
[Mitosis y meiosis] [Microorganismos] [Plantas] [Invertebrados] [Peces] [Anfibios y reptiles] [Aves] [Humanos] [Enfermedades humanas]
Índice
Índice
||1|| Introducción ||2|| Generalidades de la reproducción en vertebrados ||3|| Cilcostomos, lampreas y mixinos ||4|| Peces Cartilaginosos ||5|| Peces óseos ||6|| Gametogénesis y fecundación ||7|| Fertilización | ||9|| Las larvas ||10|| La metamorfosis en los peces ||11|| La determinación ambiental del sexo ||12|| Maduración, longevidad y vejez ||13|| Apareamiento y cambio de sexo ||14|| Caracteres sexuales secundarios y selección sexual ||15|| Desove y cuidado parental |
Portada

Portada
1. Introducción
1. Introducción
Los peces son un clado parafilético que incluye a los agnatostomos o peces sin mandíbulas y a los gnatostomos o peces con mandíbulas, pero excluyendo a sus descendientes tetrápodos terrestres y semiterrestres. Evolutivamente son el grupo de vertebrados mas antiguo que existe y por ende sus caracteristicas han tenido mas tiempo para diversificarse.
Dado lo anterior, en los peces encontramos firtualmente todas las estrategias reproductivas y de desarrollo que encontrremos en los demás vertebrados, incluyendo los comportamientos anidatorios, las metamorfosis con reorganización de sistemas de organos importantes como la alteración de la respiración branquial a la pulmonar en los peces pulmonados, y virtualmente todos los estados intermedios entre la reproducción ovípara, ovovivipara y vivipara.
Por esta amplia variedad de estrategias reproductivas, hemos decidido devidir el capítulo de reproducción en vertebrados y dar un espacio propio a la reproducción de los peces.
El éxito evolutivo está determinado por la capacidad de un individuo para colocar genes en las generaciones futuras, en relación con el éxito de las necesidades específicas inmediatas de sobrevivir el día a día. Para transmitir genes, los individuos deben emparejarse. Sin embargo, muchas especies de peces “y otros vertebrados” son relativamente solitarias como adultos. Durante las temporadas de reproducción, los peces deben superar los hábitos individualistas y buscar posibles parejas de apareamiento. Los hábitats y sustratos de reproducción adecuados deben encontrarse o incluso modificarse en nidos, las actividades de machos y hembras deben estar sincronizadas y deben emplearse tácticas para evitar la hibridación (mecanismos de aislamiento de especies).
Las agregaciones de individuos reproductivamente activos crean una competencia potencial para los sitios de desove y las parejas, provocando un comportamiento territorial y de elección de pareja. El cortejo y el desove pueden distraer la atención de los participantes y hacerlos más vulnerables a los depredadores. Después del desove, muchas especies participan en diversos grados de cuidado parental. Todas estas actividades y características constituyen los diversos sistemas de apareamiento de los peces.
Eugenie Clark (1922-2015)
Eugenie Clark, también conocida como "La Dama de los Tiburones", fue una destacada ictióloga y oceanógrafa estadounidense. Nacida el 4 de mayo de 1922, dedicó su vida al estudio y la defensa de los tiburones. Su pasión y dedicación revolucionaron nuestra comprensión de estos fascinantes depredadores marinos. A lo largo de su carrera, Clark realizó importantes descubrimientos y contribuyó al campo de la biología marina con su valiosa investigación. Su legado continúa inspirando a generaciones de científicos y conservacionistas, dejando una huella imborrable en la ciencia y la protección de los océanos.

Contexto social
Eugenie Clark nació en 1922, una época marcada por importantes eventos históricos como la Segunda Guerra Mundial y la Guerra Fría. Estos conflictos globales generaron tensiones y cambios en el panorama político internacional, que podrían haber influido en su perspectiva y motivación para estudiar la vida marina.
Durante su vida se produjo un crecimiento económico significativo, especialmente en los Estados Unidos. Este desarrollo económico proporcionó oportunidades para la investigación científica y el financiamiento de proyectos relacionados con la biología marina.
Eugenie Clark vivió en una época en la que las mujeres enfrentaban desafíos para ingresar a campos científicos dominados por hombres. Sin embargo, su dedicación y pasión por los tiburones le permitieron superar barreras y destacarse como una destacada científica en un campo tradicionalmente masculino.
Eugenie Clark creció en una sociedad predominantemente cristiana en los Estados Unidos. Si bien su enfoque científico se basó en la observación y el estudio de los animales marinos desde una perspectiva científica, es posible que sus creencias personales y valores culturales hayan influido en su conexión y respeto por la vida marina.
Infancia
La infancia de Eugenie Clark estuvo marcada por su fascinación temprana por el mundo marino. Nacida el 4 de mayo de 1922 en Nueva York, desde muy pequeña mostró un gran interés por los peces y las criaturas acuáticas. A menudo pasaba horas explorando las costas rocosas de Long Island en busca de conchas y observando la vida marina en las mareas bajas.
A pesar de los desafíos que enfrentaba como mujer en ese tiempo, su madre alentó su curiosidad y la animó a seguir sus pasiones. En su infancia, Eugenie Clark disfrutaba de visitar el Acuario de Nueva York, donde se maravillaba con la diversidad y belleza de los peces.
A medida que crecía, su pasión por el mundo submarino se intensificaba. Aprendió a bucear y comenzó a estudiar biología marina en la Universidad de Nueva York. Su infancia y su temprano contacto con el océano sentaron las bases para su carrera futura como ictióloga y defensora de los tiburones.
La infancia de Eugenie Clark fue un período formativo en el que su amor por el mar y su determinación por explorarlo se arraigaron profundamente, allanando el camino para su notable contribución al campo de la biología marina.
Logros
Eugenie Clark, reconocida como "La Dama de los Tiburones", tuvo una destacada educación y numerosos logros científicos a lo largo de su carrera. Tras graduarse en biología marina en la Universidad de Nueva York, obtuvo su doctorado en zoología en la Universidad de Hawái. Durante sus estudios, se especializó en el comportamiento y la fisiología de los peces.
Entre sus principales logros científicos, destacan sus investigaciones pioneras sobre la biología y el comportamiento de los tiburones, particularmente el tiburón toro. Descubrió su capacidad de aprender y recordar, desmintiendo así la creencia generalizada de que los tiburones eran criaturas puramente instintivas.
Eugenie Clark también fue conocida por su trabajo en el desarrollo de equipos y técnicas de buceo, como el uso de tanques de aire comprimido y trajes de neopreno, que permitieron a los científicos explorar las profundidades marinas de manera más segura y eficiente.
En cuanto a sus publicaciones, escribió numerosos artículos científicos y libros, entre ellos "El Misterio del Tiburón" y "La Vida Secreta de los Tiburones". Sus obras contribuyeron a difundir el conocimiento sobre estos animales y a cambiar la percepción negativa que se tenía de ellos.
A lo largo de su carrera, Eugenie Clark también enfrentó algunos errores y controversias. En una ocasión, se retractó de una afirmación inicial sobre la docilidad de los tiburones toro, reconociendo que habían sido casos aislados y que en general eran animales salvajes.
A pesar de los desafíos y las controversias, el legado científico de Eugenie Clark es innegable. Sus investigaciones, descubrimientos y contribuciones en el campo de la ictiología marina han dejado una huella duradera en la comprensión y conservación de los tiburones y otros peces marinos. Su dedicación y pasión por el océano la convierten en una figura destacada en el campo de la biología marina.
Comunidad científica
Eugenie Clark fue una figura influyente en la comunidad científica, especialmente en el campo de la ictiología marina. Durante su carrera, estableció numerosas colaboraciones con otros científicos, construyendo una red de colegas y amigos que compartían su pasión por la investigación marina.
Uno de sus principales colaboradores fue el renombrado ictiólogo japonés Hisashi Imamura, con quien realizó investigaciones conjuntas sobre tiburones y peces tropicales en la década de 1950. Juntos, publicaron varios artículos científicos que ampliaron el conocimiento sobre la biología y el comportamiento de estas especies.
Asimismo, Clark fue una defensora de la colaboración entre científicos y buceadores recreativos, lo que le permitió obtener valiosos datos y observaciones sobre los tiburones en su entorno natural. Esta colaboración incluyó a buzos experimentados como Hans Hass y Jacques Cousteau, quienes compartieron sus experiencias y conocimientos con Clark.
Si bien Clark fue ampliamente respetada y admirada en la comunidad científica, también enfrentó opositores y críticos en algunos aspectos de su trabajo. Algunos científicos cuestionaron inicialmente sus afirmaciones sobre la inteligencia y el comportamiento de los tiburones, considerándolas exageradas o insuficientemente respaldadas por evidencia empírica.
A pesar de las diferencias de opinión y la existencia de opositores, no se puede decir que Clark tuviera enemigos en el sentido estricto de la palabra. Siempre se mostró abierta al diálogo y dispuesta a debatir sus ideas con aquellos que cuestionaban sus hallazgos.
En general, la comunidad científica vio a Eugenie Clark como una investigadora apasionada y dedicada, cuyos logros y contribuciones en el campo de la ictiología marina fueron altamente valorados. Su trabajo inspiró a generaciones de científicos y su legado perdura en la investigación y conservación de los ecosistemas marinos.
Reconocimientos
Eugenie Clark recibió numerosos reconocimientos a lo largo de su vida y póstumos por su destacada contribución a la ictiología y la divulgación científica. Estos reconocimientos incluyeron premios, medallas, honores y la apreciación de la comunidad científica y el público en general.
En vida, Clark recibió la Medalla de Plata de la Sociedad Zoológica de Londres en 1955, en reconocimiento a sus investigaciones sobre tiburones. También fue galardonada con la Medalla de Oro de la Sociedad Geográfica Nacional en 1988 por sus contribuciones a la comprensión de los océanos y su biodiversidad.
En 1991, Clark fue nombrada "Embajadora del Océano" por la National Oceanic and Atmospheric Administration (NOAA) de Estados Unidos, en reconocimiento a su trabajo de divulgación y educación sobre los océanos.
Tras su fallecimiento en 2015, diversos homenajes y reconocimientos fueron realizados en su honor. La Universidad de Maryland estableció el "Día de Eugenie Clark" en 2016 para celebrar su legado y contribuciones a la ciencia. También se le otorgó póstumamente el Premio a la Trayectoria en Conservación de la revista Smithsonian.
En cuanto a los apodos, Clark era conocida como "La Dama de los Tiburones" debido a su dedicación y experiencia en el estudio de estas criaturas. Su pasión y valentía al investigar en el agua junto a los tiburones también le valió el apodo de "Buceadora de Tiburones".
En resumen, Eugenie Clark fue ampliamente reconocida y honrada en vida y después de su fallecimiento por sus contribuciones científicas y su defensa de los océanos. Sus premios, medallas y títulos reflejan el impacto significativo que tuvo en la comunidad científica y el aprecio que el público en general tenía por su trabajo y dedicación.
¿Por qué es importante?
Eugenie Clark y su investigación en la ictiología tienen un impacto significativo en nuestra vida cotidiana, aunque a menudo no nos demos cuenta directamente. Su trabajo en el estudio de los peces y los ecosistemas acuáticos ha contribuido a nuestro conocimiento sobre la biodiversidad marina y los ecosistemas acuáticos, lo cual tiene implicaciones importantes en diferentes aspectos de nuestra vida.
En primer lugar, su investigación nos ha proporcionado una comprensión más profunda de los océanos y su importancia para el equilibrio del planeta. Esto nos ayuda a tomar conciencia de la necesidad de conservar y proteger los ecosistemas marinos y a tomar medidas para preservarlos.
Además, su estudio de los tiburones y otros peces ha permitido mejorar la seguridad en las actividades acuáticas. Su investigación sobre el comportamiento y las interacciones de estas criaturas nos ha ayudado a comprender mejor cómo evitar encuentros peligrosos y cómo protegernos de posibles ataques.
El trabajo de Eugenie Clark también ha tenido impacto en la industria pesquera y en la gestión de los recursos acuáticos. Sus investigaciones sobre la biología y la ecología de los peces han proporcionado información clave para la conservación de especies, la pesca sostenible y la gestión adecuada de los ecosistemas acuáticos.
Además, su dedicación a la divulgación científica y la educación ha inspirado a muchas personas a interesarse por la vida marina y a cuidar del medio ambiente. Sus esfuerzos en este sentido han contribuido a fomentar una mayor conciencia ambiental y a promover prácticas más sostenibles en nuestra vida diaria.
¿Qué debemos aprender de ella?
Eugenie Clark nos brinda una poderosa inspiración a través de su vida, sus limitaciones, obstáculos, logros e impacto en el campo de la ictiología. Su historia nos enseña valiosas lecciones sobre perseverancia, determinación y pasión por la ciencia y la conservación del medio ambiente.
A pesar de los desafíos y prejuicios que enfrentó como mujer en un campo dominado por hombres, Eugenie Clark no dejó que eso la detuviera. Su dedicación y curiosidad inquebrantables la llevaron a superar barreras y convertirse en una pionera en su campo.
Su trabajo nos muestra la importancia de seguir nuestra pasión y perseguir nuestros sueños, incluso cuando enfrentamos obstáculos aparentemente insuperables. Nos enseña que, a pesar de las dificultades, podemos lograr grandes cosas si nos comprometemos y trabajamos arduamente en lo que amamos.
El impacto de la vida de Eugenie Clark también nos invita a reflexionar sobre nuestra propia responsabilidad con el medio ambiente. Su dedicación a la conservación marina nos recuerda que cada uno de nosotros puede hacer la diferencia en la protección de los océanos y la vida acuática. Nos anima a tomar medidas concretas para preservar los ecosistemas acuáticos, reducir nuestra huella ecológica y promover prácticas sostenibles en nuestras comunidades.
En última instancia, la vida de Eugenie Clark nos inspira a ser valientes, perseverantes y comprometidos con nuestros ideales. Nos muestra que no hay límites para lo que podemos lograr si nos apasionamos por una causa y nos esforzamos por alcanzarla. Su legado nos impulsa a seguir su ejemplo y marcar una diferencia en el mundo, ya sea en el campo científico, la conservación ambiental o cualquier otra área en la que estemos apasionados.
2. Generalidades de la reproducción en vertebrados
2. Generalidades de la reproducción en vertebrados
Los vertebrados son por lo general animales dioicos, pero en muchas especies de peces se da el cambio de sexo, dependiendo de la disponibilidad de individuos de un sexo determinado en una colonia. Como característica, la historia evolutiva del aparato reproductor de los vertebrados siempre se encuentra asociado al sistema excretor, es decir al desarrollo de los nefridios. Funcionalmente los dos sistemas son bastante diferentes, el sistema excretor principal está compuesto por los riñones metanefrídicos y los conductos que transportan los desechos nitrogenados.
El sistema reproductor incluye las gónadas y los conductos que transportan los huevos, la esperma y en ocasiones llevan a cabo otras funciones reproductivas. El estudio del funcionamiento del sistema reproductor es quizá más profundo en los vertebrados que en los invertebrados, lo cual se debe a una serie de similitudes funcionales en todos los vertebrados, donde el ser humano es quizá la especie mejor estudiada. Las gónadas no solo producen gametos, también son órganos endocrinos, mientras que los conductos reproductivos pueden ser reclutados para otros desarrollos embrionarios posteriores, ya sea manteniendo el huevo en el interior o desarrollando a un embrión sin cáscara de manera más directa.
Las hormonas generadas por las gónadas se encargan, al igual que en el ser humano, del desarrollo de los caracteres sexuales secundarios, necesarios para el cortejo, la fecundación y el embarazo en aquellas especies con algún grado de desarrollo interno. Las hormonas también poseen un efecto en los rasgos de comportamiento, como el cuidado de las crías y la alimentación de las crías con diversos niveles de organización.
En la presente unidad no solo estudiaremos los órganos del sistema reproductor, sino también de algunos aspectos relacionados al desarrollo de los seres vivos bajo la premisa de visualizar el ciclo de vida de los seres vivos. Hay que destacar que es muy común pensar que los ciclos de vida de los vertebrados son sencillos, pero ese no es el caso, existen diversos grados de desarrollo indirecto en los cuales existe una diferencia entre una etapa larvaria y una etapa adulta.
Fertilidad
En la mayoría de los vertebrados, la reproducción es estacional. El cortejo y la cópula se encuentra usualmente restringida a un breve momento anual que se sincroniza de modo tal que las crías nazcan durante las estaciones más favorables como las estaciones lluviosas o en la primavera. Durante la temporada de apareamiento, los conductos genitales reciben los gametos, lo cual también reactiva los comportamientos reproductivos, momento denominado celo. Los humanos “Homo sapiens” somos una excepción notable al mantener el celo durante todo el año.
La fertilidad se define como la habilidad de la hembra para producir huevos fertilizables o en el macho para producir espermatozoides de suficiente calidad y cantidad para lograr la fertilización de dichos huevos u óvulos. Un macho que produce espermatozoides en cantidades insuficientes es denominado infértil o estéril.
Fertilización interna o externa
La fertilización y el desarrollo de los vertebrados poseen diversos grados de desarrollo. Muchas especies de peces desovan en el agua y realizan una fecundación externa, pero otros poseen fecundación interna con diferentes grados a su vez de desarrollos embrionarios internos. La fertilización interna generalmente necesita algún nivel de desarrollo de órganos intromitentes que denominamos penes o claspers.
La fertilización externa tiene como principal ventaja que no involucra mayores esfuerzos fisiológicos por parte de los padres para el desarrollo de las crías, más allá de posiblemente ofrecer algunos cuidados a los huevos. Sin embargo, también involucra algunos inconvenientes. Las ranas Ascaphus spp., por ejemplo, viven en ambientes de corrientes fuertes que pueden llevarse a los gametos impidiendo su fecundación y su ubicación en un lugar adecuado. Otra inconveniente es que ni la hembra ni el macho poseen algún control de selección sexual total, por ejemplo, si un macho ha realizado todo el ritual de apareamiento y está listo para liberar su esperma, bien puede aparecer un macho de calidad inferior y liberar el esperma propio al mismo tiempo sobre los mismos huevos, lo cual crea una ascendencia mixta que no era de interés de la hembra.

Figura 2.1. Las ranas con cola del género Ascaphus son anfibios únicos que pertenecen a la familia Ascaphidae. Estas especies, conocidas comúnmente como ranas de vientre de fuego, se caracterizan por tener una cola larga y un cuerpo alargado. A diferencia de la mayoría de las ranas, los machos de Ascaphus tienen un órgano reproductor especializado llamado "cladode" que les permite realizar la fecundación interna. Estas ranas se encuentran en regiones montañosas del noroeste de América del Norte, especialmente en arroyos y ríos de agua fría y rápida. Su dieta se compone principalmente de insectos acuáticos.Las ranas con cola son fascinantes debido a sus características únicas y su adaptación a entornos acuáticos. Su estudio proporciona información valiosa sobre la diversidad y las adaptaciones de los anfibios en diferentes ecosistemas.
La fertilización interna ofrece que el cortejo y la fertilización sea eventos separados a la deposición de los huevos, adicionando el hecho de que tanto machos como hembras pueden tener grados diversos de selección sexual real. Los rituales de cortejo sirven de mecanismos de reconocimiento de especie. La fertilización puede no ocurrir en un ambiente adecuado para la deposición de los huevos. Por ejemplo, algunas salamandras se aparean en la tierra donde el cortejo puede verse fácilmente, pero la tierra seca es inviable para los huevos no amnióticos de las salamandras.
Restricciones fisiológicas pueden llegar a limitar la evolución del viviparismo en algunos grupos. Al interior de los amniotas, el calcio para la osificación del esqueleto embrionario puede ser almacenado en el vitelio o en el cascarón. En el viviparimo, el huevo calcáreo duro se pierde, permitiendo un intercambio eficiente entre los tejidos maternos y del embrión en desarrollo, pero con la pérdida del cascarón como reservorio de calcio para el desarrollo. La evolución paralela del viviparismo es común en aquellos linajes que no emplean el cascarón como reservorio de ion calcio(2+).
3. Cilcostomos, lampreas y mixinos
3. Cilcostomos, lampreas y mixinos
El sistema reproductor de los vertebrados se encuentra íntimamente vinculado a su aparato excretor, y ambos han coevolucionado, pues estructuras abandonadas en la función excretora fueron tomadas por el reproductor.
En los ciclóstomos, un único ovario se encuentra suspendido en la pared mesodorsal. Las lampreas son el grupo de ciclostomos mejor conocidos, mientras que de los mixinos aún se sabe relativamente poco a cerca de su comportamiento sexual. A diferencia de los mamíferos como el ser humano, las lampreas liberan una enorme cantidad de ovocitos en la única cohorte de su único ciclo sexual, en este sentido, cerca de 200.000 folículos pueden desarrollarse de forma sincrónica. La mayoría de las lampreas mueren después del desove. Los ciclóstomos no poseen trompas de Falopio, por lo que los óvulos son liberados en el celoma, desde donde alcanzan la cloaca o el ano a través de poros secundarios.

Figura 3.1. El sistema reproductor de los ciclóstomos (vista ventral) es básicamente el mismo para ambos sexos, una única gónada (ovario/testículo) ubicada en el interior del animal que segrega gametos (óvulos/espermatozoides) al celoma (cavidad general del cuerpo), los gametos son capturados por el embudo celómico que se conecta a los tubos excretores donde salen gracias al nefridioporo junto con la orina hacia la cloaca.
En los machos de los ciclóstomos también existe un único testículo que no está conectado a ningún ducto genital. Al igual de los óvulos, los espermatozoides son liberados al celoma desde donde los espermatozoides alcanzan la cloaca o el ovario a través de poros especializados. El poro especializado se denomina embudo celómico se encuentra vinculado al poro urinario como indica la figura. En los ciclóstomos no existe vínculo anatómico/funcional entre el sistema excretor y el sistema reproductor.
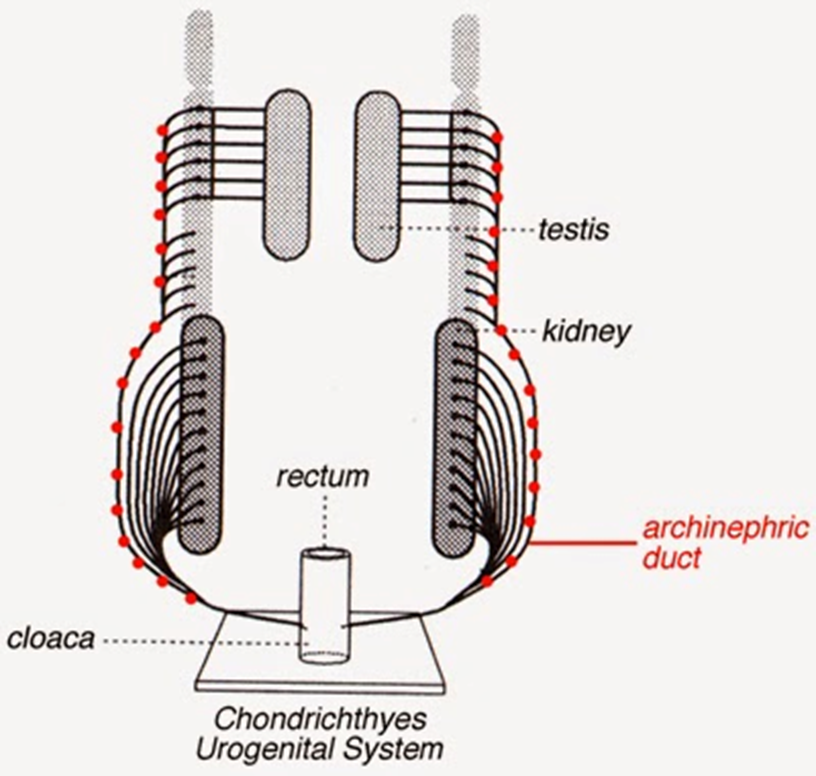
Figura 3.2. Sistema reproductor de tiburones y rayas. En los tiburones uno de los túbulos del mesonefros crece (ducto arquinéfrico "en rojo") para empotrarse con las gónadas que ya están configuradas en parejas.
Uno de los aspectos más notables de los procesos evolutivos es que las estructuras que se degeneran no se pierden del todo y gracias a un atavismo afortunado una estructura puede recuperar su desarrollo para nuevas funciones. Como vimos en el sistema excretor de los mixinos, el riñón de estos peces pasa por una etapa juvenil en la que los glomérulos de la punta o pronefros se desarrollan, y mientras el animal crece, los glomérulos del medio crecen para formar el mesonefros. El mesonefros no solo cumple la función de proyectar glomérulos excretores, evolutivamente hablando también será empleado para proyectar tubos más largos que se empotran en los testículos formando los túbulos seminíferos. El tubo del mesonefros que cumple la función de túbulo seminífero se denomina túbulo arquinéfrico, pero en los embriones en desarrollo se lo llama tubo de Leidig, tubo de Wolff o tubo nefrico. En las hembras el sistema excretor no se desarrolla a través del sistema excretor, sino por un tubo formado de novo paralelo al riñón.
4. Peces Cartilaginosos
4. Peces Cartilaginosos
Examinaremos en detalle las características anatómicas y funcionales del sistema reproductor de estos fascinantes animales acuáticos. Los elasmobranquios, que incluyen a los tiburones y las rayas, son una de las clases más antiguas y diversas de peces cartilaginosos. Su sistema reproductor presenta una serie de adaptaciones únicas que les permiten reproducirse de manera eficiente en su entorno acuático. En este artículo, se analizan aspectos como la estructura de los órganos reproductores, el proceso de fertilización, la gestación y el desarrollo de las crías. Además, se exploran las diferencias entre las especies de elasmobranquios y cómo estas adaptaciones anatómicas han contribuido a su éxito evolutivo. El conocimiento detallado de la anatomía reproductiva en estos animales es fundamental para comprender su biología, su reproducción y su conservación.
Sistema reproductor de las hembras de tiburones y rayas
Los elasmobranquios son el principal grupo de peces condrictios “esqueleto de cartílago” y está compuesto por los grupos más famosos, los tiburones y las rayas. En estos peces puede verse un aspecto interesante en la evolución del sistema reproductor y es una evolución no homóloga de los ductos reproductivos, o lo que es lo mismo, los conductos reproductivos de machos y hembras poseen un origen diferente.

Figura 4.1. Arriba; Sistema reproductor izquierdo de un tiburón, en este caso solo está presente la parte alta del útero por lo que esta especie tiene en efecto dos úteros separados. El ovario se encuentra separado del cifón que funciona como trompa de Falopio recolectando los óvulos liberados. Puede observarse que el conducto reproductor o paramesonefros es independiente del ducto urinario, lo cual convierte a los conductos reproductores masculino y femenino en análogos, pero no homólogos;

Figura 4.2. Sistema reproductor extraído de un tiburón real, en este caso si existe un útero bajo "único" y un útero alto dividido en una parte derecha y otra izquierda.
En las hembras un ducto paralelo al del mesonefros llamado por tal razón, conducto paramesonéfrico, en los humanos este ducto se denomina ducto de Muller y cumple en general las mismas funciones. El ducto paramesonéfrico crece hasta proyectarse cerca de los ovarios, su función es la de capturar los óvulos liberados al celoma y canalizarlos hacia la cloaca “Igual que en las trompas de Falopio”. El ducto mesonéfrico puede ser empleado para fecundación interna mediante coito, por lo que su sección intermedia puede almacenar semen y en las especies con reproducción ovípara, permite la formación de la cascara alrededor del embrión. Esta sección intermedia es denominada la glándula de la cascara. La parte baja se denomina útero, y en los tiburones este está compuestos por tres secciones, la parte alta es la base del tubo paramesonéfrico mientras que la parte baja es la función de los dos tubos, en las especies vivíparas y ovovivíparas es donde se desarrollan los embriones y fetos. En las hembras los arcos del mesonefros drenan únicamente al riñón funcional “opistonefros” que en su mayoría proviene del mesonefros.

Figura 4.3. Sistema reproductor interno y externo de un tiburón macho, los testiculos se encuentran en el interior del animal los cuales se conectan al arquinefros que posee función de epididimo por el arquinefros, el cual discurre de forma interconectada hasta que sale de forma definitiva y se dirige a la base del ducto urinario donde se fusiona, los espermatozoides salen por el uréter hacia la cloaca; abajo; Sistema reproductor extraído de un tiburón real.


Figura 4.4. Sistema reproductor extraído de un tiburón real.
Sistema reproductor de los machos de tiburones y rayas
El sistema de tubos seminales en los machos es absolutamente dependiente de los tubos del mesonefros, un grupo de estos que parte del arquinefros crece sin formar glomérulos para empotrarse en los testículos. Por el contrario, los tubos de Muller no se desarrollan de forma prominente. El túbulo del arquinefros no solo se empotra en los testículos, también se conecta con la estructura arcaica del pronefros que ahora cumple una función semejante a la del epidídimo y el vas deferens, es decir, almacenar los espermatzoides, madurarlos y durante la eyaculación contraerse para generar la presión de expulsión. Los espermatozoides viajan por el arquinefros hasta uréter en la cloaca, y de allí son capturados por el clasper.
El pene de los tiburones y rayas
El pene puede entenderse en sentido laxo como cualquier órgano copulatorio masculino destinado para una fertilización interna, en este sentido los ductos seminales deben conectarse desde la cloaca o ser continuos.

Figura 4.5. Los claspers son en últimas un apéndice óseo en cuya punta hay dígitos que se abren, los (radials) son las estructuras que forman la aleta pelviana como tal", aunque su función es análoga a la del pene, en realidad son homólogos a las piernas ya que emergen al igual que ellas de las aletas pelvianas, es por esto que existen reportes que dicen que los primeros penes evolucionaron como "piernas extra".
En los tiburones estas estructuras copulatorias se llaman clasper y cómo puede entenderse de cualquier órgano en términos evolutivos, estos se forman a partir de un órgano preexistente, en este caso las aletas pélvicas cercanas a la cloaca. Debido a que las aletas pelvianas son dos, de igual forma es el número de claspers, lo cual hace que el tiburón tenga dos órganos copilatorios, es decir, dos penes.

Figura 4.6. Otros peces con cláspers. Se supone que lo único lo suficientemente sólido como para fosilizar en un placodermo era la coraza del cráneo o placa dérmica dado que el resto del esqueleto era de cartílago como en los tiburones y rayas (Long et al., 2015), pero en condiciones especiales, se pueden obtener esqueletos completos, que incluyen los clásper.
Durante la copula, sin embargo, el tiburón solo puede usar uno de sus claspers de forma simultánea nadando al lado de la hembra como se muestra en el siguiente video (YouTube). Los claspers pueden ser encontrados en todos los elasmobranquios por lo que las rayas también lo poseen, resulta interesante que debido a que los claspers son modificación de una aleta, poseen hueso y son capaces de fosilizarse en caso de que tratemos con linajes de peces óseos. Un ejemplo son los placodermos “aunque no son en sentido estricto peces óseos, por lo menos si lo eran sus penes”, algunas especies han dejado sus penes fosilizados. Durante la cópula, uno de los claspers es introducido en la cloaca de la hembra y su cartílago final se abre como si se tratara de una sombrilla, lo cual ancla al macho en la hembra “algo útil cuando no hay de que sostenerse”. Cuando el esperma es emitido sale de los ductos seminíferos a la cloaca, donde ingresa por el embudo del clasper junto con agua.
Canibalismo embrionario
El canibalismo intrauterino implica que uno o más embriones fuertes compiten por recursos en el interior, y esto conlleva a la eliminación de la competencia convirtiéndola en alimento. Este canibalismo puede clasificarse en la forma fuerte y la forma débil. La forma fuerte del canibalismo intrauterino en los tiburones se denomina embriofagia o adelfofagia, la cual fue descubierta en 1948. En este caso el embrión más grande se come a sus hermanos “literalmente a mordiscos”. La forma débil del canibalismo intrauterino se llama ovifagia en la cual los embriones fertilizados emplean los óvulos no fertilizados como fuente de nutrientes dado que los embriones de los tiburones no se insertan en el útero. Este tipo de canibalismo es conocido desde 1907 (Awruch, 2015). Algunas especies reportadas con este comportamiento son:
(a) Carcharodon carcharias: También conocido como el gran tiburón blanco, es una especie ampliamente reconocida y estudiada. Se han registrado casos ocasionales de canibalismo embrionario en esta especie, donde los embriones más grandes pueden consumir a sus hermanos más pequeños dentro del útero de la madre.

Figura 4.7. El Carcharodon carcharias, conocido comúnmente como gran tiburón blanco, es una especie icónica y temida en los océanos. Se caracteriza por su imponente tamaño, poderosa mandíbula y afilados dientes serrados. Su cuerpo hidrodinámico y musculoso le permite ser un depredador ágil y eficiente. Con una longitud que puede superar los 6 metros, es considerado uno de los mayores depredadores marinos. Su dorso es de color grisáceo o marrón, mientras que el vientre es blanco, lo que le confiere un camuflaje efectivo al acechar a sus presas. Posee una excelente capacidad olfativa y visual, lo que le permite localizar y perseguir a sus presas, principalmente focas, peces y otros tiburones. Aunque su reputación está marcada por su presencia en películas y relatos de ataques a humanos, la interacción con ellos es muy rara y se cree que no consideran a los humanos como presas. La conservación del gran tiburón blanco es de vital importancia debido a su papel crucial en los ecosistemas marinos y su estatus de especie amenazada debido a la caza indiscriminada y la degradación de su hábitat.
(b) Lamna ditropis: Conocido como el tiburón mako de aleta larga, también ha sido documentado con casos de canibalismo embrionario. Los embriones más grandes pueden devorar a sus hermanos más pequeños en el útero, lo que proporciona una ventaja competitiva para su supervivencia.
(c) Sphyrna tiburo: Este es el tiburón martillo pequeño, y también se ha observado canibalismo embrionario en esta especie. Algunos estudios indican que el canibalismo intrauterino podría ocurrir cuando los recursos alimentarios son limitados, lo que brinda una mayor oportunidad de supervivencia para los embriones más grandes.
(d) Galeus melastomus: Conocido como el tiburón de aguas profundas, es una especie que habita en aguas frías y profundas. Ha sido reportado con mayor frecuencia de canibalismo embrionario, donde los embriones más grandes se alimentan de los más pequeños en el útero materno.
Nacimiento
Existen tres diferentes formas por las cuales los tiburones se desarrollan fetalmente y nacen:
Viviparismo
Los huevos eclosionan en el interior del útero de la hembra, pero se desarrolla una placenta que interacciona entre la hembra y las crías alimentándoles y liberándolas de sus desechos metabólicos. Los tiburones vivíparos generalmente dan a luz a más de una cría.
(a) Rhincodon typus: También conocido como tiburón ballena, es el pez más grande del mundo. A pesar de su enorme tamaño, es una especie completamente inofensiva para los humanos y se alimenta principalmente de plancton. Los tiburones ballena son vivíparos, lo que significa que dan a luz crías vivas en lugar de poner huevos. Las hembras llevan a las crías en su útero hasta que están completamente desarrolladas.

Figura 4.8. Rhincodon typus, conocido como tiburón ballena, es el pez más grande del mundo. Puede alcanzar una longitud de hasta 12 metros y un peso de más de 20 toneladas. Su cuerpo es grisáceo con manchas distintivas en forma de lunares o rayas. Este tiburón se alimenta de plancton y pequeños organismos marinos, filtrándolos a través de su enorme boca. A pesar de su tamaño, es amigable y pacífico, no representa peligro para los humanos. Se reproduce de manera vivípara, dando a luz crías vivas. El tiburón ballena es admirado por científicos, buceadores y amantes de la vida marina, recordándonos la importancia de conservar los océanos para proteger su diversidad y supervivencia.
(b) Carcharhinus leucas: Conocido como tiburón toro, es una especie agresiva y territorial que se encuentra en aguas tropicales y subtropicales. Son vivíparos y las hembras dan a luz a crías vivas después de un período de gestación de aproximadamente 10 meses. Las crías nacen en aguas poco profundas y son capaces de valerse por sí mismas desde el momento del nacimiento.
(c) Galeocerdo cuvier: El tiburón tigre es conocido por su distintivo patrón de rayas similares a las de un tigre. Son vivíparos y las hembras dan a luz a crías vivas después de un período de gestación de aproximadamente 14 meses. Las crías nacen en aguas costeras y son capaces de nadar y cazar por sí mismas desde el principio.

Figura 4.9. Galeocerdo cuvier, conocido como tiburón tigre, es una especie de tiburón que habita en aguas cálidas y tropicales de todo el mundo. Su cuerpo es robusto y alargado, con una coloración distintiva de rayas verticales que se asemejan a las marcas de un tigre, de ahí su nombre. Puede alcanzar tamaños de hasta 5 metros de longitud. Es un depredador ágil y poderoso, con dientes afilados y una mandíbula fuerte. Se alimenta de una variedad de presas, incluyendo peces, tortugas marinas y mamíferos marinos. El tiburón tigre es conocido por ser oportunista y adaptable en su dieta. Aunque es temido por su potencial peligrosidad, los encuentros con humanos son relativamente raros. Su presencia en los ecosistemas marinos juega un papel importante en el equilibrio y la salud de los océanos.
Oviparismo
El oviparismo en los tiburones se refiere al proceso de reproducción en el cual las hembras depositan huevos externamente, fuera de su cuerpo. A diferencia de los tiburones vivíparos, que dan a luz a crías vivas, los tiburones ovíparos producen huevos que deben ser incubados fuera del cuerpo de la madre.
Los huevos de tiburón ovíparos se conocen como "cápsulas de huevo" o "mermeladas de huevo". Estas cápsulas tienen una forma rectangular o en forma de bolsa, con dos lados planos y dos lados curvos. Generalmente son de color oscuro, lo que les permite mezclarse con el entorno y proporcionar camuflaje.
El tiempo de incubación de los huevos de tiburón ovíparos varía según la especie y las condiciones ambientales. Puede oscilar desde unas pocas semanas hasta varios meses, dependiendo de la temperatura del agua y otros factores ambientales. Durante este período, el embrión se desarrolla y crece dentro de la cápsula de huevo.
En la mayoría de las especies de tiburones ovíparos, no hay cuidado parental después de la deposición de los huevos. La hembra abandona los huevos una vez que los ha colocado en un lugar seguro, como el fondo del océano o en estructuras submarinas. Sin embargo, algunos estudios sugieren que ciertas especies de tiburones ovíparos pueden exhibir comportamientos defensivos para proteger los huevos de posibles depredadores o perturbaciones.
(a) Scyliorhinidae: Esta familia de tiburones, comúnmente conocidos como tiburones gato, son ovíparos. La hembra deposita varios huevos en un vivero y puede cuidar de ellos durante la incubación. Algunas especies de este grupo incluyen el tiburón gato común (Scyliorhinus canicula) y el tiburón gato de coral (Apristurus spp.).
(b) Orectolobiformes: Este orden de tiburones incluye varias especies ovíparas, como el tiburón alfombra (Chiloscyllium spp.) y el tiburón de bambú (Hemiscyllium spp.). Estos tiburones ponen huevos que se adhieren a objetos en el lecho marino y se incuban allí hasta que eclosionan las crías.
(c) Hemiscylliidae: Esta familia de tiburones está estrechamente relacionada con los tiburones de bambú y también son ovíparos. El tiburón nodriza de cola corta (Hemiscyllium ocellatum) es un ejemplo de especie ovípara dentro de esta familia.
(d) Familia Heterodontidae: Esta familia de tiburones, conocidos como tiburones nodriza, también incluye especies ovíparas. El tiburón nodriza común (Heterodontus francisci) es un ejemplo de especie ovípara dentro de esta familia. Las hembras de tiburón nodriza ponen huevos que se adhieren a sustratos marinos donde se incuban hasta la eclosión.

Figura 4.10. Heterodontus francisci, el tiburón nodriza común, es una especie ovípara que habita en el Pacífico oriental. Con un cuerpo robusto y una protuberancia nasal puntiaguda, este tiburón alcanza una longitud de aproximadamente 1.2 metros. Su coloración varía entre marrón y gris para mimetizarse en su entorno. Posee dientes afilados en la parte frontal de la mandíbula para morder y triturar presas como crustáceos, moluscos y peces pequeños. Durante el día, descansa en el fondo marino y muestra comportamientos defensivos cuando se siente amenazado debido a su naturaleza territorial. Las hembras de esta especie ponen huevos que se adhieren a objetos marinos para su incubación hasta que las crías eclosionan. El tiburón nodriza común es un fascinante ejemplar con adaptaciones especializadas para su supervivencia y reproducción en su hábitat marino.
Ovoviviparismo
No existe placenta “y representa una etapa evolutiva a las formas con placenta”, en este caso los huevos eclosionan en el putero, pero las crías deben valerse por sí solas en un ambiente con espacio y nutrientes extremadamente limitados, alimentándose de los óvulos no fertilizados y de sus propios hermanos.
La cápsula ovárica no es continua con el oviducto “un tema general en muchos vertebrados”, por lo que los huevos se vierten en la cavidad del cuerpo, la condición de gimnovaria. Los huevos entran en el embudo del oviducto, que es un conducto de Müller, no un conducto mesonéfrico modificado; Se desarrolla como una continuación posterior de la túnica ovárica. La parte anterior del oviducto está especializada para formar una glándula nidamental o de la concha donde tiene lugar la fertilización. La glándula nidamental secreta una membrana alrededor del óvulo fertilizado.
En los taxones ovíparos (puesta de huevos), la membrana es córnea, compuesta de queratina. La glándula nidamental puede funcionar como un recipiente seminal donde los espermatozoides se nutren antes de la fertilización. En las especies vivíparas (vivas), la parte posterior del oviducto se modifica para formar un útero, que alberga el embrión en desarrollo.
(a) Carcharias taurus (Tiburón toro): Este tiburón es conocido por su reproducción ovovivípara. Los huevos se desarrollan y eclosionan dentro del cuerpo de la hembra, y las crías nacen vivas. La gestación dura aproximadamente 12 meses.

Figura 4.11. Carcharias taurus, conocido como tiburón toro, es un tiburón ovovivíparo de gran tamaño que se encuentra en aguas tropicales y templadas de todo el mundo. Con una cabeza ancha y un cuerpo robusto, puede alcanzar hasta 3.5 metros de longitud. Su coloración varía de grisácea a marrón oscuro, con manchas más claras en el dorso y una piel rugosa. La hembra del tiburón toro da a luz crías vivas después de un período de gestación de aproximadamente 12 meses. Durante este tiempo, los huevos se desarrollan y eclosionan internamente en su cuerpo. Posee una mandíbula fuerte y dientes triangulares y afilados, adaptados para alimentarse de presas principalmente carnívoras. Este tiburón se considera un depredador tope en su ecosistema y puede ser encontrado en hábitats costeros y marinos. Es conocido por su comportamiento territorial y su naturaleza robusta. El tiburón toro juega un papel importante en el equilibrio de los ecosistemas marinos, manteniendo el control poblacional de otras especies.
(b) Triakis semifasciata (Tiburón leopardo): El tiburón leopardo también es ovovivíparo. Los huevos se incuban internamente y las crías nacen vivas. La gestación puede durar de 10 a 12 meses.
(c) Mustelus canis (Tiburón cazón): Esta especie de tiburón también sigue un ciclo reproductivo ovovivíparo. Los huevos se incuban dentro del cuerpo de la hembra y las crías nacen vivas. La gestación dura alrededor de 9 meses.
(d) Galeocerdo cuvier (Tiburón tigre): Aunque la mayoría de los tiburones tigre son vivíparos, algunas poblaciones pueden ser ovovivíparas. Los huevos se incuban dentro del cuerpo de la hembra, pero en lugar de una placenta desarrollada, las crías reciben nutrición de un saco vitelino. Las crías nacen vivas después de un período de gestación de aproximadamente 14 meses.
5. Peces óseos
5. Peces óseos
Los peces teleósteos, una de las subdivisiones más numerosas y diversas de los peces óseos, cuentan con una increíble variedad de estrategias reproductivas y sistemas reproductores especializados. Estos peces exhiben una amplia gama de características anatómicas adaptadas a sus diferentes modos de reproducción y desarrollo.
En este artículo, exploraremos la anatomía del sistema reproductor en los peces teleósteos, centrándonos en cinco aspectos clave. Analizaremos la estructura y función de los órganos reproductores, como los ovarios y los testículos, y cómo se relacionan con la producción de gametos.
Además, examinaremos los mecanismos de fecundación, desde la liberación y fertilización de los huevos hasta el desarrollo embrionario. También exploraremos las diferencias entre los peces teleósteos ovíparos, ovovivíparos y vivíparos, y cómo se lleva a cabo la incubación y el cuidado parental en cada caso.
A lo largo del artículo, destacaremos las adaptaciones evolutivas y las peculiaridades anatómicas de diferentes especies de peces teleósteos, resaltando la diversidad y la complejidad de los sistemas reproductivos en este grupo. A través de este estudio, obtendremos una visión más profunda de la anatomía y la fisiología reproductiva de los peces teleósteos y su importancia en la conservación y el entendimiento de la biodiversidad acuática.
Sistema reproductor femenino
En los peces óseos hembras, como la mayoría de las hembras anamnotas, los conductos arquinéfricos sirven a los riñones y los oviductos pareados (conductos de Müller) sirven a los ovarios pareados (Figura 5.1 y Figura 5.2).

Figura 5.1. Sistemas reproductores de varios peces óseos femeninos 1. Conducto de Müller aparece adyacente y paralelo al conducto arquinéfrico en la mayoría de los peces. En los teleósteos, el oviducto generalmente se reemplaza por un conducto ovárico que se deriva por separado.

Figura 5.2. Sistemas reproductores de varios peces óseos femeninos 2. (a) pez torpedo. (b) Protopterus. (c) pez arquero

Figura 5.3. Sistemas reproductores de varios peces óseos masculinos.
En algunos teleósteos, como los salmónidos, los huevos liberados de los ovarios llenan la cavidad del cuerpo. Eventualmente, alcanzan los remanentes cortos y en forma de embudo de los oviductos situados en la parte posterior del celoma. Sin embargo, en muchos teleósteos, los oviductos retroceden por completo, dejando el transporte de óvulos a nuevos conductos ováricos. Estos conductos ováricos no son homólogos a los oviductos (conductos de Müller) de otros vertebrados. En su lugar, se derivan de pliegues peritoneales que abrazan cada ovario y van crecido posteriormente para formar nuevos conductos.
La mayoría de los peces teleósteos ponen huevos, pero algunos dan a luz crías vivas. Entre estos teleósteos vivíparos, los tejidos maternos pueden nutrir el embrión:
(a) Poecilia reticulata (Guppy): El guppy es una especie de pez pequeño y colorido originario de América del Sur. Son famosos por su reproducción vivípara, en la que las crías se desarrollan y nacen vivas. Las hembras pueden retener espermatozoides para múltiples fertilizaciones y dar a luz a crías completamente formadas.
(b) Gambusia affinis (Mosquito fish): Este pez, también conocido como pez mosquito, es nativo de América del Norte. Es vivíparo y presenta un desarrollo interno de las crías. Las hembras pueden almacenar esperma y dar a luz a crías vivas, que son funcionales y capaces de nadar poco después del nacimiento.
(c) Xiphophorus helleri (Espada): El pez espada es originario de América Central y presenta una reproducción vivípara. Las hembras dan a luz a crías vivas después de un período de gestación interno. Los alevines nacen completamente formados y pueden cuidarse a sí mismos desde el momento del nacimiento.
(d) Poeciliopsis gracilis (Molly): El molly es un pez tropical nativo de América del Norte. Es vivíparo y las hembras dan a luz a crías vivas. Al igual que otras especies vivíparas, las hembras pueden almacenar esperma y dar a luz a crías totalmente desarrolladas. Un caso extremo se encuentra en la familia de teleósteos que incluye el pez millón “Poecilia reticulata”. En este grupo, la fertilización ocurre mientras los óvulos todavía están en los folículos ováricos. El ovario continúa sujetando los embriones durante el desarrollo posterior hasta que se liberan como pequeños alevines. Los ovocitos que no alcanzan un punto de maduración en el que pueden ser fertilizados generalmente se someten a involución y se llaman ovocitos atréticos. El reciclaje del tejido atrético proporciona nutrición para los ovocitos sobrevivientes.

Figura 5.4. Poecilia reticulata, conocido como guppy, es un pequeño pez tropical originario de América del Sur. Destaca por su belleza y facilidad de reproducción en acuarios. Los machos son más pequeños y tienen colores vivos, mientras que las hembras son más grandes y de tonos más apagados. Son peces ovovivíparos, las hembras retienen los huevos hasta que las crías están completamente formadas. Los guppies son activos y se adaptan fácilmente a diferentes condiciones de agua. Su reproducción rápida y prolífica los convierte en una especie popular en la cría en cautiverio. Son ideales para principiantes en la acuariofilia debido a su resistencia y comportamiento pacífico. Además de su atractivo estético, los guppies desempeñan un papel importante en la investigación científica debido a su capacidad de mostrar cambios rápidos en respuesta a diferentes condiciones ambientales.
Sistema reproductor masculino
En los peces óseos, la anatomía reproductiva presenta una serie de características especiales y notoriamente diferentes. Por ejemplo, en cuanto a la presencia de penes o hemipenes, es importante mencionar que en la mayoría de los peces óseos, no se desarrolla un órgano copulador similar al pene presente en otros grupos de animales.
En cambio, los peces óseos tienen una estructura llamada conducto arquinéfrico, que cumple diversas funciones, entre ellas el drenaje de los riñones y la posible recepción de esperma proveniente de los testículos. Sin embargo, los testículos de los peces óseos tienden a desarrollar conductos espermáticos separados y rutas de salida.
Un ejemplo de esta diferenciación en los conductos espermáticos se observa en la figura 9b y 9c. Allí se puede apreciar cómo los testículos desarrollan conductos espermáticos separados, distintos al conducto arquinéfrico. Esta separación permite que los espermatozoides tengan una ruta específica hacia el exterior.
En algunos teleósteos, un subgrupo de peces óseos, este sistema de conductos separados puede incluso formar un conducto testicular que no es homólogo al conducto arquinéfrico. Esta estructura puede tener su propia apertura hacia el exterior, como se muestra en la figura 9d. Esta particularidad demuestra la diversidad en la anatomía reproductiva de los peces óseos y la capacidad de desarrollar sistemas especializados para la reproducción.
Además, existen casos de teleósteos, como los salmónidos, que presentan una situación aún más única, ya que carecen por completo de conductos espermáticos. En lugar de eso, los espermatozoides se liberan directamente en la cavidad del cuerpo o celoma. Posteriormente, estos espermatozoides salen del cuerpo a través de poros ubicados cerca de la parte posterior del celoma.
Esta forma de liberación de espermatozoides en la cavidad del cuerpo es una adaptación reproductiva especializada en los salmónidos y demuestra otra variación significativa en la anatomía reproductiva de los peces óseos.

Figura 5.5. El clado de los salmónidos es un grupo de peces que pertenecen a la familia Salmonidae. Comprende una amplia diversidad de especies, incluyendo el salmón, la trucha y la trucha de mar. Estos peces son conocidos por su migración desde el océano hacia los ríos para reproducirse, en un fenómeno llamado anadromía. Los salmónidos poseen características distintivas, como aletas adiposas, que son pequeñas aletas ubicadas entre la aleta dorsal y la aleta caudal. Además, presentan un cuerpo fusiforme y escamas ctenoides. Son peces carnívoros y se alimentan principalmente de invertebrados acuáticos y peces más pequeños. Su importancia económica y ecológica es significativa, ya que son objeto de pesca comercial y recreativa, además de ser indicadores de la salud de los ecosistemas acuáticos. Los salmónidos son un clado fascinante y diverso dentro de la familia de los peces, con adaptaciones especiales para su estilo de vida y una notable capacidad de migración.
En resumen, los peces óseos presentan una diversidad notable en su anatomía reproductiva, con diferentes tipos de conductos espermáticos y rutas de salida. La presencia de penes o hemipenes es poco común en estos animales, y en su lugar, se han desarrollado estructuras especializadas, como el conducto arquinéfrico y los conductos espermáticos separados. Los salmónidos destacan aún más al liberar los espermatozoides directamente en la cavidad del cuerpo. Estas adaptaciones reproductivas únicas demuestran la increíble variedad y evolución en el mundo de los peces óseos.
6. Gametogénesis y fecundación
6. Gametogénesis y fecundación
Este capítulo se enfocará en los aspectos de los gametos, la fecundación y la embriogénesis de los peces hasta que el juvenil rompe el cascarón.
Gametogénesis en los peces

Figura 6.1. Gametogénesis en los peces. La gametogénesis masculina y femenina es muy conservada en todos los vertebrados, y sus detalles los describiremos en el sistema reproductor mamífero típico, el del ser humano. En la imagen tenemos a la izquierda la espermatogénesis y a la izquierda la ovogénesis generalizada de los peces óseos.

Figura 6.2. Amia calva, conocida comúnmente como "pez remo" o "amia", es una especie de pez de agua dulce que pertenece al orden Amiiformes. Este pez es considerado una especie fósil viviente, ya que se cree que ha existido durante más de 150 millones de años, lo que lo convierte en uno de los peces más antiguos que aún habita en nuestros días. La Amia calva tiene un aspecto único, con un cuerpo alargado y comprimido, y una aleta dorsal situada en la mitad posterior del cuerpo. Posee una mandíbula grande y una boca con dientes afilados. Es un depredador que se alimenta principalmente de otros peces, invertebrados acuáticos y pequeños vertebrados. Este pez es conocido por su capacidad de respirar aire atmosférico a través de un órgano llamado vejiga natatoria modificada, lo que le permite sobrevivir en aguas con baja concentración de oxígeno. La Amia calva es una especie interesante desde el punto de vista evolutivo y ecológico, siendo un verdadero tesoro viviente que nos conecta con el pasado distante de la vida en la Tierra.
La mayoría de los peces tienen gónadas pareadas, aunque un miembro de la pareja puede ser consistentemente más grande que el otro en algunas especies o solo una gónada puede ser funcional. Los mixinos y las lampreas son únicos en que solo se desarrolla un ovario, a partir de la fusión de dos primordios en las lampreas y de la pérdida de un ovario en los mixinos. A diferencia de los tiburones y otros vertebrados, los testículos y los ovarios en los peces sin mandíbulas y los peces óseos se desarrollan solo desde la corteza del epitelio peritoneal, no desde la corteza y la médula.
Los testiculos
Los testículos en machos inmaduros suelen ser de color rojizo y adquieren una textura suave y una coloración cremosa a medida que el pez madura y se aproxima el tiempo de desove. Los testículos generalmente representan <5% del peso corporal. Los folículos dentro de los testículos producen espermatozoides en desarrollo (= espermatogénesis) a través de una serie de transformaciones meióticas y de desarrollo típicas de los vertebrados: espermatogonias, espermatocitos primarios, espermatocitos secundarios, espermátides y espermatozoides.
Los espermatozoides de los peces varían en tamaño, forma, número de flagelos (ninguno a dos) y presencia o ausencia de acrosomas y otras estructuras. Los espermatozoides de los peces son indicadores de clasificación, incluso al nivel de especie. Las cabezas de los espermatozoides varían en longitud desde aproximadamente 2 mm (Amia calva, Lota lota, Oryzias latipes) hasta 70 mm (Neoceratodus forsteri). El acrosoma con forma de casco en el extremo anterior de la mayoría de los espermatozoides de peces primitivos se pierde en prácticamente todos los peces neopterigios (Lepisosteiformes, Amia calva y teleósteos). Dos familias africanas de peces osteoglosomorfos, los Mormyridae (peces elefante) y Gymnarchidae, carecen de flagelo. Su esperma puede moverse por algún tipo de movimiento ameboide.

Figura 6.3. El pez pulmonado de Queensland (Neoceratodus forsteri) es una especie de pez de la familia Ceratodontidae endémica de Australia, y considerada como fósil viviente ya que se han hallado restos fósiles de este grupo de hace 380 millones de años en la región australiana de Nueva Gales del Sur. También se le conoce como pez pulmonado australiano, barramunda y salmón de Burnett.
Los eyaculados típicos durante el desove contienen millones de espermatozoides. Los espermatozoides se liberan en el fluido seminal en especies con fertilización externa, o en paquetes llamados espermatóforos en fertilizantes internos. En general, se afirma que los machos producen un exceso de esperma y, por lo tanto, el éxito reproductivo de los machos está más limitado por el acceso a las hembras que por la capacidad de producir gametos (lo contrario se considera limitante en las hembras).

Figura 6.4. La lota (Lota lota), también llamada bacalao de agua dulce, es un pez adaptado a aguas frías del hemisferio norte. Posee un cuerpo alargado, cabeza grande y boca ancha con dientes afilados. Su coloración varía entre tonos pardos oscuros. Estos peces habitan en lagos, ríos y embalses, y son depredadores voraces que se alimentan de otros peces, insectos acuáticos y crustáceos. Destacan por su capacidad de sobrevivir en ambientes con bajo contenido de oxígeno y pueden respirar aire atmosférico. Durante el invierno, buscan refugio en el fondo del agua y entran en letargo. La lota es una especie fascinante de los ecosistemas acuáticos.
Sin embargo, en circunstancias en las que los machos se aparean diariamente durante una temporada de reproducción prolongada, puede producirse el agotamiento de los espermatozoides y, de hecho, el apareamiento puede retrasarse hasta que se llenen las reservas de esperma.
Ovogénesis
El desarrollo de los huevos, ocurre dentro del ovario y también progresa a través de varias etapas que involucran oogonios, ovocitos primarios y secundarios, y finalmente huevos. Los oogonios se desarrollan a partir de células sexuales primordiales en el epitelio germinal de la pared del ovario. Los gránulos proteicos de la yema se depositan alrededor de los ovocitos primarios durante la vitelogénesis, fabricándose los precursores del material de la yema en el hígado. Las gotitas de aceite se incorporan en la yema. Los huevos maduros pasan del ovario a través del oviducto, que es una continuación del tejido ovárico, hacia el exterior a través de la cloaca. En los elasmobranquios “tiburones y rayas”, no existe una conexión directa entre el ovario y el oviducto y, por lo tanto, los huevos pasan a través de la cavidad peritoneal en su camino hacia el oviducto.
En varios peces óseos, el oviducto está muy reducido o ausente y los huevos entran en la cavidad celómica peritoneal del cuerpo antes de ser arrojados. Las hembras que han engendrado se denominan gastadas; sus ovarios son sangrientos y contienen huevos residuales que son reabsorbidos por el ovario. La reabsorción de huevos es un proceso general. Por lo general, la mayoría de los huevos maduros en un ovario se engendran, mientras que una pequeña proporción se reabsorbe y las proteínas, grasas y minerales contenidos en ellos son reutilizados por la hembra para el mantenimiento, el crecimiento o la producción de más huevos. El número reabsorbido varía enormemente entre y dentro de las especies, dependiendo del tamaño de los peces, la cantidad de desoves previos en esa temporada y el estado energético de la hembra.
Fecundidad y estrategias de vida r/K
La cantidad de huevos liberados por una hembra durante un desove o un ciclo de reproducción, varía de uno a dos en algunos tiburones a decenas de millones en Megalops atlanticus y Molva molva, a 3.00×106 en el pez gigante, Mola mola; También se pueden calcular las fecundidades estacionales y de por vida. La mayoría de los peces marinos templados más grandes producen de decenas de miles a millones de huevos a la vez. La fecundidad generalmente disminuye al aumentar el tamaño del huevo y al aumentar el cuidado de los padres, pero aumenta con el tamaño corporal en un individuo.
Los criadores de boca, como el pez gato de mar y algunos cíclidos, producen solo unos 100 huevos a la vez, y los vivíparos, como el pez de cuatro ojos Anableps spp., contienen alrededor de una docena de embriones. La relación entre el número de huevos y el tamaño corporal suele ser proporcional a la masa de la hembra, lo que refleja el volumen del cuerpo de una hembra que puede transportar los huevos.

Figura 6.5. El sábalo real o tarpón (Megalops atlanticus), es un pez osteíctio elopiforme de la familia de los megalópidos. Es un pez de gran tamaño, de entre 2 y 2,5 m y alrededor de 150 a 161 kg de peso, de cuerpo fusiforme parecido a un arenque, totalmente cubierto de grandes y gruesas escamas, verde azulado en la parte superior y plateado en sus costados. La aleta dorsal empieza en la mitad del dorso y en el último radio, al igual que sucede con la aleta anal, se alarga hasta formar un filamento. Las aletas pectorales se encuentran en posición baja y las pelvianas son abdominales.
Por lo tanto, el número de huevos generalmente aumenta en relación con el cubo, la cuarta o quinta potencia de la longitud de la hembra. Además de producir más huevos, las hembras más grandes de muchas especies producen huevos más grandes y mejores que dan como resultado una mayor supervivencia larvaria: por ejemplo, en salmones, bacalao, eglefino, lubina rayada, platija.

Figura 6.6. El pez luna (Mola mola) es un pez pelágico tetraodontiforme de la familia Molidae. Es el pez óseo más pesado del mundo, con una media de 1000 kg de peso y con ejemplares que alcanzan más de 3 m de longitud y superan las 2 toneladas. Es una especie cosmopolita que habita en aguas tropicales y templadas a lo largo de todo el planeta. Tiene el cuerpo aplastado lateralmente y cuando extiende sus aletas dorsales y ventrales, el pez es tan largo como alto.
Las excepciones muestran la prima otorgada a garantizar la supervivencia de los jóvenes en lugar de producir grandes cantidades de huevos. En los cíclidos que incuban en la boca, la fecundidad aumenta en relación con el cuadrado de la longitud de la hembra debido a que el tamaño de la boca aumenta solo linealmente al aumentar la longitud del cuerpo. Debido a la reabsorción, las estimaciones de fecundidad basadas en el recuento de huevos maduros no necesariamente indican una fertilidad verdadera, que es el número de descendientes viables producidos. Las estimaciones de fecundidad para los peces vivíparos se complican aún más por el consumo de huevos durante el desarrollo de embriones.
Así podemos designar que la estrategia reproductiva de los peces es r o reproductiva, variando desde extrema con millones de crías, hasta mediana con docenas de crías, solo algunos tiburones tendrían una estrategia K con dos crías. Al interior de los reproductores tenemos a su vez que distinguir según la etapa de crecimien to de la hembra, pues las hembras más gordas dan más y mejores huevos, combinando así partes de la estrategia de vida K.
Los huevos
Ciertas generalizaciones se aplican a los huevos de los peces, con una fuerte correlación entre el hábitat y las características de los huevos fertilizados. La mayoría de los peces marinos, independientemente de las afinidades sistemáticas, los hábitos demersales o pelágicos, la distribución costera u oceánica, los rangos tropicales o boreales, desovan huevos pelágicos que se fertilizan externamente y flotan individualmente cerca de la superficie del mar.

Figura 6.7. Vinciguerria lucetia es un pez que pertenece a la familia Phosichthyidae. Esta especie es bioluminiscente, con dos hileras de pequeños fotóforos a lo largo de su cuerpo. Habita en la zona tropical del Indo Pacífico, donde hace migraciones todos los días.

Figura 6.8. El celacanto es un raro género de peces que incluye dos especies existentes: el celacanto del Océano Índico Occidental (Latimeria chalumnae) y el celacanto de Indonesia (Latimeria menadoensis). Siguen el linaje vivo más antiguo conocido de Sarcopterygii (peces con aletas en el lóbulo y tetrápodos), lo que significa que están más estrechamente relacionados con los peces pulmonares, reptiles y mamíferos que con los peces con aletas comunes.
Entre los reproductores pelágicos, los huevos son generalmente de forma esférica y tienen un único glóbulo de aceite. Sus diámetros varían desde aproximadamente 0,5 mm (Vinciguerria spp., Photichthyida hasta 5,5 mm (morenas Muraenidae), con un tamaño modal de aproximadamente 1 mm. Esta notable convergencia entre taxones filogenéticamente extendidos sugiere un conjunto común y adaptable de soluciones a las presiones de selección encontradas por los huevos que se dispersan pasivamente en un entorno cercano a la superficie y que contienen un embrión dependiente de los suministros de yema para la nutrición.
La mayoría de los peces de agua dulce y algunas especies marinas costeras divergen de este patrón y producen huevos demersales que se depositan en el fondo. Muchos engendran en nidos y participan en algún tipo de cuidado parental. Los huevos demersales tienden a ser relativamente grandes, de hasta 7 u 8 mm como en el salmón. Los huevos teleósteos más grandes son producidos por bagres marinos que crecen en la boca y varían de 14 a 26 mm. Los huevos de tiburón son generalmente más grandes que los huevos de los peces óseos, mientras que los huevos de peces óseos más grandes son producidos por el celacanto Latimeria calumnae, con un diámetro de 9 cm. Los huevos demersales a menudo tienen coriones gruesos y recubrimientos especiales que pueden brindar protección mecánica.

Figura 6.9. Blennodesmus scapularis. La anguila ocelada es una especie de pez con aletas radiadas de la subfamilia Congrogadidae, que pertenece a la familia Pseudochromidae. Es la única especie en el género monoespecífico Blennodesmus. Se encuentra a lo largo de las costas del norte de Australia, desde Australia Occidental, a lo largo de la costa del Territorio del Norte hasta Queensland.
Las desviaciones de la forma esférica se encuentran en los huevos alargados de algunas anguilas, anchoas, pececillos, peces loro y gobios. Los huevos de la anguila Blennodesmus scapularis tienen forma de cruz, y los huevos de algunos peces dardo tienen una sangría profunda y parecen casi en forma de corazón. Aunque generalmente es lisa, la membrana vitelina externa del huevo, llamada corion, puede ser esculpida (como en el pez lagarto, pez hacha de aguas profundas, salmonetes, algunos plateados), o puede tener filamentos, tallos o espinas (peces linternas mictofiformes y muchos peces aterciopelados, como el mariquita, pez volador). Los filamentos a menudo ayudan a que los huevos se adhieran a otros huevos o a estructuras como las algas marinas, como en los peces voladores.
La variación en otras estructuras de huevos puede ayudar en la identificación de especies o familias. El grado de segmentación y pigmentación de la yema difiere, ya que los teleósteos primitivos, como muchas anguilas, arenques y salmones, tienen yemas segmentadas, mientras que los teleósteos más avanzados tienen yemas homogéneas. Las yemas pigmentadas producen huevos coloridos en, bagres y salmones. La aparición, el número y la ubicación de los glóbulos de aceite en la yema difieren entre las especies. Los glóbulos de aceite pueden servir como nutrición para los embriones, como mecanismos de flotación y, cuando están pigmentados con melanina, pueden ayudar a proteger las estructuras sensibles en desarrollo de la radiación dañina. Los glóbulos de aceite en algunas especies pasan por patrones de movimiento altamente predecibles. Los huevos también varían entre las especies en términos del espacio entre el corion y la yema, denominado espacio perivitelino o espacio "alrededor de la yema".

Figura 6.10. Diagrama de un huevo de pez óseo. A. membrana vitelina B. corion C. yema de huevo D. glóbulo de aceite E. espacio perivitelino F. embrión. Los peces producen muchos huevos, generalmente de aproximadamente 1 mm de ancho, y generalmente los liberan en la columna de agua abierta.
Un aspecto interesante de la reproducción es el esfuerzo y la energía que gastan las diferentes especies e individuos, que a menudo se correlaciona con el patrón de historia de vida que una especie ha evolucionado. El esfuerzo reproductivo incluye la ingesta de alimentos y su transferencia a las gónadas, así como el gasto de energía en el crecimiento somático frente al gonadal. En las hembras, la maduración de los ovocitos implica la movilización de los primeros lípidos y luego las proteínas de otras partes del cuerpo, como los depósitos de grasa y los músculos del cuerpo, hacia el ovario. Esta maduración también se acompaña de un aumento de 10 veces en el consumo de oxígeno por el ovario hasta la etapa final de la ovogénesis, cuando la masa ovárica aumenta a través de la acumulación de agua. Los verdaderos costos de reproducción también deben tener en cuenta la energía gastada durante las migraciones reproductivas, el cortejo, el desove y la crianza interna y otras formas de cuidado de los padres, entre otros factores. La energía gastada en el crecimiento no mononadal puede ser sustancial. Por ejemplo, el esturión y el salmón, hembras migratorias utilizan el 80–90% de sus reservas de grasa corporal durante la reproducción, gran parte de la cual se gasta durante la migración a las zonas de desove.
7. Fertilización
7. Fertilización
La fecundación es un proceso fundamental en la reproducción de los seres vivos, y los peces, como grupo diverso y ampliamente distribuido en los ecosistemas acuáticos, presentan una amplia variedad de estrategias reproductivas. Desde las técnicas de fertilización externa hasta los sistemas de fertilización interna, la fecundación en peces ofrece un fascinante panorama de adaptaciones y comportamientos únicos.
En este artículo, exploraremos los diferentes métodos de fecundación utilizados por los peces, destacando las características clave de cada uno. Analizaremos tanto la fecundación externa, donde los huevos y el esperma se liberan en el agua, como la fecundación interna, donde los gametos se encuentran dentro del cuerpo de la hembra. Además, examinaremos casos especiales de peces hermafroditas y de fertilización múltiple.
Descubriremos cómo los peces han desarrollado estrategias sorprendentes para garantizar la supervivencia de sus crías, desde la producción de grandes cantidades de huevos hasta la formación de estructuras protectoras para proteger los embriones en diferentes etapas de desarrollo.
En última instancia, este artículo nos brindará una visión fascinante sobre la fecundación en peces, resaltando la adaptabilidad y la diversidad de estos animales acuáticos en sus esfuerzos por perpetuar sus especies en los variados hábitats acuáticos del mundo.
Fecundadores externos
En la mayoría de los peces, la fertilización ocurre fuera del cuerpo de la hembra (YouTube) (YouTube). En cualquier caso, la fertilización ocurre cuando un espermatozoide penetra o se le permite ingresar a la membrana del óvulo a través de un orificio en forma de embudo en la membrana llamado micrópilo. El micrópilo se encuentra sobre el polo animal del huevo y es demasiado estrecho para permitir el paso de más de un espermatozoide. Después de la entrada de esperma, el micrópilo se cierra y el corion tiende a endurecerse, lo que evita la polispermia o la entrada de más de un esperma. Los huevos de esturión tienen varios micropilas y se produce polispermia. La presencia y el tamaño de los micrópilos son diagnósticos de diferentes especies. Se forma un cigoto cuando se fusionan los pronúcleos del esperma y el óvulo.
En especies con fertilización externa, los gametos (huevos y esperma) tienen una viabilidad limitada en el agua. Su capacidad de fertilización varía dependiendo de la temperatura y puede oscilar entre menos de un minuto hasta una hora, siendo la viabilidad más prolongada en aguas más frías. Estudios sobre eventos naturales de desove han revelado que al menos el 75% y con frecuencia entre el 90% y el 95% de los huevos liberados son fertilizados. Esta proporción está directamente relacionada con la cantidad de esperma que el macho libera.
En especies que se reproducen en la columna de agua, como los lustrosos y los peces loro, los machos pueden regular el gasto de esperma en función del tamaño y la competencia de las hembras. Cuando desovan hembras más grandes que liberan una mayor cantidad de huevos, o cuando otros machos intentan desovar simultáneamente, los machos aumentan la cantidad de esperma liberado. Por otro lado, en especies que se reproducen en el fondo y son territoriales, como los gobios, el gasto de esperma no parece aumentar en respuesta a la liberación de más huevos por parte de las hembras o a la competencia de otros machos.

Figura 7.1. El clado de los peces loro, científicamente conocidos como la familia Labridae, agrupa a una diversa variedad de especies marinas. Estos peces se caracterizan por su apariencia colorida y sus bocas protuberantes en forma de pico, lo que les otorga su nombre común. Se encuentran en aguas tropicales y subtropicales alrededor del mundo, especialmente en los arrecifes de coral. Con más de 90 géneros y alrededor de 500 especies, los peces loro son uno de los grupos más numerosos de peces marinos. Destacan por su comportamiento territorial y agresivo durante la reproducción. Su papel ecológico es importante, ya que se alimentan de algas y otros invertebrados marinos, contribuyendo al equilibrio y la salud de los arrecifes de coral. Su capacidad de adaptación a diversos hábitats acuáticos los convierte en un fascinante grupo de estudio para científicos y amantes de la vida marina.
Un ejemplo interesante de fertilización externa se encuentra en los peces anfibios, como la rana de lluvia. Estas especies depositan sus huevos en charcos temporales de agua, y cuando llueve, los machos liberan su esperma directamente sobre los huevos. La fertilización ocurre externamente y los embriones se desarrollan en el agua hasta que eclosionan y se convierten en alevines.
Fecundadores internos
Las hembras de algunas especies fertilizadas internamente pueden almacenar esperma en el ovario (YouTube). En la perca enana, (Embiotocidae), los machos recién nacidos son sexualmente maduros y aunque eyaculan no fertilizan a las hembras recién nacidas inmediatamente. Las hembras almacenan este esperma durante 6 a 9 meses hasta que maduran y ovulan. El almacenamiento de esperma está muy extendido en los miembros del clado Poeciliidae que son viviparos, que a menudo involucran a más de un compañero masculino. Algunas especies almacenan esperma y lo utilizan para fertilizar varios lotes de huevos. Las hembras de Heterandria formosa, almacenan esperma de una cópula durante 10 meses y lo utilizan para fertilizar hasta nueve crías de embriones en desarrollo diferentes, varias de las cuales pueden desarrollarse simultáneamente, un fenómeno conocido como superfetación.

Figura 7.2. Heterandria formosa, comúnmente conocido como molly pigmeo o molly enano, es una especie de pez de agua dulce perteneciente a la familia Poeciliidae. Se encuentra principalmente en las regiones del sureste de los Estados Unidos y México. Estos pequeños peces alcanzan un tamaño de alrededor de 2.5 centímetros y tienen una apariencia distintiva con un cuerpo alargado y aletas transparentes. Heterandria formosa es conocido por su comportamiento social y pacífico, lo que los convierte en una opción popular para los acuarios comunitarios. Son peces vivíparos, lo que significa que las hembras dan a luz a crías vivas en lugar de poner huevos. Su dieta consiste principalmente en pequeños invertebrados acuáticos y algas. Son muy prolíficos y pueden reproducirse fácilmente en cautiverio. Además, son resistentes y pueden adaptarse a una variedad de condiciones de agua.
En algunas especies, la activación de la división celular no es sinónimo de fertilización. Algunos miembros del clado Poeciliidae que son viviparos son ginogenéticos en el sentido de que las hembras usan esperma de machos de otras especies para activar la división celular, pero en realidad no se incorpora material genético masculino al cigoto. En algunas especies fertilizadas internamente, la fertilización se produce, pero el desarrollo puede detenerse después de unas pocas divisiones celulares y luego se reanuda cuando las condiciones ambientales son más favorables para la eclosión. En algunos peces anuales, como las rivulinas sudamericanas y africanas, los huevos se fertilizan y luego se entierran; pasan la estación seca en un estado de reposo conocido como diapausa.
La fertilización interna es universal entre los tiburones, pero se limita a una docena de familias de peces óseos, especialmente los celacantos; un siluro pez gato; brotulidos; portadores de la vida, mexclapiques, tres géneros de peces mediopicos; peces de cuatro ojos; los neostethidos y phallostethidos de Asia sudoriental; scorpaenidos en los géneros Sebastodes spp., y Sebastes spp., (por ejemplo, Sebastes viviparus); Pez aceite de Baikal; embiotocido mojarras vivíparas; la anguila viruela, Zoarces viviparus; clinidos y labrisomidas. Un ciprínido asiático, Puntius viviparus, se describió originalmente como un vivíparo, pero un examen posterior del tipo de material indicó que la depredación de cíclidos jóvenes, que estaban contenidas en el estómago, se había confundido con el desarrollo de crías en el ovario.

Figura 7.3. Puntius viviparus, conocido como barbo vivíparo o barbo de Burma, es una especie de pez de agua dulce perteneciente a la familia Cyprinidae. Es originario de los ríos y arroyos de Myanmar y Tailandia. Este pez presenta una apariencia llamativa, con un cuerpo alargado y esbelto, y una coloración que varía entre tonos dorados y plateados. Una característica destacada de Puntius viviparus es su capacidad de reproducción vivípara, lo que significa que las hembras dan a luz a crías vivas en lugar de poner huevos. Esto los convierte en peces fascinantes para observar en acuarios. En su entorno natural, se alimentan de pequeños invertebrados acuáticos y material vegetal. En cautiverio, aceptan una dieta variada de alimentos comerciales y alimentos vivos o congelados.
Penes
La reproducción es un proceso vital para la supervivencia de las especies, y los peces no son una excepción. Una característica notable en muchos peces es la presencia de órganos reproductivos masculinos llamados penes. Estos penes han evolucionado de manera sorprendente en diferentes especies, mostrando una asombrosa diversidad y adaptaciones reproductivas. En este artículo, exploraremos la clasificación de los penes en los peces y proporcionaremos ejemplos concretos de especies que exhiben diferentes tipos de órganos reproductores.
Los penes en los peces se clasifican en varios tipos, según su estructura y función. A continuación, se presentan tres categorías principales:
(a) Penes externos: Algunas especies de peces poseen penes externos que son visibles a simple vista. Un ejemplo destacado es el gonopodio en los peces vivíparos del género Poecilia, como el guppy (Poecilia reticulata). Estos penes están adaptados para transferir el esperma directamente a la hembra durante la reproducción.
(b) Penes intromitentes: Muchos peces tienen penes intromitentes, que son órganos reproductores que se introducen en la hembra durante la cópula. El pez gato del Amazonas (Trachelyopterus galeatus) es un ejemplo de una especie con un pene intromitente en forma de gancho. Este tipo de pene permite una cópula segura y efectiva en ambientes acuáticos.
El clasper en los peces pertenece a la categoría de penes intromitentes. El clasper es un órgano reproductor presente en algunos grupos de peces, como los tiburones, las rayas y las quimeras. Está ubicado en la región pélvica de los machos y se utiliza durante el proceso de reproducción para transferir el esperma a la hembra.
El clasper es una estructura especializada que puede variar en forma y tamaño según la especie. Consiste en una prolongación alargada y a menudo con ganchos o espinas en su extremo, que se inserta en la hembra durante la cópula. Esta adaptación permite que el esperma se transfiera de manera eficiente a través del clasper hacia el tracto reproductor de la hembra.
La presencia del clasper en los peces es una característica distintiva y esencial para la reproducción en estas especies. Su evolución y diversificación reflejan las diferentes estrategias reproductivas y adaptaciones específicas de cada grupo de peces.
(c) Hemipenes: Algunos peces, como los lagartos acuáticos del género Lepidophyma, presentan una estructura llamada hemipene. Los hemipenes son órganos reproductores bifurcados que se insertan en la hembra durante la cópula. Un ejemplo de pez con hemipenes es el género Gambusia, que incluye especies como el mosquito de agua (Gambusia affinis).
Los peces exhiben una amplia variedad de penes adaptados para la reproducción exitosa en diferentes entornos acuáticos. Desde penes externos hasta penes intromitentes y hemipenes, estas estructuras reproductivas demuestran la increíble diversidad evolutiva en el mundo de los peces. Comprender la clasificación y las adaptaciones de los penes en los peces nos proporciona información valiosa sobre sus estrategias reproductivas y la evolución de la reproducción sexual en el reino animal.
Es importante destacar que el estudio de los penes en los peces continúa siendo un campo de investigación activo, con muchas especies aún por descubrir y analizar. Estas investigaciones contribuyen a nuestro conocimiento sobre la diversidad biológica y las adaptaciones en la reproducción, y nos permiten apreciar la asombrosa complejidad y variedad de la vida acuática.
8. Desarrollo embrionario y ciclo de vida
8. Desarrollo embrionario y ciclo de vida
En general es típico que ignoremos las complejidades de los ciclos de vida de los vertebrados, pensando de manera descuidada y superficial que son semejantes al nuestro. De hecho, solemos creer que el ciclo de vida más complejo de los vertebrados es el de los anfibios, pues a ellos asociamos la metamorfosis, como si este rasgo apareciera de la nada. En los siguientes dos capítulos discutiremos las complejidades del ciclo de vida de los peces.
Blastulación y gastrulación
Después de la fertilización, el corion del huevo se endurece, un proceso conocido como endurecimiento con agua, que sirve para proteger el embrión en desarrollo. La embriogénesis en peces procede como en la mayoría de los vertebrados. El embrión se desarrolla sobre la yema; la yema se concentra en el polo vegetativo y el óvulo fertilizado se considera telolecítico. En mixinos, elasmobranquios y teleósteos, la escisión es meroblástica, ya que la división celular se produce en la pequeña capa del citoplasma que se convertirá en un pez, pero no en la yema. Las lampreas tienen una escisión holoblástica, mientras que la aleta roja, el gar y el esturión exhiben una forma intermedia, denominada semiholoblastic. La división celular y la diferenciación continúan en secuencias bastante predecibles, con muchas diferencias interespecíficas en el tiempo de aparición de diferentes estructuras.

Figura 8.1. El centro del huevo, yema o Vitelio es un almacén de nutrientes, mientras que el embrión se desarrolla lateralmente, los huevos de los peces son permeables a los gases y a los desechos, así que permiten excretar al ambiente, pero a cambio son susceptibles a una rápida desecación.
Desarrollo de órganos
Los órganos y estructuras que se desarrollan al menos parcialmente en muchos embriones antes de la eclosión incluyen: somitas corporales, que son precursoras de los bloques musculares; conductos renales; tubo neural; vesículas ópticas y auditivas; placodios de lentes oculares; melanóforos de cabeza y cuerpo; un corazón palpitante y un sistema circulatorio en funcionamiento, muchos de los cuales están vinculados a los vasos circulatorios en la yema; Aletas pectorales y pliegues de la aleta media, pero no las aletas medianas en sí mismas; cubiertas operculares, pero no arcos branquiales y filamentos; otolitos; Órganos de los sentidos de la línea lateral, debajo de los cuales se formarán escamas; y el notocordio. En el momento de la eclosión, la cabeza y la cola se levantan de la yema aun unidos a esta, la boca y las mandíbulas apenas se han formado, los radios de la aleta pueden estar presentes en la aleta caudal, pero el esqueleto no se ha osificado. El intestino no funcional es un tubo simple y recto, y la vejiga natatoria es evidente como una pequeña evaginación del tubo intestinal. Los embriones avanzados se enrollan alrededor de sí mismos, sus cuerpos hacen más que un círculo completo. En los peces que eclosionan en una etapa relativamente poco desarrollada, el ojo rara vez está pigmentado, lo que mantiene la transparencia del embrión libre, indefenso y recién nacido, con su saco de yema muy grande y engorroso.
Eclosión ovípara
Justo antes del rompimiento, las células de las glándulas en la cabeza de algunos peces secretan enzimas proteolíticas que ayudan a romper la membrana coriónica o "cáscara de huevo". Los movimientos de sacudida de la cola y el cuerpo ayudan en el proceso de eclosión. El tiempo que pasan en el huevo antes de la eclosión varía mucho entre las especies, desde alrededor de 12 h en algunos peces de arrecife de coral; a unos pocos días como en algunos cíclidos y peces cirujano; a una o dos semanas en melones, dardos, caballas y peces planos; A varias semanas o meses en salmones y tiburones. En todos los peces estudiados, el tiempo de eclosión disminuye al aumentar la temperatura. La "eclosión" es un punto de referencia conveniente durante el desarrollo temprano, pero debe observarse que la etapa exacta de desarrollo en la cual el embrión se libera del huevo varía mucho entre las especies e incluso varía dentro de las especies, dependiendo de la temperatura y el contenido de oxígeno del agua, entre otros factores.
La identificación de especies de huevos no fertilizados o recientemente fertilizados puede ser una tarea difícil, que a menudo requiere microscopía electrónica. Los embriones, especialmente aquellos en etapas avanzadas de desarrollo, son considerablemente más fáciles de identificar. Los patrones de pigmentación específicos de la especie a menudo se desarrollan en el cuerpo y los pliegues, especialmente de las especies marinas (por ejemplo, bacalao, pez plano). Otros caracteres de uso en la identificación incluyen la morfología general de la cabeza, el intestino y la cola, el número de miómeros corporales y la existencia de radios de aleta alargados o desarrollados de manera precocial (por ejemplo, peces voladores, peces cinta, peces arena, peces de fondo).
Eclosión vivipara
Las especies pueden asignarse a categorías según el lugar donde se produce el desarrollo embrionario y si los embriones en desarrollo dependen de la provisión materna y la yema. Si la madre libera huevos que luego dependen de la yema para la nutrición, la especie es ovípara, como es el caso de las quimeras, algunos tiburones y patines, y la mayoría de los peces óseos. Si los jóvenes se desarrollan dentro de la madre y la madre proporciona nutrición a través de una conexión placentaria, secreciones o huevos no embrionados y embriones adicionales débiles que se comen al desarrollarse jóvenes más fuertes, la especie es vivípara. Las crías de peces vivíparos generalmente nacen como juveniles, habiendo pasado por alto una etapa larvaria libre.
La viviparidad caracteriza aproximadamente la mitad de las 1200 especies de peces cartilaginosos y aproximadamente 500 (2%) de las más de 25,000 especies de peces óseos. Prácticamente todas las formas conocidas de viviparidad de los vertebrados y las posibles relaciones materno-fetales se pueden encontrar en los peces. Algunos investigadores reconocen la ovovivipariedad, por lo que los jóvenes se desarrollan dentro de la madre, pero dependen principalmente de la yema que se depositó durante la ovogénesis (por ejemplo, algunos tiburones, scorpaenidos, las anguilas Zoarcidae y, posiblemente, los peces millón). Sin embargo, existen muchas condiciones intermedias que involucran secreciones de yema y maternidad, difuminando las distinciones y haciendo que la ovoviviparidad sea difícil de definir. Aunque el nacimiento vivo generalmente sigue a la fertilización interna, los dos no son sinónimos. Muchos elasmobranquios tienen fertilización interna, pero ponen huevos que se desarrollan durante muchos meses fuera de la hembra.
Variación merística
El desarrollo embrionario generalmente progresa según las instrucciones establecidas en el proyecto genético, pero el momento e incluso los detalles del desarrollo son bastante sensibles a las influencias ambientales. Los contaminantes y los cambios químicos en el agua a menudo dan como resultado anormalidades en las larvas y pueden usarse para monitorear la calidad ambiental. Pero incluso la variación natural de la temperatura, el contenido de oxígeno, la salinidad, la intensidad de la luz, el fotoperíodo o el dióxido de carbono pueden afectar el desarrollo. Se sabe que los rasgos merísticos, como el número de radios de aleta, vértebras, hileras de escamas laterales, miotomas y raspadores de branquias varían en relación con las condiciones ambientales, lo cual revela un aspecto importante de los peces, son morfológicamente plásticos y por ende tienen un mayor potencial evolutivo.
9. Las larvas
9. Las larvas
Biológicamente, la etapa larvaria o su equivalente es probablemente el período más estudiado de la historia de vida temprana de los peces. Esto se debe a la identificabilidad de las larvas, su abundancia relativa y su importancia para determinar la distribución y la abundancia posterior de muchas especies, en particular las de valor comercial. La vida larvaria generalmente comienza cuando el pez sale del huevo y pasa de las reservas internas de yema a las fuentes externas de alimentos planctónicos. Las crías que nadan libremente aún pueden tener un gran saco vitelino y ser llamadas embriones libres, o larva de saco vitelino hasta que la yema se absorba. “Fry” es un término no específico inglés sin equivalente cercano en español que se usa a menudo para larvas avanzadas o juveniles tempranos llamados alevines; la etapa de natación a menudo se refiere a embriones o larvas libres que inicialmente estaban en un nido pero que han crecido capaces de nadar por encima del nido.

Figura 9.1. Larva recién nacida. Alevín de salmón liberándose del huevo. La larva ha roto y está descartando la envoltura del huevo. En aproximadamente 24 horas, absorberá el saco de la yema restante y se convertirá en un juvenil.
La etapa larvaria continúa hasta que se completa el desarrollo del esqueleto axial, las aletas y los sistemas de los órganos. Los radios de la aleta medianos se desarrollan, primero como rayos interespinosos cortos y carnosos; Los verdaderos radios de la aleta y las espinas se desarrollan más tarde entre los radios interespinosos. Las escamas se desarrollan, primero a lo largo de la línea lateral cerca del pedúnculo caudal, luego en filas dorsal y ventral a la línea lateral, y luego se extienden hacia delante. Una vez que se alcanza el
complemento completo de escamas, el número permanece fijo. El final de la notocorda, denominado urostilo, se flexiona hacia arriba y se desarrolla una placa hipural triangular justo debajo de ella. Los rayos caudales crecen posteriormente desde los elementos de la placa hipural. Se desarrollan patrones característicos de pigmentación larvaria, los ojos se pigmentan; y la boca y el ano se abren y se vuelven funcionales. Hasta que se desarrollan los filamentos de las branquias, la larva se basa en la respiración cutánea, que involucra en gran medida la absorción de oxígeno a través de las paredes delgadas de los pliegues primordiales.
Al principio, el sistema circulatorio es relativamente abierto, y la sangre libre de corpúsculos se bombea a través de los senos alrededor de la yema y entre las membranas de la aleta. Las larvas pelágicas, que son particularmente comunes entre los peces marinos, también caracterizan a las especies de lagos y ríos en el agua dulce. Los períodos larvarios en las larvas pelágicas varían ampliamente en duración, de 1 a 2 semanas en sardinas y peces escorpóidos, a aproximadamente 1 mes en muchas especies de arrecifes de coral, a varios meses o incluso años en anguilas. Una larga existencia larvaria indudablemente ayuda en la dispersión de estos jóvenes a hábitats apropiados. Los peces corrientes, tales como muchos peces pequeños, dardos y escultinas, generalmente tienen larvas demersales con períodos larvales cortos. Los peces jóvenes permanecen en el fondo entre las rocas o la vegetación hasta que desarrollan habilidades razonables para nadar, lo que refleja las condiciones turbulentas de los arroyos que pueden dañar las larvas o llevarlas río abajo a hábitats subóptimos como los lagos o el océano.
Una vez que se forman las características musculares y esqueléticas, muchas se capturan en redes de deriva estacionarias, lo que indica una fase de dispersión juvenil, en oposición a la dispersión de larvas en especies pelágicas, lacustres y fluviales. Las larvas de algunos peces intermareales rocosos no pueden dispersarse mar adentro, sino que, en cambio, pasan la totalidad de su existencia larvaria a 5 m de la costa, lo que podría garantizar su regreso a un hábitat adecuado.
Los periodos larvarios se omiten o son muy cortos en los peces vivíparos y en los peces que todavía poseen reservas considerables de yema después de la eclosión, como los salmónidos. Podría decirse que no existe fase larvaria o juvenil en la Perca Enana vivípara, Micrometrus minimus, cuyos machos son sexualmente maduros al nacer. El desarrollo que involucra una fase larvaria con metamorfosis distinta en la etapa juvenil se denomina indirecto. El desarrollo directo ocurre si una etapa larvaria es muy breve o no se puede definir, es decir, el pez se convierte en un adulto en miniatura pero inmaduro, como ocurre en muchas formas marinas costeras y en muchos bagres, salmóniformes y cóccidos.
Formas larvarias
Mientras que los huevos tienden a ser generalmente similares en muchos taxones, las larvas que emergen son sorprendentemente distintas y, a menudo, bastante extrañas en comparación con nuestras expectativas de morfología de los peces.

Figura 9.2. Etapas de la historia temprana de la vida del Trachurus symmetricus.
Los desafíos para los ictiólogos incluyen la identificación y vinculación de larvas con sus contrapartes adultas. Igualmente, desafiante es comprender el significado adaptativo de las diversas estructuras, aparentemente incongruentes, que poseen muchas larvas y que luego pierden a medida que se metamorfosean en juveniles. Una de las claves es que las larvas, aunque son capaces de locomoción, son inicialmente, al menos, relativamente indefensas y vulnerables. Son demasiado lentos para evitar activamente a la mayoría de los depredadores, aparte de aquellos que también flotan con corrientes, como varios cnidarios con células urticantes “medusas”. Muchos depredadores de larvas pelágicas son pequeños, limitados por la mordedura y orientados visualmente a sus presas.
Para contrarrestar a estos depredadores, las larvas dependen de estructuras que los hacen espinosos y aumentan las dimensiones de su cuerpo, o imitan a los animales planctónicos potencialmente nocivos, como los sifonóforos. Algunas estructuras, como las aletas extendidas, los colgajos de piel y los recubrimientos corporales gelatinosos también pueden reducir la velocidad de hundimiento de las larvas, manteniéndolas en aguas superficiales más ricas en nutrientes. Los patrones de pigmentación, que a menudo son característicos de los estadios larvales y son útiles para la identificación, pueden proteger contra los dañinos rayos ultravioleta. Esto se aplicaría particularmente a la gran pigmentación melanística que se encuentra en muchas especies de larvas marinas neustónicas o que habitan en la superficie. Las larvas de peces marinos han demostrado ser misteriosas para los ictiólogos, y nuestro conocimiento ha crecido de manera lenta e incremental. Parte de esta historia se refleja en los nombres dados a los estadios larvales, donde se requirieron muchos años de búsqueda para vincular animales larvales y adultos.
La metamorforsis
La metamorfosis altera de manera tan radical a muchas especies que las dos etapas tienen poca semejanza obvia, atestiguando diferentes regímenes selectivos y adaptaciones de diferentes etapas de la historia de vida. La situación puede complicarse aún más por etapas intermedias entre larvas y juveniles, a veces llamados prejuveniles, que también son distintos, por lo que en los peces podríamos hablar de varias metamorfosis. Los cambios morfológicos son tan marcados que muchas especies únicas recibieron nombres taxonómicos independientes para sus etapas pre y postmetamórficas.

Figura 9.3. La metamorfosis. Diversas etapas en el desarrollo de Protopterus spp., este es un género de pez pulmonado propio de África, el único de la familia Protopteridae. Son peces alargados, con aspecto de anguila con las aletas pectorales y pélvicas muy delgadas, filiformes, y la dorsal y caudal fusionadas. Tienen el cuerpo cubierto por escamas blandas. Pueden nadar ondulando el cuerpo o reptar por el fondo usando sus aletas pares. Los ejemplares mayores alcanzan los 2 metros de longitud. Estos organismos pasan una metamorfosis semejante a la de los anfibios, con alevines (A, B, C) que respiran por branquias externas y adultos con pulmones verdaderos (D).
Las metamorfosis por definición implican cambios importantes en la anatomía, fisiología y comportamiento de un animal. La transición de larva a juvenil en muchos peces involucra complejos conjuntos de cambios que con frecuencia incluyen cambios importantes en los hábitos de alimentación y el hábitat. Estas alteraciones requieren la descomposición y reelaboración de las estructuras embrionarias y larvales y la reconstrucción de estructuras adultas que funcionarán en condiciones ambientales muy diferentes. Un breve ejemplo involucra a las lampreas de mar. Las lampreas larvales, denominadas ammocoetes, son animales sedentarios, ciegos, de agua dulce que residen en madrigueras en fondos limosos y filtran la materia suspendida del agua. En la metamorfosis a la etapa juvenil, este animal se transforma en un depredador / parásito con boca suctorial, lengua áspera, glándulas salivales que secretan anticoagulantes, ojos funcionales, ventilación de marea y la capacidad de vivir en el agua de mar.
Ciclos de vida complejos en vertebrados
Dos rasgos generales compartidos por la mayoría de los peces los diferencian de la mayoría de las especies de vertebrados y también subyacen en muchas de sus adaptaciones más interesantes. Estos dos rasgos son el crecimiento indeterminado y una etapa larvaria. Muchos peces emergen de un huevo como una larva, que tiene poca semejanza anatómica, fisiológica, comportamental o ecológica con el juvenil o adulto en el que los peces finalmente se transformarán. De hecho, el crecimiento continuo mueve a cada individuo a través de una progresión de etapas de su historia de vida, que difieren en la mayoría de los rasgos, creando un espectro de estructuras en continuo cambio, sobre los cuales ha operado la selección natural.

Figura 9.4. Los peces poseen los ciclos de vida más complejos entre los vertebrados, donde algunas especies atraviesan más de una etapa metamórfica.

Figura 9.5. Ciclos de vida de los peces 2. Los peces poseen los ciclos de vida más complejos al interior de los vertebrados, los cuales se han ido simplificando en el curso evolutivo. Dependiendo de la especie estudiada, el número de etapas inmaduras y sus nombres cambian mucho, siendo la más simple: huevo, larva, juvenil y adulto, aunque podemos tener otras nomenclaturas, a la izquierda tenemos: huevo, larva, pececillo, alevín, juvenil, adulto, adulto reproductivo. A la izquierda tenemos algunas etapas en la que puede distinguirse la larva: con Vitelio, preflexión del notocordio, durante la flexión del notocordio, posterior a la flexión del notocordio.
Crecimiento intermedio
Describe el aumento continuo en longitud y volumen que ocurre en la mayoría de los peces a lo largo de sus vidas. Aunque este crecimiento puede disminuir considerablemente a medida que el pez envejece, el potencial de un aumento continuo afecta profundamente a muchos, si no a la mayoría de los aspectos de la vida del pez. Con respecto a la mayoría de los rasgos, un tamaño corporal más grande parece conferir una ventaja, al menos dentro de una especie. La reproducción está íntimamente ligada al tamaño corporal en términos de número y tamaño de huevos, hembras más grandes producen más y más grandes huevos. La elección de pareja por parte de machos y hembras a menudo favorece a individuos más grandes, y los peces más grandes son más capaces de defender un territorio de desove.
La energía de la natación y la bajada interactúan con el tamaño corporal: los peces tienden a compartir con individuos del mismo tamaño, y los peces más grandes pueden nadar más rápido y migrar en distancias más grandes. La tasa de depredación suele ser mayor en los peces más pequeños, y los peces pequeños pueden verse obligados a alimentarse de forma rentable áreas habitadas por depredadores o conspecíficos “individuos de la misma especie” más grandes que pueden canibalizarlos. El crecimiento indeterminado conduce a poblaciones y comunidades estructuradas por tamaño, no por especie, en las que individuos de diferente tamaño funcionan esencialmente como especies diferentes, el llamado nicho ontogenético.
Las limitaciones fisiológicas del tamaño corporal pequeño pueden explicarse por el crecimiento alométrico (proporcional) de muchas estructuras, como el aumento de la agudeza visual y la sensibilidad que se producen cuando un pez crece. El forraje también se ve afectado por el tamaño del cuerpo, no solo porque muchos peces están limitados por el tamaño mandibular y la imposibilidad de masticar, por lo tanto, solo pueden comer cosas que pueden tragar enteras, sino porque muchos tipos de presas no están disponibles para los peces jóvenes hasta que los sitios de inserción y las masas musculares alcanzan un nivel capaz de superar las defensas de la presa.
Nomenclatura del ciclo de vida de un pez
Dada la diversidad y complejidad de las etapas, estados, fases o intervalos en la historia de vida temprana de los peces, no es sorprendente que se hayan desarrollado varios sistemas de clasificación para describir estas etapas, cada una de las cuales difiere ligeramente o mucho en terminología.
Todos estos esquemas intentan subdividir alrededor de una docena de eventos generales reconocibles durante el desarrollo en una progresión coherente y descriptiva. La clasificación más simple reconoce un huevo (que después de la fertilización o activación contiene un embrión en desarrollo), que se convierte en una larva, que se metamorfosea en un juvenil. Las subdivisiones de esta secuencia básica generalmente involucran eventos de punto final, algunos de los cuales ocurren rápidamente, otros gradualmente. Los eventos significativos de punto final incluyen el cierre del blastoporo y el levantamiento de la cola del embrión en desarrollo; absorción del saco vitelino, alimentación independiente y flexión del notocordio de la larva; desarrollo de radios de aletas, escamas y pigmentación; y cambios en las proporciones corporales del juvenil.
Estas descripciones generales cubren una secuencia más complicada de eventos que involucran cambios en la anatomía, fisiología, comportamiento y ecología de un pez en desarrollo. Desde un punto de vista sistemático, la mayoría de las especies de peces son fácilmente distinguibles como tales desde las primeras etapas. En consecuencia, las etapas tempranas de la historia de la vida han desempeñado un papel importante en la sistemática de los peces.
Parte de la controversia sobre la terminología del desarrollo surge de la gran diversidad de tipos embrionarios y larvales, las tasas de desarrollo y las etapas o eventos de transición que existen entre las más de 27,000 especies de peces. Los intentos de generalización se ven frustrados por la excepción y el matiz, y si la investigación se centra en especies marinas o de agua dulce, pelágicos o de profundidad; jóvenes, portadores de huevos o vivíparos entre otras categorías. Algunos autores sostienen que el desarrollo es un proceso continuo y gradual y que la designación de etapas exactas es un proceso arbitrario. Otros sostienen que el desarrollo es saltatorio, que ocurre con períodos de cambio gradual salpicado por eventos o umbrales significativos que permiten un cambio rápido, como el cambio de la dependencia de la yema o las secreciones maternas a la alimentación independiente y exógena.
En cualquier caso, la conclusión principal es que nuestro concepto de metamorfosis debe extenderse desde los anfibios a los peces y los anfibios, y que son los peces los que demuestran una mayor cantidad de etapas metamórficas, las cuales se han ido simplificando en el curso de la evolución.
Los juveniles
La eclosión o el nacimiento y el inicio de la alimentación exógena representan dos eventos históricos en la vida temprana de un pez. También es importante para muchas especies el cambio de hábitat de larvas a juveniles, una transición que a menudo implica el asentamiento de la columna de agua y el supuesto de una existencia casi bentónica. Tradicionalmente, se considera que la fase larvaria termina y la fase juvenil comienza cuando se pierden los caracteres larvarios y el esqueleto axial, los sistemas de órganos, la pigmentación, la escamación y las aletas se desarrollan por completo, momento en el que los peces parecen esencialmente un adulto en miniatura. Esta transición puede ser breve y relativamente simple, requiriendo minutos u horas en algunas damiselas, o puede ser muy larga y complicada, requiriendo varias semanas en salmones, peces ardilla, gobios y peces planos.

Figura 9.6. Los ostariofisiarios es un superorden de peces teleósteos siendo el segundo superorden de peces más grande que existe abarcando al menos 8000 especies, lo que representa alrededor del 28% de todas las especies de peces conocidos en el mundo y 68% de las especies de agua dulce, presentes en la mayoría de los continentes excepto Antártida. Poseen en común cierto número de características como una sustancia de alarma y el aparato de Weber. Miembros de este grupo incluyen peces usados en actividades humanas, como la industria alimentaria, pesca deportiva, investigación y en acuarios.
Algunas adaptaciones complejas que definen esencialmente los principales grupos taxonómicos no aparecen hasta la fase juvenil. Un ejemplo es la reacción de alarma del ostariofisiarios. Los peces ostariofisiarios tienen una respuesta de escape característica a la sustancia de alarma, una sustancia química que se libera de la piel de los conespecíficos lesionados. La reacción de alarma aparece relativamente tarde en el desarrollo, después de que se desarrolla el comportamiento de agrupación y después de que los peces ya pueden producir una sustancia de alarma en sus células epidérmicas. Sin embargo, la reacción de alarma está programada genéticamente. Después de 51 días después de la eclosión, los pececillos europeos, Phoxinus phoxinus, reaccionan a la sustancia de alarma en el agua la primera vez que la encuentran, independientemente de su experiencia con depredadores.

Figura 9.7. La trucha de manantial (juvenil arriba y adulto abajo), trucha de arroyo o salvelino (Salvelinus fontinalis) es una especie de pez de la familia Salmonidae. La trucha de arroyo, también conocida como salvelino (Salvelinus fontinalis), es una especie de pez de agua dulce que se encuentra en arroyos y ríos de América del Norte. Es apreciada tanto por su valor deportivo como por su calidad culinaria. Las truchas de arroyo tienen un cuerpo alargado y aerodinámico, con una coloración característica que varía desde tonos verdosos hasta plateados, con manchas rojas y negras en su dorso y flancos. Son excelentes nadadoras y se alimentan principalmente de insectos acuáticos y pequeños peces. Estas truchas son consideradas una especie indicadora de la calidad del agua, ya que son muy sensibles a la contaminación y a los cambios en el ecosistema acuático. Su conservación y manejo adecuado son fundamentales para mantener la salud de los ecosistemas acuáticos y garantizar su presencia en el futuro.
Aunque los huevos y las larvas son, con mucho, las etapas más vulnerables durante la historia de vida de un individuo, el logro de la etapa juvenil aún implica una fuerte selección para la adquisición exitosa de alimentos y la capacidad de evitación de depredadores. La interacción entre estos factores está ejemplificada por la trucha de arroyo juvenil, Salvelinus fontinalis. La trucha de manantial no se somete a la transformación por muda característica de muchos otros salmónidos. En cambio, eclosionan en primavera y se instalan en arroyos pequeños y poco profundos. Su tarea principal es adquirir suficientes reservas de energía durante el primer verano para superar el período invernal de escasa disponibilidad de alimentos. Los juveniles más grandes tienen una mayor probabilidad de sobrevivir al primer invierno. Para adquirir energía y crecer, deben establecer y defender un territorio de alimentación. Los mejores territorios se encuentran en aguas relativamente poco profundas, lo que expone a los peces a depredadores acuáticos y aéreos. Los depredadores pueden evitarse permaneciendo inmóviles, pero los peces inmóviles no pueden perseguir a sus presas ni repeler a los invasores territoriales. La alimentación y las peleas distraen al individuo de evitar a los depredadores.
Básicamente, los peces más pequeños corren más riesgos y tienden a alimentarse de manera más extensa y abierta, mientras que los peces más grandes están menos dispuestos a aceptar los riesgos de depredación y están más dispuestos a interrumpir su alimentación tomando medidas evasivas. La mayor probabilidad de inanición en invierno obliga a los juveniles más pequeños a hacer un intercambio entre el riesgo de depredación y la búsqueda de alimento de manera diferente a los peces más grandes de la misma edad.
10. La metamorfosis en los peces
10. La metamorfosis en los peces
La abundancia numérica de estadios larvarios de muchas especies y los procesos que reducen esta abundancia han sido un foco importante de investigación ictiológica. Pero el crecimiento y el cambio continúan durante la vida de un pez. Dado que la mayoría de los peces de importancia comercial se explotan en la edad adulta, se ha estudiado ampliamente el crecimiento y la maduración de los juveniles hasta la edad adulta. Además, muchos peces atraviesan un período posreproductivo cuando los sistemas de órganos se degeneran, lo que proporciona un modelo comparativo para estudiar el proceso de envejecimiento y la vejez, temas de importancia en la biología humana.
Etapas de transición
Las etapas de transición complican la búsqueda de una terminología descriptiva universal sobre la historia de la vida temprana. También hacen que sea difícil identificar cuándo los peces cambian de una forma de desarrollo a otra. Los estadios de transición ocurren con mayor frecuencia entre los períodos larvario y juvenil y entre los períodos juvenil y adulto. La fase de transición entre larva y juvenil en los peces de arrecife se ha denominado de diversas formas como postlarval, larvaria tardía, nuevo recluta, juvenil recluta, juvenil pelágico, juvenil de transición y colono. La fase de transición puede ser de duración variable, incluso dentro de una especie. Esta variabilidad tiene sentido cuando se da cuenta de que un pez joven puede no encontrar un hábitat apropiado para su siguiente etapa simultáneamente con su capacidad para hacer la transición a esa etapa. Por lo tanto, si se viera obligada a asentarse del plancton en el día 35 de desarrollo, o en el momento en que los elementos esqueléticos se osificaran y los rayos de las aletas se desarrollaran por completo, una larva que todavía estaba lejos en el mar podría no tener más remedio que hundirse hasta el fondo. varios kilómetros más abajo y morir de hambre o morir de frío.

Figura 10.1. Gobiosoma bosc es una especie de peces de la familia de los Gobiidae en el orden de los Perciformes. Gobiosoma bosc, conocido comúnmente como gobio rayado o gobio de rayas, es una especie de pez perteneciente a la familia Gobiidae. Se encuentra en aguas costeras del océano Atlántico, desde Carolina del Norte hasta Brasil. Este pequeño pez presenta un cuerpo alargado y estrecho, con una coloración que varía entre tonos marrones y amarillos, decorado con franjas verticales oscuras. Es conocido por su habilidad para camuflarse en su entorno rocoso o en los arrecifes de coral. El gobio rayado es un pez territorial y se alimenta de pequeños crustáceos y otros invertebrados. Es apreciado en acuarios marinos por su colorido y comportamiento interesante
La variabilidad en el período larvario es evidente en las larvas del gobio desnudo, Gobiosoma bosci. Estas larvas se asientan en el plancton y toman una existencia bentónica y escolar durante hasta 20 días antes de transformarse en juveniles solitarios. Otros gobios y un lábrido pueden tener una “ventana de oportunidad” de 20 a 40 días durante la cual pueden buscar un hábitat apropiado como larvas sin transformarse en la forma juvenil más sedentaria. Los peces planos pueden retrasar la transformación a la forma juvenil si no encuentran un hábitat juvenil apropiado; lo hacen alternando entre asentarse en el fondo y nadar por encima de él. Se han observado preferencias de sustrato, que implican la búsqueda activa de un hábitat apropiado, en numerosas larvas. Las observaciones directas del asentamiento de especies de arrecifes de coral indican que tal flexibilidad puede estar relativamente extendida y que el asentamiento y la transición de larva a juvenil no deben verse como una decisión de todo o nada. Una vez que se adquiere la competencia de asentamiento, muchas larvas pueden tener días o incluso semanas antes de que deban asentarse y asumir hábitos juveniles.
La metamorfosis
Las metamorfosis por definición implican cambios importantes en la anatomía, fisiología y comportamiento de un animal. La transición de larva a juvenil en muchos peces implica complejos conjuntos de cambios que con frecuencia incluyen cambios importantes en los hábitos de alimentación y el hábitat. Estas alteraciones requieren una descomposición y reelaboración de las estructuras embrionarias y larvarias y una reconstrucción en estructuras adultas que funcionarán en condiciones ambientales muy diferentes. Un breve ejemplo son las lampreas de mar. Las lampreas larvales, llamadas ammocoetes, son animales sedentarios, ciegos, de agua dulce que residen en madrigueras en fondos limosos y filtran la materia suspendida del agua. En la metamorfosis a la etapa juvenil, este animal se transforma en un depredador / parásito con boca succionadora, lengua áspera, glándulas salivales que secretan anticoagulantes, ojos funcionales, ventilación de las mareas y la capacidad de vivir en el agua del mar. Se podrían mencionar muchos otros taxones, pero los detalles de dos grupos bien estudiados, el salmón y los peces planos, servirán para ejemplificar la complejidad de las reelaboraciones que implican el cambio de un animal adaptado a la existencia larvaria en uno adaptado para enfrentar los desafíos de etapas posteriores en su historia de vida.
La esmoltificación
El interés generalizado en los salmónidos ha dado como resultado un conocimiento detallado y una terminología especial asociada con diferentes etapas del ciclo de vida. Por lo general, el salmón y la trucha desovan en nidos de grava llamados redds, los huevos eclosionan en alevines tipo (larvas del saco vitelino) que reabsorben la yema y se convierten en alevines tipo frey. Los alevines frey desarrollan patrones típicos de la especie de barras verticales en sus lados llamados marcas de parr, el pez ahora se llama parr. Después de unos meses o años, dependiendo de la especie y la población, la pareja de especies anádromas que pasan su vida juvenil en el mar (salmones del Pacífico y del Atlántico, trucha Steelhead) se desplaza río abajo como smolts plateados. Los procesos asociados con la migración río abajo de los smolts se encuentran entre los aspectos biológicos más intrigantes y mejor estudiados de la historia de vida temprana de los peces.

Figura 10.2. Ciclo de vida del salmón.
La esmoltificación es un fenómeno complejo que implica reelaboraciones de casi todas las características de un salmón joven. Una característica interesante de los cambios es que son preparatorios: ocurren cuando el animal cambia de un parr a un smolt en agua dulce, anticipándose a las condiciones ambientales que los peces jóvenes encontrarán más tarde después de ingresar al océano. Anatómicamente, los smolts cambian de sombreado y barrado a plateado, que es una mejor forma de camuflaje en mar abierto . También adquieren una forma más delgada y aerodinámica que implica una reducción de la condición a medida que se consumen los lípidos corporales. El plateado es el resultado de un aumento en la densidad de los cristales de purina, principalmente guanina pero también hipoxantina, que se depositan debajo de las escamas y en lo profundo de la dermis. A pesar de la pérdida de lípidos, los smolts son más flotantes que los conespecíficos no migratorios. El aumento de la flotabilidad resulta del aumento del volumen de gas en la vejiga de natatoria, lo que puede reducir los costos energéticos de la migración. La complejidad de las hemoglobinas en la sangre aumenta, lo que afecta la afinidad por el oxígeno entre otros factores respiratorios.
(a) Regulación respiratoria y osmótica: Estas alteraciones preparan un smolt migratorio para las condiciones oceánicas que a menudo incluyen una disponibilidad reducida de oxígeno en comparación con las aguas frías y turbulentas de un arroyo o río. Se producen muchos cambios en la función de las branquias, incluido un aumento del número de células de cloruro y cambios en la permeabilidad iónica y la actividad enzimática. Estos cambios anticipan el paso del ambiente hipoosmótico de agua dulce donde la pérdida de iones es el problema principal al ambiente marino hiperosmótico donde la retención de agua es el principal problema.
(b) Comportamiento: Desde el punto de vista del comportamiento, los parr del salmón del Atlántico son primero muy territoriales en aguas poco profundas, pero luego se mueven hacia aguas más profundas y forman cardúmenes, aunque con frecuencia existe una jerarquía de dominancia en los cardúmenes. Incluso esta agresión disminuye a medida que los peces comienzan a moverse hacia el mar. El movimiento se ve favorecido por una inversión en la reotaxis, la respuesta a las corrientes fluidas que mantenían incluso a los embriones dirigidos río arriba. La reotaxis positiva desaparece cuando los peces en grandes cardúmenes se desplazan río abajo con las corrientes. Es durante el smoling que los peces jóvenes aprenden o imprimen el olor de su corriente de origen, lo que les permite identificarlo entre cientos de alternativas cuando regresan del mar durante la migración de desove.
(c) Hormonas: Muchas de las transformaciones que ocurren durante la smoltificación pueden estar relacionadas con cambios en las hormonas circulantes. Los aumentos de corticosteroides, prolactina y hormona del crecimiento afectan respectivamente el metabolismo de los lípidos, la osmorregulación y el equilibrio mineral. Los niveles de cortisol y estradiol también aumentan. Los niveles de tiroxina también aumentan de forma natural, y las inyecciones experimentales de hormona estimulante de la tiroides pueden inducir muchos de los eventos fisiológicos y conductuales de la smoltificación, como la deposición de purina, la actividad de las enzimas branquiales, el aumento de la actividad de natación, el crecimiento corporal y el consumo de lípidos. Estos cambios sugieren que la hormona tiroidea, que interactúa con el fotoperíodo y los ritmos endógenos, juega un papel importante en el proceso. Tenga en cuenta que este conjunto de hormonas también es el que dispara la metamorfosis en los anfibios y la adolescencia en los seres humanos.
(d) Regulación alimentaria: La smoltificación no se fija de ninguna manera en términos de edad en una especie o incluso en una población. El salmón del Atlántico puede smoltificar entre 1 y 7 años, dependiendo de la temperatura y la latitud. Las oportunidades de alimentación parecen ser el determinante clave del inicio de la smoltificación. Los individuos bien alimentados smoltifican más jóvenes, aunque las diferencias genéticas en la actividad de alimentación pueden hacer que algunos peces dejen de alimentarse y, en consecuencia, retrasen la smoltificación. Alguna evidencia indica un umbral de tamaño: el salmón del Atlántico que no alcanza una longitud de 10 cm en el otoño de la primera temporada de crecimiento es menos probable que esmoltifique el próximo año. La tasa de crecimiento y la edad interactúan con esta longitud umbral hipotética. Los peces de crecimiento más rápido son más propensos a smoltificar, y los peces más viejos pueden hacerlo en un tamaño más pequeño.

Figura 10.3. El salmón rojo del pacífico (Oncorhynchus nerka) posee dos formas “adultas”, una de alimentación oceánica que se camufla y es azul, y otra que es netamente reproductiva, que es la que realiza el viaje de regreso a su río de nacimiento, en la cual adquiere una coloración reproductiva que no es mimética y, por lo tanto, lo convierten en una presa fácil. Lo cual implica que los salmones atraviesan más de una metamorfosis.
(b) Temporada de esmoltificado: El tiempo también es importante. La etapa de smolt en sí dura algunas semanas; si un pez no ingresa al mar, el proceso se revertirá y el pez volverá a la condición de parr. Aunque el salmón del Pacífico (Oncorhynchus spp.) y el salmón del Atlántico se consideran anádromos y, por lo tanto, se someten a smoltificación, estos peces pueden quedarse sin litoral y nunca migrar al mar. Además, algunos individuos dentro de una población pueden pasar por alto las fases migratoria y de smolt y permanecer en agua dulce. Cuando esto sucede en los machos, pueden madurar rápidamente al año de edad y desovar con hembras que regresan la próxima temporada. En el salmón del Atlántico, la proporción de machos precoces difiere entre las poblaciones, oscilando entre el 5% y> 50% de los machos. Los factores que determinan la maduración precoz en los salmones machos son ampliamente debatidos, y la evidencia sugiere que la disponibilidad de alimentos o los factores genéticos son determinantes.
Pez plano asimétrico
La simetría es una característica anatómica casi universal de los animales. La mayoría de los animales, independientemente de su filo, exhiben simetría bilateral en su morfología, con estructuras aproximadamente reflejadas en un espejo a la derecha e izquierda de la línea media. Las desviaciones de la simetría implican funciones y adaptaciones inesperadas. Los biólogos buscan comprender la causa y función de la asimetría en el nivel próximo del control genético y ambiental del desarrollo y en el nivel último de su posible adaptabilidad.
Entre los ejemplos más sorprendentes de asimetría se encuentra la "lateralidad" de los peces planos. Las 14 familias y alrededor de 680 especies de pleuronectiformes (platijas, halibuts, soles, solla, etc.) como grupo se caracterizan por adultos que se encuentran en la parte inferior de un lado de su cuerpo. Sus cuerpos aplanados son funcionalmente análogos a muchos otros peces bentónicos como los tiburones ángel; patines rayos bagres banjo, acorazados con boca de ventosa y bagres de cabeza cuadrada; peces murciélago ogcocéfalos; cabezas planas platycephalidos; y algunos peces escorpión. La principal diferencia es que todos los otros grupos están aplanados en un plano dorsal-ventral (= deprimidos), mientras que los peces planos están aplastados lateralmente (= comprimidos) (la gente suele deprimir las cucarachas y comprimir los mosquitos). Los peces deprimidos mantienen su simetría bilateral a pesar de su extrema morfología.

Figura 10.4. Ogcocephalus radiatus, conocido como pez murciélago ogcocéfalo, es una especie fascinante que habita en aguas tropicales y subtropicales del océano Atlántico occidental. Este pez se caracteriza por su cuerpo aplanado o deprimido en forma de disco y una cabeza grande con una boca extensible. Su coloración varía, pero generalmente presenta tonalidades marrones o verdosas para camuflarse en su entorno. El pez murciélago ogcocéfalo es un maestro del camuflaje, y su apariencia y comportamiento mimetizan con el entorno marino. Puede enterrarse en el lecho marino y utilizar sus aletas modificadas para moverse sobre el fondo marino. Se alimenta de pequeños peces y crustáceos que atrapa rápidamente con su boca extensible. Esta especie es fascinante debido a su morfología única y su habilidad para adaptarse a su entorno. Es un verdadero tesoro de los mares tropicales, cautivando a los observadores marinos con su apariencia y comportamiento únicos.
La mayoría de los peces comprimidos son especies de cuerpo profundo, simétricas bilateralmente que nadan en la columna de agua y usan sus cuerpos aplanados para aumentar la maniobrabilidad o para aumentar la profundidad de su cuerpo contra los depredadores (p. Ej., Carácidos serrasalmina, peces luna centrarquídeas, muchos pompanos, peces dedos monodactílidos, peces mariposa, efípidos peces murciélago y peces espada, y cirujanos). Los peces planos están comprimidos lateralmente, pero se encuentran en el fondo, ya sea en su lado derecho o izquierdo, por lo que se enfrentan al desafío de recibir información sensorial de solo la mitad de sus órganos sensoriales, mientras que la otra mitad está enterrada en la arena o el barro. Las adaptaciones más obvias a su orientación inusual se pueden ver en la estructura y desarrollo de su aparato visual.

Figura 10.5. Pez plano Platichthys stellatus. Platichthys stellatus, conocido como lenguado estrellado o platija estrellada, es una especie de pez plano que pertenece a la familia Pleuronectidae. Se encuentra en el océano Atlántico, desde el Mar del Norte hasta el Mar Báltico y el Mediterráneo. Este pez tiene un cuerpo plano, ovalado y simétrico, con los ojos en un mismo lado de la cabeza. Su coloración varía entre tonos marrones, grises y verdes, y está decorado con manchas y líneas oscuras. La platija estrellada es un depredador camuflado que se entierra en el fondo marino y se alimenta de pequeños peces, crustáceos y moluscos. Es valorada como especie comercial y es apreciada en la gastronomía debido a su carne blanca y sabrosa.
Los peces planos comienzan su vida como larvas pelágicas normales, bilateralmente simétricas. En la platija estrellada, Platichthys stellatus, las larvas emergen del huevo cuando tienen unos 3 mm de largo y comienzan a alimentarse exógenamente. Durante el próximo mes o dos, llevan una vida pelágica normal, hasta que alcanzan una longitud de 7 mm. Luego comienza la metamorfosis a una forma comprimida (el tamaño en la metamorfosis varía entre 4 y 120 mm en diferentes peces planos). La mayoría de los huesos están osificados de forma incompleta en este momento, lo que aparentemente facilita la transformación. El neurocráneo anterior, el encéfalo y las cuencas oculares (órbitas) giran.

Figura 10.6. Migración progresiva del ojo en una platija de verano en desarrollo, Paralicthys dentatus. Cuando la larva del lenguado mide unos 10 mm de largo, el ojo derecho comienza a migrar hacia el lado izquierdo del pez a través de un proceso que incluye la reabsorción ósea y la rotación del neurocráneo del pez. Todo el proceso dura de 3 a 4 semanas, durante las cuales la larva crece entre 5 y 10 mm. La posición del ojo derecho en el lado derecho del cuerpo se representa en las etapas 1 a 3 (círculo tenue). Observe otros cambios en el desarrollo, incluido el desarrollo de las estructuras oculares, la migración anterior de la aleta dorsal, el crecimiento y elaboración de las aletas pectoral y pélvica y el crecimiento de la boca.
Esto permite que un ojo migre realmente a través de la parte superior de la cabeza. En algunas especies de biótidos y paralictíidos, el ojo se mueve a través de una hendidura que aparece entre el cráneo y la base de la aleta dorsal. La aleta dorsal permanece en la línea media o, en algunas especies, crece hacia adelante hasta que la primera espina se encuentra por delante de los ojos. Todo el proceso ocurre rápidamente, durante aproximadamente un período de 5 días en Platichthys stellatus, o en menos de 1 día en algunas especies.
Ocurren otras asimetrías que reflejan la transformación a una existencia bentónica y comprimida. El órgano nasal del lado ciego migra hacia la línea media dorsal; el lado ciego generalmente no está pigmentado, puede carecer de una línea lateral, tiene aletas pectorales y pélvicas más pequeñas y la escamación con frecuencia difiere en los dos lados. Durante la metamorfosis, los conductos semicirculares experimentan un desplazamiento de 90° y la reacción de luz dorsal también cambia de manera apropiada para un pez acostado de lado. En el momento de la metamorfosis o poco después, el pez toma una existencia bentónica y pierde su vejiga natatoria.
11. La determinación ambiental del sexo
11. La determinación ambiental del sexo
El desarrollo de un individuo con funciones reproductivas implica tres procesos muy diferentes (determinación, diferenciación y maduración) que ocurren en diferentes momentos durante la historia de la vida. La determinación del sexo es el proceso mediante el cual se decide la masculinidad o feminidad (género) de un individuo, generalmente durante la ontogenia temprana. La determinación puede controlarse genética o ambientalmente. La diferenciación implica el desarrollo de estructuras gonadales reconocibles (ovarios o testículos) en un individuo, aunque los gametos en maduración no están necesariamente presentes. La maduración implica la producción real de gametos, espermatozoides u óvulos viables. El género de un individuo puede determinarse en la fertilización, el pez puede diferenciarse como juvenil, pero técnicamente no es un adulto hasta que madura, y aun después de eso, algunas especies pueden cambiar de sexo.
Tipos de determinación del sexo
Existe una relación interesante entre la determinación del sexo, el cambio de sexo y la existencia de cromosomas sexuales en los vertebrados. Las aves y los mamíferos tienen cromosomas sexuales identificables. Los mamíferos machos son del sexo heterogamético y poseen un cromosoma sexual X e Y. Lo contrario es válido para las aves, en las que la hembra es el sexo heterogamético, conocido como heterogamety ZW. En aves y mamíferos, el género se determina y se fija en la fertilización. El género de los individuos permanece constante y las condiciones ambientales no tienen ningún efecto sobre la determinación del sexo. Algunos reptiles y anfibios tienen cromosomas sexuales, otros no. En las tortugas, los machos generalmente se producen a bajas temperaturas; lo contrario es válido para los cocodrilos y las lagartijas. En taxones en los que ocurre tal determinación ambiental del sexo, los cromosomas sexuales son relativamente raros.
Grupos de peces con determinación ambiental del sexo
La determinación genética del sexo en los peces puede implicar un control monogénico o poligénico, y los genes y factores determinantes del sexo pueden localizarse en cromosomas autosómicos o en cromosomas sexuales definitivos. Los cromosomas sexuales son relativamente raros entre los peces, y caracterizan a 176 especies en 72 familias, o alrededor del 10% de las aproximadamente 1700 especies para las que se han descrito el número y la morfología de los cromosomas, aunque esto puede subestimar la frecuencia real de heterogamia. Los ejemplos de familias con cromosomas sexuales incluyen varias familias de aguas profundas, como los fundidos batilagidos, los peces hacha esternoptíquidos, los neoscopélidos, los peces linterna mictófidos y las crestas melamácidas. En aguas someras, se ha encontrado heterogamia en rayas; osteoglosidos; anguilas y congrios; carácidos; bagres, silúridos y loricáridos; truchas; pez lagarto; killis; portadores de vida; espinosos; esculpinos y cíclidos; gobios aguja blanca; peces planos; y peces gatillo.
Efectos, utilidades e inconvenientes
Como podría anticiparse en un taxón donde la determinación genética del sexo puede ser la excepción, la determinación del sexo en los peces es bastante flexible y está influenciada por una variedad de factores externos. Esta labilidad ha sido explotada en programas de acuicultura porque permite a los practicantes producir cepas monosexuales de especies económicamente valiosas donde un sexo crece más rápido, alcanza un tamaño mayor, o produce más o menos grasa que el otro sexo. Un inconveniente de la determinación ambiental del sexo generalizada es que hace que los peces sean vulnerables a la degradación ambiental, incluidos los químicos disruptores endocrinos y el cambio climático.
Causas de la ambivalencia sexual
La etapa exacta en la que se determina el género en los peces es controvertida. Aunque la determinación genética probablemente se aplica a la mayoría de las especies, en muchos peces la determinación del sexo puede no estar fijada en la fertilización o incluso durante la ontogenia temprana. Muchos peces pasan por un cambio de sexo prematuro, diferenciándose pero sin madurar primero como hembras, y algunos individuos luego se transforman en machos. Este patrón se sospecha o se conoce de los peces bruja, lampreas, pececillos, salmónidos, cíclidos, peces mariposa, lábridos, loros, gobios y peces belontíidos del paraíso. Tal ambivalencia no es del todo sorprendente cuando se recuerda que todas las gónadas en agnatos y teleóstos se desarrollan a partir de una estructura única, la corteza epitelial, que da lugar a los ovarios en los vertebrados superiores. En los tiburones, los ovarios se desarrollan a partir de la corteza, mientras que los testículos se desarrollan a partir de la médula. En consecuencia, los tiburones no muestran labilidad sexual.
Efecto de la temperatura
La temperatura puede jugar un papel importante como factor ambiental porque la determinación del sexo en los peces es sensible a la alteración térmica. Los estudios experimentales generalmente encuentran masculinización de individuos o proporciones de sexo sesgadas por machos cuando los huevos o larvas de especies de pececillos, gobios, pejerreyes, lochas, peces roca, cíclidos y flotadores se crían a temperaturas más altas, y el efecto aumenta a medida que aumenta la temperatura. La feminización o las proporciones de sexos sesgadas por las hembras han dado lugar a temperaturas más altas en lampreas, salmones, viveros, espinosos y lubinas. Los mecanismos subyacentes a estos efectos parecen implicar una alteración de la actividad enzimática o una alteración endocrina (síntesis de hormonas o alteración de la función del receptor de esteroides). La aromatasa es una enzima ovárica que convierte la testosterona en estradiol, un proceso vital para el crecimiento de los ovocitos. En la tilapia del Nilo, Oreochromis niloticus y la platija japonesa, Pleuronectes olivaceus, las temperaturas elevadas dieron como resultado una masculinización asociada con una actividad de aromatasa reducida.

Figura 11.1. La tilapia del Nilo (Oreochromis niloticus) es una especie de pez de la familia Cichlidae en el orden de los Perciformes. La tilapia del Nilo (Oreochromis niloticus) es una especie de pez de agua dulce originaria de África, pero que se ha introducido en muchas regiones del mundo debido a su valor como alimento y su adaptabilidad. Tiene un cuerpo ovalado y comprimido, con una coloración que varía desde tonos plateados hasta dorados. Es conocida por su rápido crecimiento y alta fecundidad, lo que la convierte en una especie popular en la acuicultura. Además de su importancia económica, la tilapia del Nilo desempeña un papel ecológico al controlar la vegetación acuática y servir como fuente de alimento para otras especies. Sin embargo, en algunas áreas, su introducción ha tenido impactos negativos en los ecosistemas locales.
Otras causas para la determinación ambiental del sexo
Aunque la temperatura es la causa ambiental más común o famosa para la determinación del sexo no fijada a cromosomas, sin duda no es la única. A continuación, plantearemos una lista de otras causas o determinantes ambientales para el sexo en los peces:
(a) "medio ambiente", incluido el clima,
(b) la disponibilidad de alimentos y
(c) las interacciones sociales, Lo cual involucra la presencia o ausencia de peces del sexo opuesto.

Figura 11.2. El pez del paraíso (Macropodus opercularis) es una especie de pez perciforme anabántido de agua dulce de la familia Osphronemidae que se encuentra en China (en la cuenca del Yangtsé), Taiwán, Corea y el norte de Vietnam.
En el pez paraíso, Macropodus opercularis, todos los individuos comienzan como hembras y algunos luego se diferencian como machos, pero estos cambios ocurren antes de la maduración. La determinación final se basa en el estatus social: los individuos dominantes se convierten en machos y los subordinados en hembras. Las anguilas, a pesar de tener heterogametia aparente ZW, producen más machos en poblaciones densas como una aparente respuesta al hacinamiento. La determinación ambiental del sexo también se ha documentado en salmón rojo, pez arroz o medaka (Oryzias latipes), porteadores vivos poecilíidos, rivulines y peces luchadores siameses, todos en respuesta a temperaturas extremas.

Figura 11.3. El pez arroz o medaka (Oryzias latipes) es una especie de pez de agua dulce nativo de Asia, especialmente Japón y Corea. Es conocido por su pequeño tamaño y su aspecto elegante, con un cuerpo delgado y alargado, y una coloración que varía desde tonos plateados hasta dorados, rojos o naranjas brillantes. El pez arroz es ampliamente utilizado en la investigación científica debido a su facilidad de mantenimiento en acuarios y su corto ciclo de vida. También es valorado como pez ornamental en acuarios domésticos. Además, el medaka ha sido objeto de estudios genéticos y de toxicología debido a su sensibilidad a los contaminantes ambientales. Es una especie versátil y valiosa tanto para la investigación como para la acuicultura.
12. Maduración, longevidad y vejez
12. Maduración, longevidad y vejez
No es sorprendente que la edad en la primera reproducción y la longevidad varíen mucho entre los peces, lo que dificulta identificar patrones o sacar conclusiones. La importancia adaptativa de las diferencias de edad en la primera reproducción se relaciona con las compensaciones entre dedicar energía al crecimiento somático y la reproducción, combinado con las tasas de mortalidad esperadas y la probabilidad de vivir lo suficiente para reproducirse.
Alevines precoces
Los extremos de edad en la primera reproducción incluyen algunas surfpercas embiotócidas, cuyos machos nacen produciendo espermatozoides funcionales. Se ha demostrado que los peces gobioides de los géneros Schindleria y Paedogobius maduran en menos de 2 meses, mientras que Schindleria madura en tan solo 3 semanas. Muchos peces pequeños de arroyo maduran en 1 año, siendo reproductivamente activos durante la temporada de desove después de que nacen (por ejemplo, la mayoría de los dardos), aunque la maduración puede demorar más en las poblaciones de latitudes más altas.
Adultez tardía
En el otro extremo, los esturiones y algunos tiburones pueden tardar entre 10 y 20 años en madurar. Los esturiones pueden vivir entre 80 y 150 años. El tiburón de maduración más lenta documentado es el cazón espinoso, Squalus acanthias, una especie bien conocida por los estudiantes de anatomía comparada.

Figura 12.1. La mielga, galludo o tollo de cacho (Squalus acanthias) es una especie de elasmobranquio escuálido de la familia Squalidae. Es un pequeño tiburón alcanza 1.25 metros de longitud, tiene cuerpo estilizado con espinas venenosas y muy afiladas en las aletas dorsales. El aparato toxicóforo está delante de cada aleta dorsal, con la glándula toxicófora ubicada en el surco dorsal de dicho radio. Cuando el tóxico se inyecta en la piel produce un intenso dolor de la zona, tumefacción y rubor
El cazón espinoso no madura hasta los 20 años y tiene la vida útil más larga registrada de un tiburón, más de 70 años. El récord de reproducción retardada naturalmente entre los peces óseos lo tienen aparentemente las anguilas americanas en Nueva Escocia, que pueden no madurar y emprender su migración de desove de regreso al Mar de los Sargazos hasta los 40 años.
Longevidad
Los patrones de longevidad son solo un poco más definibles. Con muchas excepciones, los peces más grandes generalmente viven más que los peces más pequeños. Los teleósteos más antiguos conocidos son los peces roca escorpénidos del Pacífico nororiental. Los análisis radioisotópicos y de otolitos indican que el pez roca de ojos rojos (Sebastes aleutianus) vive durante 140 años, el pez roca gris plateado (S. borealis) durante 120 años y el pez roca de aguas profundas (S. alutus) durante 90 años Entre las especies deportivas comunes, la perca europea puede vivir 25 años y la lobina negra puede vivir de 15 a 24 años.

Figura 12.2. El pez roca de ojos rojos (Sebastes aleutianus) es una especie de pez actinopterigio de la familia Sebastidae de color rosa, beige o marrón con manchas irregulares de color marrón oscuro y a menudo tiene un parche oscuro en la parte posterior del opérculo. La parte posterior de la línea lateral es a menudo de color rosa. Los adultos tienen un tamaño medio de unos 80 cm
Numerosas especies viven durante un año o menos, incluidos los llamados peces anuales de América del Sur y África. Varios gobios tienen tiempos de generación y vida útil notablemente cortos. El gobio australiano del arrecife de coral Eviota sigillata pasa 3 semanas como larva planctónica, se asienta y madura en 1 a 2 semanas, y vive no más de otras 4 semanas, con una vida útil total de menos de 60 días. El período de vida más corto conocido entre los peces de agua dulce ocurre en un rivulino africano Nothobranchius furzeri, con una esperanza de vida en la naturaleza de unos pocos meses y un período de vida máximo en el laboratorio de menos de 12 semanas. Otras especies de vida corta incluyen los pececillos norteamericanos del género Pimephales, varios peces galaxidos de Tasmania y Nueva Zelanda, retropinnidos, el Ayu japonés entre otros.

Figura 12.3. El rivulino africano (Nothobranchius furzeri) es una especie de pez anual endémico de África Oriental, especialmente de Mozambique y Zimbabue. Es conocido por su ciclo de vida corto y rápido, ya que vive solo por unos pocos meses. Este pez presenta una coloración vibrante y variada, con aletas alargadas y un cuerpo delgado. El rivulino africano es muy valorado en la investigación científica debido a su capacidad de envejecimiento acelerado y su susceptibilidad a enfermedades relacionadas con el envejecimiento. Se utiliza como modelo para estudiar procesos de envejecimiento, regeneración de tejidos y enfermedades relacionadas con la edad, lo que lo convierte en un organismo de gran interés en la biología y la medicina.
La vejez
La muerte de los peces suele ser el resultado de depredación, accidentes, patógenos oportunistas o mutaciones somáticas acumuladas que conducen a un deterioro lento de la salud y una mayor susceptibilidad a los factores ambientales. Sin embargo, algunos peces envejecen a través del proceso de senescencia de “muerte programada” que es más típico de los mamíferos como nosotros.
La senescencia se refiere a los cambios relacionados con la edad que tienen un efecto adverso en un organismo y que aumentan la probabilidad de su muerte. La senescencia incluye la degradación metabólica y anatómica que ocurre en los animales adultos mayores después de la maduración y reproducción. El salmón del Pacífico proporciona un ejemplo espectacular. Los peces que migran reproductivamente en condiciones físicas óptimas ingresan a su río natal, maduran, desovan, se descomponen anatómica y fisiológicamente y mueren en cuestión de semanas. Muchos de los cambios anatómicos y fisiológicos que ocurren pueden estar relacionados con los efectos combinados de la sobreproducción de esteroides y la inanición. Las células interrenales, que son células productoras de esteroides asociadas con el riñón y son homólogas con la corteza suprarrenal de los mamíferos, secretan corticosteroides, produciendo niveles sanguíneos de estas sustancias cinco o más veces superiores a los niveles normales. Este hiperadrenocorticismo produce cambios degenerativos rápidos en el corazón, hígado, riñón, bazo, timo y arterias coronarias; la última degeneración es sorprendentemente similar a la enfermedad de las arterias coronarias en humanos. El tracto digestivo, incluidas las vellosidades intestinales, se degenera, las reservas de grasa se agotan y cesa la alimentación. Se producen infecciones por hongos y una resistencia reducida a las bacterias, lo que indica una pérdida de la función inmunológica. Un conflicto entre la reproducción y la supervivencia es evidente en el colapso del sistema inmunológico: los corticosteroides elevados aparentemente sirven para acelerar la movilización de la energía almacenada hacia la actividad reproductiva, pero tienen el "efecto secundario" de suprimir la función inmunológica. En los salmones del Pacífico que se reproducen naturalmente, estos efectos secundarios son irreversibles. Los machos y hembras castrados no producen los corticosteroides elevados y no desovan, sino que continúan creciendo al doble de longitud y viven el doble de tiempo que los peces intactos. Los machos precoces, aquellos que maduraron como parr y pasaron por alto las fases de smolt y marina, pueden sobrevivir al desove y reproducirse nuevamente al año siguiente.

Figura 12.4. La reproducción es antagónica con la supervivencia del individuo, pero sin ella, la especie no sobrevive, de allí que algunos biólogos evolutivos sostengan que la unidad evolutiva son los genes que usan a los individuos como máquinas de transporte desechables.
Se produce una senescencia igualmente espectacular en varios otros taxones de peces. La reproducción tanto en lampreas parasitarias como no parasitarias implica la maduración acompañada por el cese de la alimentación y la atrofia de la mayoría de los órganos internos con la excepción del corazón y las gónadas. Las grasas y las proteínas musculares se metabolizan o transforman en productos gonadales. Tanto los machos como las hembras mueren poco después del desove, probablemente por inanición. Las anguilas viven como juveniles durante muchos años en ríos y lagos. Luego se someten a una metamorfosis reproductiva que incluye agrandamiento de los ojos, cambios en la coloración corporal y en las proporciones finas, degeneración intestinal y cese de la alimentación. Después de una migración reproductiva al mar que puede llevarlos miles de kilómetros, presumiblemente todos los adultos mueren. La manipulación de laboratorio de las funciones hormonales indica que, al igual que con los salmones, la senescencia rápida resulta de la elevación de los corticosteroides y la inanición. Durante la maduración, el congrio y la agachadiza también experimentan atrofia intestinal y además pierden los dientes. El Gobio de hielo, Leucosparion petersi, que ingresa al agua dulce para desovar y luego muere, desarrolla glándulas suprarrenales agrandadas y degeneración esplénica. Se ha observado una senescencia más gradual en muchas especies reproductoras múltiples, como arenques, eglefinos, guppies y otros portadores vivos, peces killi anuales y Medaka. Los indicadores anatómicos y fisiológicos de la senescencia gradual incluyen cambios de longitud y peso reducidos o incluso negativos, producción reducida de huevos, enturbiamiento corneal, escamas desordenadas, crecimientos malignos como melanomas, deformidades espinales y capacidad regenerativa alterada. Tales cambios senescentes son más comunes en especies pequeñas de vida corta que maduran a edades relativamente tempranas.

Figura 12.5. El gobio de hielo (Leucosparion petersi) es una especie de pez que habita en los ríos de Siberia, Rusia. Es conocido por su adaptación única a las bajas temperaturas y su capacidad de sobrevivir en aguas heladas. Tiene un cuerpo delgado y alargado, con una coloración plateada y aletas traseras largas y puntiagudas. El gobio de hielo se alimenta de pequeños invertebrados y algas que encuentra en los ríos congelados durante el invierno. Es considerado un indicador de la calidad del agua, ya que su presencia indica la pureza y el estado saludable del ecosistema fluvial. A pesar de su resistencia al frío, el gobio de hielo se encuentra amenazado debido al cambio climático y la alteración de su hábitat natural.
13. Apareamiento y cambio de sexo
13. Apareamiento y cambio de sexo
El éxito evolutivo está determinado por la capacidad de un individuo para colocar genes en las generaciones futuras, en relación con el éxito de los conespecíficos “competidores de la misma especie”. Para transmitir genes, los individuos deben emparejarse con los específicos. Sin embargo, muchas especies de peces son relativamente solitarias como adultos. Durante las temporadas de reproducción, los peces deben superar los hábitos individualistas y buscar posibles parejas de apareamiento. Los hábitats y sustratos de reproducción adecuados deben encontrarse o incluso modificarse en nidos, las actividades de machos y hembras deben estar sincronizadas y deben emplearse tácticas para evitar la hibridación (mecanismos de aislamiento de especies). Las agregaciones de individuos reproductivamente activos crean una competencia potencial para los sitios de desove y apareamiento, provocando un comportamiento territorial y de elección de pareja. El cortejo y el desove pueden distraer la atención de los participantes y hacerlos más vulnerables a los depredadores. Después del desove, muchas especies participan en diversos grados de cuidado parental. Todas estas actividades y características constituyen los diversos sistemas de apareamiento de los peces. Nuestro énfasis aquí será en la diversidad de los sistemas de apareamiento, con un enfoque en los patrones y adaptaciones donde se puedan identificar.
Apareamiento
Los componentes importantes de un sistema de reproducción incluyen la frecuencia de apareamiento, el número de parejas y el rol de género de los individuos. Los peces muestran una mayor diversidad en estos rasgos que los otros vertebrados. La mayoría de los peces siguen la norma de los mamíferos y las aves de permanecer de un género a lo largo de la vida adulta, pero muchas especies de peces cambian de sexo y algunas son partenogenéticas, produciendo crías de huevos no fertilizados. Algunos peces conservan un solo compañero, tal vez de por vida, otros se aparean promiscuamente, y algunos son harémicos. Y en algunos peces, un solo sistema de reproducción puede no caracterizar a toda la especie.
La mayoría de los peces son iteroparos, que engendran más de una vez durante su vida. Sin embargo, algunas especies conocidas son semielparas, desovan una vez y mueren. La semelparidad caracteriza a la mayoría de los salmones del género Oncorhynchus spp. Estos peces eclosionan en agua dulce, migran al mar por un período de 1 a 4 años, y luego regresan a su corriente natal (nacimiento) donde desovan y mueren. Aunque el ciclo de vida de las hembras parece ser relativamente fijo a lo largo de una especie, las diferencias intra-poblacionales en los ciclos masculinos son comunes.
Con la excepción de peces anuales como Riquulinas aplocheiloides, los peces semélparos son diádromos o incluyen al menos una fase migratoria importante en su ciclo de vida. Las anguilas muestran las diferencias de tácticas basadas en el sexo dentro de una estrategia semelparosa general. Los machos a menudo maduran rápidamente (c. 3–6 años) y en un tamaño uniformemente pequeño (30–45 cm) independientemente del lugar, mientras que las hembras son consistentemente más largas (35–100 cm) y pueden madurar rápidamente (4–13 años) en latitudes bajas o lentamente (6–43 años) en latitudes altas. Las hembras de maduración lenta crecen y producen más huevos que las hembras más pequeñas y de maduración más rápida. En las anguilas americanas, los machos tienen una distribución geográfica relativamente restringida, que se presenta principalmente en los estuarios del sureste de los Estados Unidos, mientras que las hembras se encuentran en todo el rango de especies y en todos los hábitats de América del Norte. Por lo que se sabe, todos los miembros de una especie de anguilida migran a la misma región oceánica para desovar y morir (el mar de Sargasso en el Atlántico occidental para las anguilas americanas y europeas, el mar de Filipinas para las anguilas japonesas)
Tipos de apareamiento
Los sistemas de apareamiento se definen por el número de parejas de apareamiento que un individuo tiene durante una temporada de reproducción. Las tres categorías más comunes son promiscuas, polígamas y monógamas. Los criadores promiscuos son aquellos en los que hay poca o ninguna elección de pareja obvia, donde tanto los machos como las hembras engendran con múltiples parejas, ya sea en un momento o en un período corto.

Figura 13.1. El pez anémona o pez payaso, perteneciente a la familia Pomacentridae, es un grupo diverso de peces marinos que se encuentran en los arrecifes de coral de todo el mundo. Estos peces son conocidos por su relación simbiótica con las anémonas marinas, de las cuales obtienen protección y refugio, mientras que las anémonas se benefician de la limpieza y la dispersión de nutrientes que proporcionan los peces. Los peces anémona exhiben una amplia gama de colores y patrones, y suelen ser territoriales y agresivos para defender su anémona de otros peces. Son importantes para el ecosistema de arrecifes de coral, ya que ayudan a mantener el equilibrio y la salud del coral y otros organismos marinos en la comunidad del arrecife.
La poligamia, donde solo un sexo tiene múltiples parejas, toma múltiples formas. La poliandria, donde una hembra se aparea con varios machos (y presumiblemente no al revés), es relativamente infrecuente, hasta ahora documentada solo en un pez anémona (Pomacentridae spp.) (Moyer y Sawyers, 1973). La poliandria también podría ser descriptiva de los peces rape hembra ceratioides que tienen más de un macho adjunto. La poliginia es la forma más común, involucrando a los machos como el sexo polígamo. Los machos territoriales que cuidan huevos y crías son visitados frecuentemente por varias hembras, como en esculturas, peces solares, dardos, damiselas y cíclidos. La poliginia también puede convertirse en formación de harén, donde un macho tiene derechos de reproducción exclusivos de varias hembras que puede proteger. Los harenes se han observado en numerosos cíclidos y en varias familias de arrecifes de coral como en los peces cirujano.
Muchos animales poligínicos forman leks, que son áreas tradicionales donde varios machos se congregan con el único propósito de exhibir a las hembras. Las hembras a menudo se sienten atraídas por un macho en respuesta a su posición central dentro del terreno de lekking, o al vigor de algún carácter sexual secundario que genera desventaja adaptativa “efecto cola de pavo real”. El lekking es común en aves y mamíferos, en los que solo la hembra proporciona cuidado parental. Algunos cíclidos africanos están más cerca de formar verdaderos leks. Un gran número (aproximadamente 50,000) de Cyrtocara eucinostomus macho se congregan en una plataforma poco profunda de 4 km en el lago Malawi y construyen nidos de arena y exhiben a las hembras que pasan cada mañana. Las hembras desovan y luego echan los huevos en otros lugares. Las agregaciones masculinas se rompen cada tarde, cuando los peces se alimentan.

Figura 13.2. Cyrtocara eucinostomus, también conocido como Haplochromis jumbo o pez papagayo de Malawi, es una especie de cíclido endémico del lago Malawi en África Oriental. Este pez es reconocido por su tamaño impresionante y su forma corporal alargada y comprimida lateralmente. Los machos adultos exhiben colores vibrantes, como el azul, amarillo y negro, mientras que las hembras son más discretas. Son peces herbívoros que se alimentan de algas y pequeños invertebrados encontrados en el lecho del lago. El Cyrtocara eucinostomus es muy valorado en la acuariofilia debido a su apariencia llamativa y su comportamiento interesante. Sin embargo, es importante destacar que esta especie requiere un hábitat adecuado y condiciones de agua óptimas para su cuidado en cautiverio.
En los sistemas monógamos, los peces viven en parejas que permanecen juntos y se aparean, o se aparean con el mismo individuo de forma repetida y exclusiva. Las especies que se emparejan fuertemente incluyen los peces gato de agua dulce de América del Norte, muchos peces mariposa y pez ángel, la mayoría de los protectores de sustratos y algunos cíclidos que incuban en la boca y los peces anémona; en los peces mariposa, las parejas pueden permanecer juntas durante varios años y probablemente se apareen de por vida. Los peces de arrecifes de coral monógamos comúnmente engendran con la misma pareja diariamente durante un período prolongado sin el cuidado de los jóvenes, mientras que las especies de agua dulce como los cíclidos se reproducen durante un tiempo limitado y luego ambos padres cuidan de los jóvenes. La monogamia ha evolucionado de manera independiente en muchos grupos, a menudo en conjunción con la territorialidad y el cuidado paterno.
Roles de género en los peces
Aunque la gran mayoría de los peces son gonocoristas, con el sexo determinado a una edad temprana y permaneciendo fijos como machos o hembras, un número significativo de peces puede funcionar como machos o hembras de forma simultánea o secuencial. Los correlatos ambientales y las causas evolutivas del cambio de sexo en los peces han sido objeto de considerable estudio y especulación.
La inversión sexual ha evolucionado, aparentemente de forma independiente, en quizás 34 familias pertenecientes a 10 órdenes, incluidas las anguilas (Anguilliformes), las loaches (Cypriniformes), los peces light (Stomiiformes), los killifishes (Atheriniformes), las anguilas del pantano (Synbranchiformes), las flatheads (Scorpaeniformes), pez cofre (Tetraodontiformes), y al menos 24 familias perciformes (incluidos los snooks, las lubinas, los azulejos, los emperadores, los rovers, las percas, las aletas, los pezones, los anguilas, los paracaídas, los pargos, los loros y los gobies). Los individuos que cambian de sexo pueden ser:
(a) hermafroditas simultáneas, capaces de liberar óvulos o espermatozoides viables durante el mismo desove; o
(b) hermafroditas secuenciales, que funcionan como machos durante una fase de la vida, y como hembras durante la otra. Entre los hermafroditas secuenciales, los peces protandrosos se desarrollan primero como machos y luego cambian a hembras, mientras que los peces protoginos maduran primero como hembras y luego se convierten en machos. Existen variaciones en estos patrones, como las poblaciones protóginas con algunos machos que se desarrollan directamente a partir de juveniles, o hermafroditas simultáneos que pierden la capacidad de funcionar como un solo sexo. La protoginia es, con mucho, la forma más común de hermafroditismo, que se exhibe en al menos 17 familias marinas tropicales, que es aproximadamente una quinta parte de las familias de arrecifes
Un grupo adicional de peces se aparta de los roles de género gonocorísticos normales. Los vivípados en México y Texas incluyen "especies" partenogenéticas que son todas hembras pero que requieren el esperma de machos de otras especies para activar la división celular en sus óvulos. La partenogénesis en víviparos toma dos formas: ginogénesis e hibrogénesis. Las hembras ginogenéticas suelen ser triploides y producen huevos que también son 3N. Estos huevos son activados por el esperma de otras especies, pero no se incorpora material de esperma; Por lo tanto, las hijas son genéticamente idénticas a sus madres. En cambio, las hembras hibrogenéticas son diploides y producen huevos haploides que, durante la reducción de la división de la meiosis, mantienen los genes maternos y descartan los genes paternos. Al aparearse, estos huevos se unen con el esperma de machos de otra especie, formando una nueva hija híbrida diploide (no se producen hijos). Cuando la hija se aparea, vuelve a producir huevos que son haploides y "hembras". Por lo tanto, el linaje materno se conserva y la contribución genética del macho se pierde después de una generación. Se cree que estas "especies" partenogenéticas surgieron originalmente como híbridos entre hembras de Poeciliopsis monacha y machos de cuatro congéneres, P. lucida, P. occidentalis, P. latidens y P. viriosa. Los machos de las cuatro especies son los donantes de esperma habituales durante el apareamiento.

Figura 13.3. El guatopote Jarocho (Poeciliopsis gracilis) es una especie de pez de agua dulce que se encuentra en las regiones costeras del Golfo de México, especialmente en Veracruz, México. Este pez es apreciado tanto en la acuariofilia como en la pesca deportiva debido a su belleza y comportamiento interesante. Los guatopotes Jarocho tienen un cuerpo delgado y alargado, con colores vibrantes que van desde el amarillo hasta el naranja y el rojo. Son peces omnívoros que se alimentan de pequeños invertebrados, algas y materia vegetal. Son especies vivíparas, lo que significa que dan a luz a crías vivas en lugar de poner huevos. Estos peces son resistentes y se adaptan bien a una variedad de condiciones de agua. Su presencia en los ecosistemas acuáticos es importante para mantener el equilibrio y la diversidad de especies.
14. Caracteres sexuales secundarios y selección sexual
14. Caracteres sexuales secundarios y selección sexual
Algunos rasgos de un animal funcionan principalmente para atraer parejas o para ayudar en batallas entre miembros de un sexo para tener acceso al otro sexo. Tales rasgos seleccionados sexualmente confieren una ventaja de apareamiento a un individuo; son un subconjunto de la selección natural, que generalmente involucra rasgos que confieren una ventaja de supervivencia. Los rasgos seleccionados sexualmente no tienen otro propósito que el apareamiento e incluso pueden perjudicar al poseedor con respecto a otras actividades que influyen en la aptitud física. Sin embargo, los rasgos seleccionados sexualmente también pueden conferir una ventaja de supervivencia positiva, como el gran tamaño en los machos, que proporciona una defensa física contra los depredadores y también es favorecido por las hembras durante el apareamiento. Los rasgos seleccionados sexualmente a menudo se conocen como características sexuales secundarias.

Figura 14.1. En especies donde los machos no cuidan de las crías, se tiene la tendencia al desarrollo de caracteres poco ventajosos, pues las hembras se enfocan en características que indican una buena salud.
Las características primarias incluyen ovipositores, genitales y otras estructuras de copulación, como claspers en elasmobranquios, gonopodios o priapios en peves vivíparos y faloestetidos, o parches o bolsas de cría y otras estructuras utilizadas en el cuidado parental. En algunos casos, un carácter puede ser secundario y primario, sirviendo tanto en la atracción de pareja como en la cópula o el cuidado de los padres. En los espinosos “Gasterosteiformes”, los espinosos masculinos son atraídos por las hembras con vientres hinchados, pero la hinchazón se debe a los ovarios maduros de la hembra.
Dimorfismo sexual
Las características sexuales secundarias tienen cuatro atributos generales:
(a) están restringidas o se expresan de manera diferente en un sexo (generalmente el macho),
(b) no aparecen hasta la maduración,
(c) a menudo se desarrollan durante una temporada de reproducción y luego regresan,
(d) y generalmente no mejoran la supervivencia.
Las características secundarias toman la forma de dimorfismos sexuales (diferencias en las partes del cuerpo entre los sexos), como las diferencias en el tamaño corporal, la forma de la cabeza, la forma de la aleta, la dentición y la ornamentación corporal, o como dicromatismos (diferencias en la coloración).

Figura 14.2. Dimorfismo sexual en Etheostoma caeruleum. También conocido como darda azul o pez arco iris de Carolina, es una especie de pez nativo de América del Norte. Estas coloridas criaturas habitan en arroyos y ríos de agua dulce en las regiones del este de Estados Unidos. El darda azul destaca por su belleza, con un cuerpo alargado y estilizado adornado con tonos azules iridiscentes que se intensifican durante la temporada de reproducción. Son peces de tamaño pequeño, alcanzando alrededor de 5 a 8 centímetros de longitud. Estos peces son conocidos por su comportamiento territorial y su dieta principalmente compuesta de pequeños invertebrados acuáticos, como insectos y crustáceos. La conservación de Etheostoma caeruleum es crucial para preservar la diversidad de peces nativos en los ecosistemas acuáticos. Su presencia no solo enriquece la biodiversidad, sino que también indica la salud y la calidad del agua en la que habitan.
Sabemos comparativamente poco acerca de las diferencias eléctricas, químicas y acústicas entre los sexos, aunque las diferencias en anatomía y fisiología asociadas con estos modos sensoriales son comunes (por ejemplo, peces elefante, salmones, damas, gobios). Por ejemplo, los machos de Porichthys notatus (Batrachoididae), atraen a las hembras por un "zumbido", un sonido producido al contraer grandes músculos unidos a las paredes de la vejiga natatoria. La capacidad de producir sonido y su importancia en el cortejo se reflejan en numerosas diferencias entre los sexos. Los machos difieren de las hembras en cuanto a que tienen un tamaño corporal más grande, color diferente, músculos sónicos más grandes y diferentes circuitos neuronales, que involucran cuerpos celulares más grandes, dendritas y axones en el cerebro, que los que se encuentran en las hembras.

Figura 14.3. El sapo cabezón o sapo de aleta lucia (Porichthys notatus) es una especie de pez venenoso que se encuentra en las aguas costeras del Pacífico de América del Norte. Es conocido por su aspecto peculiar, con una cabeza grande y redonda, y una aleta dorsal prominente que se extiende a lo largo de su cuerpo. Este pez tiene una coloración marrón o grisácea con manchas y rayas oscuras, lo que le proporciona un excelente camuflaje en el lecho marino. El sapo cabezón es nocturno y se alimenta principalmente de crustáceos y pequeños peces. A pesar de su apariencia inofensiva, los machos de esta especie poseen glándulas venenosas en sus aletas pectorales, las cuales pueden liberar una toxina defensiva en caso de amenaza. Estos peces son de interés científico debido a sus características únicas y su capacidad para producir sonidos utilizando músculos especializados.
Mientras que la gran mayoría de las especies de peces no muestran dimorfismos sexuales obvios, muchas especies son claramente dimorfas. Los machos son a menudo el sexo más grande como en el salmón, mientras que las hembras son más grandes en los tiburones caballa y ballena, el esturión, las anguilas verdaderas, los espinos, entre otros. El pez delfín macho (Coryphaena hippurus, Coryphaenidae) es más grande y tiene una cabeza claramente más roma que las hembras. Durante las migraciones de desove, las anguilas masculinas desarrollan ojos más grandes que las hembras, y el salmón macho desarrolla mandíbulas superiores e inferiores claramente cóncavas, llamadas kype, que impiden la alimentación.

Figura 14.4. Dimorfismo sexual en el salmón. El dimorfismo sexual en los salmones se refiere a las diferencias físicas distintivas entre los machos y las hembras de la especie. En la mayoría de las especies de salmones, los machos desarrollan características más llamativas durante la época de reproducción. Estas incluyen un cambio en la coloración del cuerpo, con tonos más vibrantes y manchas más pronunciadas, así como el crecimiento de una mandíbula superior alargada llamada quijada o gancho. Además, los machos pueden presentar una joroba en la espalda y dientes más grandes. Por otro lado, las hembras mantienen una apariencia más discreta y conservan su coloración original. Estas diferencias sexuales se relacionan con la competencia por aparearse y el cortejo entre los individuos de la especie.
Los machos pueden tener filamentos de cola en los extremos de sus aletas dorsal, anal o caudal; aletas medianas o pareadas agrandadas; aletas de la cola alargadas; o tener cubiertas de branquias o aletas pélvicas alargadas con lóbulos que se muestran frente a las hembras durante el cortejo. En los elasmobranquios, los machos pueden tener dientes más largos o más afilados que las hembras, que sirven para agarrar a la hembra durante el cortejo y la cópula; Las hembras a su vez pueden tener una piel más gruesa que los machos.
Las diferencias en la ornamentación corporal incluyen pequeñas protuberancias en el cuerpo, escamas y aletas de peces mayormente machos en 25 familias diferentes de teleósteos. Estas protuberancias se denominan tubérculos nupciales o de reproducción cuando son de origen epidérmico, o órganos de contacto cuando son de origen dérmico. Las diferencias dérmicas también incluyen el tipo de escama: los ciprodontidos masculinos tienen escamas ctenoides, mientras que las hembras tienen escamas cicloides. Otros adornos dimórficos incluyen pequeños ganchos en las aletas anales, y papilas rostrales en peces cueva machos ciegos, patrones de fotóforos en peces linterna y maniquíes de huevo pigmentados en las aletas anales de los cíclidos. Algunos peces pequeños machos, cíclidos, y peces loro desarrollan jorobas óseas o grasas en la parte frontal (región nucal) de su cabeza.
Consecuencias de la contaminación en la selección sexual
La importancia de los rasgos dimórficos seleccionados sexualmente como mecanismos de aislamiento de las especies se ha vuelto cada vez más evidente en los últimos años a medida que las alteraciones del paisaje causadas por el hombre interfieren con los patrones de apareamiento. En el lago Victoria, donde "probablemente más especies de vertebrados estén en riesgo inminente de extinción. . . que en cualquier otra parte del mundo ", los cíclidos están separados taxonómicamente y aparentemente de manera conductual por la variación de color en los machos. Las hembras discriminan entre los pretendientes varones estrechamente relacionados, al menos en parte, por las diferencias de color.
El aumento en el crecimiento de algas, como resultado de la gran escorrentía de nutrientes de las laderas deforestadas, ha aumentado la turbidez y la transmisión de luz está muy limitada, especialmente en los extremos largo (rojo) y corto (azul) de la luz visible espectro. La selección de pareja aparentemente se ve afectada por esta absorción de longitudes de onda largas y cortas porque los distintos colores rojo y azul en los machos ya no son obvios, lo que lleva a una percepción alterada de los colores de apareamiento y una mayor hibridació. “Cuando la eutrofización apaga las luces de un cuerpo de agua, la diversidad ecológica y de especies se erosiona rápidamente”.

Figura 14.5. Dimorfismo sexual extremo en un macho, Nocomis leptocephalus. Este construye nidos de guijarros en el fondo de los arroyos y atrae a las hembras con su cabeza hinchada y sus distintos tubérculos reproductores. Nocomis leptocephalus, conocido como matalote espinoso, es una especie de pez de agua dulce que se encuentra en América del Norte. Este pez posee un cuerpo alargado y delgado, con una coloración plateada y tonos azulados en su dorso. El matalote espinoso es reconocido por sus aletas pectorales largas y espinosas, las cuales utilizan para protegerse y defenderse de posibles depredadores. Habita en arroyos y ríos de corriente rápida, prefiriendo aguas frescas y limpias. Su alimentación se basa principalmente en invertebrados acuáticos, como insectos y crustáceos. Esta especie desempeña un papel importante en los ecosistemas acuáticos, ya que ayuda a controlar las poblaciones de estos organismos. La conservación del matalote espinoso es esencial para preservar la diversidad y la salud de los ecosistemas acuáticos en los que habita. Su presencia es un indicador de la calidad del agua y la salud de los hábitats acuáticos.
Otros investigadores han relacionado las alteraciones y las reducciones en el comportamiento del desove con el aumento de la turbidez. La selección sexual se relajó en condiciones de turbidez inducida experimentalmente en el Gobio de arena, Pomatoschistus minutus. En el agua clara, las hembras mostraron una fuerte preferencia por los machos más grandes, pero esta preferencia se debilitó a medida que se reducía la claridad del agua. Por lo tanto, el aumento de la turbidez puede alterar los sistemas de apareamiento al alterar la intensidad de la selección sexual. Otros hallazgos relacionados incluyen una reducción en la intensidad de la coloración roja en los espinosos de tres espaldas masculinas en el Mar Báltico, vinculados al crecimiento de algas causado por la eutrofización y la visibilidad reducida. La frecuencia de desove disminuida, el tiempo interrumpido de desove y una reducción del 93% en el número de huevos viables producidos por Cyprinella trichroistia, los cuales se interpretaron como un resultado de las señales visuales interrumpidas asociadas con el aumento de la turbidez y los cambios resultantes en la transmisión de luz.
La comunicación visual no es el único modo afectado por los impactos antropogénicos. El reconocimiento de especies en la cola de espada, Xiphophorus birchmanni, está mediado químicamente, y las hembras prefieren machos conespecíficos basados en señales químicas. Los peces probados en agua sin contaminar mantienen esa preferencia de especie. Las hembras sometidas a efluentes de aguas residuales y escorrentía agrícola con agroquímicos no discriminaron entre machos de X. malinche con los que pueden hibridar. El químico con mayor probabilidad de interferir con la comunicación se identificó como ácido húmico, "un producto ubicuo y natural elevado a niveles altos por procesos antropogénicos". Parece razonable suponer que otras modalidades sensoriales involucradas en el cortejo y el reconocimiento de especies, como la comunicación acústica y eléctrica, también podrían verse afectadas por actividades antropogénicas.
Anidación
Muchos peces desovan en nidos. Los nidos no pueden ser más elaborados que una simple depresión que uno o ambos sexos excavan en el fondo al abanicar vigorosamente con las aletas o nadar rápidamente en su lugar. En las lampreas y el salmón, los huevos se cubren con arena o grava adicional y luego se abandonan, mientras que en otras especies el macho permanece para proteger los huevos expuestos. Muchas especies desovan en una grieta o espacio de roca en o debajo de una concha o roca o en un tronco ahuecado que ha sido excavado o liberado de crecimiento y escombros. Algunos damiseos limpian la superficie de la arena con una salpicadura de arena y luego la abanican con sus aletas. La banda europea, Crenilabrus melops, recubre una grieta de roca con diferentes tipos de algas. Las algas más suaves en la parte posterior de la grieta sirven como sustrato para el desove, mientras que las algas coralinas más duras y pegajosas se empaquetan en la parte exterior de la grieta para proteger los huevos.

Figura 14.6. La banda europea (Crenilabrus melops) es una especie de pez costero que se encuentra en las aguas del Atlántico Este, desde Noruega hasta Marruecos. Este pez presenta un dimorfismo sexual marcado. Los machos tienen un colorido y patrón distintivo, con un cuerpo predominantemente azul y una franja naranja que recorre horizontalmente desde la cabeza hasta la base de la aleta caudal. También desarrollan una protuberancia en la frente durante la época de reproducción. Por otro lado, las hembras son de color marrón o verde oliva, con manchas más sutiles. Estos peces son territoriales y se alimentan principalmente de pequeños crustáceos y moluscos. La banda europea es valorada tanto en acuarios marinos como en la pesca deportiva debido a su belleza y comportamiento activo.
Las madrigueras se excavan y luego se protegen. Los machos de algunos pequeños peces construyen montículos de nidos acumulando hasta 14,500 piedras pequeñas, de 6 a 12 mm de diámetro, que se transportan desde la boca hasta el sitio del nido desde una distancia de hasta 4 m (p. Ej., Nocomis, Semotilus, Exoglossum).
Los huevos depositados por la hembra caen en los intersticios entre las piedras y están cubiertos por piedras adicionales; el nido está protegido y mantenido libre de sedimentos por el macho. En los cíclidos africanos, los machos construyen nidos en arena, algunos de diseño elaborado. Estas estructuras han sido comparadas con las bandadas de aves de corral porque son principalmente estaciones de exhibición, cortejo y desove desde las cuales la hembra recoge los huevos en su boca casi tan rápido como se ponen.
Los nidos también pueden construirse con materiales producidos intrínsecamente, a veces combinados con objetos recolectados extrínsecamente. Los gouramis y los peces luchadores siameses (Anabantoidei) y los lucios caracinos producen nidos de burbujas o espuma, que consisten en burbujas cubiertas de moco que se unen en una masa. Algunos anabantoides agregan fragmentos de plantas, detritos, arena e incluso partículas fecales a la masa de la burbuja. Las espinillas macho pasan una mucoide, una sustancia similar a un hilo que se fabrica en los riñones a través de la cloaca, y la utiliza para unir trozos de hojas, pastos y filamentos de algas que crean un nido en forma de taza o túnel en el que una hembra deposita sus huevos. Todas estas estructuras son tendidas por el macho después del desove.
Cortejo
El cortejo es la serie de acciones de comportamiento realizadas por uno o ambos miembros de una pareja de apareamiento justo antes del desove. El cortejo tiene varias funciones que maximizan la eficiencia del acto de desove. El cortejo ayuda en el reconocimiento de especies (un mecanismo de aislamiento de especies antes del apareamiento), la unión por pares, la orientación al sitio de desove y la sincronización de la liberación de gametos. El cortejo a menudo es necesario para superar la agresión territorial del macho, que de otro modo podría alejar a la hembra del sitio (en muchas especies, los machos ya tienen huevos en sus territorios y deben protegerse contra la depredación por parte de individuos de ambos sexos). El cortejo puede ser relativamente simple (como en el arenque), o puede implicar un gran número o una progresión compleja de pantallas y señales de uno o ambos sexos.

Figura 14.7. Igual que otras características como la metamorfosis, la anidación se encuentra de manera mucho más diversa en los peces.
Durante el cortejo, los individuos cambian frecuentemente el color de sus patrones normales de camuflaje, a patrones de colores más contrastados y brillantes. En muchas especies (por ejemplo, peces pequeños, plateados, cíclidos), la coloración del cuerpo existente se intensifica o la cabeza se vuelve oscura en relación con el resto del cuerpo. La producción de sonido durante el cortejo, usualmente por el macho, ocurre en muchas familias de peces, a menudo acompañados de presentaciones visuales. Patrones de natación exagerados o rápidos, erección de aletas y saltos fuera del agua. Los estimulantes químicos también están involucrados. El pez dorado macho, Danios cebra (Cyprinidae) y los gobios inician las actividades de cortejo cuando se exponen al agua que contenía a una hembra gravida, y los gobios femeninos grávidos son atraídos por sustancias andróginas producidas por el macho. Las especies y el reconocimiento sexual durante el cortejo en cíclidos ocurren más rápidamente cuando los individuos reciben señales visuales y químicas de parejas potenciales.

Figura 14.8. El pez dorado Danios cebra es una especie popular en la acuariofilia debido a su belleza y su fácil cuidado. Originario del sudeste de Asia, este pez presenta un cuerpo alargado y delgado, con una coloración llamativa de rayas horizontales que alternan entre dorado y plateado. Los Danios cebra son peces activos y sociales que se mueven rápidamente por el acuario. Son peces de cardumen, por lo que se recomienda mantenerlos en grupos para promover su bienestar. Son omnívoros y se alimentan de una variedad de alimentos, incluyendo escamas, larvas de insectos y pequeños crustáceos. Además de su atractivo estético, los Danios cebra también son conocidos por su resistencia y adaptabilidad, lo que los convierte en una opción popular para los aficionados a los acuarios.
15. Desove y cuidado parental
15. Desove y cuidado parental
El desove y el cuidado parental son aspectos fascinantes y vitales en la vida de los peces. A lo largo de la evolución, los peces han desarrollado una variedad de estrategias reproductivas para asegurar la supervivencia de sus crías. Desde el desove en masa en aguas abiertas hasta el cuidado intensivo de los huevos y las crías por parte de los padres, este comportamiento reproductivo ofrece una amplia gama de adaptaciones y comportamientos interesantes. En este artículo, exploraremos 15 ejemplos destacados de desove y cuidado parental en peces, examinando las diferentes estrategias, los factores ambientales que influyen en ellas y su importancia para la ecología y la evolución de estas especies acuáticas. A través de esta exploración, obtendremos una mayor comprensión de los complejos y sorprendentes comportamientos reproductivos de los peces.
Desove
El acto final de desove puede tener lugar en la columna de agua, sobre el fondo, en contacto con plantas y rocas, y en algunos casos especiales, fuera del agua. En los fertilizantes externos, los comportamientos asociados con el desove a menudo implican nadar rápidamente, temblar, vibrar, extender las aletas y envolver a la hembra con las aletas o el cuerpo del macho. Los tubérculos reproductores y los órganos de contacto comunes en muchos peces pueden ayudar a mantener el contacto entre los miembros de una pareja y también pueden estimular a la hembra. Las especies de fertilización interna también participan en secuencias elaboradas de cortejo. Los peces millón masculinos realizan una variedad de acciones que incluyen seguir, atraer, morder y nadar sigmoides para mostrar sus aletas y la coloración del cuerpo hasta que una hembra les permita acercarse y copular. La secuencia y los tipos de exhibiciones por parte del macho sirven como un mecanismo de aislamiento de especies en el sentido de que las hembras rechazan a los machos de la especie equivocada después de ver sus exhibiciones de cortejo.
También pueden producirse sonidos específicos de la especie durante el acto de desove. Por ejemplo, en las especies hermafroditas, el "macho" emite una llamada de cortejo y la "hembra" una llamada de desove. A medida que los individuos cambian de rol durante un desove prolongado, también cambian los sonidos que producen.
En grupo
Aunque la gran mayoría de los peces desovan como parte de grupos grandes, es común el emparejamiento de machos y hembras individuales dentro de estos grupos. La formación de parejas a corto plazo probablemente asegure la liberación eficiente de gametos y la fertilización; la liberación aleatoria de gametos podría hacer que una gran proporción de óvulos no sean fertilizados porque los espermatozoides se vuelven inviables y se diluyen rápidamente en aguas abiertas, y los óvulos se vuelven no fertilizables unos minutos después de la liberación. El desove grupal, que involucra a más de dos peces, generalmente involucra a una hembra acompañada por varios machos.
Engendrando en la superficie
Los engendradores de columna de agua en los arrecifes de coral a menudo se precipitan rápidamente hacia arriba y liberan sus gametos en la cima, a veces cerca de la superficie. Las velocidades se aproximan a 40 km / h en el pez loro rayado, Scarus croicensis. Este patrón se ha observado en más de 50 especies en más de 18 familias. Sus funciones son debatidas. El movimiento hacia arriba en la columna de agua coloca a los huevos fuera del alcance de muchos zooplanktivoros invertebrados y vertebrados bentónicos o casi bentónicos y en corrientes que promueven la dispersión. Sin embargo, al alejarse del arrecife, los adultos reproductores se enfrentan a la amenaza conflictiva de sus propios depredadores, como aves costeras. No es sorprendente que los reproductores bien defendidos se muevan más alto en el agua o desoven más lentamente que las especies más pequeñas y vulnerables. Para muchas especies, la carrera de desove puede servir como un evento final de sincronización en la secuencia de cortejo y también puede ayudar a evadir los depredadores que abundan cerca del arrecife.

Figura 15.1. El pez loro rayado (Scarus croicensis) es una especie colorida y vibrante que se encuentra en los arrecifes de coral del Caribe. Este pez herbívoro es conocido por su característica mandíbula protusible en forma de pico, que utiliza para arrancar y triturar algas y corales. Presenta un cuerpo alargado y comprimido lateralmente, con un patrón de rayas azules y amarillas distintivo que recorre su cuerpo. El pez loro rayado desempeña un papel importante en el ecosistema marino al mantener el equilibrio y la salud de los arrecifes de coral al consumir algas que podrían sofocarlos. Su comportamiento y su apariencia llamativa lo convierten en una especie popular entre los buceadores y los amantes del acuario marino.
Engendrando en el lecho
(YouTube) Es menos probable que los peces engendradores de sustrato formen grupos grandes que los reproductores de columnas de agua; también liberan menos huevos en cada desove. Los machos generalmente establecen territorios sobre el sustrato de desove apropiado, ahuyentan a los machos intrusos y atrayendo a las hembras apropiadas. Las hembras entran en el territorio y depositan uno o unos pocos huevos adhesivos, mientras que el macho pliega su cuerpo o aletas a su alrededor y la presiona contra el sustrato. La liberación de espermatozoides ocurre casi de inmediato, de nuevo en parte porque los depredadores están siempre cerca. La paternidad se puede asegurar, desde el punto de vista masculino, si las hembras facilitan la fertilización al llevar el esperma a la boca, como ocurre en muchos cíclidos.
El acto de desove reúne peces que normalmente pueden ser solitarios, territoriales o extremadamente sensibles a los depredadores, a menudo en lugares que rara vez frecuentan, dado que están infestados de depredadores. Las agregaciones de desove son especialmente comunes entre las especies de arrecifes de coral, involucradas por al menos 164 especies en 26 familias. Como un posible mecanismo para ayudar a superar comportamientos que serían contraproducentes en este momento crítico en la vida de un pez, muchos peces exhiben estupor de desove. Cuando se encuentran en una agregación o modo de desove, las especies que normalmente son difíciles de aproximar o que son muy activas, en cambio se mueven lentamente, en un estado casi similar al de un trance hipnótico. Toman poca o ninguna acción evasiva cuando son abordados por depredadores o buzos. El estupor de desove se ha observado en peces pequeños, retoños, salmonetes, silversides, lubinas, pompanos y pargos.
Cuidado parental
(YouTube) El cuidado de los padres es sorprendentemente común, generalizado y diverso en peces. Aunque la mayoría de las especies dispersan o abandonan los huevos después de la fertilización, aproximadamente 90 de los c. 460 familias de peces óseos incluyen especies que participan en algún tipo de defensa o manipulación de huevos y crías. El cuidado de los padres es más necesario para los huevos demersales o adhesivos que pueden encontrar los depredadores en la parte inferior o entre las plantas u otras estructuras. Por lo tanto, no es sorprendente que pocas especies, si las hay, con huevos pelágicos y flotantes se ocupen de ellos. El cuidado parental incluye:
Construcción y mantenimiento de un nido
La construcción y mantenimiento de nidos en peces es un comportamiento fascinante que se observa en diversas especies acuáticas. Aunque asociamos comúnmente los nidos con las aves, algunos peces también demuestran habilidades notables en la creación y cuidado de sus lugares de reproducción.
La construcción de nidos en peces varía según la especie y el entorno en el que habitan. Algunos peces, como los bagres, construyen nidos en forma de hoyos cavados en el lecho del río o lago. Utilizan sus cuerpos y aletas para desplazar el sedimento y crear un espacio protegido donde depositar los huevos.
Otros peces, como los cíclidos, construyen estructuras más elaboradas. Excavan hoyos o montículos de arena y grava para crear nidos en los que los huevos son depositados y protegidos por los padres. Algunas especies incluso utilizan su boca para transportar materiales y dar forma al nido.
Una vez construido el nido, muchos peces exhiben comportamientos de mantenimiento. Esto implica la limpieza del nido, la protección de los huevos o alevines de posibles depredadores y la oxigenación del agua circundante mediante el batido de las aletas.
El mantenimiento del nido también puede implicar la defensa activa contra otros peces intrusos o depredadores que puedan amenazar los huevos. Los peces padres pueden exhibir comportamientos agresivos, como cargas y mordiscos, para proteger su progenie.
Algunas especies de peces también cuidan de los huevos ventilándolos o agitando sus aletas para asegurar una adecuada oxigenación. Esto es especialmente importante en nidos ubicados en áreas de baja circulación de agua.
El éxito reproductivo de los peces está estrechamente relacionado con la construcción y el mantenimiento efectivo de los nidos. Un nido bien construido y mantenido proporciona un entorno seguro para el desarrollo de los huevos y los alevines, reduciendo la exposición a depredadores y condiciones adversas.
Es interesante destacar que algunos peces también muestran comportamientos cooperativos en la construcción de nidos. Pueden trabajar en equipo para construir estructuras más grandes y complejas, lo que aumenta la protección y supervivencia de los huevos.
La construcción y mantenimiento de nidos en peces es un ejemplo fascinante de comportamiento instintivo y adaptativo en el reino acuático. Estos comportamientos son fundamentales para la reproducción exitosa de las especies y juegan un papel crucial en la conservación y diversidad de los ecosistemas acuáticos.
Perseguir a los potenciales depredadores de huevos
Perseguir a los potenciales depredadores de huevos es un comportamiento defensivo observado en algunos peces para proteger sus huevos y asegurar la supervivencia de su descendencia. Aunque los peces no pueden perseguir activamente a los depredadores como lo harían las aves o mamíferos terrestres, emplean estrategias adaptativas para evitar que sus huevos sean consumidos.
Una de las estrategias comunes es la elección de sitios de desove bien protegidos. Los peces seleccionan áreas con vegetación densa, grietas en las rocas o estructuras submarinas donde pueden depositar sus huevos lejos de los depredadores. Además, algunos peces desovan en nidos construidos específicamente para proteger los huevos, como montículos de arena o grava.
Una vez que los huevos son depositados, los peces pueden exhibir comportamientos defensivos. Algunas especies vigilan activamente sus nidos y ahuyentan a los depredadores que se acercan. Pueden nadar agresivamente hacia los intrusos, hacer vibrar sus aletas o chocar contra ellos para asustarlos y alejarlos.
Además de estas estrategias activas, algunos peces cuentan con mecanismos físicos para proteger sus huevos. Por ejemplo, ciertas especies tienen huevos adhesivos que se adhieren a las superficies donde son depositados, dificultando que los depredadores los consuman. Otros peces como los gobios construyen túneles o cavidades para depositar sus huevos, proporcionando una protección adicional.
La persecución de potenciales depredadores de huevos en peces es esencial para aumentar las posibilidades de supervivencia de la descendencia. A través de estrategias defensivas y la elección de sitios adecuados para el desove, los peces buscan minimizar los riesgos y maximizar la reproducción exitosa. Estos comportamientos defensivos son fundamentales para mantener el equilibrio en los ecosistemas acuáticos y preservar la diversidad de las poblaciones de peces.
Oxigenación de los huevos
El avivar o salpicar huevos o crías con la boca, las aletas o el cuerpo es un comportamiento común observado en peces para proporcionar oxígeno y eliminar los sedimentos y los desechos metabólicos de sus huevos y crías. Este comportamiento es crucial para garantizar una adecuada oxigenación y mantener un entorno limpio y saludable para el desarrollo de la progenie.
Cuando los peces depositan sus huevos, estos pueden estar cubiertos de sedimentos o escombros. Para evitar que los huevos se asfixien o se vean afectados por la acumulación de desechos metabólicos, los padres los salpican o avivan con movimientos rápidos y vigorosos.
El avivamiento de los huevos y crías implica la utilización de la boca, las aletas o el cuerpo para crear corrientes de agua alrededor de ellos. Estas corrientes ayudan a mantener un suministro constante de oxígeno y promueven la circulación del agua, lo que ayuda a eliminar los sedimentos y los desechos.
Algunas especies de peces, como los cíclidos y los peces luchadores, son conocidas por su dedicación en avivar sus huevos. Los padres alternan entre frotar suavemente los huevos con la boca y agitar sus aletas para crear movimientos de agua que promueven la oxigenación y eliminación de desechos.
En el caso de los peces que cuidan alevines, el avivamiento también es esencial para proporcionar un suministro adecuado de oxígeno a las crías en desarrollo. Los padres pueden nadar cerca de los alevines y agitar su cuerpo o aletas para generar corrientes de agua que los mantengan bien oxigenados.
Además de la oxigenación y la eliminación de desechos, el avivamiento de los huevos y crías también puede tener beneficios sociales. Algunas especies de peces utilizan este comportamiento como una forma de comunicación entre los padres y la progenie, fortaleciendo los vínculos familiares y brindando una protección adicional.
Eliminación de huevos muertos y enfermermos
Los peces tienen instintos naturales para eliminar huevos muertos o enfermos como parte de su comportamiento parental. Esta acción es importante para mantener la salud y la viabilidad de la progenie restante.
Cuando los peces detectan huevos muertos o enfermos, los padres pueden realizar diversas acciones para eliminarlos. Usualmente, esto implica recoger los huevos con la boca y escupirlos lejos del nido o área de desove. Algunas especies también pueden utilizar sus aletas o incluso frotar sus cuerpos contra los huevos para desalojar los huevos no viables.
La eliminación de huevos muertos o enfermos es esencial para evitar la propagación de enfermedades y para garantizar que los recursos disponibles se utilicen de manera eficiente en la crianza de la progenie sana. Los huevos muertos o enfermos pueden ser presa fácil para bacterias, hongos u otros patógenos, y si no se eliminan, pueden afectar negativamente a los huevos sanos y propagar enfermedades en el nido.
Además de eliminar los huevos muertos o enfermos, algunos peces también pueden realizar comportamientos de limpieza para mantener el nido o área de desove en condiciones óptimas. Esto puede incluir remover sedimentos, desechos o algas que puedan acumularse alrededor de los huevos.
Es importante destacar que la habilidad de los peces para detectar y eliminar huevos muertos o enfermos puede variar según la especie y las condiciones ambientales. Algunas especies pueden ser más meticulosas en la limpieza y eliminación, mientras que otras pueden depender más de factores naturales como las corrientes de agua para llevarse los huevos no viables.
Algunos peces tienen la capacidad de secretar un moco especializado que desempeña un papel crucial en la protección contra patógenos y en la alimentación de sus crías. Este moco tiene propiedades antimicrobianas que ayudan a inhibir el crecimiento de bacterias, hongos y otros patógenos, brindando una defensa natural contra infecciones.
El moco secretado por los peces contiene sustancias químicas activas, como proteínas y péptidos antimicrobianos, que pueden actuar de diferentes maneras para prevenir la colonización y el crecimiento de patógenos. Estas sustancias pueden alterar la estructura de las membranas celulares de los patógenos, interferir con sus procesos metabólicos o estimular respuestas inmunológicas en los peces y sus crías.
Además de su función antimicrobiana, el moco especializado también puede desempeñar un papel en la alimentación de las crías. Algunas especies de peces, como los bagres, producen un moco nutritivo que es consumido por sus crías. Este moco contiene nutrientes esenciales que las crías necesitan para su crecimiento y desarrollo temprano. Las crías se alimentan libremente del moco secretado por sus padres, lo que les proporciona una fuente de alimento abundante y accesible durante sus primeras etapas de vida.
El moco especializado en peces es una adaptación notable que demuestra la capacidad de estas especies para proporcionar protección y nutrición a sus crías. La secreción de moco antimicrobiano es una estrategia defensiva efectiva que contribuye a la supervivencia de los peces y a la salud de sus crías. Además, la producción de moco nutritivo asegura que las crías tengan acceso a nutrientes esenciales en las primeras etapas críticas de su desarrollo. Estas adaptaciones muestran la complejidad y la diversidad de las estrategias que los peces han desarrollado para asegurar la supervivencia y el éxito reproductivo.
Nidos vivos
Los peces han desarrollado diversas estrategias para transportar huevos o crías. Algunas especies llevan los huevos o las crías en la boca o en cámaras branquiales especiales. Por ejemplo, los peces ciclídos tienen una estructura llamada bolsa bucal donde los padres guardan los huevos o las crías para protegerlos de depredadores o de condiciones desfavorables. Otros peces, como algunos tipos de tiburones y rayas, pueden llevar sus huevos o crías en una bolsa de cría ventral, que es una estructura especializada ubicada debajo del cuerpo. También hay peces que llevan sus huevos o crías externamente en la cabeza, la espalda o el vientre. Un ejemplo de esto es el pez emperador, donde el macho lleva los huevos adheridos a su piel hasta que eclosionan.
Enrollarse alrededor de la masa de huevos es otra estrategia utilizada por algunos peces para prevenir la desecación y proteger los huevos. Esto se observa en peces como los gobios y los bagres, que se envuelven alrededor de los huevos con sus cuerpos flexibles. Al hacerlo, forman una barrera física que ayuda a retener la humedad y evitar que los huevos se sequen. Además, al enrollarse, los peces también pueden proporcionar una protección adicional contra depredadores y mantener una temperatura más estable para los huevos.
Estas adaptaciones en los peces demuestran la diversidad de estrategias que han evolucionado para asegurar la supervivencia de sus huevos y crías. El transporte de huevos o crías en la boca, en bolsas especializadas o externamente, así como el enrollamiento alrededor de los huevos, son comportamientos importantes que contribuyen al éxito reproductivo y la supervivencia de las especies de peces. Cada una de estas estrategias está adaptada a las necesidades específicas de cada especie y a las condiciones ambientales en las que viven.
Aprovisionamiento y acompañamiento
Algunas especies de peces tienen comportamientos parentales que implican acompañar a sus crías mientras forrajean y proporcionarles refugio o defensa cuando los depredadores se acercan. Estos comportamientos son fundamentales para proteger a las crías y garantizar su supervivencia.
Cuando las crías de peces están en una etapa vulnerable, los padres suelen acompañarlas durante las actividades de búsqueda de alimento. Los padres nadan junto a las crías, guiándolas hacia áreas donde pueden encontrar comida adecuada. Durante esta actividad, los padres también vigilan de cerca a sus crías para asegurarse de que estén a salvo y alerta ante cualquier amenaza potencial.
Cuando los depredadores se acercan, los padres de peces desempeñan un papel crucial en la defensa de sus crías. Pueden desplegar comportamientos agresivos para ahuyentar a los depredadores, como embestirlos o mostrar señales de advertencia. Además, algunos peces construyen nidos o refugios donde las crías pueden esconderse en caso de peligro. Los padres permanecen cerca de las crías para protegerlas y, en algunos casos, incluso los cubren con sus cuerpos para proporcionarles un escudo protector.
En muchos peces, los padres también desempeñan un papel activo en el aprovisionamiento de alimentos para sus crías, ayudándolas en la captura de alimentos. Esto es especialmente común en peces que tienen dietas especializadas o que dependen de fuentes de alimento específicas.
Los padres pueden llevar a sus crías a áreas ricas en alimentos y mostrarles cómo atrapar presas. Pueden nadar en formación con las crías, enseñándoles técnicas de caza, como emboscar a las presas o coordinar movimientos para capturar alimentos en grupo. Además, algunos peces pueden regurgitar o soltar presas parcialmente capturadas para que las crías las atrapen, facilitando así el proceso de alimentación.
Este comportamiento de aprovisionamiento de alimentos por parte de los padres es esencial para el crecimiento y desarrollo adecuado de las crías de peces. Proporciona una fuente confiable de alimento durante las etapas tempranas de la vida, cuando las crías aún no son capaces de cazar o capturar presas por sí mismas. Además, al aprender de sus padres, las crías adquieren habilidades de caza y alimentación que serán cruciales para su supervivencia a largo plazo.
Referencias bibliográficas
Referencias bibliográficas
(Helfman, Collette, Facey, & Bowen, 2009; Kardong, 2011, 2014)
Awruch, C. A. (2015). Reproduction strategies. In Fish Physiology (Vol. 34, pp. 255–310). Elsevier.
Helfman, G. S., Collette, B. B., Facey, D., & Bowen, B. W. (2009). The diversity of Fishes, biology, evolution, and ecology (2nd ed.). West Sussex UK: John Wiley & Sons Ltd.
Kardong, K. V. (2011). Vertebrates, comparative anatomy, function, evolution (6th ed.). McGraw-Hill New York.
Kardong, K. V. (2014). Vertebrates: Comparative Anatomy, Function, Evolution (7th ed.). McGraw-Hill Education.
No hay comentarios:
Publicar un comentario