
9. Sistema esquelético en esponjas
|| Regresar al Índice ||
Los elementos esqueléticos de las esponjas vienen en dos tipos, que se puede clasificar como orgánicos e inorgánicos. El esqueleto orgánico está construido por fibras de colágeno, mientras que el segundo se puede formar mediante la sedimentación de dióxido de silicio y carbonato de calcio. Las esponjas son las únicas que emplean dióxido de silicio como parte de sus sistemas esqueléticos.

Figura 9.1. En la imagen
anterior podemos ver algunos modelos de la estructura de las esponjas, la parte
dedicada al esqueleto de las esponjas está en amarillo, sirve como punto de
anclaje para los coanocitos y los amebocitos.
El colágeno es la principal proteína
estructural de los invertebrados –y también hace parte del tejido conectivo de
los mamíferos dando tención a la piel, la pérdida del colágeno en la piel se
asocia a la aperción de arrugas durante el envejecimiento. En las esponjas las
fibras de colágeno pueden presentarse de forma dispersa en la matriz del
citoplasma o como unas fibras que mantienen unidas a las células de la esponja
en una estructura gelatinosa llamada espongina.
La espongina verdadera es encontrada
únicamente en los miembros de la clase Demospongiae,
mientras que las fibras de colágeno disperso están distribuidas en todos los
miembros del filum Porifera. La
cantidad de colágeno disperso en el citoplasma depende mucho de la especie, y
esto afecta la densidad y rigidez de la esponja, a mayor colágeno, la esponja
será más tensa, a menos colágeno la esponja se hace más gelatinosa.
Clásicamente el esqueleto orgánico de las esponjas ha sido denominado espongina, este término debe ser empleado en sentido estricto a la forma de colágeno que se presenta por fuera de la célula formando una red como si fuera un citoesqueleto externo. La región que rodea a las células de la esponja se llama mesófilo, y solo el grupo de las demosponjas poseen espongina en su mesófilo. La red de espongina puede cristalizar minerales como espículas de dióxido de silicio a su estructura haciéndola aún más rígida. La espongina también se encuentra presente en el caparazón de una estructura de reproducción asexual muy resistente llamada gémula. Las gémulas son producidas principalmente por esponjas de agua dulce tropical y se general usualmente al final de la estación lluviosa para resistir durante la estación seca. En la imagen siguiente tenemos fibras de espongina vistas al microscopio electrónico

Figura 9.2. La parte dura de la esponja puede transformarse en estructuras de defensa, por ejemplo, en la punta superior de las esponjas se proyectan estructuras en forma de agujas llamadas espículas.

Figura 9.3. Los esqueletos
minerales de dióxido de silicio o carbonato de calcio se encuentran
virtualmente en todas las esponjas, excepto en la clase de las Demospongiae
“que poseen el esqueleto de espongina de origen orgánico”.
9.1 Esqueleto
mineralizado de las esponjas
En la mayoría de las esponjas una serie de
células del mesófilo se especializan para formar estructuras de anclaje a los
minerales, a estas células las denominamos esclerocitos. Los esclerocitos
también segregan el carbonato de calcio o dióxido de silicio mineralizando la
estructura que pasa a denominarse espícula. En algunas ocasiones, un solo
esclerocito es capaz de producir una sola espícula; en otras ocasiones se
necesitan varios esclerocitos para sintetizar una sola espícula –generalmente
dos células por espícula. La construcción de la espícula de dióxido de silicio
comienza con la secreción de un filamento orgánico que sirve como eje de la
espícula, posteriormente el dióxido de silicio hidratado es segregado es
depositado en el filamento. En el caso de las espículas de carbonato de calcio,
estas se sintetizan in situ sin la necesidad de un marco orgánico que sirva
como eje de construcción. Estas espículas calcáreas son producidas
intracelularmente, en este sentido cada espícula calcárea es en esencia un solo
cristal de calcita o aragonita siendo sedimentado por la célula.

Figura 9.4. Cristales de
Calcita (arriba) y cristales de aragonita (abajo), ambos son creados por la
misma molécula, el carbonato de calcio precipitado. Por lo general la aragonita
es menos estable y con el tiempo sus cristales se transforman en calcita.
La morfología de la espícula parece estar
relacionada con la evolución de las esponjas, por lo que la nomenclatura
empleada para clasificarlas –que es bastante compleja, como todo lo que
proviene de la era antes del siglo XX –aún tiene peso taxonómico y filogenético
–ayuda a clasificar y a elaborar hipótesis sobre las relaciones de parentesco
entre los diferentes grupos de esponjas respectivamente.
De acuerdo con su morfología general –tamaño –las espículas se clasifican como microscleras y megascleras. Las microscleras son espículas pequeñas que sirven como refuerzo estructural. Las megascleras por el contrario son espículas grandes. Las demosponjas y los hexactinelidos poseen ambos tipos de espículas; mientras que las esponjas calcáreas solo poseen megascleras.

Figura 9.5. El esqueleto espicular debe ser visto como una estructura de soporte suplementario. Si la cantidad de material inorgánico aumenta con respecto al material orgánico, la esponja se calcifica hasta que su textura se aproxima a la de una roca como en el caso de los órdenes Choristida y Lithistida.

Figura 9.6. Las espículas
pequeñas son llamadas microscleras y son microscópicas, por lo que deben
visualizarse con microscopios. Las imagenes presentadas aquí fueron obtenidas
con microscopio electrónico.
Del mismo modo que sucede en las plantas o
en el concreto, algunas esponjas introducen fibras de polisacáridos al interior
del cristal de calcita o aragonita, esto proporciona al esqueleto calcáreo
mayor flexibilidad y menor peso sin sacrificar la resistencia del mineral. Una
ventaja adicional reside en que la presencia de fibras orgánicas le permite al
organismo moldear la espícula de muchas formas, y no solo como picos
cristalinos.
9.2 Usos del
esqueleto de las esponjas
Las especies de esponjas que carecen de los esqueletos minerales tienden a tener una textura suave, aunque fibrosa, debido a que solo cuentan con el colágeno como su mecanismo de rigidez. Estas esponjas fueron generalmente empleadas para el baño o el lavado de los utensilios de cocina por parte de algunas culturas. En la actualidad todas las esponjas que compramos son imitaciones sintéticas del esqueleto de espongina.

Figura 9.7. Las primeras
esponjas sintéticas al estar hechas de pulpa incrementaron la necesidad de
talar bosques para obtener la materia prima. Esponjas sintéticas recientes se
realizan con plásticos, pero en este caso la fuente del material es el
petróleo, adicionalmente liberan carcinógenos –sustancias que favorecen la
generación de cáncer –durante su uso, fabricación y deposición, adicionalmente
no se biodegradan adecuadamente.
Las esponjas han sido empleadas por
milenios por parte de los seres humanos como instrumentos de limpieza; Homero y
otros escritores griegos presocráticos mencionan que el comercio de esponjas
era un negocio lucrativo a través del Mediterráneo esto aproximadamente en el
siglo VIII antes de Cristo. La industria alcanzó su pico en 1938 cuando la
captura anual de esponjas –incluyendo las esponjas criadas en cautiverio –
excedió los 2.6 millones de libras. De estas 700.000 provenían de las costas de
Estados Unidos y las Bahamas. La mayoría de las esponjas comercialmente
explotadas pertenecían a los géneros Hippospongia “imagen siguiente” y Spongia.
Las consecuencias fueron previsibles, la practica extinción de las especies
pertenecientes a estos dos géneros tanto en el mar Mediterráneo como en el mar
Caribe cercano a la Florida.
Las esponjas artificiales fueron
introducidas en la década de 1940 por el gigante de la síntesis de materiales,
la compañía Du Pont –líder en la manufactura de materiales sintéticos como el
nylon “Figura 44”. Estas primeras esponjas fueron hechas de pulpa de celulosa
–la misma que se emplea para hacer papel –aunque el proceso permaneció bajo
secreto industrial hasta 1950. Para la década de los 70s las esponjas
artificiales habían reemplazado a las esponjas naturales, lo cual sin duda ha
salvado a muchas especies de la extinción, pero al mismo tiempo ha generado sus
propios problemas.
10. Sistema esquelético cnidarios
|| Regresar al Índice ||
Los Cnidarios –medusas, pólipos de coral e hidras –emplean una amplia variedad de mecanismos de soporte, anclaje y protección, en parte debido a que poseen dos formas muy diferentes en su ciclo de vida, el pólipo anclado a un sustrato y la medusa de vida libre. Los pólipos en sí mismos, así como las medusas deben confiar en el esqueleto básico de todos los invertebrados, el esqueleto hidrostático. Los Atozoos “Anthozoa” es un uno de los principales grupos de los cnidarios, generalmente nos referimos a ellos como los pólipos constructores de coral debido a que aparte de su esqueleto hidrostático, segregan un saco de quitina que pronto se mineraliza gracias a una simbiosis establecida con un protista del tipo de los dinoflagelados llamada zooxantela. Las especies de zooxantelas son fotosintéticas, emplean los desechos nitrogenados y el dióxido de carbono del pólipo junto con la energía del sol para producir azúcares que son compartidos con el pólipo –simbiosis mutualista. Por tal razón los corales solo pueden crecer en aguas someras muy iluminadas.

Figura 10.1. Los cnidarios tienen dos modos de vida, la medusa y el pólipo. Mientras que el esqueleto de la medusa es hidrostático, los pólipos pueden generar exoesqueletos duros y muy complejos llamados corales. Los corales poseen una amplia diversidad de formas a nivel macroscópico, aunque todos emergen de la misma combinación, un pólipo y su saco o exoesqueleto protector.

Figura 10.2. La mesoglea
(gris) es el tejido medio de los cnidarios, en las medusas (izquierda) es mucho
más importante y ocupa una mayor parte del volumen del cuerpo. Los pólipos
(derecha) no recurren demasiado a ella como mecanismo de soporte, ya sea porque
generan exoesqueletos como los antozoos, o porque se trata de una etapa del
ciclo de vida que no es permanente como en las medusas jóvenes.
El conjunto del pólipo y la zooxantela producen un material adicional a la quitina y es carbonato de calcio, el cual se cristaliza rápidamente endureciendo el saco y formando el coral duro. Cuando muchos pólipos crecen junto formando una enorme masa de sacos de carbonato de calcio se los denomina en conjunto arrecife de coral, cada especie de pólipo tiene una forma diferente de construir su saco, y por lo tanto la forma del arrecife cambia. Por lo general los pólipos formadores de corales “Athozoos” no poseen una forma de vida libre o medusa, pero las medusas si tienen vida anclada a sustrato. Cuando una medusa “Medusae” alcanza su etapa de vida libre “la medusa” ella debe confiar exclusivamente en su esqueleto hidrostático, pero adicionalmente en la capa media o mesoglea se presenta una modificación, la capa puede ser flexible o endurecida por mesénquima fibroso hasta el punto de tener la consistencia de un cartílago.

Figura 10.3. Los llamamos esqueletos axiales
porque forman ejes o tubos, en el caso de los gorgonianos son tubos
ramificados. En los Antipatharios –corales negros –el esqueleto axial es tan
duro y denso que puede ser empleado para hacer joyería mediante tallado.
10.1 Detalles
de los esqueletos coralinos axiales y escleróticos
La dureza o textura de las estructuras
esqueléticas duras de lo cnidarios vienen en tres tipos particulares:
esqueletos axiales callosos semejantes a la madera; esclerosas calcáreas, y
marcos conglomerantes masivos.
👉 Esqueletos
axiales: Los esqueletos axiales
callosos se presentan en muchos grupos de antozoos coloniales –pólipos
formadores de coral – como los gorgonianos, plumas de mar y corales antipatharianos.
Las células encargadas se segregar el marco de quitina son los amebocitos
presentes en el coenenquima. Las barras axiales son proteínas –complejos de
mucopolisacáridos llamados gorgonina en el orden Gorgonacea –pero su naturaleza química aún continúa siendo
estudiada (Figura 10.3).
👉 Esqueletos escleróticos calcáreos: En la mayoría de los octocorales “imagen siguiente”, las células del mesénquima son denominadas escloroblastos, los cuales segregan escleritas calcáreas de varias formas y colores. Son usualmente estas escleritas las que les dan a los corales de los gorgonianos sus colores y texturas representativos. En muchas especies, las escleritas se hacen bastante densas y pueden fusionarse para formar un marco o red más o menos sólida de tipo calcáreo –rellena de carbonato de calcio cristalizado.

Figura 10.4. Coral
esclerótico
El precioso coral rojo Corallium es un coral gorgoniano con
escleritas coenenquimales fusionadas de color rojo. En los Tubuporos –corales
con forma de pipas de órgano –las escleritas del cuerpo de las parades de los
pólipos individuales se fusionan para formar los tubos rígidos. El esqueleto de
carbonato de calcio de la mayoría de los invertebrados no posee de forma usual
fibras de colágeno, sin embargo, algunos corales gorgonianos –como en
Leptogorgia “imagen siguiente”–las espículas calcáreas incluyen un componente
de colágeno, que ayuda a flexibilizar la estructura. Los esqueletos calcáreos
masivos son producidos principalmente por los antozoos –pólipos formadores de
coral – pero los hidrozoos –las hidras –también presentan algunos linajes
capaces de formar coral, pero aún no se ha encontrado especies en las medusas –Cubozoa,
Staurozoa y Scyphozoa –que sean
capaces de calcificar sus sacos de quitina en el estado juvenil de pólipo.
10.2 Detalles
de los esqueletos coralinos verdaderos
También denominados corales duros verdaderos, son producidos por los antozoos –pólipos formadores de coral –específicamente los corales del orden Sclerectinia “imagen siguiente”. Estos corales son los más comunes y mejor estudiados de todos, en ellos las células de la epidermis en su parte más baja segregan el esqueleto de carbonato de calcio. Inicialmente el esqueleto es cubierto por una membrana viva de epidermis, lo cual técnicamente hace que la primera parte de un coral sea un esqueleto interno. Sin embargo, la parte calcárea sigue creciendo por fuera de la membrana viva construyendo un marco envolvente super-masivo externo, por lo que la mayoría de los autores se refieren al coral como un esqueleto externo. Esto representa un caso en el que las categorías que creamos los humanos no representan adecuadamente los fenómenos biológicos, ya que estamos tratando con un esqueleto que es simultáneamente externo e interno.

Figura 10.5. El Corallum
es la únidad básica de cualquier coral verdadero, compuesto por el pólipo vivo
y el exoesqueleto de carbonato de calcio.
El esqueleto completo de un
esclerectiniano se denomina corallum,
sin importar si se trata de un pólipo solitario o de una colonia; sin embargo,
el esqueleto de un solo pólipo también puede ser designado como coralito. La
parte más externa del coralito se denomina teca; el piso se denomina lámina
basal. Creciendo desde el centro de la lámina basal se encuentra una estructura
de soporte llamada columella. La
lámina basal y las paredes internas de la teca dan lugar de forma radial a
múltiples particiones calcáreas denominadas septos, los cuales se proyectan al
interior y soportan el mesénquima del pólipo. El pólipo ocupa solo la parte
superior de la superficie del corallum.
La dureza del esqueleto de coral crece a medida que crece el pólipo. El corallum puede asumir una gran cantidad
de formas y tamaños, desde estructuras simples semejantes a una copa en los
ejemplares solitarios hasta los grandes superconglomerados de los arrecifes en
las especies coloniales.
10.3 Esqueletos
de las hidras
Los hidrozoos “hidras” poseen formas de vida libre generalmente, pero dos familias –Milleporidae y Stylasteridae –también producen exoesqueletos calcáreos, en este caso el exoesqueleto es comúnmente denominado hidrocoral.

Figura 10.6. Al igual que
los corrales verdaderos, las colonias de los mileporidos pueden asumir una
variada gama de formas, desde estructuras erectas y ramificadas hasta
incrustaciones. El exoesqueleto de los mileporidos se denomina formalmente coenosteum.
El coenosteum
se caracteriza por poros de dos tamaños que acomodan a dos tipos de pólipos.
Los gastrozoides viven en los poros grandes o gastroporos. Los gastroporos se
encuentran rodeados por una serie de poros más pequeños o dactiloporos que dan
resguardo a los dactilozoides. Canales llevan desde los poros hasta el interior
en el coenosteum, estos tubos están
encerrados en túbulos calcáreos. A medida que la colonia envejece, el
hidrocoral se hace más duro, nuevos túbulos se forman, manteniendo los poros de
los pólipos a una profundidad más o menos constante. Las colonias de los
hidrocorales difieren de los corales vertederos debido a que el esqueleto está
penetrado por tejido viviente.
11. Sistema esquelético en platelmintos
|| Regresar al Índice ||
A diferencia de las plantas, las esponjas
y el resto de los seres vivos, el sistema esquelético de los animales desde los
gusanos y los cnidarios deberá responder a una nueva función biológica, el
movimiento. El esqueleto, ya sea duro o hidrostático es el que le confiere su
forma al animal, y la forma es la que le permite al animal movilizarse en un
sustrato –agua, suelo, subsuelo, aire. Un animal sin una forma definida y sin
apéndices o estructuras que le permitan afianzarse a un sustrato no pueden
moverse. Sin embargo, las piezas del esqueleto empleadas para el movimiento
solo permiten afianzar al sustrato, no se mueven por sí mismas, el sistema de
órganos y tejidos encargados de dar movimiento es el sistema muscular. Por tal
razón, a menos que el movimiento de algún animal se encuentre vinculado al
esqueleto, no se mencionara la función de movimiento, la cual será tratada
afondo cuando examinemos el sistema muscular.
Solo unos pocos gusanos planos poseen
algún tipo de elementos esqueléticos especiales. En algunos turbellarios,
pequeñas placas calcáreas o espículas están incrustadas en la pared del cuerpo.
El soporte corporal en todos los demás gusanos planos es proporcionado por las
cualidades hidrostáticas del mesénquima, la elasticidad de la pared corporal y
la musculatura general del cuerpo.
11.1 Tubelarios
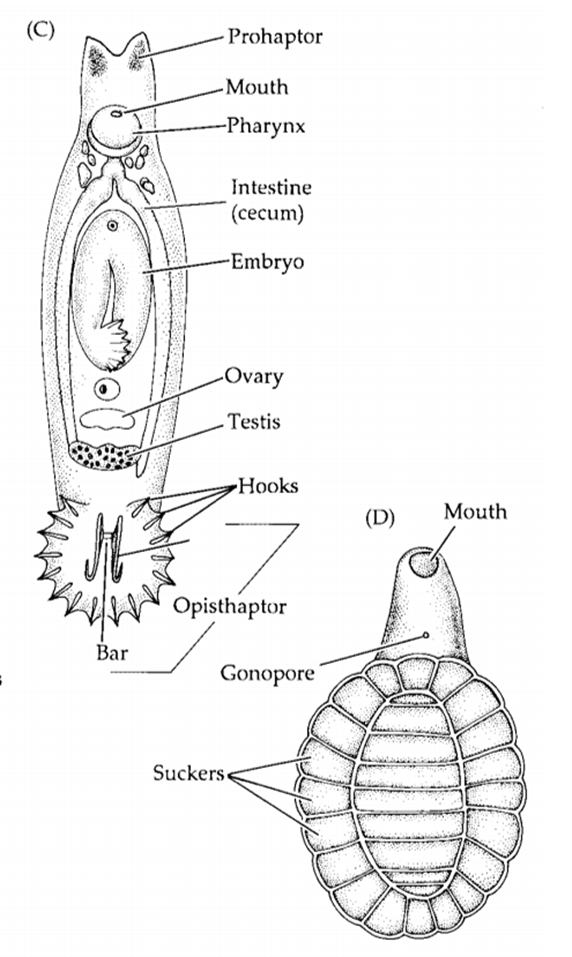
Figura 11.1. Cerca de la
boca y el gonoporo se encuentra una ventosa que permite la fijación a una
posición.
La mayoría de los turbelarios bentónicos
se mueven sobre su superficie ventral mediante un deslizamiento impulsado por
cilios. El moco proporciona lubricación a medida que el animal se mueve y sirve
como un medio viscoso contra el cual actúan los cilios. Algunas de las formas
más grandes o alargadas también usan contracciones musculares. La superficie
ventral del cuerpo se lanza en una serie de surcos transversales alternos y
crestas que se mueven como ondas a lo largo del animal, impulsándolo hacia
adelante. Las ondulaciones musculares de los márgenes laterales del cuerpo
permiten que algunos policíclados grandes naden durante breves períodos de
tiempo. La acción muscular permite que el cuerpo gire y gire, proporcionando
dirección. Algunas formas intersticiales son muy alargadas y utilizan los
músculos de la pared del cuerpo para deslizarse entre los granos de arena.
Muchos de estos tipos de gusanos planos poseen glándulas adhesivas, cuyas
secreciones proporcionan pegajosidad temporal y permiten a los animales
adquirir y aprovechar mientras se mueven. Los turbelarios muy pequeños (p. Ej.,
Acoels) nadan o se deslizan por la acción de los cilios que cubren toda
la superficie del cuerpo.
11.2 Tramátodos
Las duelas adultas carecen de cilios
externos, y su movimiento depende de los músculos de las paredes corporales de
las duelas o de los fluidos corporales de su anfitrión. Algunos se mueven
lentamente sobre o dentro de su huésped por acción muscular, y unos pocos (por
ejemplo, trematodos sanguíneos) se transportan en el sistema circulatorio del
huésped. Sin embargo, ciertas etapas larvales son muy móviles y nadan
utilizando la acción ciliar.
Una vez establecido dentro o en un anfitrión,
es ventajoso que una duela permanezca más o menos en un solo lugar. En ese
sentido, casi todos están equipados con órganos externos para la fijación
temporal o permanente. Los trematodos monogenéticos suelen tener un órgano
adhesivo anterior y posterior llamado prohaptor y opisthaptor, respectivamente.
El prohaptor consiste en un par de estructuras adhesivas, una a cada lado de la
boca, con ventosas o almohadillas adhesivas simples. El opisthaptor suele ser
el principal órgano de unión e incluye uno o más retoños bien desarrollados con
ganchos o garras.
Las duelas digeneticas poseen dos ventosas
sin anzuelo. Uno, el succionador oral, rodea la boca, y el otro, el acetábulo,
se encuentra en la superficie ventral. Estas ventosas generalmente se
suministran con células de glándulas adhesivas, aunque las bien desarrolladas operan
principalmente con succión producida por la acción muscular. Los trematodos
aspidogastreanos carecen de una ventosa oral, pero tienen una ventosa ventral
grande y subdividida.
11.3 Céstodos
Las tenias adultas no se mueven mucho, pero son capaces de ondulaciones musculares del cuerpo. Permanecen fijadas a la pared intestinal del huésped por el escólex (o, en el caso de los miembros de la subclase Cestodaria, por un adhesivo anterior o gancho) y por los microtriches.

Figura 11.2. Escólex de
varios eucestodos. (A) Escólex "típico" con rostelo, anzuelos y
ventosas (Taenia solium). (B) Escólex complejo con ventosas en forma de
hoja (Myzophyllobothrium).
Los detalles de la anatomía escólex Figura
11.2 son extremadamente variables y de importancia crítica en la taxonomía de Eucestoda. La punta del escólex en
muchos cestodos (p. Ej., Taenia) está
equipada con un rostellum móvil con
gancho, que a veces es retráctil en el escólex. En otros (por ejemplo, Cephalobothrium), el extremo anterior
tiene una ventosa protrusible, o almohadilla adhesiva, llamada myzorhynchus. El resto del escólex tiene
varias ventosas o estructuras similares a ventosas y, a veces, ganchos o
espinas. Hay tres categorías de ventosas adhesivas en las que se basa
parcialmente la clasificación ordinal y subordinada de los cestodos.
12. Sistema esquelético nemátodos
|| Regresar al Índice ||
Las técnicas modernas de microscopía
revelan que los órganos de la mayoría de los nematodos ocupan casi todo el
espacio interno. La cutícula proporciona la mayor parte del soporte corporal en
nematodos. En ausencia de músculos circulares de la pared del cuerpo, algunos
tipos de locomoción, como la excavación peristáltica, son imposibles. El patrón
típico de locomoción de nematodos implica contracciones de los músculos
longitudinales, produciendo un movimiento ondulatorio en forma de latigazo.
Entre los nematodos de vida libre, este patrón de movimiento se basa en el
contacto con sustratos ambientales, contra los cuales el cuerpo empuja. Los
músculos actúan contra el esqueleto hidrostático y la cutícula, que sirven como
fuerzas antagónicas a las contracciones musculares. Las fibras colágenas
cruzadas de la cutícula no son elásticas, pero su disposición permite cambios
de forma a medida que el cuerpo se ondula. Cuando se colocan en un ambiente
fluido y privados de contacto con objetos sólidos, los nematodos bentónicos se
agitan de manera bastante ineficiente. Algunos realmente nadan (pero no muy
bien), y algunos pueden arrastrarse usando varias espinas cutáneas, hendiduras,
crestas y glándulas para ganar la compra en el sustrato.
13. Sistema esquelético anélidos
13.1 Poliquetos
Los poliquetos proporcionan un ejemplo
clásico del empleo de espacios celómicos como esqueleto hidrostático para
soporte corporal. Junto con la musculatura bien desarrollada, el plan corporal
metamérico y los parapodios, esta cualidad hidrostática proporciona la base
para comprender la locomoción en estos gusanos. Existen especies de anélidos
que emplean esqueletos calcáreos como guaridas ancladas a un sustrato de forma
permanente, informalmente se los denomina gusanos tubo, aunque son varios los
grupos de anélidos que han desarrollado esta forma de vida de forma
independiente.
13.2 Oligoquetos
Los oligoquetos dependen en gran medida de
su esqueleto hidrostático bien desarrollado para el apoyo y la locomoción. La
acción de los músculos de la pared corporal sobre los fluidos celómicos
proporciona los cambios hidráulicos asociados con el patrón típico de
locomoción de oligoquetos. En ausencia de "paletas" parapodiales, los
oligoquetos dependen del peristaltismo y la manipulación del quetal, para mover
escombros de fondo o para gatear sobre las superficies.
13.3 Hirudineos
El soporte corporal en sanguijuelas se
proporciona por la construcción corporal más o menos sólida, el tejido
conectivo fibroso y las bandas musculares incluidas, y las cualidades
hidrostáticas de los canales celómicos. La ausencia de compartimientos
celómicos aislados, espaciosos y dispuestos segmentariamente impide ciertos
tipos de locomoción que se ven en muchos poliquetos y oligoquetos. Los espacios
celómicos en las sanguijuelas son reducidos y continuos, por lo que estos
animales no pueden moverse como un gusano verdaderamente segmentado. Podemos
ver los músculos circulares y longitudinales como actuando antagónicamente
contra un espacio interno funcionalmente único cuyo volumen permanece constante
(al igual que el volumen de todo el cuerpo).
14. Sistema esquelético moluscos 1, de aplacóforos a bivalvos
|| Regresar al Índice ||
A Los moluscos dependen de dos tipos de
esqueletos, uno es el típico esqueleto hidrostático que está presente en todos
sus linajes, y el segundo es un esqueleto externo que forma una concha
protectora presente en la mayoría de sus linajes. El grupo más grande de
moluscos que no presenta un exoesqueleto es el filum Aplacophora “sin placas” el cual también retiene el plano corporal
de un gusano.
14.1 Caparazón
de los aplacóforos
Los aplacóforos poseen una estructura corporal vermiforme "forma de gusano", pero son formalmente clasificados como moluscos en base a sus estructuras internas.

Figura 14.1. Algunos
Aplacoforos como los solenogastrios pueden segregar placas rudimentarias sobre
sus dorsos o incluso espinas, pero no son comparables a las conchas del resto
del grupo en términos de sofisticación.
El exoesqueleto o concha de los moluscos
es calcáreo y se forma por la acumulación de minerales inorgánicos de calcita o
aragonita, producidas por glándulas especializadas cerca del manto. A pesar de
lo anterior los miembros del filum Aplacophora
tienen la capacidad de segregar espículas calcáreas de forma extracelular.
14.2 Diversidad
de la concha de los moluscos
A diferencia de las rudimentarias placas
de los aplocoforos, las conchas de los moluscos poseen una enorme variabilidad
en tamaño, forma y color, pero todas se adhieren a un mismo plano de
construcción basado en la acumulación de carbonato de calcio producido
extracelularmente, aplicado por capas y en ocasiones cubierto por una membrana
orgánica viva muy delgada denominada periostraco o hipostraco. Lo anterior
convierte a estas conchas técnicamente en un endoesqueleto, pero debido a la
delgadez de dicha membrana nos seguiremos refiriendo a las conchas de los
moluscos como exoesqueletos.
El periostraco está compuesto por proteínas
semejantes a la quitina similares a las que se encuentran en la cutícula de su
epidermis general más blanda. Las capas de calcio son generalmente de dos
tipos, una porción externa muy prismática y otra porción interna laminar y
necrótica. La porción interna se ha perdido evolutivamente de forma secundaria
en muchos grupos “la evolución no siempre es ganancia”. Ambas capas incorporan
conchina –una matriz extracelular sobre la cual los cristales inorgánicos
forman sus núcleos y se expanden –de múltiples formas. Las conchas de los
diferentes tipos de moluscos están compuestas por una cantidad variable de
capas inorgánicas calcáreas.
