio de una investigación de
la Universidad de La Sabana ha consumido o consume alcohol; el 43 % fuma
cigarrillo; el 11 % marihuana; el 7 % aspira inhalantes; el 6 %, ácido LSD, y
el 4% inhala cocaína, prueba éxtasis o consume bazuco. Las alarmantes
estadísticas fueron reveladas por 1.600 estudiantes, hombres y mujeres, de
sexto a once de bachillerato, entrevistados por Ángela Trujillo y Diana Obando,
profesoras de la Facultad de Psicología de la Universidad de la Sabana, en un
estudio que además evidencia que algunas conductas antisociales se relacionan
directamente con el consumo de estupefacientes. Esta investigación,
desarrollada en 2014 y conocida en exclusiva por EL TIEMPO, se da a conocer una
semana después de que Santiago Isaac Sánchez, un joven de 14 años, perdiera la vida al consumir una mezcla de droga,
licor y polvo de extintor en las instalaciones del colegio distrital Marco
Fidel Suárez, en el barrio El Tunal (ELTIEMPO.COM, 2015; López-Quintero
& Neumark, 2012; Mejía & Gómez, 2005; Wiesner & Peñaranda, 2002)
El incidente, que dejó intoxicados a otros
21 alumnos, prendió las alarmas sobre la realidad del consumo de drogas en los
jóvenes estudiantes y las redes delincuenciales de microtráfico que acechan a los colegios
públicos. Según la investigación de la Universidad de La Sabana, a
los 14 años, la misma edad de Santiago Isaac, los estudiantes de colegios
públicos de Bogotá inician a
experimentar con la marihuana y el alcohol, las dos sustancias que
más consumen los niños y adolescentes junto a los ácidos, la cocaína, el
éxtasis y el bazuco (ELTIEMPO.COM, 2015; López-Quintero
& Neumark, 2012; Mejía & Gómez, 2005; Wiesner & Peñaranda, 2002).
En su orden y en números, el estudio
indica que las sustancias más comunes son el licor (53%); el cigarrillo (14%);
la marihuana (6%); los inhalantes (bóxer, dick, productos de limpieza, entre
otros); el ácido LSD (4%), y el bazuco, el éxtasis y la cocaína (3%). Estas
cifras hacen referencia al consumo que habían hecho los entrevistados en los
últimos 30 días a partir del momento en que se realizó la encuesta”, dijeron
las investigadoras. Según la investigación, los niños no solo consumen por
primera vez alcohol entre los 12 y 13 años, sino que comienzan a hacerlo de
manera permanente, por lo menos una vez a la semana, a partir de los 14. En el
estudio se evidencia que el cannabis es la droga de más fácil acceso dentro y
fuera de la institución educativa”, advierten Obando y Trujillo (ELTIEMPO.COM, 2015; López-Quintero
& Neumark, 2012; Mejía & Gómez, 2005; Wiesner & Peñaranda, 2002)
La investigación de la Sabana muestra
claramente que el consumo de alcohol y drogas es la puerta de entrada para los
jóvenes a conductas delictivas como conflictos, robos, agresiones y arrestos.
Las expertas afirmaron que fumar, tomar trago y probar otras sustancias
psicoactivas tiene una relación directa con la presencia de comportamientos
antisociales. “Especialmente se encontró que el consumo de marihuana está
relacionado con la conducta de pertenecer a una pandilla”, dijeron. El 31 % de
los consultados señaló que ha participado en el robo simple a tiendas o
supermercados. Y de ahí en adelante, confesaron otros delitos: daño a propiedad
ajena como hacer grafitis, pinchar un carro o romper una ventana (22 %); haber
sido suspendido del colegio (16 %); agresiones a otras personas (14 %); robos
de objetos de mayor valor (14 %); haber sido arrestado (9 %); pertenecer a una
pandilla (7 %), y vender drogas ilegales (5%) (ELTIEMPO.COM, 2015; López-Quintero
& Neumark, 2012; Mejía & Gómez, 2005; Wiesner & Peñaranda, 2002)
De acuerdo con la investigación, el
principal factor de riesgo que se asocia con el consumo de drogas y el
desarrollo de conductas antisociales es el desinterés que tienen los padres en
saber en dónde y en qué andan sus hijos. En ese sentido, el 50 % de los jóvenes
respondió que a sus familiares no les interesa sus actividades o se muestran
permisivos ante las acciones que realizan, así estas sean ilegales o pongan en
riesgo su integridad y seguridad. Ante esto, el estudio plantea a las
autoridades soluciones que aborden de manera integral todos los ambientes en
los que los jóvenes y niños conviven. Concretamente, piden implementar planes
de prevención y de intervención que involucren a estudiantes, instituciones
educativas y padres de familia que impacten a largo plazo. (ELTIEMPO.COM, 2015; López-Quintero
& Neumark, 2012; Mejía & Gómez, 2005; Wiesner & Peñaranda, 2002)
25.3 Otros comportamientos adictivos
Dado que el cuerpo produce sus propias
sustancias adictivas que dependen de los sentidos y del comportamiento, es
particularmente evidente que pueden existir otros mecanismos que generan
adicciones. Los comportamientos adictivos se relacionan con actividades,
objetos, comportamientos o sustancias que generalmente no se considerarían
adictivos. La adicción se señala en el momento en que dichos comportamientos
empiezan a obstruir los lazos sociales y las responsabilidades cotidianas, o
hacen daño en nuestra anatomía y fisiología.
Por lo general los comportamientos
adictivos crean una dependencia psicológica de forma exclusiva pero no
necesariamente anhedonia, a diferencia de las sustancias adictivas que crean
una dependencia tanto psicológica como fisiológica. Sin embargo, de los dos
mecanismos de dependencia, la psicológica es la más difícil de curar ya que sus
causas son el inicio mismo del comportamiento adictivo. Las adicciones se consideran como un tipo de
compulsión, es decir, comportamientos sobre los que se tiene poco o ningún
control, pero caracterizado por refuerzos principalmente positivos “dependencia
psicológica” y en algunos casos negativos “síndrome de abstinencia”. Las
compulsiones en general se realizan sin tener en cuenta la recompensa o
negatividad del comportamiento. La compulsión relacionada al síndrome
obsesivo-compulsivo y las adicciones comparten rutas nerviosas comunes ubicadas
en los circuitos de recompensa del cerebro, señalados de forma concreta en el
núcleo Accumbens.
26. Adicción al alcohol
|| Regresar al Índice ||
En química orgánica se describe una cantidad cuasi-infinita de sustancias denominadas alcoholes, sin embargo, para el el neurotransmisor glutamato hay dos moléculas importantes, las cuales son también las más simples, el etanol y el metanol. El etanol es el alcohol más ligero y simple, también es el más parecido al agua, por lo que el cuerpo tiende a confundirlo con esta, puede atravesar fácilmente la membrana celular e interferir en las reacciones metabólicas que involucran al agua, que son casi todas, por esa razón sus efectos tóxicos son rápidos. También es más fácil de producirlo, por lo que es una buena manera que producir dinero, matando a tus consumidores, en Colombia lo denominamos alcohol adulterado, un peligro constante en las navidades.

Figura 26.1. Bebidas alcohólicas. Las
bebidas alcohólicas se pueden obtener de cualquier cosa que tenga azúcar
fermentable, pero la destilación artesanal tiende a concentrar metanol que es
tóxico (YouTube).
La segunda
molécula es el alcohol de dos carbonos, al ser más pesado y más grande, el cuerpo
no lo interpreta como el agua, aun así, posee efectos tóxicos y de generación
de dependencia. Este alcohol de dos carbonos es llamado metanol, siendo el
agente activo de todas las bebidas alcohólicas de recreación. Nunca bebemos
alcohol puro al 100%, eso sería peligroso “el ron al 80% podría tumbar a un
hombre adulto con pocos sorbos”, por lo general se lo consume diluido en agua y
otras moléculas orgánicas que dan sabor, textura y color. La razón para esto no
es médica, sino de productividad.
Originalmente el
alcohol de bebida se producía como el subproducto de la fermentación
alcohólica, es un desecho del metabolismo de las bacterias, y en consecuencia
es tóxico para ellas. Normalmente las levaduras o bacterias alcohólicas se
mueren cuando el porcentaje de alcohol alcanza entre un 15% a un 20%. Para
obtener bebidas más potentes hay que destilar, es decir calentar la mezcla y
recolectar el gas, el alcohol se evapora a poco menos de 65°C, junto con trazas
de agua en el goteo; o empleando filtros que retienen el vapor de alcohol y
eliminan el agua “generalmente barriles de madera”. La destilación puede
producir bebidas hasta un 80% como es el caso del ron, en este punto se emplea
agua purificada para rebajarlo a un 40%, porcentaje aproximado de todas las bebidas
destiladas como el whisky o el aguardiente.
El efecto del
alcohol varía de persona a persona dependiendo del tipo de bebida alcohólica
consumida, de la cantidad y el tiempo de bebida, de otras sustancias
consumidas, en especial las de alto contenido en grasa y del metabolismo propio
de los individuos. Un ejemplo es el Sake, los japoneses tienen un pobre
metabolismo para el alcohol, por lo que muchos pueden morirse bebiéndolo aun
cuando se trata de una bebida fermentada con un contenido de alcohol que
generalmente se ajusta a un 15%, que es semejante a una cerveza.
26.1 Absorción
del alcohol
(YouTube) Los efectos tóxicos del alcohol comienzan cuando
este es absorbido por el sistema digestivo y es depositado en el sistema
circulatorio. A diferencia de los nutrientes normales, el alcohol puede ser
absorbido directamente en el estómago, casi el 20% del alcohol puede ser
absorbido allí, mientras que el restante es absorbido principalmente en el
jejuno.
La tasa de absorción depende principalmente de la concentración de la bebida, las bebidas fermentadas y/o diluidas se absorben con mayor lentitud que las bebidas alcohólicas destiladas puras como el aguardiente o el vodka. Corroborando un mito popular, la tasa de absorción del alcohol disminuye si el estómago está lleno justo antes de la ingesta, eso se debe a que el alimento diluye el alcohol, y a que la grasa de la comida puede impermeabilizar parcialmente la pared gastrointestinal, impermeabilizándola al alcohol. Decimos parcialmente debido a que el metanol posee cierto nivel de permeabilidad ante los lípidos. Debido a que el alcohol puede ser absorbido por el estómago, algo que no hacen la mayoría de los nutrientes o toxinas, sus efectos se pueden manifestar relativamente rápido después del inicio de su ingesta, lo cual usualmente son 15 minutos.

Figura 26.2. La borrachera. La
ebriedad puede verse como un mecanismo de defensa del cuerpo para evitar la
muerte por sobredosis.
26.2 Distribución del alcohol y la borrachera
(YouTube) El metanol al ser soluble en agua y en grasas puede afectar a cualquier órgano del cuerpo, y atravesar la barra hematoencefálica. Por lo general las drogas de abuso se clasifican como estimulantes o depresoras, sin embargo, muchas personas piensan que el alcohol es un estimulante del sistema nervioso central. En la realidad tratamos ante una sustancia con efectos típicos de depresión del sistema nervioso central, especialmente afectando las regiones corticales prefrontales encargadas del pensamiento lógico, de la corteza motora encargada de los movimientos consientes, de la corteza sensorial, encargada de la sensibilidad, y del cerebelo en sus regiones motoras y del equilibrio. En consecuencia, el alcohol deprime las neuronas que inhiben comportamientos, permitiendo que las neuronas exitatorias tomar control absoluto, creando el efecto desinhibitorio que muchas personas confunden por estimulante, sin tomar en cuenta la obstrucción motora y sensorial que genera.

Figura 26.3. Mecanismo de acción. El
alcohol es un depresor, que limita el efecto de los neurotransmisores
depresores, por lo que genera, inicialmente la ilusión de ser un estimulante (YouTube)
(YouTube).
En general, el
alcohol restringe a la corteza cerebral, afectando las regiones sensoriales
consientes, la memoria consiente y el movimiento consiente, hasta que finalmente
toda la zona conciencie del cerebro es inhibida. Cabe anotar que debido a que
las zonas emocionales del cerebro no se ubican en la corteza, estas quedan
libres del control de la conciencia. Al aumentar la concentración del alcohol,
no solo la corteza comienza a ser inhibida, el tallo y la zona límbica pueden
verse afectados, lo cual incluye los circuitos parasimpáticos que controlan la
respiración y el ritmo cardíaco. Esto puede provocar la muerte por ingesta de
alcohol también denominada intoxicación por alcohol.
La intoxicación
por alcohol es evadida gracias al efecto sedante del mismo, al final el cuerpo
en su corteza motora está tan sedado que no es capaz de coordinar el movimiento
de una copa hacia la boca. A pesar de esto las personas se pueden morir debido
a un desarrollo de tolerancia, al consumo de drogas que si estimulan el sistema
nervioso o a que carecen de enzimas encargadas de la metabolización del
alcohol.
26.3 Efectos neuronales del alcohol
El alcohol es una molécula de abuso bastante extraña debido a que afecta a dos neurotransmisores que no están implicados directamente en el placer o la felicidad, como si lo hacen otras sustancias como los alucinógenos, los opioides o los canabinoides (McBride, Murphy, Lumeng, & Li, 1990; Morrow, Montpied, Lingford-Hughes, & Paul, 1990; Roberto, Madamba, Moore, Tallent, & Siggins, 2003).

Figura 26.4. El trago estándar. Un
trago estándar representa la cantidad de alcohol que el cuerpo de un hombre
adulto puede metaboliar en una hora sin que este afecte las habilidades
cognitivas, asumiendo una estatura de casi 1,8 metros, bajo índice de grasa
corporal y un hígado sano.
El alcohol afecta
los neurotransmisores relacionados con las marchas del cerebro. Los dos
neurotransmisores involucrados en esta historia son GABA y Glutamato. El
glutamato es el neurotransmisor que controla los cambios de marcha hacia la
aceleración de los procesos cognitivos, si hiciéramos una analogía, muchos
procesos del cerebro funcionan como un motor de dos cambios, la primera
velocidad se encuentra en la corteza cerebral, y la segunda velocidad se
encuentra en el cerebelo. Por ejemplo, el movimiento cambia de consiente y
burdo a fluido y concentrado a medida que su control pasa de la corteza motora
a la región motora del cerebelo. El glutamato regula estas transiciones como si
fuera un embrague, permitiendo pasar de la primera velocidad a la segunda.
Por otro lado, el
GABA funciona como el freno de la corteza motora y otras regiones del cerebelo,
el etanol es capaz de imitar estructuralmente al GABA uniéndose a los
receptores de este, por lo que frena de forma directa la actividad
neuronal al activar canales ionicos de cloro que hiperpolarizan la neurona. El
alcohol estimula en la corteza al GABA y bloquea al glutamato, por lo que la
corteza se encuentra incapaz de
👉 funcionar de forma consiente,
👉 pasar a funciones inconscientes de alto grado de
control en el hipotálamo y
👉 bloqueada por la acción del GABA.
Una vez bloqueado el cerebro no puede pasar de las funciones consientes a las funciones fluidas inconscientes, con lo que se pierden las habilidades finas en los aspectos motriz, y cognitivo “lo cual incluye hablar de forma asertiva y elocuente”. No todo el cerebro sufre estos problemas, las regiones límbicas relacionadas con las emociones se afectan mucho más lentamente, lo cual crea el efecto desinhibitorio típico de la intoxicación por etanol.

Figura 26.5. Degradación de etanol a etanal.
Degradación de etanol a etanal, es la típica oxidación de un alcohol primario a
un aldehído por deshidrogenación.
26.4 Excreción del alcohol del cuerpo humano
El alcohol es una toxina metabólica, siendo el principal producto de finalización y excreción de la fermentación alcohólica. En consecuencia, se trata de una molécula de muy baja energía que es capaz de bloquear las rutas metabólicas de la glucolisis, lo cual puede ser mortal ya sea para una colonia de bacterias fermentadoras o para los tejidos de un órgano humano. Como en todas las toxinas ingeridas por el ser humano corresponde al hígado su metabolización para no afectar a la sangre y los tejidos que esta alimenta. El hígado tiene una eficacia del 95% en la descomposición del alcohol que es degradado a dióxido de carbono y agua, pero su unidad de tiempo es patética, aproximándose a una copa estándar por hora.

Figura 26.6. Segunda oxidación.
Estas dos oxidaciones son típicas de los alcoholes primarios, primero a etanal
y luego a ácido etanoico, el cual puede integrarse de manera segura al
metabolismo (YouTube).
Por lo general
las bebidas alcohólicas se sirven en copas estándar, entre mayor es el
contenido alcohólico más pequeña es la porción de cada copa –compare el tamaño
de una copa de cerveza y una copa de Whisky –de forma tal que la ingesta de
alcohol alcance una cantidad estándar o copa estándar. En otras palabras, solo
la primera copa es metabolizada durante la primera hora, y evidentemente
bebemos más de una copa por hora, esto hace que el hígado tenga que decantar el
sobrante en la sangre mientras degrada lentamente lo que puede.
Al ser una toxina
no puede ser almacenado de forma permanente en ningún tejido sin que genere
problemas metabólicos con la glucólisis, por lo que el exceso de alcohol en
cada ciclo de purificación debe circular por la sangre y regresar al hígado
para un nuevo ciclo de destrucción. Si en una hora se consume más de una onza
de alcohol puro “copa estándar”, la concentración de alcohol en la sangre
se elevará rápidamente. No existen mecanismos para acelerar el catabolismo del
alcohol en el hígado, y por lo tanto no existe un mecanismo para mantenerse
sobrio por más tiempo sin correr peligros metabólicos. Pero si existen
sustancias que te permiten mantenerte sobrio, pero te expones a la muerte por
intoxicación de alcohol y otros problemas secundarios.
Las moléculas que
sirven como estimulante del sistema nervioso pueden servir como una
contramedida de los efectos del GABA en el sistema nervioso, estimulando la
producción de neurotransmisores estimulantes, pero el efecto tóxico-metabólico
del alcohol continuará siendo el mismo, o peor. De cierta forma la ebriedad es
un reflejo del cuerpo que impide una intoxicación mortal por alcohol, al
consumir estimulantes se puede consumir más alcohol, más allá del límite de
seguridad fisiológico y causar muertes por intoxicación de alcohol con efectos
semejantes al consumo del alcohol adulterado o metanol.
El etanol es una
molécula de desecho de la glucólisis en su fermentación alcohólica, pero los
humanos fermentamos lácticamente produciendo ácido láctico. Como lo que producimos
naturalmente es ácido láctico, nuestros cuerpos son capaces de convertirlo en
glucosa sacrificando ATP proceso llamado gluconeogénesis. Lamentablemente no
poseemos las enzimas para hacer lo mismo con el alcohol, eso lo convierte en
tóxico, pues como se ha mencionado reiterativamente, su acumulación paraliza el
metabolismo de la glucosa inhibiendo la producción de energía. Aproximadamente
el 5% del alcohol es eliminado por los riñones o por los pulmones. El alcohol
liberado por los pulmones sirve como mecanismo de medida para las pruebas de
alcoholemia por aspiración. El restante debe eliminarse por catabolización
hepática durante varias horas.
26.5 Genética del alcohol en el cuerpo humano
Cuando hablamos
de la genética del alcohol debemos repasar antes el último paso de la biología
del alcohol, es decir su bioquímica. Las células hepáticas degradan el alcohol
en un catabolismo de dos pasos principales que lo integran al catabolismo
del ácido acético. En primera instancia el etanol se transformado a aldehído
mediante la enzima alcohol deshidrogenasa. Como muchos intermediarios
metabólicos, el aldehído es tremendamente tóxico, por lo que el cuerpo vigila
que nunca se acumule en altas cantidades al interior de la célula, e
inmediatamente lo conduce a la segunda reacción.
Aunque existen
otras reacciones como la de la catalasa y el retículo endoplasmático, la
reacción que ocurre en el citoplasma es la más importante. El alcohol no se
integra al anabolismo, pero si puede ser catabolizado, extrayendo su energía en
horma de protones de alta energía transportados por el NAD. En la segunda
reacción el aldehído es convertido a ácido acético por medio de la enzima
aldehído deshidrogenasa. El ácido acético o acetato es un componente normal del
catabolismo celular que va directo a la degradación oxidativo, produciendo
dióxido de carbono y agua.
La segunda
reacción ocurre en la mitocondria, nuevamente se obtiene energía por
transferencia de pasos intermedios, uno de los hidrógenos del agua es
reabsorbido por el etanal y cargado de energía, para luego ser transferido al
NAD "reacciones intermedias no mostradas aquí". El producto
porta oxígenos y es el ácido etanoico que ingresa en equilibrio con el ion
etanoato que puede ingresar a diversas rutas metabólicas.
El punto es que la
parte química hace omitir el biológico. Como químicos muchos justan a las
enzimas como moléculas estandarizadas omitiendo que, como entidades biológicas,
estas poseen una estructura interna mediada por una secuencia genética que
posee variaciones entre individuos, familias, poblaciones y etnias. El alcohol
fue empleado como el principal desinfectante del agua por milenios en el mundo
occidental, en oriente se empleó al te para el mismo propósito. Debido a que él
te desinfecta por calor y no por una toxina como el alcohol, los orientales
desarrollaron un polimorfismo de las enzimas que degradan el alcohol menos
efectivo “versiones secundarias” (Day &
Bassendine, 1992; de Alwis & Day, 2007; Vasiliou & Pappa, 2000; Vasiliou,
Pappa, & Estey, 2004; Yoshida, Rzhetsky, Hsu, & Chang, 1998).
Por lo general un
individuo más propenso al alcoholismo cuando porta variantes del alcohol
deshidrogenasa y la aldehído deshidrogenasa más eficaces, lo cual le ayuda a
mantenerse sobrio por más tiempo, y en consecuencia a ingerir alcohol por más
tiempo después de que su unidad social ya no puede beber más. Lo cual nos lleva
al segundo factor, el alcoholismo es una conducta aislante y aislada, los
alcohólicos no saben cuándo para de beber y beben solos.
Por el contrario,
los individuos con poca tolerancia se caracterizan porque hay un desbalance en
la acción de las enzimas que conlleva a una acumulación de aldehído, este al
ser muy tóxico crea el reflejo de ebriedad a mayor velocidad, limitando la
ingesta de alcohol. Estos individuos nunca se quedan solos bebiendo, beben
menos y tienden a tener efectos secundarios más severos debido a una mayor
acumulación de aldehído. En otras palabras, beben menos, tienen una menor
probabilidad de hacerse alcohólicos, pero también tienen una mayor tendencia a
morirse por intoxicación alcohólica debido a la acumulación de aldehído (Day &
Bassendine, 1992; de Alwis & Day, 2007; Vasiliou & Pappa, 2000;
Vasiliou et al., 2004; Yoshida et al., 1998).
26.6 El alcohol y la cirrosis hepática
El primer órgano en ser afectado por el alcohol es el que se encarga de su eliminación, es decir el hígado. Recordemos que el hígado es el primer filtro de toxinas endógenos y exógenos, cuando su función disminuye se sobrecargan sus funciones en el otro sistema de órganos de filtrado, los riñones. El hígado no solo metaboliza las grasas, también regenera el ácido láctico a glucosa, en ese sentido filtra las toxinas “ácido láctico” producidas por el ejercicio, y adicionalmente se encarga de regular las grasas mediante las lipoproteínas de transporte de alta y baja densidad (Adami et al., 1992; Chuang, La Vecchia, & Boffetta, 2009; Deugnier et al., 1993; O’shea, Dasarathy, & McCullough, 2010; Tsochatzis, Bosch, & Burroughs, 2014). Cuando el hígado es afectado por el alcohol lo primero que sucede es una disminución en la capacidad para procesar las grasas, lo cual genera una acumulación de lípidos en los hepatocitos. Cuatro o cinco tragos al día por varias semanas, son más que suficientes para comenzar a dañar el hígado e iniciar el proceso de acumulación de grasas. En esta primera etapa las células de hígado aún no están dañadas y con una abstinencia de varias semanas su funcionamiento se restaura a la normalidad.

Figura 26.7. Cirrosis hepática.
Aunque el hígado es el órgano más resistente y regenerable, tiene sus límites,
sin él no tenemos escudo contra las toxinas que producimos (YouTube) (YouTube) (YouTube).
Adicionalmente el
alcohol incrementa la acumulación de grasas por dos rutas. La primera es que
las bebidas alcohólicas poseen un contenido de carbohidratos no fermentados
residuales que contribuye a su sabor, pero que pueden ser anabolizados a grasas
por el cuerpo humano. Adicionalmente cada molécula de alcohol puede en potencia
generar dos NADH+ que sirven para crear energía en la cadena de
transporte de electrones, disminuyendo la necesidad de quemar glucosa o grasa
por parte del cuerpo. Cuando los hepatocitos alcanzan su límite de acumulación
de grasas explota, y el sistema inmune los encierra en un tejido cicatrizante
que no posee función de filtro metabólico al cual llamamos quiste. A medida que
los quistes se acumulan, la función de filtro disminuye y la acumulación de
grasas aumenta, el riñón también se sobre-esfuerza lo cual retroalimenta la
acumulación de toxinas en el cuerpo. Con el tiempo se genera la inflamación
generalizada del hígado, proceso denominado hepatitis alcohólica.
A medida que los
quistes se fusionan en cicatrices más profundas la función hepática disminuye
aún más, condición conocida cono cirrosis, impidiendo que la sangre fluya por
el hígado, lo cual es especialmente para el sistema porta que viene del
intestino con los nutrientes. Con el atascamiento del sistema circulatorio
porta intestinal el intestino se atora y empiezan a reventare sus capilares, lo
cual causa sangrado intestinal, mientras que los riñones sobrecargados
renuncian a su función por sobrecarga y agotamiento, lo cual crea
desequilibrios osmóticos y acumulación de líquidos. Una vez que los riñones y
el hígado fallan es cuestión de tiempo hasta que las toxinas del metabolismo
basal destruyan el cuerpo conllevando a la muerte. Adicional a la cirrosis,
estadísticamente se ha demostrado que el alcoholismo es una causa de riesgo
para el desarrollo de cáncer del sistema gastrointestinal, y que al combinar
los factores de riesgo del alcoholismo con el consumo de tabaco el índice de
formación de tumores aumenta exponencialmente.
26.7 Efectos del alcohol en el sistema circulatorio y el cáncer
(YouTube) El alcohol tiene una relación ambivalente con el sistema circulatorio que depende de la cantidad que se consume al día. UNA sola copa al día estadísticamente disminuye la probabilidad de un ataque al corazón o de un derrame cerebral. Los mecanismos de este proceso aún no están del todo claros, pero se hipotetiza que el alcohol mejora la producción de lipoproteínas de alta densidad en el hígado. Las lipoproteínas de alta densidad o HDL por sus siglas en inglés son conglomerados de proteínas, fosfolípidos y colesterol que recolecta grasa acumulada en las arterias y las reenvía al hígado. Eso tiene sentido con los inicios de la cirrosis que es una acumulación de grasas en el hígado (Dai, Mukamal, Krasnow, Swan, & Reed, 2015; Matsumoto, Miedema, Ofman, Gaziano, & Sesso, 2014; Rader & Hovingh, 2014).

Figura 26.8. Corazón alcohólico. El
corazón de un alcohólico puede ser del doble del tamaño de un corazón normal.
En la imagen podemos apreciar a la izquierda un corazón no alcohólico y a la
derecha un corazón alcohólico.
En consecuencia,
consumir suficiente alcohol para enviar más grasa al hígado es bueno ya que
impide que estas se acumulen en capilares donde pueden provocar la enfermedad
de los vasos sanguíneos que denominamos arteriosclerosis. Cuando el consumo de
alcohol aumenta el hígado recibe más grasa, pero ya no puede procesarla,
provocando la acumulación, la explosión de los hepatocitos saturados de grasa,
la enquistación y la cirrosis.
El aumento
crónico de alcohol en la sangre favorece la mutación de los tejidos en contacto
con él, lo cual favorece la formación de tumores en cualquier parte del cuerpo,
al ser una toxina lastima los vasos sanguíneos y el corazón, por lo que estos
aumentan su tamaño ara mantener la homeostasis del cuerpo.
26.8 Síndrome de abstinencia y tolerancia en el
alcoholismo
(YouTube) El síndrome de abstinencia del alcoholismo va de
leve “resaca o guayabo” a severa y afecta principalmente al sistema nervioso
central. El síndrome leve está caracterizado a la dependencia fisiológica,
mientras que el síndrome severo está caracterizado por la dependencia
psicológica, característica compartida por cualquier sustancia de abuso. Sin
embargo algunos factores fisiológicos pueden contribuir a la dependencia a
largo plazo como la anhedonia por tolerancia cruzada a los neurotransmisores
endógenos (Bell et al., 2017;
Most, Ferguson, & Harris, 2014).
La dependencia
leve o guayabo inicia entre 6 y 24 horas después de la última ingesta para
individuos que aún no presentan problemas de alcoholismo. Sin embargo, a medida
que los episodios de consumo se hacen más habituales, el guayabo tiende a
prolongarse por más de un día y llegar a una semana de dependencia física. Como
cualquier síndrome de abstinencia, el guayabo decae si el cuerpo consume
alcohol, reforzando la dependencia. Una vez que el individuo experimenta un
guayabo que dura más de un día y que por lo tanto puede abarcar dos episodios
de ingesta de alcohol se encontrará enganchado fisiológicamente a este, y
seguirá bebiendo para no sentir dolor.
El síndrome de
abstinencia leve se caracteriza por espasmos musculares “temblores”, vómitos,
dolores de cabeza por deshidratación e hipersensibilidad a la luz, pero una vez
que el síndrome de abstinencia se prolonga y agrava empieza a incluir temblores
prolongados en las manos, ansiedad, insomnio, alucinaciones –auditivas,
visuales o táctiles –e inestabilidad del sistema nervioso autónomo.
Este es el
síndrome de abstinencia fisiológico, el cual crea un refuerzo negativo del
comportamiento –si no bebes de va a doler. El alcohol es un sedante-hipnótico
empleado para aliviar penas psicológicas, por lo tanto, crea una situación de
dependencia psicológica a largo plazo que se mantiene mucho tiempo después de
haberse liberado del síndrome de abstinencia. Esto crea un refuerzo positivo
del comportamiento –si bebes te sentirás mejor. La tolerancia al alcohol
también involucra una tolerancia cruzada a glutamato y el GABA en situaciones
de normalidad, en consecuencia, las experiencias placenteras previas a la
exposición por alcohol disminuyen su efecto después, en términos poéticos es
como si el mundo aumentara su color con el consumo, pero después fuera aún más
gris que antes de la ingesta. A este efecto del alcohol y otras drogas se lo
denomina anhedonia y puede durar para el alcohol por casi un año.
La tolerancia es
causada no solo por la degradación de los receptores para GABA, sino por
alteraciones en los patrones de expresión genética, ergo, el alcohol posee
efectos epigenéticos sobre el núcleo de las neuronas, lo cual intensifica el
síndrome de abstinencia, prolongándolo por meses después de la última ingesta.
27. Adicción a la marihuana
|| Regresar al Índice ||
A diferencia del alcohol, la marihuana es una sustancia ilegal, sin embargo estadísticamente es la sustancia ilegal de mayor consumo en los Estados Unidos y en el mundo, adicionalmente se trata de la primera droga de abuso con la cual inician los consumidores asiduos antes de pasar a sustancias más potentes (Diaz & Fernandez, 2013; Hasin et al., 2015; Mendoza, Romero, Mendoza, & Posada, 2015; Scoppetta Díaz-Granados, Pérez Gómez, & Muñoz, 2013). La marihuana se obtiene de las hojas, las flores y los tallos de una planta de origen hindú llamada Cannabis sativa. La molécula activa principal es un canabinoide, una molécula relacionada con los fosfolípidos de membrana por lo que puede atravesar la barrera hematoencefálica a pesar de ser una molécula de gran tamaño. La molécula en si se denomina delta-9-tetrahidrocanabinol o THC por sus siglas en el nombre principal. Los efectos de la marihuana dependen de la concentración de THC, los cuales dependen a su vez de la concentración en el Cultivar específico empleado para fabricar la dosis, y de la cantidad de dosis administradas por unidad de tiempo. En pequeñas dosis el TCH provoca euforia y sensación de bienestar, pero a mayores concentraciones se generan alucinaciones y paranoia con eventos de ansiedad tan fuertes que llegan al pánico.

Figura 27.1. Endocanabinoides.
Aunque los canabinoides están hechos a partir de grasa y deberían poder
atravesar la membrana, su receptor esta acoplado a una proteína G de membrana (YouTube).
Al igual que con el alcohol el síndrome de abstinencia
fisiológico no es demasiado severo y la limpieza puede realizarse en poco
tiempo. Se caracteriza por incapacidad para descansar, irritabilidad, ansiedad
leve, insomnio, nausea y calambres en las articulaciones. El verdadero problema
es la dependencia psicológica por retroalimentación positiva, que usualmente
provoca que el consumidor regrese a la sustancia. Estudios sugieren que los
exocanabinoides de la marihuana pueden causar efectos a largo plazo en los
cerebros de individuos adolescentes (Rey, Martin, & Krabman, 2004;
Schneider, 2008; Vandrey & Budney, 2015).
27.1 Mecanismo de acción de la marihuana
Al ser una droga ilegal, la investigación sobre los
mecanismos de acción de la marihuana ha sido un problema por décadas, por lo
que el entendimiento de sus mecanismos moleculares aún se encuentra en pañales (Davies & Graham, 1980; Di Marzo & Matias,
2005; Freeman et al., 2014; Martin & Wiley, 2003).
Los endocanabonoides se sintetizan a partir de lípidos de
membrana "morado" cuando la neurona postsináptica se estimula
demasiado por ingreso del ion calcio(2+). Los endocanabinoides una vez
sintetizados "azul" viajan por la sinapsis para unirse a los
receptores (CB1), los cuales bloquean los canales de calcio(2+) de la neurona
presináptica bloqueando la comunicación entre las neuronas sensoriales. El
canabinoide más relevante en la marihuana es el tetrahidrocannabinol “THC”
aunque no es el único.
A pesar de que el THC puede atravesar libremente la
barrera hematoencefálica, estos se unen en su tejido de blanco a receptores
específicos que inducen la liberación de dopamina en los núcleos de recompensa
del cerebro. La verdadera pregunta es ¿y porque tenemos esos receptores? Una
respuesta evolucionista es que estos deben haber evolucionado como parte del
mecanismo natural de recompensas del cerebro durante millones de años, lo cual
trae al menos dos consecuencias procedimentales: (1) Deben existir
endocanabonoides, los efectores naturales para los receptores de THC que
inducen la liberación normal de dopamina y (2) que otros mamíferos también
deben poseer endocanabinoides.
Lo anterior permite hipotetizar la existencia de los
endocanabonoides y emplear modelos biológicos para la investigación. La
búsqueda no fue sencilla, se emplearon cientos de moléculas para ver cual tenía
alta afinidad con los receptores de THC en conejillos de indias hasta que
dieron con una. La molécula fue denominada anandamida, molécula propia del
cuerpo que se libera cuando el cuerpo experimenta estados de concentración que
inducen experiencias semejantes a las de fumar marihuana, aunque menos potentes
y relacionadas al medioambiente.
El mecanismo de acción de los endocanabinoides es el de
un freno de mano para señales del dolor. Los receptores de endocanabinoides se
ubica en la materia gris de la médula y el tallo cerebral, donde regulan las
señales de dolor. Los endocanabinoides son producidos por una neurona
postsináptica de transmisión sensorial cuando se encuentra sobreexitada, los
endocanabinoides atraviesan libremente la membrana postsináptica y viajan por
la sinapsis a los receptores endocanaboniudes. Cuando la pareja efecto-receptor
se acopla, los receptores bloquean los canales de sodio evitando que la neurona
presináptica descarge neurotransmisores de dolor creando un efecto analgésico.
Al igual que con los opioides, si la pareja
efector-receptor permanece ensamblada por demasiado tiempo, los receptores son
endocitados y destruidos en las vesículas de tráfico de ser necesario, lo cual
conlleva al síndrome de abstinencia permanente o anhedonia, el cuerpo pierde el
potencial de ser activado por los endocanabinoides por lo que tenderá a sentir
más dolores en estado de normalidad que antes de haber iniciado el consumo.
27.2 Efectos del uso a largo plazo de la
marihuana
La marihuana es capaz de inducir síndrome de abstinencia
agudo por refuerzo negativo, pero también el síndrome de abstinencia crónica
por degradación de los receptores de endocanabinoides, lo cual desencadena
anhedonia, que es una degradación de la capacidad natural para sentir los andocanabonoides
naturales (Celofiga, Koprivsek, & Klavz, 2014). La anhedonia está enlazada a la dependencia positiva conectada al sistema
límbico y las emociones, lo cual crea el enganche a largo plazo.
Adicionalmente existe una serie de efectos secundarios
relacionados con otras moléculas asociadas a la marihuana, las cuales dependen
del mecanismo de dosificación. Los efectos nocivos de la marihuana están
directamente relacionados a su mecanismo usual de administración, fumar. No es
de sorprender entonces que los mismos peligros que se asocian al cigarrillo
estén unidos al consumo de marihuana con una pequeña diferencia, la formación
del cachos “cigarrillos de marihuana” de marihuana no está realizada por
profesionales, y al emplearse cualquier papel y aditivos no estandarizados, la
proporción de sustancias carcinogénicas en el humo de marihuana es usualmente
más grande, especialmente de alquitrán (Callaghan, Allebeck, & Sidorchuk, 2013; Huang et
al., 2015; Volkow, Baler, Compton, & Weiss, 2014; Zhang et al., 2015).
Como resultado, los fumadores de marihuana a pesar de
consumir menos cigarros al día que un fumador de tabaco, acumulan
estadísticamente tres veces más alquitrán en sus pulmones, y cinco veces más
monóxido de carbono fluyendo en la sangre. El monóxido de carbono se une
irreversiblemente a la hemoglobina, disminuyendo el poder respiratorio de la
sangre con independencia a los pulmones, en otras palabras, aunque los pulmones
no fueran lastimados por el alquitrán, deberían trabajar más fuerte debido a
que la sangre puede trasportar menos oxígeno. El alquitrán por otra parte es
una de las moléculas carcinogénicas más importantes, si no es que la más
importante en el humo de los cigarrillos legales del tabaco, y eso que estos
cigarros legales si poseen filtros contra el alquitrán. Los cigarros de la
marihuana no tienen filtros, lo cual explica el flujo de alquitrán
El THC también puede consumirse por ingestión, pero el
hígado degradará una gran cantidad, por lo cual la marihuana en pasteles genera
efectos más lento, con menor intensidad, pero también con mayor seguridad ya
que no afectas negativamente tus tejidos con gases de combustión
carcinogénicos.
El THC tiene similitudes estructurales con el estradiol,
en consecuencia tiene efectos endocrinos opuestos a la testosterona (Hillard, 2015; Takeda, 2014). En los hombres reduce la cantidad de testosterona y la producción de
espermatozoides, mientras que en las mujeres altera los ciclos menstruales
creando irregularidades en su ciclo. Sin embargo no todo es malo, si se
dosifica la marihuana de forma legal y regulada medicamente se convierte en un
analgésico efectivo, así como un inductor del apetito en individuos que sufren
tratamientos por radiación contra algunos tipos de cáncer y los que sufren los
efectos del VIH en su fase de SIDA (Naguib & Foss, 2015; Parmar, Forrest, &
Freeman, 2016).
27.3 La marihuana sintética
Los cannabinoides sintéticos se refieren a un número creciente de compuestos químicos artificiales que alteran la mente, que se rosean sobre material vegetal seco o desmenuzado, para ser fumado (como incienso herbal). También pueden ser vendidos en forma líquida para ser vaporizados e inalados en cigarrillos electrónicos y otros aparatos (inciensos líquidos). Estos químicos son llamados cannabinoides porque pertenecen a la familia de compuestos químicos encontrados en la planta de marihuana. Debido a esta similitud, los cannabinoides sintéticos algunas veces son llamados, engañosamente, "marihuana sintética" (o "fake weed"), y son publicitados con frecuencia como alternativas legales y "no peligrosos" de la marihuana. Pero de hecho, pueden afectar el cerebro mucho más potentemente que la marihuana; los efectos verdaderos pueden ser impredecibles y en algunos casos, severos o hasta fatales (Castaneto et al., 2014; Hermanns‐Clausen, Kneisel, Szabo, & Auwärter, 2013; Mesa & Bello, 2015; Musshoff et al., 2014; Tortajada, San Miguel, Domenech, Cucarella, & Aliño, 2015).

Figura 27.2. Laboratorio artesanal. No
esperen que un traficante ilegal tenga tecnología de punta o control de
calidad.
Los productos cannabinoides sintéticos son frecuentemente etiquetados como "no aptos para el consumo humano". Las etiquetas también frecuentemente dicen que contienen materiales "naturales" tomados de una variedad de plantas. Sin embargo, la única parte de estos productos que son naturales son los materiales de planta seca. Exámenes químicos muestran que los ingredientes activos, alterantes de la mente en los compuestos de cannabinoides, son hechos en laboratorios improvisados (Castaneto et al., 2014; Hermanns‐Clausen et al., 2013; Mesa & Bello, 2015; Musshoff et al., 2014; Tortajada et al., 2015).

Figura 27.3. Marihuana sintética. Los
compuestos químicos de la marihuana sintética tienen un alto potencial de abuso
y ningún beneficio médico
Los cannabinoides sintéticos pertenecen a una clase de
drogas llamadas "nuevas sustancias psicoactivas" ("new
psychoactive substances" – NPS). Las NPS son sustancias psicoactivas no
reguladas que se han vuelto recientemente asequibles en el mercado y están
supuestas a imitar los efectos de las drogas ilegales. Algunas de estas
sustancias pueden haber estado en circulación por muchos años, pero han
reingresado al mercado en forma de compuestos químicos alterados o a su
creciente popularidad. Los fabricantes venden estos productos de inciensos
herbales en paquetes coloridos de papel metalizado, y productos similares de
inciensos líquidos, como otros líquidos de cigarrillos electrónicos, en
botellas plásticas. Estos productos se ofrecen bajo una amplia variedad de
marcas. En años recientes, "K2" y "Spice" eran las marcas
más comunes. Ahora se pueden encontrar cientos de marcas distintas, como
"Joker", "Black Mamba", "Kush" y
"Kronic" (Castaneto et al., 2014; Hermanns‐Clausen et al.,
2013; Mesa & Bello, 2015; Musshoff et al., 2014; Tortajada et al., 2015).
Por muchos años, ha sido muy fácil comprar mezclas de
cannabinoides sintéticos en tiendas de drogas parafernalia, tiendas de
novedades, estaciones de gasolina y a través del internet. Pero como los
compuestos químicos usados tienen un alto potencial de abuso y ningún beneficio
médico, las autoridades han hecho que sea ilegal vender, comprar, o tener en
posesión cualquiera de estos compuestos. Sin embargo, los fabricantes continúan
de sobre pasar estas leyes cambiando las formulas químicas de sus mezclas. Es
probable que el acceso fácil y la creencia que los cannabinoides sintéticos son
"naturales", y por lo tanto inofensivos, haya contribuido a su
popularidad creciente entre los jóvenes. Otra razón para su uso es que muchos
de los compuestos químicos usados en estos productos no son fácilmente
detectados en las pruebas estandarizadas de drogas (Castaneto et al., 2014; Hermanns‐Clausen et al.,
2013; Mesa & Bello, 2015; Musshoff et al., 2014; Tortajada et al., 2015).
Los cannabinoides sintéticos actúan en los mismos
receptores celulares del cerebro que
el delta-9-tetrahydrocannabinol (THC), el ingrediente de la marihuana
que altera la mente. Hasta ahora, ha habido pocos estudios científicos sobre
los efectos de los cannabinoides sintéticos en el cerebro humano, pero los
investigadores saben que varios de estos compuestos se unen más fuertemente a
los receptores celulares que el THC de la marihuana, por lo que podrían
producir efectos más fuertes. Las consecuencias adversas para la salud podrían
ser impredecibles. Como la composición química de muchos cannabinoides
sintéticos es desconocida y/o cambiante de un lote a otro, es muy posible que
estos productos contengan sustancias que causan efectos dramáticamente
diferentes de los que espera el consumidor. Los consumidores de los
cannabinoides sintéticos reportan varios efectos similares a los efectos
producidos por la marihuana, tales como: Emociones elevadas; Relajación;
Percepción alterada—conciencia aumentada de las condiciones y objetos
circundantes; Síntomas de psicosis—delirios o pensamientos desorganizados o
desconectados de la realidad; Ansiedad extrema; Confusión; Paranoia—extrema e
injustificada desconfianza de otros; Alucinaciones—sensaciones e imágenes que
parecen reales pero no lo son. (Castaneto et al., 2014; Hermanns‐Clausen et al.,
2013; Mesa & Bello, 2015; Musshoff et al., 2014; Tortajada et al., 2015)
Las personas que han usado cannabinoides sintéticos y han
sido llevados a salas de emergencia han mostrado varios efectos severos, que
incluyen: Aumento del ritmo cardiaco; Vómito; Comportamiento violento;
Pensamientos suicidas (Castaneto et al., 2014; Hermanns‐Clausen et al.,
2013; Mesa & Bello, 2015; Musshoff et al., 2014; Tortajada et al., 2015). Los cannabinoides sintéticos también pueden subir la presión arterial y
reducir el suministro de sangre al corazón, al igual que daños a los riñones y
convulsiones. El uso de estas drogas está asociado con un número creciente de
muertes.
27.4 Los ganchos de consumo
Si bien la marihuana es ilegal, sus efectos en sí no son
tan graves o marcados como los de otros tipos de sustancias, su síndrome de
abstinencia puede superarse sin mayores dificultades, sin embargo hay que
recordar que el expendedor desea ganar dinero, y al ser una sustancia ilegal,
esta puede mezclarse con otras drogas fuertemente adictivas, de esta forma una
posible forma de adquirir más clientes permanentes es vender mezclas de
marihuana con bazuco, pues este último es absolutamente adictivo en comparación
con la marihuana, combinación que en las calles de Bogotá hacia mediados de la
década del 2010 es denominada muy apropiadamente DIABLO.
Sin embargo, aun cuando el expendedor no sea mañoso y
comience vendiendo únicamente marihuana, el panorama no es para nada alentador.
Estudios a largo plazo de estudiantes de secundaria y sus patrones de uso de
drogas muestran que muy pocos de ellos llegan a usar otras drogas ilegales sin
haber usado primero la marihuana. Por ejemplo, el riesgo de usar cocaína es
mucho mayor para las personas que han probado marihuana en comparación con las
que nunca la han probado. El consumo de marihuana pone a los niños y jóvenes en
contacto con personas que usan y venden esta y otras drogas. En este sentido,
sí existe mayor riesgo de que los jóvenes que usan marihuana estén más
expuestos y tengan mayor tentación de probar otras drogas (la Haye, Green, Kennedy, Pollard, & Tucker, 2013;
Savage, King, Clark, & Cropsey, 2017).
En este caso la causalidad no es biológica o química, sino meramente social, quien vende marihuana vende otras drogas más potentes, así que el paso de la una a la otra es causado no por el canabinoide en sí, sino por la proximidad social dada por los expendedores, que aconsejan el consumo de sustancias más fuertes, y una vez que han localizado a individuos vulnerables a los cuales pueden explotar con mayor facilidad.

Figura 27.4. Ganchos de consumo. La
marihuana muchas veces se emplea como gancho de consumo por dos rutas,
vendiéndola mezclada con sustancias que generan dependencia más rápido o, por
el simple hecho de que alguien bajo sus efectos tomará decisiones que no
realizaría estando sobrio.
27.5 La legalización
Dado lo anterior, el debate de la legalización se hace
particularmente pertinente, si se legaliza la marihuana, que no es lo mismo que
legalizar todas las drogas, se arrebata una puerta de entrada a las drogas,
aunque claro, esto no eliminaría el problema, pues igual hay adictos a drogas
legales. En la actualidad su uso legal ha sido permitido para fines medicinales
por sus efectos analgésicos en Colombia u otros países del mundo. En noviembre
de 2015, mediante Decreto Presidencial fue aprobada y legalizada la producción
y comercialización de cannabis con fines medicinales y científicos. El 25
de mayo de 2016 el congreso de la república (Cámara de Representantes, Senado y
cooperación de él joven Juan José Arredondo Zapata), han legalizado el uso
medicinal del cannabis con el fin de ampliar el marco legal y blindar
jurídicamente el decreto presidencial de noviembre de 2015.
28. Adicción a los estimulantes
|| Regresar al Índice ||
Las moléculas de
abuso generalmente se han distinguido en dos grupos principales, estimulantes
del sistema nervioso central y depresores del sistema nervioso central. Los
estimulantes se caracterizan por incrementar la conexión de la corteza cerebral
con los sentidos, así como incremento en el metabolismo basal. Los estimulantes
aumentan la concentración, mejoran el rendimiento motor e incrementan la
sensibilidad. En la actualidad la molécula estimulante del sistema nervioso
central universalmente distribuida y consumida es la cafeína, a cuál se
encuentra en los frutos de las plantas del café o del té. La cafeína no está
relacionada con los receptores de dopamina y tampoco estimula su liberación,
esto explica porque a pesar de que sus efectos estimulantes son marcados, no
crean una adicción potente (Creese, 1983; George, 2000; McKim
& Hancock, 2012; Solanto, 2001; Weil & Rosen, 1983).
La cafeína es un
antagonista de la adenosina, una base nitrogenada con función de
neurotransmisión en la sinápsis. Como antagonista bloquea la función de los
receptores de adenosina. La adenosina al igual que GABA es un bloqueador de
varias funciones neuronales, por lo que la cafeína elimina los frenos en el
sistema nervioso. El principal neurotransmisor bloqueado por la adenosina es la
acetilcolina, por lo que el consumo de cafeína genera que los músculos
respondan con mayor facilidad cuando el individuo está despertando. Otro
mecanismo de estimulación de la acetilcolina es que la cafeína bloquea la
acetilcolinesterasa momentáneamente, favoreciendo al tono muscular. A
continuación, examinaremos específicamente las moléculas de abuso y en
consecuencia excluimos a la cafeína, las sustancias que describiremos son la
cocaína, las anfetaminas y la nicotina.
28.1 La cocaína
La cocaína es una
molécula extraída de las hojas de la planta de coca “Erythroxylon coca”,
que crece de forma natural en las montañas de Sur América, especialmente en
Perú, Bolivia, Colombia y Ecuador. Su consumo natural se hace masticando la
hoja de coca, en cuyo caso sus efectos son semejantes a los de la cafeína, pero
algo más fuertes. La cocaína es la molécula activa purificada a partir de las
hojas de coca, sería necesario masticar cerca de cuatro toneladas de hoja de
coca para consumir la misma cantidad de cocaína que una línea de polvo
purificado (Bradberry, 2002,
2002; Grinspoon & Bakalar, 1981).
El polvo purificado es inhalado sin que exista combustión, este se diluye rápidamente en las mucosas y las membranas celulares, por lo que atraviesa la barrera hematoencefálica estimulando al cerebro en segundos. Los efectos de la cocaína inyectada son semejantes, por lo que su consumo en polvo tiende a ser más económico y, en consecuencia, más común. Las formas de cocaína desarrolladas para su consumo por combustión poseen un efecto más fuerte y más rápido, son particularmente adictivas y presentan los riesgos asociados a cualquier sustancia fumada, como la acumulación de alquitrán y monóxido de carbono. Algunas formas de cocaína para combustión son el Crack y el Bazuco, económicas de fabricar y extremadamente adictivas.

Figura 28.1. La planta de coca Erythroxylum coca.
28.2 Efectos neuronales de la cocaína
Cuando ingerimos la cocaína, esta crea una intensa sensación de placer, aumenta la autoconfianza e incrementa el metabolismo basal mejorando el rendimiento físico, la capacidad cognitiva y el vigor físico.

Figura 28.2. Sinapsis de
dopamina. La cocaína básicamente bloquea
el transportador de la dopamina, impidiendo que esta se recicle de la sinapsis,
el resultado es que los receptores queden activados durante más tiempo y en
mayor número (YouTube) (YouTube).
La cocaína
incrementa los niveles de neurotransmisores relacionados con el placer, la
recompensa y el reflejo de Combate y Escape, específicamente la dopamina y la
norepinefrina “noradrenalina”. La euforia es causada principalmente por los
efectos en la dopamina, el principal neurotransmisor de los centros de
recompensa (Cagniard,
Sotnikova, Gainetdinov, & Zhuang, 2014; Freyberg et al., 2016; Ritz, Lamb,
Goldberg, & Kuhar, 1987; Rossum, Schoot, & Hurkmans, 1962; Wise, 1984;
Yasui & Su, 2016).
Normalmente la
dopamina es inmediatamente reabsorbida al interior de los nervios, por lo que
sus efectos se acaban rápidamente. La cocaína sin embargo interfiere con el
reciclaje de la dopamina aumentando y prolongando el efecto normal de la
dopamina, se trata de un efecto antagonista tapando las proteínas que reciclan
la dopamina. La cocaína crea un efecto paradójico, por un lado, otorga placer
inmediato, y por otro somete al cuerpo a estrés por la acción de la
norepinefrina, manteniendo al cuerpo listo para afrontar situaciones de
emergencia, lo cual en este caso también mejora el desempeño sexual. Los
efectos de la cocaína son más fuertes con las primeras dosis, durando hasta
unos noventa minutos, pero decrece paulatinamente a medida que el cuerpo aumenta
sus niveles de tolerancia hasta alcanzar unos dos minutos.
Cuando el
individuo regresa a la realidad no lo hace al estado homeostático estándar, por
el contrario, su cerebro cae en un estado de depresión fuerte, ansiedad y
fatiga extrema. Estos efectos son mucho más marcados que con el alcohol o la
marihuana y solo cesan de forma efectiva con el consumo de más cocaína, los
ciclos incrementan la tolerancia y aumentan los efectos secundarios, a este
fenómeno lo denominamos síndrome de abstinencia. Un factor que favorece la
recaída más allá del síndrome de abstinencia agudo es la anhedonia, esta droga
te arrebata la capacidad de sentir placer por medios normales por años. La
tolerancia y la anhedonia están relacionadas, si la dopamina no es reciclada el
receptor dopaminérgico es percibido como defectuoso por la neurona
postsináptica y lo endocita. Debido a que la vesícula encierra dopamina, el
receptor dopaminérgico no se libera y por lo tanto es degradado.
Entre más alto
sea el “vuelo” más fuerte será el “estrellón” y por lo tanto más fuerte la
necesidad por más cocaína. El Crack y el Bazuco son especialmente adictivos al
ser 75% cocaína, en comparación con el 10% al 35% del polvo de cocaína que se
obtiene en las calles.
28.3 Efectos secundarios de la cocaína
(YouTube) La cocaína como cualquier estimulante aumenta
los peligros de ataque al corazón, los cuales se ven exacerbados por un
mantenimiento del estado de Combate o Escape del cuerpo. La presión sanguínea
aumenta mucho por dos factores, el aumento en los latidos del corazón y una
fuerte constricción de los capilares sanguíneos. El ritmo cardiaco puede
hacerse irregular, y alterar el estado inconsciente del sistema nervioso
autónomo, algunos adictos reportan la sensación de estar conscientes de los
latidos de su corazón. Las sobredosis pueden afectar los nervios del marcapaso
causando un paro cardíaco.
La depresión después de la caída de un vuelo por cocaína es literalmente fisiológica. Tendemos a señalar los estados de la mente como la depresión en sentido de la moral cristiana y el libre albedrío, sin embargo, esto tiene muy poco que ver en esto, el individuo puede racionalizar lo que sucede en su cerebro, pero no tiene un control en lo que sucede en su mente o en sus sentimientos. La depresión involucra los centros autónomos de la respiración, por lo que el ritmo respiratorio también se deprime, al punto en que la respiración puede resultar inhibida. Esto se causa debido a que cuando pasa el efecto de la cocaína, los receptores dopaminérgicos se encuentran en pleno proceso de endocitosis, y a medida que decaen en las neuronas la excitabilidad neuronal decae radicalmente.

Figura 28.3. Destrucción de tejidos. La cocaína esnifada afecta degradando el cartílago nasal y el paladar secundario.

Figura 28.4.
El consumo de esta y otras sustancias, junto con sus estilos de vida
asociados, aceleran el proceso de envejecimiento.
La cocaína y todas
las drogas relacionadas como las anfetaminas tienen un potente efecto
degradante sobre los tejidos orgánicos como los epitelios, el cartílago e
inclusive partes tan duras como los dientes. Evidentemente las neuronas también
se disuelven en el proceso, por lo que la tolerancia, el síndrome de
abstinencia y la anhedonia se generan no solo por degradación de la
neurotransmisión, sino porque ya no hay nada que neurotransmita.
El daño en el
sistema respiratorio o cardiovascular también está influido por los mecanismos
de dosificación. La cocaína en polvo destruye los nervios nasales, la capa
mucosa y los vasos sanguíneos de la nariz. En consecuencia, la esta se torna
seca debido al tejido cicatrizante “cayo” que crece para protegerse. Sin
mucosas los índices de infecciones respiratorias aumentan, así como los índices
de jaquecas debido a que el aire frio extrae calor del cerebro a través del
hueso que separa el canal respiratorio de la bóveda craneana. El tabique de
bifurcación nasal tiende a corroerse y desintegrarse. Uno de los aspectos del
sistema nervioso autónomo simpático es que disminuye la sensación de dolor, y
la cocaína estimula al sistema simpático al máximo, en consecuencia, es un
analgésico potente, por lo que todos estos síntomas pasarán desapercibidos
hasta que se haya causado un daño considerable.
La degradación no
se limita a unos cuantos sistemas, todos los tejidos son afectados lo cual
acelera el ritmo de envejecimiento biológico, al final representas más años
biológicos que lo que tienes cronológicamente. Las inyecciones por otra parte
favorecen las infecciones secundarias del sistema circulatorio, ya sea con
bacterias ambientales que pueden generar septicemia “que trae como consecuencia
la amputación del miembro” o con patógenos oportunistas que infectan la sangre
como muchos microorganismos de trasmisión sexual tales como la hepatitis y el
VIH. Fumar crack/bazuco por otro lado daña rápidamente los pulmones y las vías
respiratorias debido a la acumulación de alquitrán, plásticos y los demás
aditivos químicos empleados para crear la mezcla. Las consecuencias son
semejantes a las de fumar tabaco solo que mucho más poderosos debido tanto a la
mala fabricación de los cigarros como a la propiedad analgésica de la cocaína
que hace que la patología sea detectada cuando ya está muy avanzada.
28.4 El bazuco
(YouTube) (YouTube) La pasta base de cocaína (llamada también paco,
bicha, bazuco, bazuco, base o carro) es una droga de bajo costo similar al
crack elaborada con residuos de cocaína y procesada con ácido sulfúrico y
queroseno. En ocasiones suele mezclarse con cloroformo, éter o carbonato de
potasio, entre otras cosas (Arias Dioses,
2014; Carmona & Novoa, 2012; Cervantes, 2003; Jonathan, 2006; Rojas Valero,
2015; Sabogal-Carmona & Urrego-Novoa, 2012).
La dosis pesa
solo entre 0,01 y 0,03 gramos. Es una droga estimulante del sistema nervioso
central, cuyo componente activo es el alcaloide cocaína, proveniente de una
planta natural de América, la Erythroxylum
coca, al que "estiran" o
adulteran con otros componentes como cafeína, manitol y bicarbonato de sodio,
entre otros. Ingresa al organismo luego de ser fumado en instrumentos
especialmente diseñados para tal fin, con el objeto de obtener euforia y
placer; pero tiene un efecto tan efímero como deletéreo para todo el organismo,
lo que genera una rápida dependencia, con aumento de la frecuencia de su uso,
llevando a las personas a consumir decenas de dosis diarias.
En los minutos que siguen a su absorción, aparecen los efectos sobre el sistema nervioso central, con riesgo de pérdida de conocimiento y convulsiones; luego se afecta el sistema cardiovascular, produciendo alteraciones del ritmo cardíaco y aumento de la tensión arterial, lo que pone en riesgo la vida. Si el consumo está asociado a bebidas alcohólicas u otras drogas, el efecto perjudicial se multiplica. Con el uso prolongado, aun en corto tiempo, el deterioro neurológico y por ende intelectual del sujeto se hace evidente, se acompaña también de alteraciones pulmonares y cardíacas, marcada pérdida de peso y estado de abandono personal. Existen consecuencias indirectas asociadas como los traumatismos o lesiones por accidentes o riña, en ocasiones, graves.

Figura 28.5. Atrapados por el bazuco.
Se suele consumir
fumándola en pipas (generalmente caseras), con marihuana (mixto, combi, flapy,
fillywey, maduro o banana) o con tabaco en forma de cigarrillo (pecoso,
tabacazo, tabaquito, tola, marciano, maduro, bazuco, pistolo, calavera,
marrueco, Free o ensacado). Debido a su composición química, es altamente
tóxica y al ser muy breve su efecto (entre 10 y 15 minutos) es extremadamente
adictiva. Se obtiene a través de la extracción de alcaloides de la hoja de coca
que no llegan a ser procesados hasta convertirse en el clorhidrato de cocaína,
que es la forma más común y buscada de presentación de esa sustancia. La
extracción de los mismos se realiza mediante la maceración de la hoja de coca
en queroseno u otros solventes, pero por la escasez de precursores químicos
para convertir este grupo de alcaloides en la sal de cocaína por reacción con
el ácido clorhídrico, se vende de esta forma en buena parte de Sudamérica.
Sus efectos
secundarios son muy similares a los de la cocaína, el riesgo a la salud es
mucho más alto por residuos de disolventes y otras sustancias nocivas que
contiene. Debido a que la cocaína, componente activo, no está refinada, se le
suelen llamar "fondo de olla", haciendo referencia a que son los
restos que quedan de la conversión de la pasta base a clorhidrato de cocaína.
El consumo de pasta base se realiza a través de pipas caseras (que pueden ser
latas agujereadas, antenas de televisión, tubos de aluminio, entre otras),
donde se mezcla el producto con viruta de metal y ceniza de cigarrillo de
tabaco o virulana metálica a modo de filtro. Es muy común entre los sectores
más pobres por su bajo costo y fácil adquisición en las calles. Por otro lado,
es cada vez más común el consumo de "freeway o mixto", es decir
marihuana mezclada con pasta base y consumida como cigarrillo en lugar de pipa,
ya que con ello el efecto es más suave y duradero. Un error muy común es llamar
crack a la pasta base. El crack básicamente se hace con clorhidrato de cocaína
con bicarbonato de sodio y agua o amoniaco. En cambio, la pasta base es la
pasta de cocaína previa al refinamiento o a veces el sobrante de éste. Puede
decirse que la pasta base está en el paso previo a la cocaína y el crack está
en un paso posterior.
En Colombia, Venezuela y Perú se le conoce como bazuco o bazuco, ya que hace parte de un juego de
palabras "base sucia de coca". Los consumidores frecuentes
de paco, especialmente aquellos con escasos recursos económicos, suelen
cometer delitos, prostituirse o vender sus pertenencias para
obtener más de esta droga. La socióloga Alcira Daroqui, empleada en los
juzgados de menores de Quilmes, cita a una joven adicta al paco luego de
su internación: "Yo agradecí haberle dejado mi piba “hija” a mi papá,
aunque él era violento, porque si no, yo era capaz de haberla vendido".
28.5 Testimonios del bazuco en Colombia
Hace tres años
comenzó a llevar una doble vida. Todas las noches, vestido de paño, sale del
trabajo y camina hasta una esquina del barrio Garcés Navas, en el occidente de
Bogotá, para comprar unas ‘bichas’ de bazuco, que no le cuestan más de 15.000
pesos. El caso de este hombre es similar al de otros 10.000 consumidores
colombianos, que sin ser habitantes de la calle están esclavizados por una de
las drogas más destructivas que existen. La mayoría de los adictos pertenecen a
los estratos 1, 2 y 3, un sector de la población que cada día hace más
lucrativo el negocio de la venta de drogas ilegales (Serrano, 2014).
“Le coge el alma a la persona que muerde el
anzuelo y es muy difícil liberarse. Se sufre un deterioro cognitivo y cerebral
muy grave. Si se consume bazuco o bóxer, el daño es superior al que generan
otras drogas”, explica Felipe Cárdenas, antropólogo y experto en temas de
salud de la Universidad de la Sabana. El bazuco, una sustancia compuesta por
cocaína, ladrillo molido y hasta detergente, es altamente adictivo. Causa
alucinaciones, agresividad y delirio de persecución; al principio, genera una
sensación de placer y luego, de angustia (Serrano, 2014).
Según Cárdenas, “los
procesos de adicción duran toda la vida”. Eso, sumado a su bajo costo
(entre 1.000 y 2.000 pesos la dosis) y a la facilidad para adquirirla, ha hecho
que se abra camino en la clase media. Y los datos así lo indican: según el
Estudio Nacional de Consumo de Sustancias Psicoactivas en Colombia 2013, aunque
la mayoría de la población consumidora de bazuco se concentra en los estratos 1
y 2 (13.810 y 24.734 personas, respectivamente), en el 3 ya hay motivos para
alarmarse. En sus cuentas, de las 49.756 personas que usaron esa droga el año
pasado en Colombia, 6.387 pertenecen al estrato 3 y 4.825 a los estratos 4, 5 y
6 (Serrano, 2014).
Un panorama
igualmente dramático se presenta en Bogotá, donde el mercado de esta droga,
según la Fundación Ideas para la Paz (Fip), mueve al menos 508.000 millones de
pesos al año, es decir, unos 1.392 millones al día –un consumidor habitual
compra entre 10 y 20 papeletas diarias–. Aunque no hay precisión sobre las
cifras de consumidores de bazuco en los estratos 4, 5 y 6, se calcula que en
toda la ciudad hay unos 22.000, y que cerca de 2.700 son de estrato 3 (12 por
ciento). Y hay más: el Centro de Estudio y Análisis en Convivencia y Seguridad
reportó el año pasado 3.000 nuevos casos respecto del 2009. Por ejemplo
–sostiene Jonathan Nieto, subsecretario de Convivencia y Seguridad–, existen
adictos de localidades como Engativá y Suba que han terminado por vivir en la
calle (Serrano, 2014).
Ese es el caso de
David, de 37 años, quien cayó en el bazuco hace seis. Vivía con sus padres y
tenía un empleo como preparador físico, que abandonó por la droga. No le
importaba vender sus cosas o empeñarlas para conseguir unas dosis. “Salía a la
calle y me escondía de la Policía y de mi familia –recuerda–. Me daba miedo
todo. Me vestía bien, con ropa de marca y relojes finos. Cuando se me acababa
la plata, entregaba lo que fuera”. En ocasiones, la adicción viene acompañada
de delitos menores y mayores. En ciertos sectores, la droga está relacionada
estrechamente –según Nieto– con las lesiones personales, los homicidios y la
lucha territorial. Incluso, en la entrada de algunos colegios los ‘jíbaros’
regalan droga a los niños como ‘estrategia de mercado’ (Serrano, 2014).
Y la
rehabilitación, ¿qué? Pese a que la marihuana y la cocaína siguen siendo las
drogas que más se consumen en Bogotá, el número de adictos al bazuco es
crítico, y hay carencia de programas para tratarlos. Mientras el Distrito les
apostó a los Centros de Atención Móvil para Drogodependientes (Camad), con el
fin de asistir a habitantes de calle, y la Secretaría de Salud informa que hace
un trabajo de prevención en los colegios, no hay una política que abarque el
fenómeno totalmente. “No hay programas efectivos en la solución o
rehabilitación del bazuco. No hemos mejorado en su diseño porque no se ha
realizado una planeación participativa. Hay muchas personas que lo enfrentan y
no hay solución”, explica Amy Ritterbusch, profesora de la Escuela de Gobierno
de la Universidad de los Andes (Serrano, 2014).
‘Vi a niñas
vender su cuerpo por bazuco’ “Comencé a consumir marihuana a los 14 años. A los
18, conocí a alguien y me fui a vivir con él; vendía bazuco y yo le ayudaba. Me
dio curiosidad de saber qué se sentía, y probé. “Al principio, era uno, dos o
tres, y luego hasta 10 gramos en una noche”. “Cuando lo metieron a la cárcel,
yo me hundí en el bazuco; me dejó de importar todo porque me sentía sola.
Entonces, alquilaba cuartos, por 5.000 pesos la noche, para meterme la droga”.
“Al final, ya era capaz de entrar sola al sector de la L (en el centro de
Bogotá) con 200 o 300 mil pesos que conseguía trabajando o diciéndoles a mis
hermanos que necesitaba algo. Allá me los fumaba en tres o cuatro días”. “A mí
no me tocó vender mi cuerpo, pero sí vi a niñas, de unos 7 años, llevadas por
la droga y haciéndolo por unas pocas ‘bichas’”. Testimonio de Carolina, en
proceso de rehabilitación en la Fundación la Luz (Serrano, 2014).
‘Yo era estrato 6
cuando lo probé’ “Caían las 5 o 6 de la tarde y comenzaba a soplar. Lo hacía
toda la noche y me regresaba a la casa sin un peso en el bolsillo. “Empecé
fumando cigarrillos con bazuco; podía fumarme hasta 100 en tres horas. Cuando
me pasé a la pipa, bajó la frecuencia porque andaba asustado por más de 40
minutos, entonces ‘metía’ menos. “Venía de un estrato 6 en Cali, tenía plata y,
desde los 11 años, andaba en barrios pesados. Conocí la marihuana, la cocaína y
las pastillas. Todo lo pagaba yo. “Consumía entre clases. Llegó un punto en el
que ya no volví al colegio. Después me interné y duré tres años limpio. Pero
recaí y a los 19 o 20 años probé el bazuco para ver qué se sentía y solo paré
cuando la mamá de mi hija quedó embarazada, de eso hace ya cuatro años.
“Tristemente, hace cinco meses otra vez busqué la droga. Mi mamá falleció,
luego de luchar tres años contra un cáncer, y eso me dio muy duro. Caí de nuevo
en la cocaína y en el alcohol, y estuve a un paso de coger la pipa. Siento tristeza
y resentimiento conmigo mismo. “Hace mes y medio volví a un centro de
rehabilitación para curarme; aunque sé que no es fácil, no quiero volver a una
vida de perdición”. Andrés, consumidor desde hace 14 años (Serrano, 2014).
¿Qué está
pasando? Mientras para Isaac de León Beltrán, consultor de la Fundación Ideas
para la Paz (FIP), el mercado del bazuco sigue afectando básicamente a los
estratos 1 y 2 y al habitante de calle, hay quienes insisten en que el problema
está oculto en los demás estratos y que hay que prestarle atención. David
González, psicólogo de la Fundación la Luz –especializada en rehabilitación–,
cree, por su parte, que pocas veces los casos de consumo de bazuco en los
estratos 3, 4, 5 y 6 son públicos. Esto, porque algunas familias, con el ánimo
de proteger al adicto, permiten que se droguen en casa. Solo en ese centro hay
110 pacientes, en su mayoría adictos al bazuco y de estratos medios y altos.
Amy Ritterbusch, docente de la Universidad de los Andes, asegura que esta
sustancia es principalmente de la calle. Pero coincide con los otros expertos
en que se han conocido múltiples casos en otros círculos sociales (Serrano, 2014).
28.6 Anfetaminas
(YouTube) Las anfetaminas se producen sintéticamente como
moléculas que imitan la estructura y función de la dopamina y la norepinefrina
(los mismos neurotransmisores que son segregados cuando los receptores de
cocaína son estimulados). Existen muchas formas de anfetaminas, incluyendo la
deztroamfetamina y la metanfetamina, también conocida como metanfetamina
cristalizada. Al igual que la cocaína, la metanfetamina puede ser administrada
por diversos mecanismos de dosificación: inhalación de polvo, bebida del polvo
diluido, ingerida en pastillas, inyectada o fumada (Cao, Shi, Hao, Wu,
& Li, 2016; Covey, Bunner, Schuweiler, Cheer, & Garris, 2016;
Fleckenstein, Volz, Riddle, Gibb, & Hanson, 2007; Steinkellner et al.,
2014). Tres son los efectos de la metanfetamina de cristal:
👉 al igual que la cocaína bloquea el mecanismo de
reciclado de dopamina,
👉 estimula la secreción de dopamina y
👉 imita a la dopamina y la norepinefrina el cuerpo
experimenta sensación de placer físico y emocional por una parte y un estímulo
del sistema nervioso simpático por otro, al activar al simpático y su respuesta
de Combate o Escape, las capacidades físicas y cognitivas se aumentan
repentinamente por el tiempo en que el “vuelo” dure.
La sinergia de los tres efectos explica cómo es que estas drogas son más potentes que la cocaína. Las anfetaminas al ser sustancias totalmente sintéticas son más difíciles de degradar, por lo que sus efectos pueden durar por periodos más prolongados, horas en lugar de los minutos que proporciona la cocaína.

Figura 28.6. Envejecimiento acelerado. Una
combinación de mal estilo de vida, con el consumo de estas sustancias, acelera
radicalmente el proceso de envejecimiento.
Al igual que con
la cocaína, entre mayor es la potencia en que el sistema nervioso simpático es
estimulado, mas es el impacto que sufre el individuo en su fisiología
cardiovascular, que a parte de la arteriosclerosis y la vasoconstricción,
genera arrestos en el flujo cardíaco y acumulación de fluidos en los pulmones.
El síndrome de abstinencia y la anhedonia se desencadenan de forma tan marcada
como con la cocaína en la medida que el sistema nervioso aprende a regular la
sustancia para alcanzar el equilibrio nuevamente mediante la degradación de los
receptores dopaminérgicos. El síndrome de abstinencia se caracteriza por
dolores y espasmos musculares, depresión física “respiratoria” y emocional, así
como un aumento en el apetito.
La metanfetamina de cristal al igual que la
cocaína disuelve los tejidos orgánicos, acelerando el proceso de envejecimiento
biológico. Adicionalmente, la anfetamina puede cruzar la placenta y causar
efectos en el feto en desarrollo, causando partos prematuros con infantes muy
débiles y de bajo peso.
28.7 La adicción a la nicotina
(YouTube) (YouTube) La
nicotina tiene una ruta de acción diferente a la de la cocaína y anfetaminas.
Los receptores de nicotina mejoran la afinidad de los receptores de
acetilcolina en el sistema nervioso periférico, aumentando el ritmo cardíaco,
la presión sanguínea y la excitabilidad del tejido muscular. En el sistema
nervioso central los receptores de acetilcolina facilita la liberación de
dopamina y serotonina, creando sensaciones de placer y relajación (Herman, DeVito,
Jensen, & Sofuoglu, 2014; Picciotto & Kenny, 2013; Prochaska &
Benowitz, 2016; Subramaniyan & Dani, 2015). Los receptores de nicotina experimentan
desensibilización, lo cual contribuye tanto al síndrome de abstinencia agudo
como a una anhedonia crónica.
Es la nicotina la
que causa que los fumadores queden enganchados a los cigarrillos, asegurando
una exposición continúa a otras sustancias que contiene el cigarro. Aunque el
cerebro interpreta una sensación de relajación, el cuerpo realmente recibe
estímulos en el corazón y los vasos sanguíneos. Bajo la influencia de la
nicotina el corazón ate al menos 33 veces más por minuto. Al mismo tiempo los
vasos sanguíneos se constriñen, por lo que la presión sanguínea aumenta. La
nicotina también afecta las plaquetas causando que se ornen más pegajosas,
incrementando la probabilidad de formación de trombos combinados de plaqueta y
grasa LDL que puede conllevar a la enfermedad cardiovascular o arteriosclerosis
que conlleva tarde o temprano a un derrame cerebral o un ataque al corazón.
La nicotina puede ser
legal, pero también es poderosamente adictiva, casi el 95% de los fumadores son
fisiológicamente dependientes de ella, al igual que con la cocaína en la medida
que el cuerpo “vuela” también cae, por lo que cuando sus efectos pasan, los
neurotransmisores del placer y el empuje emocional para realizar las acciones
normales baja, creando una depresión que limita las actividades diarias. El
síndrome de abstinencia puede ser emocional o fisiológico, se caracteriza por
dolor muscular, espasmos, vómito, diarera, insomnio, irritabilidad entre otros.
Muchos fumadores siguen consumiendo por el solo hecho de no sentir el síndrome
de abstinencia. Los síndromes de abstinencia pueden durar desde unas cuantas
semanas hasta varios meses, lo cual explica la dificultad con la que los
adictos pueden zafarse de una adicción.
29. Adicción a los opioides
|| Regresar al Índice ||
Las moléculas de abuso generalmente se han distinguido en dos grupos principales, estimulantes del sistema nervioso central y depresores del sistema nervioso central. Los depresores del sistema nervioso central se caracterizan por una desconexión generalizada del cerebro con respecto a los órganos de los sentidos, deprimiendo las capacidades cognitivas, sensoriales y motoras. La marihuana y el alcohol son depresores del sistema nervioso central, pero sin duda las moléculas estándar en esta categoría son los opiáceos (Grenald, Largent-Milnes, & Vanderah, 2014; Moreira & Dalley, 2015; Torrens, Fonseca, Galindo, & Farré, 2015; Wikler, 2013).

Figura 29.1. Flor de amapola. El
producto de esta planta es referenciado en Game of Thrones como “leche
de la flor”, es una referencia al uso real que se le dio a la amapola por
milenios en Eurasia.
29.1 El opio
Los opiáceos son
moléculas naturales o sintéticas que afectan el cuerpo de forma similar o
mediante los mismos mecanismos moleculares que la morfina, el principal
componente activo del opio o planta de amapola "Papaver somniferum".
Los opiáceos tienen una relación de amor y odio con la medicina, por una parte,
son sustancias de peligroso abuso debido a su poderosísimo síndrome de
abstinencia, pero por otro lado son los analgésicos más fuertes conocidos por
la medicina, por lo que son empleados para aliviar dolores severos.
La morfina y la
codeína que se extraen de la planta de amapola se encuentran entre los primeros
opiáceos en ser reportados. La heroína es un derivado sintético de la morfina
con un efecto al menos el doble de potente. Las dosis de heroína y de los
opiáceos se dan regularmente mediante inyección intravenosa, pero existen
formas de dosificación mediante combustión “fumadas” o mediante inhalación por
la cavidad nasal. Independientemente del mecanismo de dosificación, la heroína
alcanza el cerebro muy rápidamente, produciendo una sensación que generalmente
es descrita en términos de éxtasis religioso y/o sexual. El cuerpo produce
moléculas opiáceas endógenas: endorfinas, enquefalinas y dinomorfinas; todas
relacionadas con la actividad física, el dolor o el miedo.
Todos los opiáceos comparten efectos semejantes: euforia, supresión del dolor y reducción de la ansiedad. Al ser depresores directos del sistema nervioso, disminuyen el ritmo respiratorio y cardiaco, lo cual puede inducir el coma o el paro cardiorrespiratorio. La sobredosis con los opiáceos es siempre un problema debido a su producción ilegal, que impide una aseguración de estándares de calidad continuos, la pureza de los agentes activos varía de acuerdo al lote de producción, por lo que la dosis administrada contiene cantidades variables y potencialmente mortales de opiáceos.

Figura 29.2. Endoopioides. Los
receptores opioides son un sistema de dendritas que permiten la disminución del
dolor, los opioides vegetales simplemente piratean este mecanismo para provocar
un estado de somnolencia de sus herbívoros para que sean más vulnerables a los
carnívoros (YouTube).
Las pupilas
extremadamente cerradas en un individuo inconsciente son un fuerte indicativo
de sobredosis por heroína. A parte del paro cardiorrespiratorio, los adictos a
la heroína se exponen a diversos peligros dependientes del mecanismo de
dosificación. Las inyecciones incrementan la probabilidad de septicemia e infecciones
de la sangre como la hepatitis y el VIH, los fumadores experimentan daños en su
tracto respiratorio y bucofaríngeo; mientras que los inhaladores experimentan
problemas en la cavidad nasal.
Los efectos
neuronales ya los hemos discutido en general en capítulos previos, estas
sustancias imitan a las endorfinas, que son narcóticos endógenos que el cuerpo
produce para disminuir el dolor después de fuertes entrenamientos. Si alguien
hace mucho ejercicio notará una leve sensación de euforia y que el cuerpo no
duele como debería, pero a la mañana siguiente al levantarse la cosa es
completamente diferente. Eso se debe al efecto de las endorfinas. Los derivados
del opio imitan a las endorfinas, y por lo tanto se caracterizan por un efecto
calmante y de euforia mucho más marcado, pero que degenera rápidamente en los
conocidos síndrome de abstinencia, tolerancia y anhedonia severas. Ahora, con
tan nocivos efectos, uno esperaría que los opioides hubieran estado muy
regulados desde hace muchos años, después de todo la planta de opio se conoce
desde hace miles de años, sin embargo, su historia es un poco más
convulsionada.
29.2 Las guerras del opio
Hubo una época en que los grandes imperios buscaron subyugar a naciones en vías de desarrollo vendiéndoles opio, y cuando estas se negaron, hubo guerra, una guerra en pro de las drogas duras. La guerra del opio fue el conflicto que mantuvieron China y Gran Bretaña, dividiéndose en dos etapas, siendo la primera entre los años 1839 y 1842; mientras que la segunda, en la que Francia se implicó con los británicos, estalló en 1856 y duró hasta 1860. El desencadenante del mismo fue la introducción en China de opio cultivado en la India y comercializado por la compañía británica de las Indias Orientales, administradora de la India. Esta sustancia (una droga estupefaciente) constituía una importante fuente de ingresos para los británicos y servía para equilibrar su balanza de pagos con China al compensar el gasto de las ingentes cantidades de té chino que Gran Bretaña importaba (Brook & Wakabayashi, 2016; Foxcroft, 2016).

Figura 29.3. Las guerras del opio.
Actualmente, comerciar con sustancias adictivas como el opio es un anatema, el
peor crimen internacional, pero hubo una época donde los imperios coloniales
traficaron, y peor aún, forzaron el comercio y consumo bajo represalia de
guerra en caso contrario, el caso más famoso fueron las guerras del opio, donde
los Chinos pelearon por su derecho a no ser envenenados (YouTube) (YouTube).
El comercio del
opio fue rechazado y prohibido por el gobierno chino. La puesta
en práctica de la abolición recayó sobre el comisionado imperial Lin Zexu.
Los emisarios enviados por los comerciantes británicos e indios quejándose por
el quebranto que tal prohibición causaba a sus intereses decidió a la Corona
británica a enviar una flota de guerra que
finalmente derrotó a la China. Como consecuencia de este
descalabro el emperador chino hubo de firmar el Tratado de Nanking,
por el que se obligaba a China al libre comercio -el del
opio incluido- con Inglaterra, a través de cinco
puertos (el más importante de ellos Cantón) así como a la cesión de
la isla de Hong Kongdurante 150 años.
29.3 Los tónicos
En el siglo XIX
el opio y la cocaína fueron empleados para la producción de tónicos milagrosos
que eran vendidos por comerciantes itinerantes, “quien haya visto el capítulo
de los Simpson sobre el tónico vigorizante del abuelo Simpson tendrá una idea
del asunto”. Aunque había tónicos de cocaína he decidido colocar el asunto en
esta sección para no saturar el capítulo de los estimulantes.
Figura 29.4. Los tónicos. Es
evidente que el tónico del abuelo (YouTube) es algún tipo de
mezcla estimulante como la coca, pero también existieron analgésicos en base a
morfina; siendo una referencia del vino Mariani o a la propia coca-cola, ambos
hechos con extracto de coca a principios del siglo XX.
Los expertos en
Historia de la Farmacia saben bien que los remedios antiguos para paliar
ciertas dolencias eran útiles a la par que de uso común. Algunos pintores
Impresionistas eran consumidores de absenta, una bebida que se consumía en los
cafés parisinos y tenía un cierto poder alucinógeno (Radenkova-Saeva,
2008). Otros destilados, tónicos y bebidas se
podían adquirir en las oficinas de farmacia o en establecimientos de
ultramarinos porque no estaban prohibidas, a pesar de que entre sus
ingredientes entraban lo que hoy conocemos como drogas duras, es decir,
la cocaína y la heroína. La Coca-Cola en sus orígenes estaría entre
estos remedios medicinales (Jay, 2015). Uno de los ejemplos más populares era
el Vino Mariani, una bebida de la que la publicidad de la época afirmaba
que restauraba la salud, la fuerza, la energía y la vitalidad, y era buena para
el cuerpo y la mente. Incluso se recomendaba para los pacientes que habían
tenido la gripe (Radenkova-Saeva,
2008).
El consumo de
este vino venía avalado porque el papa León XIII, cuyo pontificado entre 1878 y
1903 es uno de los más largos de la historia de la Iglesia, era un consumidor
habitual. Incluso algún folleto publicitario que se repartía del Vino Mariani
mostraba la imagen del pontífice. Los farmacéuticos lo recomendaban y
el público lo consumía, pero esta bebida francesa, vino-tónico a la coca del
Perú como rezaba en la etiqueta, estaba elaborada a base de vino de Burdeos y
hojas de coca. Contenía una cantidad elevada de cocaína, unos 6 miligramos por
onza líquida en la fórmula original y 7.2 miligramos en los envases que
llegaban al mercado internacional.
La cocaína era un
ingrediente fundamental en las bebidas, tónicos y tabletas comercializados en
farmacias y tiendas a finales del siglo XIX para paliar catarros, problemas de
garganta, insomnio, irritaciones de bronquios o nervios. No solo estaba
indicada para adultos, sino que también se encontraba en pastillas recetadas
para niños pequeños, píldoras para la garganta o pastillas para la rinitis
alérgica o la fiebre del heno. Y de esta droga solo
se hablaba bondades.
Casi lo mismo
ocurría con la heroína, indicada para aliviar dolores corporales, dolor de
cabeza y para niños pequeños a los que les estaban saliendo los dientes. A los
más pequeños de la casa se les administraba el popular Mrs. Winslow’s
Soothing Syrup, una bebida que contenía 65 gramos de sulfato de morfina por
cada onza líquida (Albert, 2000;
Christen & Christen, 2000; Finch, 1999). Pero este sirope, que se empezó a vender
en 1849, también causó la muerte de algunos bebes, y así algún médico lo
argumentó porque provocaba sobredosis y narcotizaba a los niños.
Sin embargo, el
opio era una sustancia habitualmente consumida por los adultos e incluso los
gobernantes, escritores y personajes influyentes presumían de ello. Éste es el
caso del Primer Ministro británico William Gladstone, que lo añadía
regularmente al café, John Keats o Florence Nightingale.
29.4 Heroína y morfina
La morfina es
una potente droga opiácea usada frecuentemente en medicina como analgésico. La morfina fue nombrada así por el
farmacéutico alemán Friedrich Wilhelm Adam Sertürner en honor a Morfeo, el dios griego de los sueños. La morfina es
un alcaloide fenantreno
del opio, siendo preparado el sulfato por neutralización
con ácido sulfúrico. Es una sustancia controlada, opioide
agonista utilizada en premedicación, anestesia, analgesia, tratamiento del
dolor asociado a la isquemia miocárdica y para la disnea asociada al fracaso
ventricular izquierdo agudo y edema pulmonar. Es un polvo blanco, cristalino,
inodoro y soluble en agua. Actualmente sigue siendo el analgésico clásico más
eficaz para aliviar los dolores agudos, pero su utilización va decreciendo a
medida que aparecen nuevas drogas sintéticas, las cuales se supone son menos
adictivas y permiten que personas alérgicas a ella puedan aliviar igualmente
sus dolores. La morfina también se usa para paliar la adicción a ciertas drogas
como la heroína y la cocaína, aunque por si misma también es fuertemente
adictiva (Boerner, Abbott,
& Roe, 1975).
La heroína,
o diacetilmorfina en su Denominación
Común Internacional, es un opioide con
propiedades analgésicas que también se utiliza de forma
menos común como supresor de la tos y antidiarreico. Por sus efectos eufóricos, la heroína se
utiliza como droga recreativa semisintética. Derivada de la morfina y originada a partir de la adormidera, planta de la que se extrae el opio, su administración frecuente y regular está asociada a la tolerancia y a
una fuerte dependencia física. En algunos países se emplea en terapias
en usuarios crónicos como sustituto de opiáceos en combinación con labores de asistencia y asesoramiento al paciente.
Administrada por vía intravenosa mediante inyección, la heroína puede ser entre
dos y cuatro veces más potente que la morfina y es más rápida en comenzar sus
efectos. La heroína ilícita suele presentarse en un polvo blanco mate al que se
añaden diversos adulterantes. Una fracción importante de los consumidores
de opioides, más de treinta millones de personas al año en todo el mundo según
la ONU, consumen heroína, la cual está relacionada
con un efecto narcótico pronunciado, se clasifica dentro de las drogas
depresoras del sistema nervioso
central y se
caracteriza por producir una dependencia psicológica y física intensa a un
ritmo muy acelerado, siendo considerada una de las drogas más adictivas y
dañinas (Boerner et al., 1975).
La heroína fue
sintetizada por primera vez por Charles Romley Alder Wright a finales del siglo XIX, que consiguió
aislarla gracias a la acetilación del clorhidrato de morfina, un producto
obtenido de la adormidera. «Heroína» fue el nombre comercial que la
empresa Bayer puso
a la nueva sustancia, que la lanzó al mercado como sustituta de la morfina, creyéndola
menos adictiva. A nivel internacional, la heroína está controlada por las
Listas I y IV de la Convención Única
sobre Estupefacientes y por lo general es ilegal su fabricación, posesión y venta sin
licencia (Akhgari,
Etemadi-Aleagha, & Jokar, 2016).
29.5 Calmantes
A partir de la
morfina es posible fabricar una amplia gama de calmantes, que como su sustancia
base, son tremendamente adictivos y causan muchas muertes (Joranson, Ryan,
Gilson, & Dahl, 2000; Pasternak, 1993; Portenoy & Foley, 1986). Ejemplos de estas sustancias son MS
Contin, Oramorph, Avinza, Kadian, Roxanol (nombre genérico: morfina); codeína;
Dolophine (nombre genérico: metadona); Opana, Opana ER, Numorpahn HCI (nombre
genérico: oximorfona); Dilaudid (nombre genérico: hidromorfona); Levo-Dromoran
(nombre genérico: levorfanol); OxyContin, OxyIR o Roxicodone (nombre genérico:
oxicodona); Demerol (nombre genérico: meperidina); Duragesic, Actiq o Fentora
(nombre genérico: fentanilo). Algunos analgésicos narcóticos combinan un
opiáceo con aspirina, paracetamol o ibuprofeno. Entre los ejemplos, se
incluyen: Percodan (nombre genérico: oxicodona y aspirina), Percocet y Roxicet
(nombre genérico: oxicodona y acetaminofeno), Vicodin, Lorcet y Lortab (nombre
genérico: hidrocodona y paracetamol)
Los opiáceos se
usan para aliviar el dolor agudo relacionado con la cirugía y otras
intervenciones médicas, así como para aliviar el dolor persistente (crónico) y
el dolor súbito de moderado a intenso. El dolor persistente se trata
habitualmente con opiáceos de acción prolongada que se liberan en el organismo
lentamente y controlan el dolor durante largos períodos. Se administran de modo
permanente siguiendo un cronograma establecido, ya sea que sientas o no dolor
en ese momento. Si se presentan eventos de dolor súbito, también puede
indicarse un segundo opiáceo de acción inmediata. Los opiáceos de acción inmediata
hacen efecto con más rapidez y no permanecen en el organismo por un tiempo
prolongado. A veces, los opiáceos se administran en combinación con no
opiáceos.
Los medicamentos
opiáceos calmantes no son malos en sí, pero deben ser tomados con mucho respeto
y bajo el cronograma estricto, una vez que se nota el desarrollo de tolerancia,
la cual se manifiesta por sentir un efecto más leve que el que se tenía antes,
debe consultarse al médico inmediatamente, y una vez que el tratamiento
finaliza, el médico receta un cronograma de desmonte paulatino, es importante
no terminar de golpe su consumo, so pena de sufrir peores dolores de su vida
vía el síndrome de abstinencia agudo, lo cual iniciaría ciclos no controlados
de consumo y en consecuencia el inicio de una adicción.
30. Adicción a los alucinógenos
|| Regresar al Índice ||
Los alucinógenos
pueden ser considerados un subtipo de estimulante si poseen efectos sobre el
sistema nervioso simpático mediante la liberación de epinefrina y
norepinefrina, a pesar de esto su característica de agrupación depende de la
fuerte desconexión que causan entre el cerebro y el sistema sensorial. El
cerebro interpreta los efectos de estas sustancias como si se tratara de
estímulos visuales o auditivos tan intensos como si se tratara de fenómenos
reales, así como alteraciones de los núcleos de recompensa a través de la
dopamina que induce sensaciones de placer físico y emocional (Harner, 1973; Nichols, 2004;
Schultes & Hofmann, 1973).
Estas drogas se
hicieron famosas en los 60s por lo que reciben un nombre más idiosincrático,
las drogas psicodélicas. En la naturaleza existen al menos seis moléculas
conocidas. Entre las más conocidas son la mezcalina que se produce a partir del
cactus de peyote; y la psilocibina que se obtiene en algunos hongos. Los hongos
son particularmente peligrosos debido a que es fácil confundir los hongos
psilocibinógenos con otros que son extremadamente venenosos. También existen
formas sintéticas más potentes como el LCD “ácido lisérgico de dietilamida”
mejor conocido como ácido; y el MDMA que se produce popularmente bajo el nombre
de éxtasis.
Estas drogas
mejoran la actividad de la serotonina, norepinefrina y acetilcolina. La ruta de
neurotransmisores placenteros es diferente, lo cual explica los efectos
alucinógenos, pero para el caso de la mezcalina su efecto sobre los receptores
de norepinefrina implican un estímulo sobre el sistema nervioso simpático
activando la respuesta de Combate o Escape. Si la molécula no imita la
norepinefrina o no estimula su receptor, los efectos fisiológicos de los drogas
psicodélicas no son particularmente dañinas para el sistema cardiorespiratorio,
pero las distorsiones de la realidad y la desconexión entre los sentidos con el
cerebro se crean comportamientos que pueden ser particularmente peligrosos.
Adicionalmente nada asegura que la alucinación sea placentera, por lo que el
consumidor puede verse encerrado en un “mal viaje”.
Los síntomas de
un “mal viaje” incluyen paranoia, pánico, depresión y confusión. La tolerancia
a las drogas alucinógenas se desarrolla de forma rápida y cruzada, por lo que
el consumo de un alucinógeno crea tolerancia a los demás.
El síndrome de
abstinencia fisiológico de las moléculas que no estimulan el sistema nervioso
simpático son poco claros, pero si se genera dependencia psicológica debido a
la estimulación de los núcleos de recompensa del hipotálamo. Entre los
estimulantes el éxtasis merece particular atención, debido a sus efectos
fisiológicos y a la facilidad de su síntesis, por lo que su consumo tiende a
popularizarse con gran rapidez. Los efectos del éxtasis incluyen una emoción de
paz interna y comunitaria, por lo que se la ha denominado la “droga de la paz”.
El éxtasis
también estimula la norepinefrina activando al sistema nervioso simpático y la
respuesta de Combate o Escape. Los individuos experimentan una resistencia
aumentada, lo cual permite bailar toda la noche, aunque sufriendo en algunas
ocasiones de deshidratación y golpe de calor. La deshidratación es
particularmente problemática ya que el alcohol es un deshidratante, cuando
ambas cosas se mezclan se puede causar la muerte por desequilibrios osmóticos
en el cerebro.
Cuando los
efectos sobre la serotonina decaen se genera una carencia de serotonina basal
desencadenando un síndrome de abstinencia caracterizado por depresión y
ansiedad, así como nausea, vómito y pereza. Otros problemas relacionados con el
éxtasis son sustancias contaminantes o aditivas que se encuentran en las
píldoras como cafeína purificada, efedrina y pseudoefedrina. En algunos casos
se han encontrado sustancias verdaderamente venenosas como la estricnina.
Referencias Bibliográficas
|| Regresar al Índice ||
Adami, H., Hsing,
A. W., McLaughlin, J. K., Trichopoulos, D., Hacker, D., Ekbom, A., &
Persson, I. (1992). Alcoholism and liver cirrhosis in the etiology of primary
liver cancer. International Journal of Cancer, 51(6), 898–902.
Akhgari, M., Etemadi-Aleagha,
A., & Jokar, F. (2016). Street Level Heroin, an Overview on Its Components
and Adulterants.
Albert, M. R. (2000).
Nineteenth-century patent medicines for the skin and hair. Journal of the
American Academy of Dermatology, 43(3), 519–526.
Alberts, B., Johnson, A.,
Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molecular
biology of the cell (5th ed.). Garland Science.
Allan, R. B. (1997).
Alcoholism, drug abuse and lawyers: Are we ready to address the denial. Creighton
L. Rev., 31, 265.
ALLIS, E. P. (1923). THE
CRANIAL ANATOMY OF CHLAMYDO‐SELACHUS ANGUINEUS. Acta Zoologica, 4(2‐3),
123–221.
Allmon, W. D. (2011). Why
don’t people think evolution is true? Implications for teaching, in and out of
the classroom. Evolution: Education and Outreach, 4(4), 648–665.
Ansel, H. C., Popovich, N. G.,
& Allen, L. V. (1995). Pharmaceutical dosage forms and drug delivery
systems. Lippincott Williams & Wilkins.
Argyropoulos, S. V, &
Nutt, D. J. (2013). Anhedonia revisited: Is there a role for dopamine-targeting
drugs for depression? Journal
of Psychopharmacology, 27(10),
869–877.
Arias Dioses, L. P. (2014). Manejo odontológico de
lesiones orales en pacientes con adicciones: revisión bibliográfica.
Universidad de Guayaquil. Facultad Piloto de Odontología.
Baker, T. B., Piper, M. E., McCarthy, D. E., Majeskie,
M. R., & Fiore, M. C. (2004). Addiction motivation reformulated: an affective
processing model of negative reinforcement. Psychological Review, 111(1),
33.
Belk, C., & Maier, V. B.
(2013). Biology Science for Life with physiology. (Pearson, Ed.) (4th
ed.).
Bell, R. L., Hauser, S. R.,
Liang, T., Sari, Y., Maldonado-Devincci, A., & Rodd, Z. A. (2017). Rat
animal models for screening medications to treat alcohol use disorders. Neuropharmacology.
Benowitz, N. L. (2008).
Clinical pharmacology of nicotine: implications for understanding, preventing,
and treating tobacco addiction. Clinical Pharmacology & Therapeutics,
83(4).
Berta, A., Sumich, J. L.,
& Kovacs, K. M. (2015). Marine mammals: evolutionary biology.
Academic Press.
Bhullar, B.-A. S., Hanson, M.,
Fabbri, M., Pritchard, A., Bever, G. S., & Hoffman, E. (2016). How to make
a bird skull: Major transitions in the evolution of the avian cranium,
paedomorphosis, and the beak as a surrogate hand. Integrative and
Comparative Biology, icw069.
Boerner, U., Abbott, S., &
Roe, R. L. (1975). The metabolism of morphine and heroin in man. Drug
Metabolism Reviews, 4(1), 39–73.
Bonomo, Y., & Proimos, J.
(2005). 8 Substance misuse: alcohol, tobacco, inhalants, and other drugs. ABC
of Adolescence, 98, 26.
Bourgois, P., & Schonberg,
J. (2007). Intimate apartheid ethnic dimensions of habitus among homeless
heroin injectors. Ethnography, 8(1), 7–31.
Bradberry, C. W. (2002). Book
Review: Dynamics of Extracellular Dopamine in the Acute and Chronic Actions of
Cocaine. The Neuroscientist, 8(4), 315–322.
Brook, T., & Wakabayashi,
B. T. (2016). The Opium Wars. Shooting Up: A Short History of Drugs and War,
58.
Brusca, R., Brusca, G. J., & Haver, N. J. (2003). Invertebrates (2nd ed.). Sinauer
Associates.
Burish, M. J., Kueh, H. Y.,
& Wang, S.-H. (2004). Brain architecture and social complexity in modern
and ancient birds. Brain, Behavior and Evolution, 63(2), 107–124.
Butler, A. B., & Hodos, W.
(2005). Comparative vertebrate neuroanatomy: evolution and adaptation.
John Wiley & Sons.
Buvinic, M., Morrison, A.,
& Orlando, M. B. (2005). Violencia,
crimen y desarrollo social en América Latina y el Caribe. Papeles de
Población, 11(43), 167–214.
Cagniard, B., Sotnikova, T. D., Gainetdinov, R. R.,
& Zhuang, X. (2014). The dopamine transporter expression level differentially
affects responses to cocaine and amphetamine. Journal of Neurogenetics, 28(1–2),
112–121.
Callaghan, R. C., Allebeck,
P., & Sidorchuk, A. (2013). Marijuana use and risk of lung cancer: a
40-year cohort study. Cancer Causes & Control, 24(10),
1811–1820.
Campbell, M. K., &
Farrell, S. O. (2009). Biochemistry (6th ed.). USA: Thomsom Brooks/Cole.
Cao, D.-N., Shi, J.-J., Hao,
W., Wu, N., & Li, J. (2016). Advances and challenges in
pharmacotherapeutics for amphetamine-type stimulants addiction. European
Journal of Pharmacology, 780, 129–135.
Carmona, J. S. S., &
Novoa, J. R. U. (2012). Composición
química de muestras de bazuco incautado en Colombia primer semestre de 2010. Revista de Salúd
Publica= Journal of Public Health, 14(6), 1010.
Castaneto, M. S., Gorelick, D.
A., Desrosiers, N. A., Hartman, R. L., Pirard, S., & Huestis, M. A. (2014).
Synthetic cannabinoids: epidemiology, pharmacodynamics, and clinical
implications. Drug and Alcohol Dependence, 144, 12–41.
Cecinato, A., Balducci, C., & Nervegna, G. (2009).
Occurrence
of cocaine in the air of the World’s cities: An emerging problem? A new tool to
investigate the social incidence of drugs? Science of the Total Environment,
407(5), 1683–1690.
Celofiga, A., Koprivsek, J.,
& Klavz, J. (2014). Use of synthetic cannabinoids in patients with
psychotic disorders: case series. Journal of Dual Diagnosis, 10(3), 168–173.
Cervantes, M. L. (2003). Características
neuropsicológicas en adultos policonsumidores de alcohol, marihuana, bazuco y
cocaína entre las edades de 20-50 años de la ciudad de Barranquilla. J.
Acosta, M. Cervantes, P. Puentes, M. Núñez, G. Jiménez, M. Sánchez, A. Berrío,
M. Zapata (Comps.), Grupo Neurociencias Del Caribe-Trayectoria Investigativa,
2005, 35–85.
Chaisson, R. E., Bacchetti,
P., Osmond, D., Brodie, B., Sande, M. A., & Moss, A. R. (1989). Cocaine use
and HIV infection in intravenous drug users in San Francisco. Jama, 261(4),
561–565.
Chapman, L. J., Chapman, J.
P., & Raulin, M. L. (1976). Scales for physical and social anhedonia. Journal
of Abnormal Psychology, 85(4), 374.
Chapman, L. J., Edell, W. S.,
& Chapman, J. P. (1980). Physical anhedonia, perceptual aberration, and
psychosis proneness. Schizophrenia Bulletin, 6(4), 639.
Christen, A. G., &
Christen, J. A. (2000). Sozodont powder dentifrice and Mrs. Winslow’s Soothing Syrup:
dental nostrums. Journal of the History of Dentistry, 48(3),
99–105.
Chuang, S.-C., La Vecchia, C.,
& Boffetta, P. (2009). Liver cancer: descriptive epidemiology and risk
factors other than HBV and HCV infection. Cancer Letters, 286(1), 9–14.
Concha, A. (2002). Impacto social y económico de la
violencia en las Américas. Biomédica, 22(Sup2), 347–361.
Covey, D. P., Bunner, K. D.,
Schuweiler, D. R., Cheer, J. F., & Garris, P. A. (2016). Amphetamine
elevates nucleus accumbens dopamine via an action potential‐dependent mechanism
that is modulated by endocannabinoids. European Journal of Neuroscience,
43(12), 1661–1673.
Cox, M. M., Doudna, J. A.,
& O’Donnell, M. (2012). Molecular biology (1st ed.). Freeman.
Creese, I. (1983). Stimulants,
neurochemical, behavioral, and clinical perspectives (Vol. 4). Raven Press
(ID).
Dai, J., Mukamal, K. J.,
Krasnow, R. E., Swan, G. E., & Reed, T. (2015). Higher usual alcohol
consumption was associated with a lower 41-y mortality risk from coronary
artery disease in men independent of genetic and common environmental factors:
the prospective NHLBI Twin Study. The American Journal of Clinical Nutrition,
102(1), 31–39.
Daniel, J. F. (1922). The
elasmobranch fishes. University of California Press.
Davies, J. A., & Graham,
J. D. P. (1980). The mechanism of action of Δ 9-tetrahydrocannabinol on body temperature in mice. Psychopharmacology,
69(3), 299–305.
Day, C. P., & Bassendine,
M. F. (1992). Genetic predisposition to alcoholic liver disease. Gut, 33(11),
1444–1447.
de Alwis, N. M. W., & Day,
C. P. (2007). Genetics of alcoholic liver disease and nonalcoholic fatty liver
disease. In Seminars in liver disease (Vol. 27, pp. 44–54). Copyright©
2007 by Thieme Medical Publishers, Inc., 333 Seventh Avenue, New York, NY 10001,
USA.
Deugnier, Y. M., Guyader, D.,
Crantock, L., Lopez, J.-M., Turlin, B., Yaouanq, J., … Halliday, J. W. (1993).
Primary liver cancer in genetic hemochromatosis: a clinical, pathological, and
pathogenetic study of 54 cases. GASTROENTEROLOGY-BALTIMORE THEN
PHILADELPHIA-, 104, 228.
Di Marzo, V., & Matias, I.
(2005). Endocannabinoid control of food intake and energy balance. Nature Neuroscience, 8(5), 585–589.
Diaz, J. H. Q., & Fernandez, M. C. O. (2013).
Consumo de marihuana en estudiantes de una universidad colombiana. Revista de Salúd
Publica= Journal of Public Health, 15(1), 32.
Donoghue, P. C. J., &
Keating, J. N. (2014). Early vertebrate evolution. Palaeontology, 57(5),
879–893.
Elman, I., Borsook, D., &
Volkow, N. D. (2013). Pain and suicidality: insights from reward and addiction
neuroscience. Progress in
Neurobiology, 109,
1–27.
ELTIEMPO.COM. (2015, August 18). Alarmante radiografía
de consumo de drogas en colegios distritales. El Tiempo. Retrieved from
http://www.eltiempo.com/bogota/estudio-sobre-consumo-de-drogas-en-colegios-de-bogota/16249537
Falk, D., Redmond, J. C.,
Guyer, J., Conroy, C., Recheis, W., Weber, G. W., & Seidler, H. (2000).
Early hominid brain evolution: a new look at old endocasts. Journal of Human
Evolution, 38(5), 695–717.
Ferré, S., & O’Brien, M.
C. (2011). Alcohol and caffeine: the perfect storm. Journal of Caffeine
Research, 1(3), 153–162.
Finch, L. (1999). Soothing
syrups and teething powders: Regulating proprietary drugs in Australia,
1860–1910. Medical History, 43(01), 74–94.
Fleckenstein, A. E., Volz, T.
J., Riddle, E. L., Gibb, J. W., & Hanson, G. R. (2007). New insights into
the mechanism of action of amphetamines. Annu. Rev. Pharmacol. Toxicol.,
47, 681–698.
Foxcroft, L. (2016). The
Making of Addiction: The’Use and Abuse’of Opium in Nineteenth-Century Britain.
Routledge.
Freeman, D., Dunn, G., Murray,
R. M., Evans, N., Lister, R., Antley, A., … Williams, J. (2014). How cannabis
causes paranoia: using the intravenous administration of∆ 9-tetrahydrocannabinol
(THC) to identify key cognitive mechanisms leading to paranoia. Schizophrenia
Bulletin, sbu098.
Freyberg, Z., Sonders, M. S.,
Aguilar, J. I., Hiranita, T., Karam, C. S., Flores, J., … Chen, A. (2016).
Mechanisms of amphetamine action illuminated through optical monitoring of
dopamine synaptic vesicles in Drosophila brain. Nature Communications, 7.
Garrick, J. A. F. (1960).
Studies on New Zealand Elasmobranchii. Part XII. The species of Squalus from
New Zealand and Australia; and a general account and key to the New Zealand
Squaloidea. In Transactions of the Royal Society of New Zealand (Vol.
88, pp. 519–557).
George, A. J. (2000). Central
nervous system stimulants. Best Practice & Research Clinical
Endocrinology & Metabolism, 14(1), 79–88.
Goodenough, J., & McGuire,
B. (2012). Biology of Humans, Concepts, Applications and Issues (4th
ed.). San Francisco: Pearson, Benjamin Cummings.
Granger, C. D., & Simon,
T. H. (1992, September 22). Misuse-resistive transdermal opioid dosage form.
Google Patents.
Grenald, S. A.,
Largent-Milnes, T. M., & Vanderah, T. W. (2014). Animal models for opioid
addiction drug discovery. Expert Opinion on Drug Discovery, 9(11),
1345–1354.
Grinspoon, L., & Bakalar,
J. B. (1981). Coca and cocaine as medicines: an historical review. Journal
of Ethnopharmacology, 3(2–3), 149–159.
Hall, B. K., & Gillis, J.
A. (2013). Incremental evolution of the neural crest, neural crest cells and
neural crest‐derived skeletal tissues. Journal of Anatomy, 222(1),
19–31.
Harner, M. J. (1973). Hallucinogens
and shamanism. Oxford U. Press.
Hartmann, T., & Ober, D.
(2000). Biosynthesis and metabolism of pyrrolizidine alkaloids in plants and
specialized insect herbivores. In Biosynthesis (pp. 207–243). Springer.
Hasin, D. S., Saha, T. D.,
Kerridge, B. T., Goldstein, R. B., Chou, S. P., Zhang, H., … Smith, S. M.
(2015). Prevalence of marijuana use disorders in the United States between
2001-2002 and 2012-2013. JAMA Psychiatry, 72(12), 1235–1242.
Herculano-Houzel, S. (2012).
The remarkable, yet not extraordinary, human brain as a scaled-up primate brain
and its associated cost. Proceedings of the National Academy of Sciences,
109(Supplement 1), 10661–10668.
Herman, A. I., DeVito, E. E.,
Jensen, K. P., & Sofuoglu, M. (2014). Pharmacogenetics of nicotine
addiction: role of dopamine. Pharmacogenomics, 15(2), 221–234.
Hermanns‐Clausen, M., Kneisel,
S., Szabo, B., & Auwärter, V. (2013). Acute toxicity due to the confirmed
consumption of synthetic cannabinoids: clinical and laboratory findings. Addiction,
108(3), 534–544.
Hillard, C. J. (2015).
Endocannabinoids and the endocrine system in health and disease. In Endocannabinoids
(pp. 317–339). Springer.
Hoefnagels, M. (2015). Biology:
concepts and investigations (3rd ed.). McGraw-Hill New York.
Hopenhayn, M. (1997). La grieta de las drogas:
desintegración social y políticas públicas en América Latina. Cepal.
Huang, Y.-H. J., Zhang, Z.-F.,
Tashkin, D. P., Feng, B., Straif, K., & Hashibe, M. (2015). An
epidemiologic review of marijuana and cancer: an update. Cancer Epidemiology
and Prevention Biomarkers, 24(1), 15–31.
Hurlburt, G. R., Ridgely, R.
C., & Witmer, L. M. (2013). Relative size of brain and cerebrum in
tyrannosaurid dinosaurs: an analysis using brain-endocast quantitative
relationships in extant alligators. Tyrannosaurid Paleobiology, 1–21.
Jay, M. (2015). Chapter
Three-Miracle or Menace?: The Arrival of Cocaine 1860–1900. International
Review of Neurobiology, 120, 27–39.
Jerison, H. J. (1969). Brain
evolution and dinosaur brains. The American Naturalist, 103(934),
575–588.
Jonathan, O. (2006). Escenarios farmacófilos de la cocaína en
Manizales:“patraseando” por la “pocaína” a la “cocaína”: del bazuco al ladruco.
revista Cultura y Droga.
Joranson, D. E., Ryan, K. M., Gilson, A. M., &
Dahl, J. L. (2000). Trends in medical use and abuse of opioid analgesics. Jama,
283(13), 1710–1714.
Justicia. (2016, December 26). Los millonarios hilos
que mueven la indigencia en el país. El Tiempo. Retrieved from
http://www.eltiempo.com/politica/justicia/cifras-de-habitantes-de-calle-en-colombia/16780702
Jylhä, K. M., Cantal, C., Akrami, N., & Milfont,
T. L. (2016). Denial of anthropogenic climate change: Social dominance
orientation helps explain the conservative male effect in Brazil and Sweden. Personality
and Individual Differences, 98, 184–187.
Kaas, J. H. (2016). of Brain
Evolution. The Wiley Handbook of Evolutionary Neuroscience, 38.
Kardong, K. V. (2011). Vertebrates,
comparative anatomy, function, evolution (6th ed.). McGraw-Hill New York.
Kardong, K. V. (2014). Vertebrates:
Comparative Anatomy, Function, Evolution (7th ed.). McGraw-Hill Education.
Karp, G. C. (2013). Cell
and Molecular Biology, Concepts and Experiments (7th ed.). USA: Wiley
Online Library.
Kerr, T., Tyndall, M., Li, K.,
Montaner, J., & Wood, E. (2005). Safer injection facility use and syringe
sharing in injection drug users. The Lancet, 366(9482), 316–318.
la Haye, K., Green, H. D.,
Kennedy, D. P., Pollard, M. S., & Tucker, J. S. (2013). Selection and
influence mechanisms associated with marijuana initiation and use in adolescent
friendship networks. Journal of Research on Adolescence, 23(3),
474–486.
Larsson, H. C. E., Sereno, P.
C., & Wilson, J. A. (2000). Forebrain enlargement among nonavian theropod
dinosaurs. Journal of Vertebrate Paleontology, 20(3), 615–618.
Launchbaugh, K. L., Provenza,
F. D., & Pfister, J. A. (2001). Herbivore response to anti-quality factors
in forages. Journal of Range Management, 431–440.
Levin, D. A. (1976). The
chemical defenses of plants to pathogens and herbivores. Annual Review of
Ecology and Systematics, 7(1), 121–159.
Lewandowsky, S., Oreskes, N.,
Risbey, J. S., Newell, B. R., & Smithson, M. (2015). Seepage: Climate change
denial and its effect on the scientific community. Global Environmental Change, 33, 1–13.
López-Quintero, C., & Neumark, Y. (2012).
Membresía a barras bravas y su influencia en el uso de drogas. Revista
Peruana de Medicina Experimental y Salud Pública, 29(1), 21–27.
Lovette, I. J., & Fitzpatrick, J. W. (2016). Handbook of bird
biology. John Wiley & Sons.
Mackean, D. G., & Hayward,
D. (2014). Biology (3rd ed.). IGCSE Cambridge.
Mader, S. S. (2010). Biology
(10th ed.). McGraw-Hill Education.
Mader, S. S., &
Windelspecht, M. (2015). Biology (12th ed.). McGraw-Hill Education.
Mader, S. S., &
Windelspecht, M. (2018). Essentials of biology (5th ed.). McGraw-Hill
Education.
Maertens, R. M., White, P. A.,
Rickert, W., Levasseur, G., Douglas, G. R., Bellier, P. V, … Desjardins, S.
(2009). The genotoxicity of mainstream and sidestream marijuana and tobacco
smoke condensates. Chemical Research in Toxicology, 22(8),
1406–1414.
Marieb, E. N., Wilhelm, P. B.,
& Mallatt, J. (2017). Human Anatomy (8th ed.). Pearson.
Marino, L., Uhen, M. D.,
Pyenson, N. D., & Frohlich, B. (2003). Reconstructing cetacean brain
evolution using computed tomography. The Anatomical Record, 272(1),
107–117.
Markowitz, G., & Rosner,
D. (2013). Deceit and denial: The deadly politics of industrial pollution.
Univ of California Press.
Marselos, M., &
Karamanakos, P. (1999). Mutagenicity, developmental toxicity and
carcinogenicity of cannabis. Addiction Biology, 4(1), 5–12.
Martin, B. R., & Wiley, J.
L. (2003). Mechanism of action of cannabinoids: how it may lead to treatment of
cachexia, emesis, and pain. The Journal of Supportive Oncology, 2(4),
305–314.
Martínez-González, J. M.,
Vilar López, R., Becoña Iglesias, E., & Verdejo-García, A. (2016).
Self-deception as a mechanism for the maintenance of drug addiction. Psicothema, 28(1).
MARTÍNEZ, I. P., & AMAR, J. A. (n.d.).
DISTORSIONES COGNITIVAS EN LOS GRUPOS DE DROGODEPENDIENTES CON DIAGNÓSTICO
DUAL.
Martínez Ruiz, M., & Rubio Valladolid, G. (2002). Manual
de drogodependencias para enfermería. Ediciones Díaz de Santos.
Mason, K. A., Losos, J. B., Singer, S. R., &
Raven, P. H. (2014). Biology (7th ed.). McGraw-Hill New York.
Matsumoto, C., Miedema, M. D.,
Ofman, P., Gaziano, J. M., & Sesso, H. D. (2014). An expanding knowledge of
the mechanisms and effects of alcohol consumption on cardiovascular disease. Journal
of Cardiopulmonary Rehabilitation and Prevention, 34(3), 159–171.
McBride, W. J., Murphy, J. M.,
Lumeng, L., & Li, T.-K. (1990). Serotonin, dopamine and GABA involvement in
alcohol drinking of selectively bred rats. Alcohol, 7(3),
199–205.
McKim, W. A., & Hancock,
S. (2012). Drugs & Behavior. Pearson Higher Ed.
Mei, W., Zhang, J. X., &
Xiao, Z. (2010). Acute effects of sublingual buprenorphine on brain responses
to heroin-related cues in early-abstinent heroin addicts: an uncontrolled
trial. Neuroscience, 170(3), 808–815.
Mejía, I. E., & Gómez, A. P. (2005). La inyección
de drogas en Bogotá: una amenaza creciente. Adicciones, 17(3),
251–260.
Mendoza, A., Romero, J. F. R., Mendoza, G. A. G.,
& Posada, I. C. (2015). Significados del consumo de sustancias psicoactivas
en indígenas estudiantes de una universidad de Medellín, Colombia. Universitas
Psychologica, 14(3).
Mesa, A. F. C., & Bello, S. M. L. (2015). Cannabis
sintético: aspectos toxicológicos, usos clínicos y droga de diseño. Revista
de La Facultad de Medicina, 63(3), 501.
Mithöfer, A., & Boland, W. (2012). Plant defense
against herbivores: chemical aspects. Annual Review of Plant Biology, 63,
431–450.
Moreira, F. A., & Dalley,
J. W. (2015). Dopamine receptor partial agonists and addiction. European
Journal of Pharmacology, 752, 112–115.
Morrow, A. L., Montpied, P.,
Lingford-Hughes, A., & Paul, S. M. (1990). Chronic ethanol and
pentobarbital administration in the rat: effects on GABA A receptor function
and expression in brain. Alcohol, 7(3), 237–244.
Most, D., Ferguson, L., &
Harris, R. A. (2014). Molecular basis of alcoholism. Handbook of Clinical
Neurology, 125, 89.
Murray, R. K., Bender, D. A.,
Botham, K. M., Kennelly, P. J., Rodwell, V., & Weil, A. (2012). Harpers
Illustrated Biochemistry (29th ed.). McGraw-Hill Medical.
Musshoff, F., Madea, B.,
Kernbach-Wighton, G., Bicker, W., Kneisel, S., Hutter, M., & Auwärter, V.
(2014). Driving under the influence of synthetic cannabinoids (“Spice”): a case
series. International Journal of Legal Medicine, 128(1), 59–64.
Naguib, M., & Foss, J. F.
(2015). Medical use of marijuana: truth in evidence. LWW.
Neumark, Y. D., Delva, J.,
& Anthony, J. C. (1998). The epidemiology of adolescent inhalant drug
involvement. Archives of Pediatrics & Adolescent Medicine, 152(8),
781–786.
Nichols, D. E. (2004).
Hallucinogens. Pharmacology & Therapeutics, 101(2), 131–181.
Northcutt, R. G. (1984).
Evolution of the vertebrate central nervous system: patterns and processes. American
Zoologist, 24(3), 701–716.
Northcutt, R. G. (2001).
Changing views of brain evolution. Brain Research Bulletin, 55(6),
663–674.
Northcutt, R. G. (2002).
Understanding Vertebrate Brain Evolution1. Integrative and Comparative
Biology, 42(4), 743–756.
O’shea, R. S., Dasarathy, S.,
& McCullough, A. J. (2010). Alcoholic liver disease. Hepatology, 51(1),
307–328.
Olin, J. T., Schneider, L. S.,
Katz, I. R., Meyers, B. S., Alexopoulos, G. S., Breitner, J. C., … Devanand, D.
P. (2002). Provisional diagnostic criteria for depression of Alzheimer disease.
The American Journal of Geriatric Psychiatry, 10(2), 125–128.
Oreskes, N., & Conway, E.
M. (2011). Merchants of doubt: How a handful of scientists obscured the
truth on issues from tobacco smoke to global warming. Bloomsbury Publishing
USA.
Panksepp, J., Knutson, B.,
& Burgdorf, J. (2002). The role of brain emotional systems in addictions: a
neuro‐evolutionary perspective and new ‘self‐report’animal model. Addiction,
97(4), 459–469.
Parmar, J. R., Forrest, B. D.,
& Freeman, R. A. (2016). Medical marijuana patient counseling points for
health care professionals based on trends in the medical uses, efficacy, and
adverse effects of cannabis-based pharmaceutical drugs. Research in Social
and Administrative Pharmacy, 12(4), 638–654.
Pasternak, G. W. (1993).
Pharmacological mechanisms of opioid analgesics. Clinical Neuropharmacology, 16(1), 1–18.
Peruaga, A., Rincón, A., & Selin, H. (2002). El
consumo de sustancias adictivas en las Américas. Adicciones, 14(2),
227–238.
Pesta, D. H., Angadi, S. S.,
Burtscher, M., & Roberts, C. K. (2013). The effects of caffeine, nicotine,
ethanol, and tetrahydrocannabinol on exercise performance. Nutrition &
Metabolism, 10(1), 71.
Picciotto, M. R., & Kenny,
P. J. (2013). Molecular mechanisms underlying behaviors related to nicotine
addiction. Cold Spring Harbor Perspectives in Medicine, 3(1),
a012112.
Pickard, H. (2016). Denial in
addiction. Mind & Language, 31(3), 277–299.
Pollard, T. D., Earnshaw, W.
C., Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell Biology
(3rd ed.). Elsevier.
Portenoy, R. K., & Foley,
K. M. (1986). Chronic use of opioid analgesics in non-malignant pain: report of
38 cases. Pain, 25(2), 171–186.
Prausnitz, M. R., &
Langer, R. (2008). Transdermal drug delivery. Nature Biotechnology, 26(11),
1261–1268.
Prochaska, J. J., &
Benowitz, N. L. (2016). The past, present, and future of nicotine addiction
therapy. Annual Review of Medicine, 67, 467–486.
Puig, S., & Gutstein, H.
B. (2017). Opioids: keeping the good, eliminating the bad. Nature Medicine.
Radenkova-Saeva, J. (2008).
Recreational Drugs and its Impact on Music Literature and Art. Biotechnology
& Biotechnological Equipment, 22(2), 656–659.
Rader, D. J., & Hovingh,
G. K. (2014). HDL and cardiovascular disease. The Lancet, 384(9943),
618–625.
Reece, J. B., Urry, L. A.,
Wasserman, S. A., Cain, M. L., Minorsky, P. V, & Jackson, R. B. (2014). Campbell
Biology (10th ed.). Pearson Higher Ed.
Rey, J. M., Martin, A., &
Krabman, P. (2004). Is the party over? Cannabis and juvenile psychiatric
disorder: the past 10 years. Journal of the American Academy of Child &
Adolescent Psychiatry, 43(10), 1194–1205.
Richardson, J. L., &
Illum, L. (1992). (D) Routes of delivery: Case studies:(8) The vaginal route of
peptide and protein drug delivery. Advanced Drug Delivery Reviews, 8(2–3),
341–366.
Rinn, W., Desai, N.,
Rosenblatt, H., & Gastfriend, D. R. (2002). Addiction denial and cognitive
dysfunction: a preliminary investigation. The Journal of Neuropsychiatry and
Clinical Neurosciences, 14(1), 52–57.
Ritchison, G. (2015). Bird
biogeography. RITCHISON, G. Avian Biology: Everything You Wanted to Know
about Birds… and More. Eastern Kentucky University,[Sd]. Disponível Em:<
Http://People. Eku. Edu/Ritchisong/Birdbiogeography1. Htm>. Acesso Em, 9.
Ritz, M. C., Lamb, R. J.,
Goldberg, S. R., & Kuhar, M. J. (1987). Cocaine receptors on dopamine
transporters are related to self-administration of cocaine. Science, 237,
1219–1224.
Roberto, M., Madamba, S. G.,
Moore, S. D., Tallent, M. K., & Siggins, G. R. (2003). Ethanol increases
GABAergic transmission at both pre-and postsynaptic sites in rat central
amygdala neurons. Proceedings of the National Academy of Sciences, 100(4),
2053–2058.
Rogers, S. W. (1999).
Allosaurus, crocodiles, and birds: evolutionary clues from spiral computed
tomography of an endocast. The Anatomical Record, 257(5), 162–173.
Rojas Valero, M. (2015). Pasta Básica de Cocaína:
cuatro décadas de historia, actualidad y desafíos.
Rosslenbroich, B. (2014).
Nervous Systems and the Flexibility of Movements. In On the Origin of
Autonomy (pp. 131–147). Springer.
Rossum, J. M. van, Schoot, J.
B., & Hurkmans, J. A. T. M. (1962). Mechanism of action of cocaine and
amphetamine in the brain. Cellular and Molecular Life Sciences, 18(5),
229–231.
Sabogal-Carmona, J. S., &
Urrego-Novoa, J. R. (2012). Quantifying the chemical composition of
crack-cocaine (bazuco) samples seized in Colombia during the first half of
2010. Revista de Salud
Pública, 14(6),
1014–1025.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of
Biology (10th ed.). Sinauer & MacMillian.
Sadler, T. W. (2012). Medical
Embryology. (C. Taylor, Ed.) (12th ed.). Baltimore: Lippincott Williams
& Wilkins.
Sakar, S. (2009). Doubting
Darwin?: Creationist designs on evolution. John Wiley & Sons.
Savage, R. J., King, V. L.,
Clark, C. B., & Cropsey, K. L. (2017). Factors associated with early
marijuana initiation in a criminal justice population. Addictive Behaviors,
64, 82–88.
Schneider, M. (2008). Puberty
as a highly vulnerable developmental period for the consequences of cannabis
exposure. Addiction Biology, 13(2), 253–263.
Schultes, R. E., &
Hofmann, A. (1973). The botany and chemistry of hallucinogens. Springfield,
Illinois.: Charles C. Thomas 267pp.. Hallucinogens: Fungi.
Schütz, C. G., Chilcoat, H.
D., & Anthony, J. C. (1994). The association between sniffing inhalants and
injecting drugs. Comprehensive
Psychiatry, 35(2),
99–105.
Scoppetta Díaz-Granados, O., Pérez Gómez, A., &
Muñoz, V. H. (2013). Sobre la supuesta inocuidad del consumo de marihuana:
diferencia entre consumidores y no consumidores en encuestas nacionales en
Colombia. Liberabit, 19(1), 55–66.
Scott, E. C. (1997).
Antievolution and creationism in the United States. Annual Review of Anthropology, 26(1), 263–289.
Serrano, A. P. (2014, September 1). Adicción al bazuco
va más allá de los estratos 1 y 2. El Tiempo. Retrieved from
http://www.eltiempo.com/bogota/relatos-de-consumidores-de-bazuco/14462035
Shah, V. P., Maibach, H. I.,
& Jenner, J. (1993). Topical drug bioavailability, bioequivalence, and
penetration. Springer.
Shier, D., Butler, J., &
Lewis, R. (2015). Hole’s Human Anatomy & Physiology (14th ed.).
McGraw-Hill Education.
Simon, E. J., Dickey, J. L.,
& Reece, J. B. (2013). Essential biology (Pearson).
Solanto, M. V. (2001). Stimulant
drugs and ADHD: Basic and clinical neuroscience. Oxford University Press,
USA.
Solomon, E., Martin, C.,
Martin, D. W., & Berg, L. R. (2014). Biology (10th ed.). Cengage
Learning.
Starr, C., Evers, C., &
Starr, L. (2013). Biology: Today and Tomorrow With Physiology (4th ed.).
Brooks/Cole.
Steinkellner, T., Mus, L.,
Eisenrauch, B., Constantinescu, A., Leo, D., Konrad, L., … Kong, E. (2014). In
vivo amphetamine action is contingent on αCaMKII. Neuropsychopharmacology, 39(11),
2681–2693.
Stern, K. R., Bidlack, J. E.,
& Jansky, S. H. (2008). Introductory Plant Biology (11th ed.).
McGraw-Hill New York.
Subramaniyan, M., & Dani,
J. A. (2015). Dopaminergic and cholinergic learning mechanisms in nicotine
addiction. Annals of the New York Academy of Sciences, 1349(1),
46–63.
Takeda, S. (2014). Δ9-Tetrahydrocannabinol targeting estrogen receptor
signaling: the possible mechanism of action coupled with endocrine disruption. Biological
and Pharmaceutical Bulletin, 37(9), 1435–1438.
Tashkin, D. P. (1990).
Pulmonary complications of smoked substance abuse. Western Journal of
Medicine, 152(5), 525.
Tashkin, D. P. (2001). Airway
effects of marijuana, cocaine, and other inhaled illicit agents. Current
Opinion in Pulmonary Medicine, 7(2), 43–61.
Taylor, M. R., Simon, E. J.,
Dickey, J. L., Hogan, K., & Reece, J. B. (2018). Biology, Concepts and
conections (9th ed.). Boston: Pearson education.
Tiffany, S. T., Cox, L. S.,
& Elash, C. A. (2000). Effects of transdermal nicotine patches on
abstinence-induced and cue-elicited craving in cigarette smokers. Journal of
Consulting and Clinical Psychology, 68(2), 233.
Torrens, M., Fonseca, F.,
Galindo, L., & Farré, M. (2015). Opioid Addiction: Short-and Long-Acting
Opioids. Textbook of Addiction Treatment: International Perspectives,
467–500.
Tortajada, R. E., San Miguel,
J. P., Domenech, M. G., Cucarella, J. O., & Aliño, M. (2015). Drogas legales emergentes: marihuana
sintética, kratom, salvia divinorum, metoxetamina y los derivados de la
piperazina. Revista Española de Drogodependencias, (1), 46–61.
Tsochatzis, E. A., Bosch, J., & Burroughs, A. K.
(2014). Liver cirrhosis. The Lancet, 383(9930), 1749–1761.
Ubeda-Bañon, I., Pro-Sistiaga, P., Mohedano-Moriano,
A., Saiz-Sanchez, D., de la Rosa-Prieto, C., Gutierrez-Castellanos, N., … Martinez-Marcos, A.
(2014). Cladistic analysis of olfactory and vomeronasal systems. Adaptive
Function and Brain Evolution, 193.
Vandrey, R. G., & Budney,
A. J. (2015). Cannabis Use Disorders. Encyclopedia of Psychopharmacology,
336–341.
Vasiliou, V., & Pappa, A.
(2000). Polymorphisms of human aldehyde dehydrogenases. Pharmacology, 61(3),
192–198.
Vasiliou, V., Pappa, A., &
Estey, T. (2004). Role of human aldehyde dehydrogenases in endobiotic and
xenobiotic metabolism. Drug Metabolism Reviews, 36(2), 279–299.
Vitellone, N. (2015). Syringe
sociology. The British Journal of Sociology, 66(2), 373–390.
Vitt, L. J., & Caldwell,
J. P. (2014). Herpetology An Introductory Biology of Amphibians and Reptiles
(4th ed.). Oklahoma: Elsevier Inc.
Volkow, N. D., Baler, R. D.,
Compton, W. M., & Weiss, S. R. B. (2014). Adverse health effects of
marijuana use. New England Journal of Medicine, 370(23),
2219–2227.
Watters, J. K., Estilo, M. J.,
Clark, G. L., & Lorvick, J. (1994). Syringe and needle exchange as HIV/AIDS
prevention for injection drug users. Jama, 271(2), 115–120.
Wayne, R. (2009). Plant
Cell Biology (1st ed.). San Diego: Elsevier.
Weaver, R. F. (2012). Molecular
biology (5th ed.). McGraw-Hill Education.
Weil, A., & Rosen, W.
(1983). Chocolate to morphine. Houghton Mifflin Company.
Wiesner, C., & Peñaranda, D. (2002). Encuesta
mundial de tabaquismo en jóvenes reporte de Bogotá, Colombia. Revista Colombiana
de Cancerología, 6(4), 5–14.
Wikler, A. (2013). Opioid
dependence: Mechanisms and treatment. Springer Science & Business
Media.
Wise, R. A. (1984). Neural
mechanisms of the reinforcing action of cocaine. Cocaine: Pharmacology,
Effects, and Treatment of Abuse. National Institute on Drug Abuse Research
Monograph, 50, 15–33.
Wise, R. A., & Koob, G. F.
(2014). The development and maintenance of drug addiction. Neuropsychopharmacology,
39(2), 254–262.
Yasui, Y., & Su, T.-P.
(2016). Potential Molecular Mechanisms on the Role of the Sigma-1 Receptor in
the Action of Cocaine and Methamphetamine. Journal of Drug and Alcohol
Research, 5.
Yoshida, A., Rzhetsky, A.,
Hsu, L. C., & Chang, C. (1998). Human aldehyde dehydrogenase gene family. European
Journal of Biochemistry, 251(3), 549–557.
Young, S. J., Longstaffe, S.,
& Tenenbein, M. (1999). Inhalant abuse and the abuse of other drugs. The
American Journal of Drug and Alcohol Abuse, 25(2), 371–375.
Zhang, L. R., Morgenstern, H.,
Greenland, S., Chang, S., Lazarus, P., Teare, M. D., … Brhane, Y. (2015).
Cannabis smoking and lung cancer risk: Pooled analysis in the International
Lung Cancer Consortium. International
Journal of Cancer, 136(4),
894–903.
No hay comentarios:
Publicar un comentario