
[Mitosis y meiosis] [Microorganismos] [Plantas] [Invertebrados] [Peces] [Anfibios y reptiles] [Aves] [Humanos] [Enfermedades humanas]
Índice
Índice
Portada

Portada
1. Introducción
1. Introducción
Los pavos reales machos usan sus coloridas plumas para atraer a las hembras. Las hembras tienden a aparearse con los machos con las plumas más grandes y brillantes. Una cola grande y brillante indica que el macho está sano y es probable que produzca descendencia saludable. Probablemente también hayas escuchado a los pájaros "cantar". Las águilas hacen acrobacias aéreas y caídas. Estas son otras formas de atraer parejas. ¿Cómo se reproducen las aves? Sabemos que los pollos ponen huevos. Pero, ¿cómo lo hacen? Todo comienza con un comportamiento dirigido a atraer a una pareja. En aves, esto involucrará un tipo de exhibición, generalmente realizada por el macho. Algunas demostraciones son muy elaboradas y pueden incluir bailes, vuelos aéreos o tambores de ala o cola. La mayoría de los pájaros machos también cantan un tipo de canción para atraer a las hembras. Si tienen éxito en atraer a una hembra, esto conducirá a la reproducción. Las aves se reproducen por fertilización interna, durante la cual el óvulo se fertiliza dentro de la hembra. Al igual que los reptiles, las aves tienen cloaca o una sola salida y entrada para esperma, huevos y desechos. El macho lleva su esperma a la cloaca hembra. El esperma fertiliza el huevo.
Entonces el huevo de cáscara dura se desarrolla dentro de la hembra. Los huevos de cáscara dura tienen un amnios lleno de líquido, una membrana delgada que forma un saco cerrado alrededor del embrión. Los huevos se ponen generalmente en un nido. ¿Por qué crees que los huevos vienen en tantos colores diferentes? Las aves que hacen nidos al aire libre tienen huevos camuflados. Esto le da a los huevos protección contra la depredación. Algunas especies, como los jarros nocturnos que anidan en el suelo, tienen huevos pálidos, pero las aves camuflan los huevos con sus plumas. Para proteger a sus crías, diferentes especies de aves hacen diferentes nidos. Aves de todo tipo, desde colibríes hasta avestruces, hacen nidos. Muchos pueden ser elaborados, con forma de tazas, cúpulas, platos, montículos o madrigueras. Sin embargo, algunas aves, como las guillemot comunes, no usan nidos. En su lugar, ponen sus huevos en acantilados desnudos. Los pingüinos emperadores no tienen ningún nido; se sientan en los huevos para mantenerlos calientes antes de que eclosionen, un proceso llamado incubación. ¿De qué otra manera podría un ave ayudar a proteger a sus crías de los depredadores? La mayoría de las especies ubican sus nidos en áreas que están ocultas, para evitar depredadores. Las aves grandes, o las que anidan en grupos, pueden construir nidos a cielo abierto, ya que son más capaces de defender a sus crías.
En las aves, del 90% al 95% de las especies son monógamas, lo que significa que el macho y la hembra permanecen juntos para reproducirse durante algunos años o hasta que una pareja muere. Las aves de todo tipo, desde loros hasta águilas y halcones, son monógamas. Normalmente, los padres se turnan para incubar los huevos. Las aves suelen incubar sus huevos después de que el último ha sido puesto. En las especies polígamas, donde hay más de una pareja, la madre realiza toda la incubación. El pavo salvaje es un ejemplo de un pájaro polígamo. La duración y el tipo de cuidado de los padres varía ampliamente entre las diferentes especies de aves. En un extremo, en un grupo de aves llamadas magapodos (que son aves parecidas a pollos), el cuidado de los padres termina con la eclosión. En este caso, el pollito recién salido del montículo del nido sin la ayuda de los padres y puede cuidarse a sí mismo de inmediato. Estas aves se llaman precoces. Otras aves precoces incluyen el pollo doméstico y muchas especies de patos y gansos. En el otro extremo, muchas aves marinas cuidan a sus crías durante largos períodos de tiempo. Por ejemplo, los pollitos de fragata común reciben cuidado intensivo de los padres durante seis meses, o hasta que estén listos para volar, y luego toman 14 meses adicionales de ser alimentados por los padres. Estas aves son lo opuesto a las aves precoces y se llaman altriciales. En la mayoría de los animales, el cuidado parental masculino es raro. Pero es muy común en las aves. A menudo, ambos padres comparten tareas como la defensa del territorio y el sitio del nido, la incubación y la alimentación de los polluelos. Dado que las aves a menudo cuidan mucho a sus crías, algunas aves han desarrollado un comportamiento llamado parasitismo de cría. Esto sucede cuando un pájaro deja sus huevos en el nido de otro pájaro. El ave anfitriona a menudo acepta y levanta los huevos del ave parásito.
Ernst Walter Mayr
Ernst Walter Mayr (1904-2005) fue un renombrado biólogo y ornitólogo alemán-estadounidense. Reconocido como uno de los fundadores de la moderna síntesis evolutiva, sus contribuciones a la biología evolutiva y la taxonomía lo convirtieron en una figura influyente en el campo. Mayr también desempeñó un papel destacado en la preservación de la biodiversidad y la promoción de la conservación. Su legado perdura como un hito en la comprensión de la evolución y la diversidad biológica.

Contexto
Ernst Walter Mayr vivió en un momento de intensos cambios políticos, económicos, sociales, religiosos y culturales. Nació en 1904, durante el apogeo del Imperio Alemán, en una época de creciente nacionalismo y tensiones geopolíticas que finalmente desembocaron en la Primera Guerra Mundial. Durante su juventud, presenció los estragos de la posguerra y el surgimiento del movimiento nazi en Alemania.
En términos económicos, Mayr fue testigo de la Gran Depresión en la década de 1930, una crisis económica mundial que tuvo un impacto significativo en la sociedad y la política. También vivió la Segunda Guerra Mundial y su posterior reconstrucción, en la cual se produjeron cambios socioeconómicos y políticos significativos en Europa y Estados Unidos.
En cuanto a lo social, Mayr experimentó una época de rápidos cambios en las normas y valores sociales. Presenció el surgimiento del feminismo y el movimiento por los derechos civiles, así como los avances en la medicina y la tecnología, que transformaron la sociedad en varios aspectos.
Desde el punto de vista religioso, Mayr creció en una sociedad predominantemente cristiana, pero también fue testigo de un aumento en la secularización y la diversidad religiosa durante su vida. Estos cambios en la religión y la espiritualidad influyeron en la forma en que se abordaron los temas científicos y filosóficos, como la evolución y la existencia de Dios.
Culturalmente, Mayr fue influenciado por la rica tradición intelectual alemana, así como por los desarrollos científicos y culturales en Europa y Estados Unidos. También fue testigo de la emergencia del cine, la música y otras formas de entretenimiento como parte integral de la cultura popular.
Infancia
La infancia de Ernst Walter Mayr estuvo marcada por el ambiente cultural y científico en el que creció. Nació el 5 de julio de 1904 en Kempten, en el Reino de Baviera, Alemania. Su familia pertenecía a la clase media-alta y tenía un profundo interés por la música, la literatura y las ciencias naturales.
Mayr desarrolló una pasión por la ornitología desde temprana edad. Durante su infancia, pasaba largas horas observando aves y recolectando especímenes. Su interés por la naturaleza fue fomentado por su padre, un abogado y apasionado naturalista aficionado.
Además de su fascinación por la naturaleza, Mayr también fue educado en un entorno intelectualmente estimulante. Su madre era hija de un distinguido profesor de filosofía, lo que le brindó acceso a una amplia gama de conocimientos y discusiones intelectuales.
A pesar de su entorno favorable, la infancia de Mayr no estuvo exenta de dificultades. Durante la Primera Guerra Mundial, Alemania se sumió en el caos y la escasez, y Mayr experimentó las privaciones de la guerra. Sin embargo, esta experiencia también le permitió desarrollar una mentalidad resiliente y una apreciación por la estabilidad y la paz.
Logros
La educación de Ernst Walter Mayr sentó las bases para su carrera científica y sus notables logros en el campo de la biología evolutiva. Después de completar su educación secundaria, Mayr ingresó a la Universidad de Greifswald en 1923, donde estudió zoología, botánica y geología. Posteriormente, continuó sus estudios en la Universidad de Berlín y obtuvo su doctorado en 1926.
Uno de los principales logros científicos de Mayr fue su contribución a la síntesis evolutiva moderna. A mediados del siglo XX, Mayr fue uno de los principales defensores de la teoría de la evolución de Darwin y trabajó en la integración de los principios genéticos de Mendel con la teoría de la evolución de las especies. Sus investigaciones y publicaciones sobre la especiación y la genética de poblaciones fueron fundamentales para desarrollar una comprensión más completa de los mecanismos de la evolución.
Mayr también realizó importantes contribuciones a la taxonomía y la sistemática. Propuso el concepto biológico de especie y desarrolló el concepto de barrera reproductiva, que ayudó a explicar la formación de nuevas especies. Su trabajo en este campo sentó las bases para la clasificación moderna de organismos y tuvo un impacto duradero en la disciplina.
Entre sus publicaciones más destacadas se encuentra su libro "Systematics and the Origin of Species" (1942), en el cual desarrolló sus ideas sobre la especiación y la evolución. También escribió "Population, Species, and Evolution" (1970), considerado uno de los libros fundamentales en el campo de la biología evolutiva.
Aunque Mayr fue un científico muy respetado, no estuvo exento de controversias y errores. En su obra, a veces fue criticado por su énfasis en la selección natural como el principal motor de la evolución, dejando de lado otros factores importantes. También hubo debates en torno a sus interpretaciones de la genética de poblaciones y la definición de especie.
Sin embargo, a pesar de estas controversias y errores, el legado de Ernst Walter Mayr es innegable. Su enfoque integral y su capacidad para integrar diferentes campos de la biología sentaron las bases para nuestra comprensión moderna de la evolución. Su trabajo sigue siendo una referencia crucial en la biología evolutiva y su influencia perdura en la disciplina.
Comunidad científica
La comunidad científica en la que Ernst Walter Mayr se desarrolló fue un entorno intelectualmente estimulante y dinámico, compuesto por prominentes científicos de diversos campos. Durante su carrera, estableció relaciones significativas con colaboradores, amigos, opositores y, en ocasiones, incluso enemigos dentro de la comunidad científica.
Uno de los colaboradores más destacados de Mayr fue Theodosius Dobzhansky, con quien mantuvo una relación de trabajo cercana y duradera. Juntos, contribuyeron de manera significativa a la síntesis evolutiva moderna, combinando los principios genéticos con la teoría de la evolución. Su colaboración llevó a la publicación conjunta de trabajos científicos y a la organización de conferencias y simposios.
Otros colaboradores y amigos cercanos de Mayr incluyeron a Julian Huxley, George Gaylord Simpson y David Lack, quienes también desempeñaron roles destacados en la biología evolutiva y la ornitología. Estos científicos compartían intereses científicos similares y colaboraron en diversos proyectos y publicaciones.
Sin embargo, no todas las interacciones de Mayr con otros científicos fueron armoniosas. Tuvo desacuerdos y debates con algunos de sus contemporáneos, como Richard Goldschmidt y Sewall Wright. Goldschmidt defendía la teoría del "salto macroevolutivo" y criticaba la importancia de la selección natural en la formación de nuevas especies, mientras que Wright tenía enfoques teóricos diferentes en cuanto a la genética de poblaciones. Estos desacuerdos dieron lugar a debates intensos y a veces a rivalidades científicas.
En términos de opositores y enemigos, Mayr enfrentó críticas y disputas con científicos que se oponían a sus ideas o enfoques científicos. Algunos detractores argumentaban en contra de sus interpretaciones de la especiación y la genética de poblaciones. Sin embargo, es importante destacar que estas controversias y desacuerdos son inherentes al proceso científico y no necesariamente indican una enemistad personal.
En general, la comunidad científica en la que Mayr se insertó fue un entorno altamente competitivo y colaborativo al mismo tiempo. Si bien tuvo colaboradores cercanos y amistades duraderas, también enfrentó desafíos y desacuerdos con científicos que tenían puntos de vista diferentes. A pesar de estas dinámicas, Mayr se destacó como una figura influyente en la biología evolutiva y dejó un impacto duradero en la comunidad científica.
Reconocimientos
Ernst Walter Mayr fue ampliamente reconocido y honrado tanto en vida como póstumamente por sus contribuciones sobresalientes a la biología y la ciencia en general. A continuación, se detallan algunos de los reconocimientos más destacados que recibió:
(a) Medalla Darwin: En 1959, Mayr fue galardonado con la prestigiosa Medalla Darwin por la Real Sociedad de Londres, en reconocimiento a su destacada labor en el campo de la biología evolutiva.
(b) Medalla de la Academia Nacional de Ciencias de Estados Unidos: En 1965, Mayr fue honrado con la Medalla de la Academia Nacional de Ciencias de Estados Unidos, uno de los mayores reconocimientos científicos en ese país.
(c) Medalla Balzan: En 1983, recibió la Medalla Balzan en el campo de la biología evolutiva y la genética de poblaciones. Esta distinción es otorgada anualmente por la Fundación Internacional Balzan para reconocer logros destacados en diversas áreas de estudio.
(d) Medalla Crafoord: En 1999, la Real Academia Sueca de Ciencias concedió a Mayr la Medalla Crafoord en Biología Evolutiva, en reconocimiento a su sobresaliente investigación en este campo.
(e) Honores y membresías académicas: Mayr fue elegido miembro de numerosas academias y sociedades científicas, incluyendo la Academia Nacional de Ciencias de Estados Unidos, la Sociedad Filosófica Americana, la Real Sociedad de Londres y la Academia de Ciencias de Baviera.
Además de estos reconocimientos formales, Mayr recibió numerosos honores y títulos honoríficos de universidades y organizaciones de todo el mundo en reconocimiento a su destacada carrera científica. Su trabajo también fue ampliamente celebrado por la comunidad científica y se le considera uno de los biólogos más influyentes del siglo XX.
Tras su fallecimiento en 2005, se llevaron a cabo varios eventos en su honor, incluyendo conferencias, simposios y homenajes. Su legado continúa siendo celebrado y su trabajo sigue siendo referente en el campo de la biología evolutiva.
¿Por qué él es importante?
Ernst Walter Mayr y su contribución a la biología evolutiva tienen un impacto significativo en nuestra vida cotidiana, aunque a menudo no seamos conscientes de ello. Aquí hay algunas formas en las que su trabajo influye en nuestras vidas:
(a) Comprender la evolución: El trabajo de Mayr ayudó a desarrollar y establecer la teoría de la evolución de Darwin como el marco central para comprender cómo cambian y se adaptan las especies a lo largo del tiempo. Esta comprensión nos permite apreciar cómo los organismos evolucionaron y se diversificaron, incluyendo a los humanos. Nos ayuda a entender nuestra relación con otras especies y cómo estamos conectados a través de la historia de la vida en la Tierra.
(b) Medicina y salud: La comprensión de los principios evolutivos es fundamental en la medicina. Ayuda a entender la evolución de enfermedades, como la resistencia a los antibióticos y la adaptación de los virus. Además, la investigación evolutiva en genética y biología molecular ha llevado a avances en diagnóstico, terapias y desarrollo de medicamentos.
(c) Conservación de la biodiversidad: La comprensión de la evolución y la diversidad biológica que promovió Mayr es esencial para la conservación de la naturaleza. Nos ayuda a apreciar el valor de la biodiversidad y a comprender los procesos evolutivos que dan forma a los ecosistemas. Esto a su vez nos permite tomar decisiones informadas y responsables para proteger y preservar la biodiversidad y los hábitats naturales.
(d) Agricultura y seguridad alimentaria: La investigación en evolución y genética ha mejorado significativamente la agricultura y la seguridad alimentaria. El conocimiento de la selección natural y la diversidad genética ha permitido el desarrollo de cultivos más resistentes a enfermedades y plagas, así como la cría selectiva de animales para obtener características beneficiosas. Esto contribuye a una producción agrícola más eficiente y sostenible.
(e) Tecnología y diseño: El estudio de la evolución también ha inspirado avances en campos como la inteligencia artificial y el diseño de sistemas complejos. Los algoritmos evolutivos se utilizan para resolver problemas de optimización y diseño, y se han aplicado en campos tan diversos como la ingeniería, la arquitectura y la informática.
¿Qué debemos aprender de él?
Ernst Walter Mayr nos ofrece valiosas lecciones e inspiración a través de su vida, limitaciones, obstáculos, logros e impacto en el campo de la biología evolutiva. Aquí hay algunas formas en las que podemos aprender de su ejemplo:
Mayr demostró una dedicación y pasión inquebrantables por la biología y la ornitología a pesar de las limitaciones y dificultades. A pesar de las restricciones impuestas por el contexto político y social en el que vivió, siguió persiguiendo su pasión por la ciencia. Su historia nos enseña la importancia de perseverar a pesar de las barreras y las limitaciones que puedan surgir en nuestro camino.
A lo largo de su vida, Mayr se enfrentó a obstáculos y críticas en su trabajo científico. Sin embargo, no se dejó desanimar y continuó desafiando las convenciones establecidas en su campo. Su capacidad para enfrentar y superar obstáculos nos inspira a ser valientes y resilientes en nuestras propias metas y proyectos, incluso cuando encontramos resistencia o desafíos inesperados.
Mayr dedicó su vida al estudio y la investigación científica, y su legado perdura hasta el día de hoy. Nos muestra la importancia de seguir aprendiendo y creciendo intelectualmente, así como la capacidad de generar un impacto duradero a través de nuestras contribuciones. Su trabajo nos anima a buscar el conocimiento y a dejar una marca significativa en nuestro campo de interés.
Mayr combinó diferentes disciplinas científicas, como la genética, la biología y la ornitología, para desarrollar su comprensión de la evolución. Su enfoque integral nos enseña la importancia de adoptar una perspectiva amplia y considerar múltiples facetas de un tema para obtener una comprensión más completa. Nos inspira a ser curiosos y a buscar conexiones entre diferentes áreas de conocimiento.
Mayr no solo fue un destacado científico, sino también un comunicador efectivo. Fue capaz de transmitir sus ideas y conocimientos de manera accesible para un público más amplio. Su habilidad para comunicar conceptos complejos nos recuerda la importancia de compartir el conocimiento y hacerlo comprensible para diferentes audiencias, lo cual nos motiva a mejorar nuestras habilidades de comunicación y divulgación científica.
2. Temporadas y territorios reproductivos
2. Temporadas y territorios reproductivos
Las aves se adaptan para reproducirse en una asombrosa diversidad de hábitats, y la biología de la reproducción aviar es, por consiguiente, variada y compleja. Para apreciar parte de esta maravillosa diversidad en cómo se reproducen las aves, considere por un momento las vidas de tres tipos de entornos muy diferentes en todo el mundo.

Figura 2.1. El Aptenodytes forsteri, comúnmente conocido como pingüino emperador, es una especie emblemática de la Antártida. Es el pingüino más grande y alto, alcanzando alturas de hasta 1,2 metros y un peso de alrededor de 40 kilogramos. Su plumaje distintivo es de color negro en la espalda y cabeza, mientras que su pecho y vientre son de un blanco brillante. Estos majestuosos pájaros son conocidos por su habilidad para nadar y bucear, pudiendo sumergirse a profundidades de hasta 500 metros y permanecer bajo el agua durante más de 20 minutos. Su ciclo de vida incluye la incubación de huevos en el frío extremo y la formación de grandes colonias para enfrentar los desafíos del clima antártico.
Primero, imagine que es la mitad del invierno justo afuera de la estación antártica francesa en Terre Adélie. Un grupo de pingüinos emperador macho (Aptenodytes forsteri) se encuentra amontonado en el viento helado y aullando. Estas aves permanecerán en este lugar durante más de 100 días, cada uno incubando su único huevo encima de sus pies dentro de una solapa gruesa y aislada de carne y plumas. El huevo eclosionará justo cuando la luz del sol regresa en la primavera antártica, lo que le da al nuevo polluelo pingüino la mayor parte del verano altamente productivo para crecer, desarrollarse y nadar antes de que el sol desaparezca nuevamente.
Figura 2.2. Alectura lathami, conocido como el pavo pincel, es un ave nativa de Australia. Los machos tienen un plumaje único y llamativo, con un copete de plumas en forma de pincel en la cabeza. Su cuerpo es de color negro con motas blancas y marrones. Habitan en bosques y selvas del este de Australia, donde se alimentan de insectos, semillas y pequeños invertebrados. Estas aves son terrestres y pasan la mayor parte del tiempo en el suelo. Los machos realizan elaboradas exhibiciones de cortejo, sacudiendo su copete y mostrando su plumaje colorido para atraer a las hembras. Son una especie emblemática de la fauna australiana.
Luego, imagínese en un bosque húmedo costero mientras observa a un macho australiano de Pavo Pincel (Alectura lathami) que clava su pico en la parte superior de una pila gigante de tierra y hojas. Este macho es el constructor, tierno y protector de este impresionante montículo. En el interior, el proceso de descomposición genera calor para incubar los huevos que las hembras han puesto y enterrado dentro del montículo. Tan pronto como eclosionen, estos huevos producirán la mayor independencia de las aves jóvenes, los pollitos que caminarán o volarán hacia el bosque para tener una vida por sí mismos.

Figura 2.3. El Palmero Africano (Cypsiurus parvus) es una pequeña ave migratoria que se encuentra en diversas regiones de África. A pesar de su nombre, no está relacionado con las palmeras, sino que es un miembro de la familia de los vencejos. Estas aves tienen un plumaje marrón oscuro en la espalda y una cola larga y bifurcada. Son conocidas por su vuelo ágil y rápido, realizando acrobacias en el aire mientras cazan insectos. Los palmeros africanos construyen sus nidos en acantilados, estructuras rocosas o edificios altos, utilizando fibras vegetales y saliva. Son aves sociales y a menudo se congregan en grandes grupos. Su presencia añade vida y movimiento a los cielos africanos.
Finalmente, imagine que se encuentra en una arboleda de palmeras en la orilla de un río africano, donde ve al palmero africano (Cypsiurus parvus) regresando a su nido inusual. La hembra ha usado su saliva pegajosa para pegar un estante poco profundo de pelusa y plumas al lado de una hoja de palma que cuelga hacia abajo, y luego ha pegado sus huevos de manera similar en este nido mínimo. Ella y su pareja se turnan para incubar estos huevos acurrucándose junto a ellos en la hoja de palma. Tan pronto como eclosionen, los polluelos se agarrarán a este nido precario y permanecerán allí durante semanas; mientras tanto, sus padres regresarán cientos de veces para aprovisionar a los polluelos a medida que crezcan lo suficientemente, como para alimentarse de insectos voladores por su cuenta.

Figura 2.4. Temporada de reproducción y disponibilidad de alimentos. Los jilgueros americanos (Spinus tristis) se reproducen más tarde en el verano que la mayoría de las aves insectívoras que ocupan el mismo hábitat, ya que dependen de las semillas maduras de los cardos y otras plantas para construir sus nidos y alimentar a sus crías.
Estas son solo tres de las muchas formas en que diferentes aves han evolucionado para criar en diferentes entornos. Podemos decir que sabemos más sobre la biología reproductiva de las aves que sobre cualquier otro aspecto de sus vidas, y este capítulo describe algunas de las generalidades compartidas por todas las aves reproductoras junto con algunas de las fascinantes variaciones en las estrategias reproductivas de los diferentes grupos de aves.
Temporadas de reproducción
La mayoría de las aves se reproducen en un tiempo limitado que depende de la estacionalidad del ambiente local y de los aspectos de la historia natural de cada especie, como su dieta, depredadores, tipo de nido y hábitat. La mayoría de las aves en las regiones templadas deben reproducirse dentro del período rico en recursos en la primavera que es impulsado por una inundación estacional en la vegetación, la incubación de insectos y el calor del verano que se aproxima.

Figura 2.5. El Hormiguero Moteado (Hylophylax naevioides) es una especie de ave paseriforme que se encuentra en América del Sur, especialmente en países como Brasil, Bolivia y Perú. Estas aves tienen un plumaje marrón oscuro en la espalda y una serie de manchas blancas en el pecho y la garganta, de ahí su nombre. Son conocidas por su comportamiento único de buscar alimento, ya que se especializan en forrajear entre las hojas y ramas de los árboles en busca de insectos y arañas. Además, tienen un canto distintivo y melódico que utilizan para comunicarse y marcar su territorio. El Hormiguero Moteado es una especie fascinante que contribuye a la diversidad y equilibrio de los ecosistemas boscosos sudamericanos.
Las fluctuaciones estacionales de la temperatura se vuelven menos dramáticas hacia el ecuador, y en muchos hábitats tropicales es la variación estacional en la precipitación “lluvias” lo que impulsa los ciclos anuales en la disponibilidad de recursos. Por lo tanto, el tiempo de reproducción suele ser menos limitado para las aves que se reproducen en los trópicos, donde algunas especies se reproducen exclusivamente durante una estación seca, otras encuentran condiciones óptimas durante la transición entre las estaciones seca y húmeda, y algunas pueden reproducirse durante todo el año.
El momento de disponibilidad de alimentos para sus crías es el factor que parece tener la mayor influencia general sobre cuándo se reproducen las aves. Idealmente, los polluelos eclosionan justo cuando esos recursos alimenticios son más abundantes. Las aves a menudo deben poner huevos mientras los suministros de alimentos siguen aumentando, especialmente en entornos con fuertes cambios estacionales en la disponibilidad de recursos. Dado que muchas especies, especialmente las paseriformes “con forma de gorrión”, alimentan principalmente de insectos a sus polluelos, estos padres aviares a menudo ponen huevos justo antes de la abundancia máxima de insectos. En los lugares más estacionales hacia los polos de la tierra, la temporada de reproducción de los insectívoros aviares puede durar solo un mes aproximadamente debido al período correspondiente breve de la abundancia de insectos en los hábitats árticos.

Figura 2.6. El Jilguero Yanqui o Jilguero Norteamericano (Spinus tristis) es un pequeño pájaro cantor que se encuentra en América del Norte. Los machos tienen un plumaje amarillo brillante en el cuerpo, con una mancha negra en el pecho y rayas negras en las alas. Las hembras son más apagadas en color, con tonos verdes y marrones. Estas aves son conocidas por su hermoso canto melódico y su capacidad para imitar otros sonidos. Se alimentan principalmente de semillas y frutas. El Jilguero Yanqui es popular entre los observadores de aves y se mantiene como mascota en algunos lugares. Su belleza y melodioso canto lo convierten en un verdadero tesoro de la fauna norteamericana.
Las especies de aves que alimentan a sus polluelos con alimentos que no sean insectos a menudo se reproducen en diferentes momentos que los insectívoros en la misma comunidad aviar. Por ejemplo, los jilgueros americanos (Spinus tristis) de América del Norte dependen casi exclusivamente de las semillas de cardo para alimentar a sus crías, y por lo tanto esta especie se reproduce a mediados del verano cuando las semillas de cardo maduran, en lugar de a principios de la primavera. En otros lugares, muchas especies de mielero en Australia se reproducen cuando y donde pueden encontrar las abundantes flores de las especies clave de árboles de eucalipto. Del mismo modo, los Halcones de Eleonora (Falco eleonorae) en el Mediterráneo oriental ponen huevos a fines del verano, asegurando que las demandas de alimentos máximas de sus polluelos coincidan con la temporada principal de caza de los padres durante la migración de aves terrestres de otoño.
Cuanto más regulares sean los cambios estacionales, más fácil será para un ave potencialmente reproductora predecirlos. En la mayoría de los entornos, las fluctuaciones de un año a otro son lo suficientemente pequeñas como para que la temporada de reproducción se encuentre dentro de un período predecible dentro de cada año. La mayoría de los sistemas reproductivos de las aves están preparados para estar activos solo durante estos meses. El reloj interno aviar se basa en los cambios en la duración del día, o fotoperíodo, que dan lugar a una cascada de preparaciones fisiológicas para la reproducción. Las aves generalmente usan el fotoperíodo para establecer su período de reproducción general, pero a menudo se basan en otras indicaciones para afinar el inicio de la reproducción. Por ejemplo, en los bosques húmedos de Panamá, la presencia de insectos vivos ayuda a estimular las aves macho moteadas (Hylophylax naevioides) en condiciones de alta reproducción.

Figura 2.7. El Quelea Común (Quelea quelea) es un ave pequeña y gregaria que se encuentra en varias partes de África. Con su plumaje marrón y su pico corto y cónico, estos pájaros son conocidos por su gran capacidad de adaptación y su habilidad para formar enormes bandadas. Son considerados una de las aves más abundantes del mundo. Sin embargo, también se les considera una plaga agrícola, ya que su gran número puede causar daños significativos en los cultivos. A pesar de ello, el Quelea Común desempeña un papel ecológico importante como polinizador y dispersor de semillas, y su comportamiento colectivo es un espectáculo impresionante en la naturaleza africana.
Cuanto más estrechamente se relaciona una señal ambiental con una característica ambiental crítica, mejor sirve como base para la programación reproductiva. Por ejemplo, muchas aves que anidan en ambientes áridos, como la Quelea (Quelea quelea) de África, son impulsadas a reproducirse después de una fuerte lluvia. Lo mismo ocurre con muchas aves que anidan en el desierto, como los pinzones de la cebra (Taeniopygia guttata) de Australia central y varias especies de pinzones de tierra en las tierras bajas áridas de las Islas Galápagos (los de Darwin). Para todas estas especies, la lluvia es un indicio que predice la disponibilidad futura de semillas e insectos relacionados con las plantas que crecen solo cuando hay agua disponible.

Figura 2.8. El Diamante Cebra (Taeniopygia guttata) es un pequeño pájaro originario de Australia, conocido por su plumaje blanco y negro con distintivas rayas en forma de "cebra". Son aves sociales y gregarias, y se encuentran comúnmente en áreas abiertas como pastizales, praderas y arbustos. Son apreciados como mascotas debido a su belleza y su capacidad de adaptación en cautiverio. Además, son reconocidos por su melodioso canto y su capacidad de aprender y imitar otros sonidos. Los Diamantes Cebra se alimentan principalmente de semillas y pequeños insectos. Su presencia añade un toque de color y alegría a los paisajes australianos y son un favorito entre los amantes de las aves.
Debido a que la producción de semillas de estas varía mucho de un lugar a otro y de un año a otro, las loxias cruzadas pueden reproducirse tan pronto como encuentren un suministro adecuado de alimentos, sin importar en qué mes del calendario se encuentre. Las aves, como las loxias cruzadas que están listas para reproducirse al encontrar condiciones favorables, deben pagar los costos de mantener sus sistemas reproductivos durante todo el año. En contraste, la mayoría de las aves, particularmente aquellas en ambientes fuertemente estacionales, cierran su sistema reproductivo durante la temporada de no reproducción.
Territorios reproductivos
Casi todas las aves defienden algún tipo de territorio durante la temporada de reproducción. Aunque son costosos en términos de tiempo, energía y riesgo personal, los beneficios de defender un territorio a menudo superan estos costos. El tamaño y la riqueza de los recursos de un territorio con frecuencia influyen en la elección de pareja y el éxito de reproducción posterior. Los machos de muchas especies, por lo tanto, defienden grandes territorios de usos múltiples para atraer a una o más hembras y proporcionar alimentos suficientes para la cría de pollos.

Figura 2.9. El Colibrí Caribeño Gorgimorado (Eulampis jugularis) es una especie de colibrí que habita en las islas del Caribe. Este hermoso pájaro destaca por su plumaje brillante y colorido, con un tono azul iridiscente en la cabeza y una garganta de un intenso color morado. Su pequeño tamaño y su vuelo ágil les permiten alimentarse del néctar de las flores mientras flotan en el aire. Son polinizadores clave en su hábitat, y también complementan su dieta con insectos pequeños. El Colibrí Caribeño Gorgimorado es un símbolo de la belleza y la diversidad de la fauna caribeña y representa la delicadeza y la elegancia en el reino de las aves.
Por ejemplo, las hembras de los chipes que anidan en los pantanos en Eurasia y los mirlos en América del Norte pueden elegir parejas en función del tamaño del territorio de un macho. De manera similar, los caribes de garganta púrpura (Eulampis yugularis) de las Antillas Menores prefieren a los machos con una mayor disponibilidad de néctar en sus territorios. Los machos en esta especie de colibrí defienden mucho más néctar del que necesitan para su propio uso, reservando partes de sus territorios para las hembras.
Los territorios también reducen la posibilidad de que otros individuos de la misma especie interfieran con las actividades de reproducción del propietario del territorio. Este tipo de interferencia de reproducción es un gran problema para algunas especies. Por ejemplo, los pergoleros satinados (Ptilonorhynchus violaceus) machos de Australia y Nueva Guinea elaboran estructuras elaboradas para atraer a las hembras, a menudo robando materiales de machos vecinos; si el dueño masculino no lo vigila por un solo día, un nido puede desaparecer por completo.

Figura 2.10. El Pergolero Satinado (Ptilonorhynchus violaceus) es una especie de ave nativa de Australia. Los machos de esta especie son conocidos por su asombrosa habilidad para construir estructuras elaboradas llamadas pergolas. Utilizan palos, ramas y otros objetos para crear plataformas ornamentadas que decoran con objetos coloridos como flores, bayas, piedras y plumas, con el objetivo de atraer a las hembras durante el cortejo. Estos pájaros tienen un plumaje de tonos oscuros y brillantes, con destellos violetas y azules. Además de su habilidad constructiva, tienen un canto distintivo y melodioso. El Pergolero Satinado es una maravilla de la naturaleza y ejemplifica la creatividad y el esplendor del reino animal.

Figura 2.11. El Periquito Coliverde (Forpus passerinus) es una pequeña especie de loro originaria de América Central y del Sur. Estos encantadores pájaros destacan por su plumaje vibrante, siendo principalmente de color verde con un tono azul en las alas y una mancha roja en la frente de los machos. Son aves sociales y activas, formando bandadas y comunicándose entre sí con chillidos agudos y llamativos. Son capaces de aprender a imitar algunas palabras y sonidos. Se alimentan principalmente de semillas, frutas y flores. Los Periquitos Coliverde son populares como mascotas debido a su tamaño compacto y a su naturaleza juguetona. Su belleza y vivacidad los convierten en una adición colorida a la familia de las aves.
Muchas aves también pueden intentar destruir los huevos de los competidores cercanos. Por ejemplo, en América del Norte, los paseriformes pertenecientes a las especies (Cistothorus palustris) y (Troglodytes aedon) perforan de forma rutinaria los huevos de los conespecíficos y otros competidores potenciales en el sitio del nido; Las roselas escarlata (Platycercus elegans) hace lo mismo en Australia. Las aves reproductoras a menudo mitigan el riesgo de este tipo de perturbaciones al proteger sus nidos. Por ejemplo, en el norte de América del Sur, las parejas de (Forpus passerinus) resguardan sus huevos con diligencia. Para probar la velocidad de destrucción de la nidada sin la defensa de los padres, los investigadores establecieron nidos experimentales llenos de réplicas de los huevos de esta especie. Estos nidos falsos no estaban defendidos, y los vecinos cercanos destruyeron el 40% de estas nidadas en 3 días, en comparación con el 5% de los nidos reales que fueron defendidos activamente.
3. Anatomía del sistema reproductor de las aves
3. Anatomía del sistema reproductor de las aves
Las gónadas masculinas se denominan testículos. De forma ovalada o elíptica, los testículos aviares están emparejados y se encuentran dentro de la cavidad del cuerpo en el extremo anterior de cada riñón. En general, los testículos producen uno de los dos tipos de células espermáticas, dependiendo de los pecies: un tipo corto y simple en órdenes no paseriformes y un tipo más largo en forma de espiral en paseriformes (Figura 3.1).

Figura 3.1. Espermatozoides (izquierda), Protuberancia cloacal (derecha).
Con el enfoque de la temporada de reproducción, los testículos aviares aumentan enormemente en tamaño. Conductos deferentes transportan espermatozoides desde los testículos hasta la cloaca.
La configuración de cada conducto crea la ilusión de estrías. En la mayoría de las aves, una inflamación terminal en forma de huso en el extremo de cada conducto, llamada ampolla “hemipene de las aves”, se abre como una papila en la cloaca. Cuando el macho invade su cloaca durante la copulación, esta papila entra en la cloaca hembra y puede incluso entrar en contacto con la abertura del oviducto. Durante la temporada de reproducción, la región cloacal de los paseriformes machos se hincha tanto que la protuberancia sirve como un indicador de la condición reproductiva de un ave (y por representación, su sexo).

Figura 3.2. Sistema urogenital aviar. Dado que los sistemas urinario y reproductivo funcionan juntos en las aves y en general, en todos los vertebrados, los componentes principales incluyen las gónadas, los riñones, los uréteres y la cloaca. Aunque sus anatomías urogenitales son similares, (A) los machos tienen testículos y un conducto eyaculador, y (B) las hembras tienen un ovario y un oviducto.
Sistema reproductor femenino
La gónada hembra u ovario (Figura 12) contiene muchos óvulos. En este contexto, el término "huevo" es confuso, ya que puede referirse a la célula reproductiva femenina en general o a la versión descascarada de la misma célula después de que haya pasado por el tracto reproductivo. Para evitar confusiones, en este capítulo usamos el término "óvulo" (óvulos plurales) mientras estas células están en el ovario femenino, en lugar de "huevo”. La mayoría de las aves solo tienen un ovario funcional, el izquierdo. Durante la ovulación, un óvulo sale del ovario hacia la cavidad abdominal general o celoma.

Figura 3.3. La ovulación y el paso de un óvulo a través del tracto reproductivo. La mayoría de las aves hembras solo tienen un ovario funcional. Para iniciar la ovulación, el infundíbulo se abre temporalmente para permitir la entrada de un óvulo. A partir de ahí, viaja hasta el Magnum, donde se secreta la albumina (clara de huevo) alrededor del óvulo y la yema. En el istmo, se depositan más albúmina acuosa y membranas de la cáscara. La cáscara se forma a medida que el óvulo pasa a través de un istmo corto hacia el útero, donde se agregan la cáscara y los pigmentos. Luego, el óvulo normalmente gira 180 grados en la vagina para colocarse con el extremo romo primero, a través de la cloaca (YouTube).
Suspendido de la pared dorsal del cuerpo por un ligamento, el oviducto comienza como una abertura aplanada, en forma de embudo, llamada infundíbulo. El infundíbulo luego "traga" el óvulo “igual que en los mamíferos”.
Después de ingresar al infundíbulo, una larga región glandular llamada magnum segrega albúmina (clara de huevo rica en proteínas), que protege el óvulo y eventualmente sirve como suministro de agua del embrión. Las membranas de la cáscara se forman cuando el óvulo pasa a través de un istmo corto. El óvulo luego viaja al útero bien vascularizado o glándula de la concha. Aquí, las papilas secretoras depositan calcio mineral, que forma la cáscara.
Dependiendo del ave, esta glándula puede segregar pigmentos también. Un óvulo generalmente pasa la mayor parte de su tiempo (18-20 horas) en la glándula de la concha. En la última parte del oviducto, la mayoría de las aves giran el huevo ya formado 180 grados antes de pasarlo por los extremos romos hacia la cloaca.
Copulación y fertilización
La inseminación ocurre cuando el esperma masculino ingresa a la cloaca de una hembra. En este proceso, el ave macho generalmente se para en la espalda de la hembra mientras que ambas aves giran sus cloacas y las presionan juntas “monta”. El esperma se transfiere cuando la papila del conducto deferente masculino entra en contacto con el revestimiento de la cloaca hembra o, en algunos casos, entra en la abertura de su oviducto. A menudo denominado "beso cloacal", este contacto entre un par de apareamiento es breve.

Figura 3.4. Falo cloacal. Algunas especies de aves tienen un falo cloacal, una adaptación que mitiga las situaciones morfológicas o ecológicas que dificultan el montaje y la copulación (como el apareamiento en el agua). Algunos patos tienen falos cloacales inusualmente largos; este Pato del lago (Oxyura vittata) tiene el récord del falo aviar más largo con 42,5 cm.
Avestruces, rheas, estaamas, patos y gansos tienen un órgano copulador más avanzado, el falo cloacal (Figura 3.4). Aunque a menudo se denomina "pene", esta estructura difiere del equivalente de los mamíferos: ya que carece de una uretra interna, los espermatozoides deben viajar a lo largo de la superficie externa del falo.
Debido a que muchas aves hembras almacenan el esperma en los túbulos ubicados dentro del oviducto, los espermatozoides generalmente sobreviven más tiempo dentro de las aves que en los mamíferos. Es probable que los espermatozoides se liberen de estos túbulos después del paso de un óvulo. Un pavo hembra puede poner hasta 15 huevos fértiles después de una sola inseminación; Incluso después de 30 días, el 83% de sus huevos pueden ser fértiles. En contraste, las especies que ponen un solo huevo en cada temporada, como el Griffon Euroasiático (Gyps fulvus) de Europa y África, pueden copular con frecuencia durante un mes antes de la puesta. Un período de copulación prolongado puede proporcionar más oportunidades para la competencia de espermatozoides, unión de parejas y / o garantía de paternidad.

Figura 3.5. El Griffon Euroasiático (Gyps fulvus) es una imponente especie de buitre que se encuentra en Europa, África y Asia. Con una envergadura de alas de hasta 2,8 metros, estos buitres son conocidos por su distintivo plumaje color canela y su cabeza cubierta de plumas blancas. Se alimentan principalmente de carroña, desempeñando un papel crucial en el ecosistema al limpiar los restos de animales muertos. Son aves gregarias que forman colonias de cría en acantilados y árboles. Desafortunadamente, el Griffon Euroasiático ha enfrentado amenazas significativas debido a la pérdida de hábitat y la intoxicación por sustancias químicas. Sin embargo, los esfuerzos de conservación están ayudando a proteger a esta majestuosa especie y preservar su importancia en los ecosistemas.
Los espermatozoides depositados en la cloaca femenina entran al oviducto y nadan hasta el infundíbulo, donde la fertilización se lleva a cabo antes de que la albúmina cubra el óvulo. Los núcleos de la célula espermática y el óvulo se unen para formar el cigoto. La división celular avanza a medida que el óvulo fertilizado viaja por el oviducto. Después de que la hembra pone un huevo, el crecimiento del cigoto cesa hasta que comienza la incubación.
4. Control hormonal del comportamiento sexual
4. Control hormonal del comportamiento sexual
El mundo de las aves está lleno de una amplia variedad de comportamientos fascinantes, y uno de los aspectos más intrigantes es el control hormonal del comportamiento sexual. Las aves exhiben una amplia gama de comportamientos reproductivos, desde el cortejo elaborado y las exhibiciones de apareamiento hasta la construcción de nidos y el cuidado de los polluelos. Detrás de esta diversidad de comportamientos se encuentran complejos procesos hormonales que regulan y desencadenan estas conductas específicas.
El papel de las hormonas en la regulación del comportamiento sexual en aves ha sido objeto de numerosos estudios y descubrimientos científicos. Las hormonas, como la testosterona y los estrógenos, desempeñan un papel fundamental en la diferenciación sexual, la maduración sexual y la expresión de comportamientos relacionados con la reproducción.
Este artículo se adentrará en el emocionante campo del control hormonal del comportamiento sexual en aves, explorando los mecanismos biológicos subyacentes, los factores que influyen en la producción hormonal y cómo estas hormonas modulan el cortejo, la elección de pareja, la construcción de nidos y el cuidado parental en diferentes especies de aves.
A través de una revisión de estudios científicos recientes, se examinarán los avances en nuestra comprensión de cómo las hormonas influyen en el comportamiento sexual en aves y cómo este conocimiento puede tener implicaciones más amplias en el campo de la biología del comportamiento y la conservación de aves.
Adentrémonos en el apasionante mundo de las aves y descubramos cómo las hormonas moldean su comportamiento sexual, desvelando los misterios de la conexión entre los mecanismos hormonales y las conductas reproductivas en estas criaturas aladas.
Determinación del sexo
Un pájaro comienza la vida como un huevo fertilizado que ya tiene un sexo, las aves no poseen hermafroditismo. Las aves hembras tienen cromosomas sexuales ZW, mientras que las aves machos tienen cromosomas sexuales ZZ. Las hembras producen óvulos que contienen un cromosoma Z o W, que luego son fertilizados por el esperma Z, por lo que las hembras son el parental que determina el sexo. Debido a que algunas aves ponen huevos con relaciones sexuales que se desvían de radio esperado 1:1, existe un interés considerable en comprender qué determina qué cromosoma sexual ingresa en un óvulo, cómo el sistema de una hembra podría sesgar ese proceso y si las hormonas sexuales de la madre son parte de este proceso. Las hormonas de la madre incluyen aquellas en su circulación, pero también las que pasan a las yemas de los huevos, que han demostrado ejercer efectos de larga duración en otros aspectos de la descendencia. En este momento aún no está claro cómo se producen los sesgos en la proporción de sexos o en qué medida están involucradas las hormonas en el proceso.

Figura 4.1. Los ejes endocrinos HPA y HPG. Los estímulos ambientales desencadenan la regulación hormonal interna a través de los ejes endocrino hipotálamo-pituitario-adrenal (HPA) e hipotalámico-pituitario-gonadal (HPG). La regulación hormonal es imperativa para las respuestas adaptativas, incluidas las conductas reproductivas y territoriales, las respuestas de lucha o huida a las amenazas y la inquietud migratoria.
El eje HPG
Las hormonas gonadales y el eje HPG “hipotalámico-pituitaria-gonadal” que las regula están involucrados en la mayoría de los aspectos de la reproducción aviar y de los mamíferos, incluida la producción de óvulos y espermatozoides, el cortejo y el comportamiento de apareamiento, la construcción de nidos y la incubación. También sirven para promover otros tipos de comportamiento relacionados con el éxito reproductivo, como la defensa territorial. Además, estas hormonas son responsables de algunas de las formas en que los machos y las hembras se ven diferentes y difieren en la anatomía reproductiva interna.
Diferenciación sexual
Durante el desarrollo en el óvulo, el embrión aviar forma testículos u ovarios y luego los órganos internos masculinos o femeninos, un proceso llamado diferenciación sexual. Estos órganos incluyen los gonaductos masculinos y femeninos, que eventualmente se convertirán en los vasa deferentia en los machos y en el oviducto de las hembras. Las gónadas embrionarias comienzan a parecerse al tejido ovárico o testicular y producen hormonas al principio del desarrollo, después de unos pocos días de incubación. En las especies de aves mejor estudiadas (pollos, codornices y patos domésticos), las gónadas de ambos sexos producen andrógenos y estrógenos, pero los ovarios embrionarios producen mucho más estrógeno, por lo que la proporción de estrógenos a andrógenos es mayor en los embriones femeninos.
Activación reproductora
Estas gónadas embrionarias también producen una hormona no esteroidea que causa la regresión de los gonaductos embrionarios femeninos en los embriones masculinos y la regresión del gonaducto derecho en las hembras. Todo esto tiene lugar antes de la eclosión. Más adelante en sus vidas, a medida que las aves entran en la madurez reproductiva, los aumentos de los esteroides sexuales gonadales causan el crecimiento de los órganos que se organizaron durante la vida embrionaria. En las hembras, los estrógenos ováricos agrandan el oviducto izquierdo, y en los machos, los andrógenos testiculares agrandan los vasos deferentes. Esta segunda etapa de la acción hormonal en los órganos reproductivos se conoce como activación.
Caracteres sexuales secundarios
Algunas especies de aves tienen caracteres anatómicos adicionales que difieren según el sexo, como el falo de las aves acuáticas masculinas (pero no las hembras), los panales de pollos (más grandes y más rojos en los machos) y los plumajes más coloridos y ornamentados de los machos de muchas especies. Los experimentos realizados en la década de 1950 por el pionero endocrinólogo francés Etienne Wolff mostraron que los embriones de pato desarrollan un falo si las hormonas ováricas no están presentes, y no desarrollan un falo si están presentes. En este caso, la ausencia de hormonas da el fenotipo masculino, y su presencia da el fenotipo femenino. Otras diferencias sexuales surgen de diferencias más sutiles en los niveles hormonales. Por ejemplo, ambos sexos de pollos (y su ancestro salvaje, Gallus gallus) tienen peines, y la diferencia de sexo en el tamaño y color del peine es principalmente una respuesta a los niveles más altos de testosterona en los machos adultos.

Figura 4.2. Caracteres sexuales secundarios. La cresta o peine es un carácter sexual secundario que ayuda a diferencia entre machos y hembras en los pollos comunes, así como la cola.
Existe una notable variación entre los grupos de aves en cuanto a la forma en que las hormonas están involucradas en la generación de diferencias sexuales en el plumaje. En aves acuáticas y galliformes (grupos hermanos en el árbol filogenético de aves vivas), las aves adultas tienen el plumaje masculino siempre que no tengan los altos niveles de estrógeno asociados con los ovarios.
Si las gónadas se eliminan de ambos sexos, los machos conservan una apariencia masculina y las hembras adquieren el plumaje masculino. Los andrógenos como la testosterona no están involucrados en la producción de las lujosas plumas masculinas. En algunos pájaros cantores, los machos tienen características debido a los niveles más altos de testosterona, como un babero negro del que carecen las hembras (como en los gorriones caseros, Passer domesticus), o los machos que tienen un color más rojo que las hembras (como en los pinzones domésticos, Haemorhous mexicanus).

Figura 4.3. Los pinzones domésticos (Haemorhous mexicanus), también conocidos como pinzones de casa o pinzones mexicanos, son aves nativas de América del Norte. Estos pequeños pájaros se caracterizan por su plumaje robusto y de tonos rosados, grises y marrones. Son aves comunes en entornos urbanos y suburbanos, adaptándose fácilmente a la presencia humana. Los pinzones domésticos son reconocidos por su canto alegre y su capacidad de formar parejas monógamas a largo plazo. Se alimentan principalmente de semillas y brotes, pero también pueden consumir insectos pequeños. Su presencia en los jardines y parques agrega color y alegría a nuestro entorno diario.
Un tercer patrón ocurre en otras especies paseriformes, como los pinzones de la cebra (Taeniopygia guttata), en los que las hormonas gonadales parecen no tener nada que ver con las diferencias de sexo en los adornos de plumaje. Las hembras genéticas con testículos funcionales generan el plumaje femenino, y los machos genéticos con ovarios funcionales retienen el plumaje masculino. Por lo tanto, no es posible hacer afirmaciones generales sobre todas las aves sobre la relación entre las hormonas y el dimorfismo sexual, como "la testosterona hace que los machos se parezcan a los machos".
Comportamiento hormonal-sexual
Los machos y las hembras se comportan notablemente diferente en la mayoría de las especies de aves, especialmente durante la temporada reproductiva. A veces solo los machos cantan. A veces las hembras tienen una postura especial de solicitud de copulación. Algunas veces las hembras se preocupan por las crías solas, pero en ocasiones los machos crían a los polluelos. El acto de apareamiento difiere entre los sexos, con los machos generalmente montando a la hembra.
Es fácil suponer que las hormonas tienen algo que ver con estas diferencias de sexo, pero muy pocas veces se comprende realmente el papel de las hormonas. La mayor parte de lo que se ha establecido proviene de un pequeño número de aves domesticadas. En algunos casos, la diferencia de sexo es principalmente un fenómeno de activación hormonal. Por ejemplo, si a las canarias adultas (Serinus canaria) se les administra testosterona, comienzan a cantar como los machos. Si se le administra estradiol a la codorniz japonesa macho (Coturnix japonica), adoptan la postura receptiva femenina y permiten que otros machos se apareen con ellos. Por lo tanto, estas diferencias de sexo parecen ser una consecuencia del dimorfismo sexual en los niveles de hormonas en individuos adultos. La razón por la cual las hembras muestran la postura receptiva durante el apareamiento y los machos no, es porque las hembras tienen más estrógeno (la hormona de la receptividad o feminidad) que los machos.

Figura 4.4. La codorniz japonesa (Coturnix japonica) es una especie de ave originaria de Asia Oriental, especialmente de Japón. Estas aves son conocidas por su tamaño compacto y su plumaje moteado en tonos marrones y beige. Son criadas tanto por su carne como por sus huevos, siendo consideradas una especie de ave de granja. Además, son populares en la avicultura de compañía debido a su fácil cuidado y reproducción. La codorniz japonesa es famosa por su capacidad de puesta de huevos, pudiendo poner una gran cantidad de huevos pequeños en un corto período de tiempo. Su adaptabilidad y su importancia económica han contribuido a su presencia en diferentes partes del mundo.
Sin embargo, el hecho de que los machos se monten y las hembras no surge antes en el desarrollo; Al igual que las hormonas embrionarias organizan la anatomía sexual interna, también tienen un efecto permanente sobre si las aves mostrarán un montaje durante el apareamiento (Figura 5.1).
5. La gametogénesis en las aves
5. La gametogénesis en las aves
Las gónadas de las aves tienen funciones duales: producen hormonas, pero también gametos (óvulos y espermatozoides). Las hormonas gonadales viajan a otras partes del cuerpo para actuar sobre órganos diana distantes y participan en la regulación de la producción de óvulos y espermatozoides.
La espermatogénesis
Tamaño testicular
Los espermatozoides aviares, como los de otros grupos de vertebrados, se producen en grandes cantidades, y muchos miles o incluso millones están contenidos en un solo eyaculado. La producción de esperma se correlaciona con el tamaño general de los testículos, que a su vez se asocia con el sistema de apareamiento de las especies y el grado de competencia espermática entre los eyaculados de diferentes machos. Los testículos son relativamente pequeños en especies de aves que son estrictamente monógamas y más grandes en aquellas en que las hembras se aparean con múltiples machos.
La formación de los espermatozoides
Al igual que en los mamíferos, la espermatogénesis en aves se define como la formación de espermatozoides maduros a partir de espermatogonias e incluye tres pasos secuenciales: espermatocitogénesis, espermatidogénesis y espermiogénesis. Las características anatómicas y el control fisiológico de la espermatogénesis aviar han sido objeto de extensos estudios. Los primeros trabajos identificaron uno o dos tipos de espermatogonias, pero los trabajos posteriores en codornices japonesas encontraron que los túbulos seminíferos contenían cuatro tipos de espermatogonias (Ad, Ap1, Ap2 y B) que difieren con respecto a su intensidad de tinción y características ultraestructurales. Más recientemente, se describieron tres tipos de espermatogonia en el pavo. Durante la espermatocitogénesis, un espermatogonio (célula germinal Ad) sufre mitosis para generar una nueva célula Ad y una célula Ap1, que se divide en dos células Ap2 hijas. Cada célula Ap2 se divide en dos espermatogonias B, que a su vez se dividen en dos espermatocitos primarios. Los espermatocitos primarios experimentan dos divisiones meióticas, dando a luz a espermatocitos secundarios, que se dividen en espermátidos (espermatidogénesis). Como resultado de estas divisiones celulares, cada Ad spermatogonia puede generar 32 espermátidas (Deviche, Hurley, & Fokidis, 2011).
Las espermátidas se transforman en células de esperma durante la espermiogénesis, que se ha estudiado mejor en codornices japonesas y pollos. La espermiogénesis se caracteriza por una reducción dramática de la cantidad de citoplasma y una profunda transformación morfológica que da como resultado células espermáticas maduras que consisten en una cabeza (acrosoma, núcleo cilíndrico y mitocondria) y cola (flagelo), y la espermiación es la liberación de esperma de Células de Sertoli en el lumen de los túbulos seminíferos. La estructura de los espermatozoides maduros en los paseriformes está relativamente conservada, pero difiere en varios aspectos de la de los no paserinos. La información limitada en aves sugiere una duración más corta y quizás más consistente de la espermatogénesis que en los mamíferos (pollo: 12 a 13 días; codorniz japonesa: 13 días) (Deviche et al., 2011).
Maduración, transporte y almacenamiento
Los espermatozoides liberados en los túbulos seminíferos se suspenden en un líquido seminífero y se transportan a la cloaca a través de conductos excurrentes que consisten en el rete testis, que comprende una región intra y extratesticular; el conducto eferente; el conducto de conexión el epidídimo; y el conducto deferente. La ontogenia y las características citológicas adultas de los conductos excurrentes se han descrito en detalle. El esperma en las aves se transporta rápidamente a lo largo del tracto reproductivo. En las codornices japonesas, este transporte requiere aproximadamente 24 horas. Durante este tiempo, los espermatozoides experimentan una maduración final, la mayor parte del líquido seminífero se reabsorbe por pinocitosis y la composición del líquido se altera para producir plasma seminal. Los espermatozoides en los túbulos seminíferos, el epidídimo y el conducto deferente en los pollos carecen de motilidad, pero, al ser transportados a lo largo de los conductos excurrentes, los espermatozoides de pollo y codorniz adquieren la posibilidad de motilidad debido a la influencia de factores, como el Ca2þ y el glutamato, que se segregan en la glándula accesoria los fluidos reproductivos (Deviche et al., 2011).

Figura 5.1. La manipulación hormonal altera el comportamiento de la monta. Normalmente, solo los machos se montan para iniciar la copulación. Sin embargo, cuando los embriones de codorniz japonesa (Coturnix japonica) se tratan con estradiol, no se pueden montar como adultos. Los varones tratados son efectivamente "demasculinizados", aunque genéticamente masculinos, pero conductualmente femeninos. Los embriones hembras tratados con un inhibidor de estrógeno no se someten al proceso normal de demasculinización y, por lo tanto, exhiben un comportamiento masculino en la adultez.
En el pollo, el movimiento flagelar depende de la temperatura y se cree que la motilidad real del esperma se adquiere por una disminución de la temperatura en el momento de la eyaculación y la transferencia de la eyaculación a la cloaca hembra. La motilidad de los espermatozoides en la paloma (Columba livia) es más alta entre 18 y 24 ºC y disminuye a 28 ºC. La motilidad del esperma en el pollo también depende de los factores genéticos que influyen en la función y la morfología mitocondrial (Deviche et al., 2011).
En los paseriformes, el extremo distal del conducto deferente forma el glomus seminal, una estructura especializada de almacenamiento de espermatozoides. El glomus en los paseriformes puede haber evolucionado como resultado del estilo de vida aéreo de estas aves, que es incompatible con los testículos muy grandes. El número de espermatozoides en el glomus seminal del gorrión doméstico disminuye durante el transcurso del día. Por lo tanto, el glomus puede acumular esperma durante la noche para usarlo durante el día en el curso de múltiples copulaciones. Los no paseriformes tienen una capacidad limitada para el almacenamiento extragonadal de esperma (Deviche et al., 2011).
Ovogénesis y la formación del huevo
A diferencia de los mamíferos hembra, las aves hembras pueden almacenar esperma viable durante días o incluso semanas, dependiendo de la especie. Una parte del oviducto contiene túbulos especiales de almacenamiento de esperma, y los espermatozoides almacenados se liberan desde allí para viajar hasta el sitio de fertilización cerca de la parte superior del oviducto. La forma en que funcionan estos túbulos y cuál es su función en la competencia de esperma o en las estrategias de fertilización femenina es poco conocida. Lo que está claro, sin embargo, es que, aunque el óvulo solo es fertilizable durante un breve período de tiempo a través del oviducto superior, las hembras no necesariamente tendrían que aparearse en un momento determinado del día o incluso el día de la semana para obtener una fecundación exitosa. Toda camada de huevos para ser fertilizada. Los espermatozoides que ya se han almacenado pueden liberarse para hacer ese trabajo.
Los huevos aviares son grandes, y se requieren varios días para que un huevo que esté listo para ser puesto. La formación de los huevos, la ovulación y la secuencia de puesta se entienden mejor en los pollos domésticos, pero es probable que muchas características del proceso sean similares en otras aves que ponen una serie de huevos aproximadamente a diario hasta que se completa su nidada.
Acumulando la yema
Antes de la aparición de la cohorte de óvulos, el ovario contiene folículos primordiales. Uno a la vez al dia, un folículo comienza a madurar y acumular la yema. El ovario de un pájaro ponedor contiene un conjunto secuencial de folículos acumulando yema, hasta que alcanzan el tamaño que vemos en los huevos puestos.
Fertilizando el óvulo
En la ovulación, la yema se libera del ovario y entra en la parte superior del oviducto. A medida que baja por el oviducto, primero se fertiliza cerca de la parte superior del oviducto, si la hembra se ha apareado.
Acumulando la clara, las membranas y la cáscara
A continuación, adquiere una capa de albúmina ("clara de huevo"). Más cerca de la parte inferior del oviducto, se agregan las membranas de la cáscara y las capas de la cáscara, luego (si el huevo está pigmentado) los colores se agregan a la cáscara, y finalmente se pone el huevo. Todo el proceso, desde la ovulación (cuando los óvulos maduros salen del ovario) hasta la oviposición (cuando se pone un óvulo) toma aproximadamente 24 horas, y en la mayoría de las especies que se han estudiado, la ovulación del siguiente óvulo comienza dentro de una hora después de la oviposición del anterior.
Control de la puesta
Las hormonas del eje HPG están involucradas en todos los aspectos del proceso de producción de huevos y en el ciclo de ovulación y oviposición. A medida que madura el folículo ovárico, las células que lo rodean producen cantidades crecientes de estrógeno y andrógeno. Los estrógenos estimulan la producción de los principales componentes de la yema en el hígado, que se transportan a las yemas en desarrollo. A medida que se acerca la ovulación, el ambiente de la hormona sexual del folículo consiste principalmente en progesterona (Figura 5.2).
Una oleada de LH “hormona luteinizante” de la pituitaria dispara la ovulación, pero solo del folículo que está en línea para ovular. ¿Qué hace que un ave hembra comience a poner huevos en primer lugar? ¿Tiene que haber un macho alrededor para que la hembra inicie una cohorte? ¿Ella tiene que aparearse?
Las respuestas a estas preguntas dependen de las especies de aves, por razones interesantes. Como cualquiera que críe pollos sabe, no es necesario que haya un gallo en el rebaño para que las gallinas comiencen a poner huevos. Solo tienen que tener la edad suficiente para que los ovarios maduren, es decir, para que el eje HPG se acelere a un nivel suficiente; Una vez que eso sucede, las gallinas ovulan y ponen huevos espontáneamente.
Sin embargo, se sabe desde hace siglos que las palomas no suelen poner huevos a menos que haya un macho cortejando presente. Las palomas tortuga anilladas (las palomas africanas domesticadas, Streptopelia roseogrisea) responden a un macho cortejando, y el sonido de los coos de las hembras en respuesta al estímulo masculino aumenta la producción de LH su glándula pituitaria, que luego estimula el crecimiento de los folículos ováricos y la deposición de la yema.

Figura 5.2. Cambios hormonales antes de la oviposición. Los niveles circulantes de progesterona y otros esteroides aumentan unas pocas horas antes de que una hembra ponga un huevo.
Como es de esperar, los diferentes sistemas de apareamiento y cuidado de los padres ayudan a determinar qué especies de aves ovulan espontáneamente o en respuesta a una interacción con un macho. Las aves, como las gallinas, tienen el cuidado de los padres solo por parte de las hembras, y las aves no forman parejas monógamas. En este tipo de situación, la ave hembra debe producir una cohorte en un momento que sea oportuno para ella; Casi siempre hay un macho cerca para fertilizar los huevos. Sin embargo, las aves, como las palomas, tienen cuidado biparental y forman parejas macho-hembra que necesitan estar bien sincronizadas para compartir la incubación y criar a los pichones. La hembra necesita tener un compañero dispuesto a compartir esos deberes antes de proceder a producir una cohorte ovular, de lo contrario los huevos se perderán.
6. Emparejamiento, cortejo y selección sexual
6. Emparejamiento, cortejo y selección sexual
El mundo de las aves está lleno de una increíble variedad de estrategias de apareamiento y comportamientos de cortejo, que reflejan la diversidad de formas y colores que adornan sus plumajes. Desde elaboradas exhibiciones de danza y vuelo hasta la entrega de regalos y la construcción de nidos elaborados, las aves han desarrollado una amplia gama de tácticas para atraer a sus parejas potenciales y garantizar el éxito reproductivo.
Este artículo explorará el apasionante tema del emparejamiento, el cortejo y la selección sexual en aves, examinando los intrincados patrones de comportamiento que involucran a machos y hembras en la búsqueda de pareja. Desde las llamativas exhibiciones de los machos hasta las sutiles señales de elección de las hembras, descubriremos cómo las aves emplean una variedad de señales visuales, auditivas y táctiles para comunicarse y evaluar la calidad de sus posibles compañeros.
Además, se analizarán los factores que influyen en la elección de pareja, como la selección basada en el plumaje, la territorialidad, la competencia por recursos y las interacciones sociales. También se abordarán los aspectos evolutivos y adaptativos de estos comportamientos, así como su importancia en la supervivencia y el éxito reproductivo de las aves.
Mediante la revisión de estudios científicos actuales y ejemplos de diversas especies de aves, este artículo busca profundizar nuestra comprensión de los procesos que subyacen al emparejamiento, el cortejo y la selección sexual en las aves, revelando la riqueza y complejidad de la vida amorosa alada en el reino animal.
Prepárese para adentrarse en el mundo de los rituales de apareamiento y las intrigas de la selección sexual en el fascinante reino de las aves, descubriendo los secretos y las maravillas de sus comportamientos de emparejamiento únicos y cautivadores.
Elección femenina
Entre las aves hembra y macho, es difícil creer que hasta mediados de la década de 1900, muchos biólogos creían que las hembras en general (¡incluso en nuestra propia especie!) carecían de preferencias por atributos particulares en sus parejas potenciales. La visión estándar solía ser que los machos controlaban el juego de apareamiento al competir entre ellos por hembras pasivas.

Figura 6.1. El pavo real (Pavo cristatus) es una de las aves más hermosas y majestuosas del reino animal. Originario de India y Sri Lanka, destaca por su plumaje exuberante y llamativo. El macho, conocido como pavo real, despliega un abanico de plumas de colores vibrantes y patrones complejos durante el cortejo, creando un espectáculo visual deslumbrante. Estas impresionantes exhibiciones son un medio para atraer a las hembras, que presentan un plumaje más discreto. Además de su belleza, los pavos reales tienen una vocalización característica, emitiendo fuertes llamadas y vibrantes chillidos. Su elegancia y singularidad los convierten en un símbolo de gracia y esplendor en el mundo aviar (YouTube).
Charles Darwin, por el contrario, argumentó que las hembras de muchas especies de aves tenían lo que él denominaba "sensibilidades estéticas" que emplean al considerar a sus compañeros de pareja. Darwin sabía que en parejas de aves en las que los sexos diferían en rasgos como la habilidad para cantar, las danzas de exhibición o la intensidad de su plumaje, el macho sería casi siempre el sexo con la mayor capacidad para cantar, las exhibiciones de cortejo más extrañas, y el plumaje más llamativo. Darwin se preguntó de forma algo retórica si “todos los machos de la misma especie excitan y atraen a las mujeres por igual. ¿O es que ella elige, y prefiere a ciertos machos?” y se respondió a si mismo afirmando que las hembras sí tienen preferencias por ciertos machos.
El canto masculino
En los últimos años, una explosión de investigaciones ha demostrado que Darwin estaba completamente en lo cierto al observar que las hembras están lejos de ser espectadores neutrales cuando se trata de seleccionar parejas. Considere, por ejemplo, los experimentos que han demostrado cómo los pinzones cebra (Taeniopygia guttata) femeninas evalúan a los machos a través de sus canciones (Woolley & Doupe, 2008). Cuando los machos de pinzones cebra cantan en presencia de una hembra, tienden a producir canciones que son más rápidas y más estandarizadas que las que cantan cuando no hay hembras cerca. Al usar dos parlantes para reproducir canciones más rápidas y más lentas, los investigadores pueden evaluar cuál de estas variantes prefieren las hembras. Resulta que cuando a los pinzones cebra se les da esta opción, generalmente se acercan al orador que toca el tipo de canción más rápida, demostrando así su preferencia por este tipo de canción.
La presunción subyacente de tal experimento es que, bajo condiciones naturales, los pinzones cebra femeninos que buscan parejas tienden a acercarse y elegir machos capaces de cantar la calidad de la canción preferida. Esta suposición también se ha probado en experimentos con pinzones cautivos. En otras palabras, la preferencia de asociación de una hembra también revela su preferencia sexual. Muchos otros experimentos sobre cómo las hembras responden a las señales sexuales de los machos han demostrado que las hembras suelen prestar mucha atención incluso a las diferencias sutiles en los comportamientos masculinos u otros rasgos. Las vocalizaciones masculinas con frecuencia son un componente de las exhibiciones sexuales que las aves hembra usan para evaluar a sus parejas.
Aunque la canción masculina en general es una señal sexual muy común en muchos grupos de aves, los rasgos de canción precisos preferidos por las hembras varían ampliamente de una especie a otra: algunas aves hembras evalúan a los machos en función de su velocidad de producción de señales, otras por la presencia de las frecuencias sonoras preferidas. Otros prefieren las vocalizaciones masculinas con un amplio rango de frecuencia o un gran número total de tipos de canciones en el repertorio masculino.
Ninguno de estos atributos de canción se favorece universalmente en todas las aves. Aún queda mucho por aprender acerca de cómo un tipo particular de preferencia de señal por parte de las hembras de una determinada especie se traduce en beneficios reproductivos para esas hembras. Al igual que ningún elemento acústico predomina en las preferencias de pareja femenina, tampoco un aspecto visual del cortejo cumple con todas las especies de aves.
La danza de cortejo
En algunas aves, las preferencias femeninas giran en torno a la intensidad y la frecuencia con que los machos realizan presentaciones visuales que muestran plumas de la cola, plumas de las alas o bolsas de garganta. Por lo tanto, cuando el gallo lira común (Tetrao tetrix) se reúne en grupos en campos finlandeses para competir por los territorios de exhibición visitados por hembras selectivas, los machos difieren en su actividad general, y los exhibidores frecuentes en territorios centrales relativamente grandes aseguran más parejas que machos periféricos menos activos. Este hallazgo sugiere que las hembras evalúan a sus parejas en parte por las propiedades de sus presentaciones visuales, que determinan el éxito territorial de un macho y su capacidad para estimular a las hembras observadoras.

Figura 6.2. El gallo lira común (Tetrao tetrix), también conocido como urogallo, es una especie de ave galliforme que habita en bosques y áreas montañosas de Europa. Los machos de esta especie son notables por su apariencia impresionante, con un plumaje oscuro y plumas alargadas en el cuello que se asemejan a una lira. Durante la época de reproducción, los gallos lira realizan exhibiciones de cortejo espectaculares, desplegando sus plumas en forma de abanico y emitiendo llamadas resonantes. Estas exhibiciones tienen como objetivo atraer a las hembras y demostrar su fortaleza y dominio del territorio. Lamentablemente, el gallo lira común enfrenta amenazas debido a la pérdida de hábitat y la caza, y su conservación es de suma importancia para preservar la biodiversidad de los bosques europeos. (YouTube).
El adorno masculino
Para otras especies, el tamaño de un adorno masculino especial afecta más fuertemente a la elección de pareja femenina. Por ejemplo, algunos investigadores han encontrado una relación entre la longitud de la asombrosa cola del pavo real (Pavo cristatus) y el éxito reproductivo masculino. Las hembras pueden prestar atención a este aspecto del macho o algún otro componente de la cola, como la densidad de los ojos de pluma, al tomar una decisión sobre cuál de los diferentes machos seleccionará como compañero. Las hembras de pavo real no son las únicas hembras con tales preferencias.

Figura 6.3. El ave del paraíso de Goldie (Paradisaea decora) es una especie asombrosa que habita en las selvas tropicales de Papúa Nueva Guinea. Los machos de esta especie son conocidos por su plumaje colorido y sus elaboradas exhibiciones de cortejo. Durante el apareamiento, despliegan sus plumas largas y brillantes, formando una figura espectacular que parece un abanico radiante. Acompañan este despliegue con movimientos de danza y llamadas vocales únicas. Estas exhibiciones buscan atraer a las hembras y competir con otros machos. El ave del paraíso de Goldie es un ejemplo deslumbrante de la belleza y la sofisticación de los rituales de apareamiento en el reino animal.

Figura 6.4. El pájaro moscón europeo (Remiz pendulinus) es una pequeña y encantadora ave que se encuentra en diversas regiones de Europa y Asia. Destaca por su plumaje distintivo, con tonos de gris, blanco y negro, y por su característica cresta en forma de corona. Estas aves construyen nidos en forma de bolsa, suspendidos de las ramas de árboles, utilizando fibras vegetales y telas de araña. Los machos realizan vuelos acrobáticos durante el cortejo, mostrando sus habilidades aéreas y emitiendo llamadas melódicas. El pájaro moscón europeo es admirado por su ingenio arquitectónico y su comportamiento juguetón, añadiendo un toque de alegría y curiosidad a los entornos naturales donde se encuentra. (YouTube).
En el pájaro moscón europeo (Remiz pendulinus), las hembras selectas prefieren a los machos que tienen rayas oscuras ("máscaras") más grandes que a los machos con máscaras menos prominentes. Como resultado, los machos con máscaras grandes se aparean más rápido que sus rivales menos enmascarados.
El “matrimonio aviar”
La señal sexual que ocurre cuando los machos cortejan a las hembras puede servir para inducir a una hembra a aparearse de inmediato, pero las señales de este tipo también pueden ayudar a establecer un vínculo de pareja entre un macho y una posible pareja como preludio a la eventual cópula y reproducción. En muchas aves, los machos y las hembras forman una sociedad que dura por lo menos una temporada de reproducción, y algunas veces por más de una. En el pequeño pingüino (Eudyptula minor) de Australia y Nueva Zelanda, por ejemplo, algunas parejas permanecen juntas durante hasta 13 años.
Cortejo por comida
Un comportamiento que en algunas aves ayuda a formar y mantener una relación de pareja es la alimentación con cortejo, en la cual un macho lleva comida a una hembra durante el cortejo. Estos regalos pueden proporcionar información que influye en la intensidad del vínculo entre el macho y la hembra y, por lo tanto, la receptividad sexual de la hembra. La cantidad de alimentos entregados puede tener estos efectos en las hembras, como lo han demostrado los estudios en Polonia para alcaudón boreal Lanius excubitor). Es más probable que se recompense con sexo a los machos que ofrecen a sus parejas un gran regalo de comida, es decir, un gorrión o lagarto de tamaño considerable en lugar de un pequeño grillo.

Figura 6.5. El alcaudón boreal (Lanius excubitor) es una ave migratoria que se encuentra en regiones templadas y frías de Europa y Asia. Con su aspecto distintivo, presenta una combinación de colores en tonos de gris, blanco y negro, y un pico fuerte y curvado. Este pájaro carnívoro es conocido por su comportamiento único de empalar a sus presas en espinas o ramas afiladas para guardarlas y consumirlas más tarde. Son depredadores habilidosos que se alimentan principalmente de insectos, pequeños roedores y aves. Además de su habilidad para cazar, los alcaudones boreales son excelentes imitadores de sonidos y emiten llamadas melódicas. Su presencia en los paisajes naturales añade una nota intrigante a la fauna aviar.
Del mismo modo, los estudios a largo plazo de gaviotas tridactilas (Rissa tridactyla) en la costa de Francia han encontrado que en años sucesivos las hembras tienen más probabilidades de copular con los machos que hicieron un mejor trabajo de alimentación durante el año anterior.

Figura 6.6. La gaviota tridáctila (Rissa tridactyla), también conocida como gaviota tridáctila ártica o gaviota de patas negras, es una especie de ave marina que se encuentra en las regiones árticas y subárticas del Hemisferio Norte. Estas aves destacan por su plumaje blanco y su tamaño mediano, con alas afiladas y un característico pico amarillo con una mancha roja en la base. Su nombre se debe a que poseen solo tres dedos en las patas en lugar de los cuatro habituales en las gaviotas. Son expertas voladoras y buceadoras, alimentándose principalmente de peces y moluscos. La gaviota tridáctila es un símbolo icónico de los paisajes árticos y desempeña un papel importante en el ecosistema marino.
Los divorcios
Aunque a menudo se piensa popularmente que muchos pares de aves se “aparean de por vida”, en la mayoría de las especies a menudo se “separan” entre o incluso dentro de las estaciones de reproducción. Estas rupturas pueden ocurrir después de que muera un miembro de la pareja o cuando uno o ambos miembros se emparejan posteriormente con un individuo diferente. Los ecologistas del comportamiento generalmente se refieren a este cambio entre parejas emparejadas como divorcio, y en las aves, la hembra generalmente toma la iniciativa al cambiar de pareja.

Figura 6.7. El carbonero (Poecile atricapillus) es un pequeño pájaro que se encuentra en América del Norte. Con su plumaje distintivo, presenta una cabeza negra y una mancha blanca en las mejillas, contrastando con su cuerpo de tonos grises y crema. Estas aves son conocidas por su energía y agilidad, escalando ágilmente los troncos de los árboles en busca de insectos y semillas. Además, el carbonero es reconocido por su llamativo canto, lleno de trinos y gorjeos melódicos. Son aves sociales y forman bandadas durante el invierno para buscar alimento y protegerse del frío. El carbonero es un visitante frecuente en jardines y parques, donde alegra el entorno con su presencia y su animado comportamiento.
Las aves hembras pueden iniciar un divorcio incluso a mitad de temporada, como lo demuestra un experimento que involucra a los carboneros (Poecile atricapillus) de cabeza negra en Ontario, Canadá. Dos investigadores, Ken Otter y Laurene Ratcliffe (1996), capturaron hembras que se habían apareado con machos de alto estatus (dominantes) o de bajo estatus (subordinados). Estas hembras se mantuvieron en cautiverio el tiempo suficiente para permitir que siete hembras vecinas abandonen a su pareja actual y se muevan a territorios con los machos experimentalmente "viudos".
Seis de los siete divorciados se emparejaron con un macho de alto estatus; solo una aceptó a un miembro del grupo subordinado. Este elegante experimento muestra que las aves hembras a menudo son conscientes del estado de dominación de varios machos cercanos, o quizás están atentos a la calidad de sus territorios, y están preparadas para divorciarse de su pareja actual de menor calidad en caso de que se presente una mejor oportunidad.
Controlando el tamaño de la nidada
En otras especies, las hembras pueden evitar el divorcio, pero hacen que la cantidad de huevos que ponen dependa del estado fisiológico de un macho. Se sabe que esto ocurre en collalba negra (Oenanthe leucura), una especie que se encuentra en el suroeste de Europa y el noroeste de África. En esta especie, las hembras parecen esperar hasta que hayan observado a sus parejas llevando piedras a las cavidades de su nido antes de decidir cuántos huevos pondrán.

Figura 6.8. La collalba negra (Oenanthe leucura) es una pequeña ave migratoria que se encuentra en Europa y partes de Asia. Su nombre se debe a su distintivo plumaje, con el macho presentando una cabeza y pecho negros, mientras que la hembra tiene tonos más apagados. Estas aves se encuentran en hábitats abiertos, como pastizales y páramos, donde cazan insectos y pequeños invertebrados. Son conocidas por su vuelo ágil y su capacidad para atrapar presas en el aire. Además, las collalbas negras son reconocidas por su canto melodioso, lleno de trinos y gorjeos. Estas aves migran hacia el sur durante el invierno, regresando a sus áreas de reproducción en primavera. Su presencia en los paisajes naturales añade un toque de encanto y elegancia.
Las hembras producen más huevos cuando sus parejas se han transportado en muchas rocas pesadas durante el período de construcción del nido, mientras que los machos que no pueden, o no están dispuestos a, llevar rocas grandes a las cuevas del nido, en promedio, tienen menos intentos de copulación. Por lo tanto, el éxito reproductivo de un macho collalba negra parece depender de las decisiones de su compañero, que a su vez están influenciadas por su comportamiento.
La evolución, la calidad de la progenie y la elección femenina
Evidencia adicional ayuda a explicar por qué las hembras de collalba se basan en el carreteo de piedras como un indicador de la calidad masculina. Los machos que llevan piedras muy pesadas tienen sistemas inmunitarios más fuertes que los machos que solo tienen piedras más pequeñas. Un macho sano puede ayudar a la hembra a levantar una nidada más grande. Alternativamente, una hembra con un macho sano puede ganar trabajando más duro en un año cuando ha adquirido una pareja con "genes buenos" en lugar de frenarse para aumentar las probabilidades de que sobreviva para reproducirse nuevamente en un año futuro. Otros caracteres como la coloración también han sido vinculados con sistemas inmunes superiores.
Sin embargo, este patrón en el que las hembras emparejadas con los machos superiores ponen más huevos también podría surgir si las hembras en las mejores condiciones adquirieran machos más fuertes como compañeros. Este resultado dejaría a las hembras que no pueden producir una gran cohorte con machos de menor calidad.
Elección masculina
Muchos menos estudios han considerado si las aves macho, como las hembras, son selectas con respecto a sus parejas. Como señaló el mismo Darwin, esta relativa falta de atención a la elección de pareja por parte de los machos se debe al hecho de que los machos son mucho más propensos a cortejar a las hembras y no al revés. Por lo tanto, tiene sentido general concentrarse primero en las señales que usan las hembras cuando evalúan el cortejar de los machos en lugar de investigar el fenómeno intuitivamente menos probable de la elección de pareja por parte de los machos.
No obstante, existen razones teóricas para pensar que la elección de pareja masculina también ocurre. La monogamia social es extremadamente común entre las aves, y en la medida en que los machos se limitan a un compañero social y sexual por temporada de reproducción, ciertamente podrían ganar al seleccionar un compañero superior, uno con el cual un macho probablemente genere un número superior al promedio de descendiente sobreviviente.
En una impresionante diversidad de aves, los machos monógamos cooperan con sus compañeras para compartir la mayoría o todas las tareas asociadas con la reproducción (excepto, por supuesto, la puesta de huevos). Para que un macho así maximice el rendimiento de sus inversiones paternas, puede beneficiarse al identificar una hembra especialmente fértil o una que sea una pareja inusualmente buena. Por ejemplo, en la población europea de papamoscas cerrojillo (Ficedula hypoleuca) en el centro de España, solo algunas hembras tienen un parche blanco en la frente, y estas crían más polluelos en promedio que las hembras que carecen de este parche. Si los machos prefieren a las hembras con parches blancos, bien podrían obtener un beneficio reproductivo de su preferencia.

Figura 6.9. El papamoscas cerrojillo (Ficedula hypoleuca) es un ave migratoria que se encuentra en Europa y Asia occidental. Su plumaje contrasta con su cabeza y espalda negras, mientras que su pecho y vientre son de color blanco. Estas aves son conocidas por su comportamiento único de atrapar insectos al vuelo, volviendo rápidamente a su percha para consumir su presa. También son reconocidas por su distintivo canto, una serie de notas agudas y melodiosas. Durante la temporada de cría, construyen nidos en huecos de árboles o grietas de edificios. El papamoscas cerrojillo es un visitante bienvenido en jardines y parques, añadiendo una nota alegre y activa al entorno natural.
En muchas especies de aves, las hembras se parecen más a los machos, y esta semejanza sugiere que los sexos pueden involucrarse en la elección mutua de pareja basada en señales de apariencia. Esta afirmación puede ser particularmente cierta cuando los machos varían mucho en su atractivo, de modo que algunos machos tienen la opción de rechazar a algunas hembras en favor de otras.

Figura 6.10. El herrerillo común (Cyanistes caeruleus) es un pequeño pájaro que se encuentra en gran parte de Europa y Asia. Su plumaje característico combina tonos de azul intenso en la cabeza, alas y cola, con un pecho amarillo y blanco en el vientre. Estas aves son conocidas por su energía y agilidad, escalando troncos de árboles en busca de insectos y semillas. Además de su alimentación, el herrerillo común es reconocido por su llamativo canto, lleno de trinos y gorjeos melódicos. Son aves sociales y forman bandadas durante el invierno para buscar alimento y protegerse del frío. Su presencia en jardines y bosques añade un toque de color y alegría a los paisajes naturales.
Un experimento que documentó definitivamente las preferencias masculinas exploró los patrones de apareamiento en el herrerillo común (Cyanistes caeruleus), cuyos machos tienden a asociarse más con las hembras cuyas plumas de la corona reflejan la radiación UV y se asocian menos con las hembras cuyo plumaje UV ha disminuido experimentalmente. Los machos en esta especie, por lo tanto, seleccionan hembras en base a un rasgo de plumaje, al igual que las hembras a menudo eligen a los machos con adornos de plumas especiales. Del mismo modo, otro estudio, este en el gorrión chillón.

Figura 6.11. El gorrión chillón (Petronia petronia), también conocido como gorrión moruno, es un ave que se encuentra en diversas regiones de Europa, África y Asia. Su apariencia distintiva incluye un plumaje grisáceo, con manchas oscuras en las alas y una cara pálida con una lista negra en los ojos. Estos gorriones son conocidos por su llamado estridente y penetrante, de ahí su nombre. Son aves sociales que se encuentran en grandes bandadas y prefieren hábitats abiertos, como campos agrícolas y áreas rocosas. Se alimentan principalmente de semillas y pequeños invertebrados. El gorrión chillón es una especie adaptable y resistente, que demuestra su capacidad para prosperar en diversas condiciones y entornos.
El gorrión chillón (Petronia petronia), un pariente del gorrión común doméstico (Passer domesticus) sumamente extendido y más sombrío, documentó que las hembras con un parche de pecho amarillo reducido experimentalmente fueron cortejadas con menos frecuencia y se aparearon en una fecha significativamente más tardía que aquellas con un parche de pecho más grande. Tales estudios demuestran que la elección de pareja masculina es una realidad en al menos algunas especies de aves y sugiere que, en general, es más común de lo que generalmente se cree.

Figura 6.12. El gorrión común (Passer domesticus) es una especie de ave que se encuentra en diversas partes del mundo, desde Europa y Asia hasta América del Norte. Esta especie es ampliamente reconocida y familiar para muchos, ya que es comúnmente encontrada en áreas urbanas y rurales, donde ha logrado adaptarse a una variedad de entornos. El gorrión común ha adquirido un interés particular en el ámbito científico debido a su papel como "especie tipo" en la definición de clados de dinosaurios. La especie tipo es una especie seleccionada para representar un grupo más amplio de organismos, y en este caso, se considera que el gorrión común representa a las aves modernas.
Las aves modernas son descendientes directas de los dinosaurios, específicamente del grupo terópodo. Los terópodos incluyen dinosaurios famosos como el Tyrannosaurus rex y el Velociraptor spp. La relación entre las aves y los dinosaurios se establece mediante la observación de características anatómicas y genéticas compartidas.
El gorrión común se seleccionó como especie tipo debido a su posición evolutiva y su relación cercana con otros grupos de aves. Además, es una especie abundante y ampliamente estudiada, lo que facilita su uso como punto de referencia para comparaciones.
Los estudios paleontológicos y genéticos han demostrado que las aves modernas comparten características clave con los dinosaurios, como la presencia de plumas, estructuras esqueléticas similares y un metabolismo aviar único. Estas similitudes respaldan la teoría de que las aves son descendientes directas de los dinosaurios y que evolucionaron a partir de un ancestro común.
La elección del gorrión común como especie tipo no significa que sea el único representante de las aves modernas, sino que se utiliza como un punto de referencia para definir el grupo en estudios científicos y taxonómicos. Es importante tener en cuenta que existen muchas otras especies de aves modernas con sus propias características y adaptaciones únicas.
El primer uso del gorrión común (Passer domesticus) como especie tipo para definir taxones de dinosaurios se atribuye a Thomas Henry Huxley, un famoso biólogo y paleontólogo británico del siglo XIX. Huxley, conocido como el "Bulldog de Darwin" por su defensa de la teoría de la evolución de Charles Darwin, utilizó al gorrión común como punto de referencia en su trabajo pionero sobre la relación entre las aves y los dinosaurios. En su obra "On the Classification of Birds, and on the Taxonomic Value of the Modifications of Certain of the Cranial Bones in that Order" publicada en 1867, Huxley examinó las similitudes anatómicas entre las aves y los dinosaurios y argumentó que las aves deberían ser consideradas descendientes directas de los dinosaurios terópodos. Huxley destacó el gorrión común como una especie representativa de las aves modernas en su clasificación y discusión. Si bien Huxley fue uno de los primeros en utilizar al gorrión común de esta manera, su trabajo sentó las bases para futuras investigaciones y estudios sobre la relación entre las aves y los dinosaurios (Huxley, 1867). Desde entonces, el gorrión común ha sido ampliamente aceptado como un punto de referencia útil para comprender la evolución de las aves a partir de los dinosaurios y ha sido utilizado como especie tipo en la definición de diversos taxones dentro del grupo de los dinosaurios.
7. La importancia de la selección sexual
7. La importancia de la selección sexual
¿Qué pueden ganar las hembras selectas (y los machos selectos) de sus preferencias para las parejas capaces de proporcionarles canciones atractivas, adornos, exhibiciones y similares? Esta pregunta realmente se relaciona con el valor adaptativo de la elección de pareja y aborda las causas evolutivas a largo plazo del comportamiento femenino o masculino de una elección, no las causas fisiológicas inmediatas a corto plazo del comportamiento de un individuo determinado. La distinción es considerada esencial por la mayoría de los biólogos del comportamiento, que distinguen entre dos tipos de preguntas: a saber, preguntas próximas que tratan con los mecanismos internos que permiten a los individuos realizar ciertas acciones, en lugar de preguntas finales que examinan el evolutivo a largo plazo, es decir explicaciones de comportamientos.
Esta sección se enfoca principalmente en las causas últimas del comportamiento de las aves; por qué, por ejemplo, los individuos que prefieren ciertos atributos en sus parejas tienen más probabilidades de dejar descendientes sobrevivientes que si tuvieran preferencias diferentes. La investigación sobre el valor adaptativo de las características de los animales complementa, pero no reemplaza, los estudios sobre las causas inmediatas del comportamiento. Ambos son necesarios para pintar una imagen completa del comportamiento animal.
Beneficios genéticos
Hay muchas razones por las cuales una hembra podría obtener beneficios reproductivos de su elección de pareja o parejas. Al estudiar la elección de pareja femenina, debemos considerar la posibilidad de que las preferencias femeninas no sean realmente adaptativas, sino que surjan a través de la selección que favorezca a los machos más capaces de persuadir a las hembras para que se junten con ellos.
La hipótesis de la publicidad vacía y el hijo sexy
Esta “hipótesis de publicidad vacía” sugiere que las hembras son atraídas a aceptar espermatozoides de parejas persuasivas que realmente no tienen nada de valor que ofrecerles, solo el cortejo que activa los mecanismos próximos subyacentes en el comportamiento de apareamiento femenino. Sin embargo, bajo este escenario, incluso si las hembras no ganan nada directamente de su pareja sexual, las hembras que prefieren a los machos más estimulantes tienden a tener hijos que han heredado ese rasgo preferido y que, por lo tanto, son especialmente capaces de adquirir parejas en la próxima generación. Dadas estas condiciones, la hipótesis del anuncio vacío se convierte en la hipótesis del hijo sexy más ampliamente aceptada, con hembra selectas ganando hijos atractivos (incluso si nada más ofrecen los machos) de sus preferencias de pareja.

Figura 7.1. El estornino europeo (Sturnus vulgaris) es una especie de ave migratoria que se encuentra en Europa, Asia y partes de África. Con su plumaje iridiscente y su característico vuelo en forma de murmurationes, los estorninos europeos son reconocidos por su comportamiento social y su habilidad para formar grandes bandadas. Estas aves tienen un canto variado y melodioso, y son conocidas por su capacidad para imitar otros sonidos. Se alimentan principalmente de insectos, frutas y semillas, y su presencia en los paisajes naturales añade un toque de vitalidad y encanto. El estornino europeo es una especie adaptable y exitosa que ha logrado adaptarse a diversos entornos, incluyendo áreas urbanas.
Un ejemplo que apoya la hipótesis del hijo sexy proviene de los estorninos europeos (Sturnus vulgaris) estudiados en el sur de Alemania, donde los hijos de padres políginos defienden más cajas nido y producen más canciones de cortejo que los hijos de padres monógamos. En otras palabras, la aparente herencia de los hijos de los comportamientos de apareamiento de sus padres permitiría a las hembras producir hijos atractivos al aparearse con machos que pueden adquirir varias parejas en lugar de un solo socio social.
La hipótesis del hijo sano
La hipótesis del hijo sexy no es la única explicación adaptativa para la elección de pareja que se centra en un posible beneficio hereditario del comportamiento. Por ejemplo, una hembra selecta podría obtener esperma de un macho hereditario sano cuyos genes, cuando se transmiten a su descendencia, los ayudará a desarrollar un sistema inmunológico fuerte. Las hembras que tomaron decisiones de pareja basadas en señales de salud masculina podrían dar a su progenie un beneficio genético. Del mismo modo, cuando la coloración del plumaje masculino refleja el contenido de carotenoides en los alimentos consumidos, una preferencia femenina por rojos brillantes o amarillos en parejas potenciales podría llevar a las hembras a aparearse con aves que son buenas recolectoras. Si esta capacidad tiene un componente hereditario, las hembras selectivas podrían asegurar parejas con genes que promueven el hábil forrajeo en sus descendientes.
Algunas de las pruebas más claras de los beneficios genéticos provienen de especies en las que los machos no brindan atención parental después de aparearse con la hembra. Las hembras en estas especies a menudo son muy selectivas sobre qué machos escogen como parejas, probablemente para obtener los mejores genes para su descendencia. En el gallo lira común (Tetrao tetrix) de Europa, por ejemplo, las hembras observan atentamente a muchos machos en exhibición antes de elegir uno para fertilizar sus huevos. El mismo macho a menudo es elegido por varias hembras, mientras que la mayoría de los machos en el área de exhibición nunca son seleccionados por ninguna hembra visitante. Los machos con más éxito reproductivo son aquellos que dominan a otros machos y evitan que los adornos de su cola se dañen en las peleas. Por lo general, también son los machos que ocupan los territorios de exhibición central, un logro que a menudo requiere años de intensa competencia.

Figura 7.2. La agachadiza real (Gallinago media) es un ave migratoria que se encuentraen Eurasia, desde Europa hasta Asia. Con su plumaje camuflado y un cuerpo delgado, estas aves son expertas en esconderse en hábitats de humedales y pastizales. La agachadiza real es conocida por su distintivo comportamiento de agacharse y desplazarse rápidamente, lo que dificulta su detección. Además, emiten un llamado característico durante su vuelo migratorio. Se alimentan principalmente de insectos, gusanos y pequeños invertebrados que encuentran en el suelo. La agachadiza real es un símbolo de los entornos húmedos y su presencia añade una nota de misterio y belleza a los paisajes naturales.
La preferencia femenina por el gallo del territorio central sugiere fuertemente que obtengan buenos genes para su descendencia de estos machos, genes que promueven el desarrollo de descendencia atractiva y / o saludable. En la agachadiza real (Gallinago media), una ave costera de Europa y el norte de Asia, los machos elegidos con mayor frecuencia por las hembras tienen una respuesta inmunitaria más fuerte que sus vecinos menos exitosos, lo que sugiere que se encuentran en la mejor condición corporal general. Una vez más, este resultado sugiere un beneficio indirecto (genético) de la elección de pareja, aunque el hallazgo no descarta absolutamente un beneficio directo (material) en forma de un riesgo reducido de adquirir una enfermedad contagiosa o parásito de las parejas sexuales preferidas.
Con respecto a los beneficios para los machos de la elección de pareja, recuerde que, como se describió anteriormente en este capítulo, los herrerillos comunes masculinos (Cyanistes caeruleus) prefieren las hembras con coronas con reflejos UV relativamente intensas. En un estudio, las hembras de este tipo sobrevivieron mejor que aquellas con menos reflectancia UV en sus plumas. Además, los herrerillos comunes femeninos con un plumaje amarillo más brillante (basado en carotenoides) produjeron garras más grandes y tuvieron mayor éxito en el desarrollo que las hembras con una coloración amarilla más opaca. En la medida en que los machos pueden elegir entre hembras brillantes y opacas como parejas, este estudio sugiere que una preferencia masculina por hembras más brillantes se traduciría en más progenie para el macho. Esta conclusión también puede aplicarse a el gorrión chillón (Petronia petronia), en el cual las hembras con un parche de pecho amarillo más grande pesaron más y tuvieron más crías por año que las hembras con un parche amarillo naturalmente más pequeño. Las preferencias de pareja masculina, como la selección de pareja por parte de las hembras, parecen tener el potencial de aumentar el éxito reproductivo de individuos selectos.
Induciendo la responsabilidad en el macho
Además, al igual que las hembras pueden ejercer un tipo de elección de pareja después de emparejadas con un macho inferior modificando su inversión en descendencia dependiendo de la calidad de su pareja, también algunas aves macho parecen ajustar su inversión en el cuidado parental en relación con la calidad de su compañera Por ejemplo, la variación en el color de los huevos depositados por las hembras sin manchas (Sturnus unicolor) en España afecta las decisiones masculinas sobre el cuidado de la descendencia resultante.

Figura 7.3. El estornino negro (Sturnus unicolor) es una especie de ave que se encuentra en gran parte de Europa y Asia. A diferencia de su pariente el estornino europeo, el estornino negro tiene un plumaje completamente negro y un brillo metálico en el sol. Estas aves son conocidas por su capacidad para formar grandes bandadas y realizar impresionantes exhibiciones de vuelo sincronizado. Además, poseen un canto melodioso y pueden imitar una amplia variedad de sonidos. Se alimentan de insectos, frutas y semillas, y su presencia en los paisajes naturales añade un toque de elegancia y encanto. El estornino negro es un espectáculo cautivador y una adición bienvenida a la avifauna local.
Los estorninos hembras a los que se les ha quitado algunas plumas de las alas, lo que reduce su condición haciéndoles más difícil volar, ponen huevos azules más pálidos que las aves de control con un complemento completo de primarias. Los machos emparejados con hembras que pusieron huevos pálidos eran padres menos activos cuando comenzaron a aprovisionar sus crías. Se han documentado respuestas masculinas similares al color del huevo en el pájaro gato gris (Dumetella carolinensis), una especie de América del Norte que también pone huevos azules. Las hembras en estas y otras especies probablemente estén bajo presión para proporcionar señales que alienten a sus compañeros sociales a trabajar arduamente para su progenie.

Figura 7.4. El pájaro gato gris (Dumetella carolinensis), también conocido como zorzal o mirlo gris, es una especie de ave nativa de América del Norte. A pesar de su nombre, no tiene ninguna relación cercana con los gatos. Su plumaje presenta tonos grises y negros, con un distintivo parche blanco en sus alas. Estas aves son conocidas por su canto melodioso y variado, que incluye una mezcla de trinos y gorjeos. Se alimentan principalmente de insectos, bayas y frutas. El pájaro gato gris es adaptable y se encuentra en diversos hábitats, desde bosques y áreas suburbanas hasta jardines y parques. Su presencia añade una nota musical y una belleza natural a los paisajes donde se encuentran.
Beneficios directos
En el ámbito de las decisiones de apareamiento, los beneficios directos son aquellos que devuelven una ventaja directamente al individuo selecto.
Alimentación
Por ejemplo, las hembras pueden preferir los machos que les dan mayores regalos nutricionales a través de la alimentación de cortejo. En el caso de la gaviota patiamarilla (Larus michahellis) estudiada en islas en el noroeste de España, las hembras que habían sido bien alimentadas consistentemente por su compañero de cortejo tenían menos probabilidades de moverse durante la copulación, por lo que era más probable que el macho transfiriera esperma durante el coito y, por lo tanto, era más probable que engendraran las crías de esa hembra. Al hacer que la cópula efectiva dependiera de la capacidad de su pareja para proporcionarle alimentos, una hembra podría usar la elección de pareja para obtener un beneficio material, a saber, las calorías y los nutrientes necesarios para mejorar su condición personal o su capacidad para producir descendencia.

Figura 7.5. La gaviota patiamarilla (Larus michahellis) es una especie de gaviota que se encuentra en las costas del Mediterráneo, Europa occidental y algunas partes de África. Estas gaviotas se caracterizan por su tamaño mediano, su plumaje predominantemente blanco y sus patas amarillas distintivas. Son aves marinas muy adaptables que pueden encontrarse tanto en áreas costeras como en entornos urbanos, donde aprovechan los desechos alimentarios y los vertederos como fuente de alimento. Son conocidas por su llamativo llamado y su habilidad para volar en busca de comida. La presencia de gaviotas patiamarillas añade un encanto costero a los paisajes marinos y ciudades costeras.
Del mismo modo, en Columbia Británica, Canadá, la tasa a la que las águilas pescadoras (Pandion haliaetus) proporcionan peces a sus compañeros durante la fase de cortejo se relaciona con la probabilidad de que la hembra se deje fecundar. Además, la frecuencia de alimentación de cortejo por parte del macho, que es una función de su capacidad de captura de peces, se correlaciona con la velocidad a la que luego aprovisiona a su descendencia. Por lo tanto, al evaluar la naturaleza de la alimentación de cortejo de un macho, las águilas hembra pueden obtener un forrajero competente como socio en la alimentación de su descendencia. Además, es probable que un forrajero altamente exitoso tenga buena salud en general y, por lo tanto, sea poco probable que transmita enfermedades contagiosas y debilitante a su pareja o su descendencia.
Calidad de plumaje y salud
Dependiendo de la especie de ave, la apariencia de un macho, la calidad de sus ornamentos de plumaje, el brillo de los rojos y amarillos en sus plumas, la intensidad de sus exhibiciones de comportamiento, la cantidad de rocas grandes que lleva a su compañero, la naturaleza de sus canciones, e incluso el tamaño del nido que construye para su compañero,

Figura 7.6. El cernícalo americano (Falco sparverius) es una especie de halcón pequeño y colorido que se encuentra en América del Norte y partes de América Central y del Sur. Estas aves tienen un plumaje llamativo, con patrones de colores como el blanco, el negro, el gris y el óxido. Son expertos cazadores y se alimentan principalmente de pequeños roedores, aves, insectos y reptiles. Los cernícalos americanos son conocidos por su vuelo rápido y ágil, así como por su capacidad para mantenerse en el aire en un solo lugar, conocido como "planeo". Su presencia en los paisajes naturales y áreas urbanas añade un toque de majestuosidad y control aéreo.
proporcionan indicadores potenciales de su condición fisiológica pasada, actual e incluso futura, y por lo tanto de su salud y capacidad para el cuidado de los pichones. Por ejemplo, en los cernícalos americanos (Falco sparverius) estudiados en Saskatchewan, Canadá, los varones con piel de color naranja brillante (producto de niveles altos de carotenoides) sobre la base del pico tienen menos probabilidades de tener infecciones de parásitos sanguíneos más adelante durante la época de reproducción, cuando el cuidado parental masculino se vuelve importante para la supervivencia de los hijos. Por lo tanto, una preferencia por los machos con este colorante ayudaría a las hembras a adquirir parejas capaces.

Figura 7.7. El pinzón doméstico (Haemorhous mexicanus), también conocido como pinzón mexicano o pinzón de casa, es una especie de ave nativa de América del Norte. Estos pinzones son reconocidos por su plumaje de tonos rojizos, con el macho exhibiendo un intenso color rojo en la cabeza, pecho y espalda. Son aves sociales y abundantes, y se pueden encontrar en una variedad de hábitats, desde bosques y praderas hasta áreas urbanas y jardines. Su alimentación consiste principalmente en semillas, pero también consumen insectos y frutas. El pinzón doméstico es apreciado por su hermoso canto y su presencia alegre y animada en los paisajes naturales y humanizados.
Del mismo modo, en un experimento en el que se inocularon machos pinzones (Haemorhous mexicanus) con un patógeno bacteriano infeccioso, las aves con plumaje más rojo se recuperaron más rápidamente que las aves con plumas más pálidas y amarillas. Debido a que la elección de pareja femenina en esta especie está sesgada a favor de los machos más rojos, las hembras podrían emparejarse con los machos capaces de ser mejores padres porque eran resistentes a ciertas enfermedades.
Calidad del canto y de crianza previa
La calidad de la canción de un macho a veces sirve como otro indicador potencial de su capacidad paterna. En este contexto, es sorprendente que la capacidad de un macho para aprender canciones a temprana edad a menudo sea muy sensible a las condiciones que se experimentan en ese momento. Por ejemplo, los pichones que reciben relativamente poca comida podrían verse perjudicados cuando se trata de imitar las canciones de sus especies. Como resultado, una hembra que escucha a un adulto cantar puede aprender algo sobre su historia pasada de desarrollo, lo que a su vez podría influir en su salud actual y en su capacidad para tener un comportamiento paternal competente. En este sentido, una serie de estudios de laboratorio cuidadosos han demostrado que los machos del gorrión cantor (Melospiza melodia) que están en buenas condiciones aprenden y copian las canciones que escuchan con mayor precisión que los gorriones jóvenes que están en malas condiciones.
Estas mejores canciones proporcionan más adelante información para el apareamiento de hembras: cuando las hembras adultas que reciben una dosis de hormonas para que sean sexualmente sensibles escuchan las canciones grabadas, es más probable que eleven sus colas a la posición precopulatoria cuando escuchan copias precisas de la canción de su especie que cuando escuchan las imitaciones más pobres hechas por los machos desfavorecidos. Este tipo de preferencia podría llevar a la hembra a aparearse y unirse con un macho mejor que el promedio, uno con una buena copia de la canción de su especie, que se correlaciona con su capacidad para brindar una asistencia mejor que el promedio a la hembra y su cría porque fue bien alimentado cuando fue un pichón.

Figura 7.8. El gorrión cantor (Melospiza melodía) es una especie de ave paseriforme nativa de América del Norte. Estos gorriones son conocidos por su distintivo canto melódico, compuesto por una variedad de trinos y notas musicales. Tienen un plumaje pardo con rayas en el pecho y una mancha blanca en la garganta. Son aves migratorias y se pueden encontrar en una amplia variedad de hábitats, desde praderas y bosques hasta arbustos densos y áreas urbanas. Se alimentan principalmente de semillas e insectos. El gorrión cantor es apreciado por su encantador canto y su presencia vibrante y animada en los paisajes naturales y los jardines. (YouTube)
Selección sexual
La elección de pareja, ya sea por hembras o por machos, tiene consecuencias evolutivas si las diferencias entre los individuos sujetos a elección son hereditarias. Los individuos con los atributos hereditarios favorecidos pueden experimentar un mayor éxito reproductivo que otros de su especie. Con el tiempo, la base genética de la capacidad de cantar un tipo de canción en particular, o de exhibir un adorno en particular, o de participar en ciertas exhibiciones de cortejo, o de suministrar a los compañeros sus alimentos favoritos puede propagarse a través de la especie.
(a) Los rasgos sexualmente atractivos generalmente tienen costos:
(b) las canciones pueden atraer tanto a los depredadores como a los compañeros, los adornos hacen que las personas sean más conspicuas para sus enemigos,
los animales enfocados en mostrar a los socios potenciales están menos alertas al peligro,
(c) y los individuos que ofrecen comida a sus compañeros pasan un tiempo valioso y energía para recoger lo que luego regalan.
Charles Darwin se dio cuenta de que tales rasgos reductores de la supervivencia requerían una explicación especial, que proporcionó en la forma de su teoría de la selección sexual. Argumentó que los ornamentos elaborados, por ejemplo, podrían poner a un macho en un riesgo especial de ataque por parte de los depredadores y, por lo tanto, podría pensarse que está inadaptado, pero si estos ornamentos también aumentan las probabilidades de que un macho adquiera un compañero y se reproduzca con éxito, entonces los efectos sexuales se imponen sobre los de las demás interacciones ecológicas de la selección sexual, esto se refuerza más en los machos al ser el sexo desechable, pues un macho puede fertilizar a muchas hembras, por lo que una especie con pocos machos, pero muy hábiles puede sobrevivir. El mismo argumento se puede aplicar a cualquier atributo que parezca funcionar estrictamente en términos de la competencia entre parejas, ya sean características o rasgos sexualmente atractivos que permitan a los individuos prevalecer sobre otros del mismo sexo en luchas relacionadas con el apareamiento.
La selección sexual ahora se considera generalmente como un componente de la selección natural, en el sentido de que las condiciones requeridas para ambas formas de selección son fundamentalmente similares. Tanto la selección natural como la sexual actúan sobre la variación hereditaria entre individuos en su capacidad para transmitir sus genes, pero la selección sexual lo hace en el contexto estrecho de la competencia por el acceso a las parejas. Sin embargo, la teoría de la selección sexual es valiosa porque ayuda a centrar la atención en el significado de la competencia reproductiva pura. Uno de los beneficios de esta perspectiva es que los investigadores que están alerta a la teoría pueden identificar los rompecabezas que, de otro modo, podrían ignorar, particularmente cuando los machos o las hembras parecen comportarse de maneras que a primera vista parecen reducir sus posibilidades de sobrevivir en primera instancia, o de autolimitación en la fertilización de parejas potenciales secundarias. Como se mencionó anteriormente en el capítulo, la monogamia por parte de los machos es un ejemplo de este tipo de rompecabezas, porque el macho verdaderamente monógamo parece renunciar a las oportunidades de aparearse con más de una hembra.
El fugitivo loco de Fisher
También llamada selección descontrolada es un mecanismo de selección sexual propuesto por el biólogo matemático Ronald Fisher a principios del siglo XX para explicar la evolución de la ornamentación ostentosa en los machos a través de la persistente elección direccional de las hembras. Un ejemplo es el llamativo y elaborado plumaje del pavo real en comparación con el plumaje relativamente discreto de la pava; los costosos ornamentos, como la cola extremadamente larga del ave, parecen ser incompatibles con la selección natural. Se postula que la selección descontrolada incluye rasgos fenotípicos dimórficos sexualmente, como el comportamiento expresado por un sexo en particular.
El dimorfismo sexual extremo y (aparentemente) maladaptativo representaba una paradoja para los biólogos evolutivos desde la época de Charles Darwin hasta la síntesis moderna. Darwin intentó resolver la paradoja asumiendo la herencia tanto de la preferencia como del ornamento, y supuso un "sentido estético" en los animales superiores, lo que condujo a una selección poderosa de ambas características en las generaciones siguientes. Fisher desarrolló aún más la teoría al asumir una correlación genética entre la preferencia y el ornamento, que inicialmente el ornamento señalaba un mayor potencial de aptitud (la probabilidad de dejar más descendencia), por lo que la preferencia por el ornamento tenía una ventaja selectiva. Posteriormente, si la preferencia femenina por la ornamentación exagerada en la selección de pareja era lo suficientemente fuerte, podría ser suficiente para socavar la selección natural incluso cuando el ornamento se volviera no adaptativo. A lo largo de las generaciones posteriores, esto podría llevar a una selección descontrolada mediante retroalimentación positiva, y la velocidad con la que el rasgo y la preferencia aumentan podría aumentar exponencialmente hasta que la contraselección interfiera.
La descripción moderna del mismo mecanismo utilizando modelos genéticos cuantitativos y poblacionales fue establecida principalmente por Russell Lande y Mark Kirkpatrick en la década de 1980, y ahora se conoce más comúnmente como la hipótesis del hijo sexy (Pomiankowski & Iwasa, 1998).
8. Desarrollo embrionario en el pollo
8. Desarrollo embrionario en el pollo
Desde que Aristóteles siguió por primera vez su desarrollo de 3 semanas, el pollo doméstico ha sido un organismo favorito para los estudios embriológicos. Es accesible todo el año y se cría fácilmente. Además, a cualquier temperatura particular, su etapa de desarrollo puede predecirse con precisión. Así, se pueden obtener grandes cantidades de embriones en la misma etapa. El embrión de pollo se puede manipular quirúrgicamente y, dado que forma la mayoría de sus órganos de manera muy similar a la de los mamíferos, a menudo ha servido como sustituto de los embriones humanos.

Figura 8.1. Escisión meroblastica discoidal en un huevo de gallina. (A-D) Cuatro etapas vistas desde el polo animal (el lado dorsal futuro del embrión). (E) Un embrión de escisión temprana visto desde el lado.
La fertilización del huevo de gallina ocurre en el oviducto, antes de que la albúmina y la cáscara se segreguen sobre él. El huevo es telolecítico (como el del pescado), con un pequeño disco de citoplasma sentado sobre una gran yema.
Clivajes
Al igual que los huevos de pescado, los huevos de yema de las aves se someten a escisión meroblastica discoidal “La división solo afecta a un disco citoplasmático próximo al polo animal y la parte inferior (polo vegetativo) no se segmenta; a continuación, se forma un disco de blastómeros, el blastodermo o blastodisco, a partir del cual se formará el embrión, que reposará sobre la masa vitelina. Se da en peces, reptiles y aves, entre otros”. La escisión se produce solo en el blastodisco, un pequeño disco de citoplasma de 2 a 3 mm de diámetro en el polo animal de la célula del huevo. El primer surco de escisión aparece centralmente en el blastodisco, y otras escisiones siguen para crear un blastodermo de una sola capa (Figura 8.1). Al igual que en el embrión de pescado, estas escisiones no se extienden en el citoplasma de la yema, por lo que las células de escisión temprana son continuas entre sí y con la yema en sus bases (Figura 8.2-E).
Posteriormente, las divisiones ecuatoriales y verticales dividen el blastodermo en un tejido de cinco a seis capas de células de espesor. Estas células se unen entre sí mediante uniones estrechas.

Figura 8.2. Formación del blastodermo de dos capas del embrión de pollo. (A, B) Las células primarias de hipoblasto se deslaminan individualmente para formar islas de células debajo del epiblasto. (C) Las células de hipoblasto secundarias del margen posterior (la hoz de Koller y las células marginales posteriores detrás de él) migran debajo del epiblasto e incorporan las islas de polinvaginación. A medida que el hipoblasto se mueve anteriormente, las células epiblastas se acumulan en la región anterior a la hoz de Koller para formar la veta primitiva. (D) Esta sección sagital de un embrión cerca del margen posterior muestra una capa superior que consiste en un epiblasto central que se introduce en las células de la hoz de Koller (ks) y la zona marginal posterior (mz). Ciertas células se deslaminaron del epiblasto (ep) para formar islas de polinvaginación (pi) de 5 a 20 células cada una. Estas células se unirán a aquellas células hipoblastas (hip) que migran anteriormente de la hoz de Koller para formar la capa inferior (hipoblástica secundaria). (sc, cavidad subgerminal; gwm, margen de la pared germinal.)
Gastrulación
Hipoblasto
Cuando una gallina ha puesto un huevo, el blastodermo contiene unas 20,000 células. En este momento, la mayoría de las células de la zona pelúcida permanecen en la superficie, formando el epiblasto, mientras que otras células de la pelúcida área se deslaminaron y migraron individualmente hacia la cavidad subgerminal para formar las islas de polinvaginación (hipoblasto primario), un archipiélago de agrupaciones desconectadas que contiene 5-20 células cada una (Figura 8.2-B). Poco después, una lámina de células del margen posterior del blastodermo (que se distingue de las otras regiones del margen por la hoz de Koller, un engrosamiento local) migra anteriormente para unirse a las islas de polinvaginación, formando así el hipoblasto secundario.

Figura 8.3. Movimientos celulares de la línea primitiva del embrión de pollo. (A-C) Vista dorsal de la formación y elongación de la veta primitiva. El blastodermo se observa en (A) 3-4 horas, (B) 7-8 horas y (C) 15–16 horas después de la fertilización. Los primeros movimientos de las células epiblastas que migran se muestran mediante flechas. (D-F) Formación de somitas de notocordio y mesodérmicas a medida que la raya primitiva retrocede, mostrada en (F) 19–22 horas, (E) 23–24 horas, y (F) la etapa de cuatro somitas. Los mapas de destino del epiblasto de pollo se muestran para dos etapas, la etapa de raya primitiva definitiva (C) y la neurulación (F). El endodermo ya ha penetrado debajo del epiblasto, y se observa una extensión convergente en la línea media.
El blastodermo de dos capas (epiblasto e hipoblasto) se une en el margen del área de opaca, y el espacio entre las capas forma un blastoceloma. Por lo tanto, aunque la forma y la formación del blastodisco aviar difieren de las de la blástula de anfibios, peces o equinodermos, se mantienen las relaciones espaciales generales.
El embrión aviar proviene enteramente del epiblasto. El hipoblasto no contribuye con ninguna célula al embrión en desarrollo. Más bien, las células hipoblastas forman porciones de las membranas externas, especialmente el saco vitelino y el tallo que une la placa de la yema al tubo digestivo endodérmico. Las tres capas germinales del embrión propiamente dicho (más una cantidad considerable de membrana extraembriónica) se forman a partir de las células epiblastas. Los mapas de destino del epiblasto de pollo se muestran en (Figura 8.3).
La principal característica estructural de la gastrulación aviaria, reptiliana y de mamíferos es el tallo primitivo. Esta raya se ve primero como un engrosamiento del epiblasto en la región posterior del embrión “hacia el dorso”, justo delante de la hoz de Koller (Figura 8.3-A). Este engrosamiento es causado por la entrada de precursores endodérmicos desde el epiblasto al blastoceloma y por la migración de células desde la región lateral del epiblasto posterior hacia el centro (Figura 8.3-B). Cuando estas células entran en la línea primitiva, la línea se alarga hacia la región de la cabeza futura. Al mismo tiempo, las células de hipoblasto secundarias continúan migrando anteriormente del margen posterior del blastodermo. El alargamiento del tallo primitivo parece ser coextensivo con la migración anterior de estas células de hipoblasto secundario. El tallo eventualmente se extiende 60–75% de la longitud del área pelúcida.
El tallo primitivo define los ejes del embrión. Se extiende desde la parte posterior a la anterior; las células migratorias entran a través de su lado dorsal y se mueven hacia su lado ventral; y separa la parte izquierda del embrión de la derecha. Los elementos cercanos al tallo serán las estructuras mediales (centrales), mientras que los más alejados serán las estructuras distales (laterales) (Figura 8.3-C, E).
Cuando las células convergen para formar el tallo primitivo, se forma una depresión dentro del propio tallo. Esta depresión se denomina surco primitivo y sirve como una abertura a través de la cual las células migratorias pasan al blastoceloma. Por lo tanto, el surco primitivo es análogo al blastoporo de anfibios. En el extremo anterior del tallo primitivo hay un engrosamiento regional de las células llamado nudo primitivo o nodo de Hensen. El centro de este nodo contiene una depresión en forma de embudo (a veces llamada fosa primitiva) a través de la cual las células pueden pasar al blastoceloma. El nodo de Hensen es el equivalente funcional del labio dorsal del blastoporo de anfibios (es decir, el organizador) y el escudo embrionario de los peces.

Figura 8.4. Migración de células endodérmicas y mesodérmicas a través del tallo primitivo. (A) La micrografía electrónica de barrido muestra que las células epiblastas pasan al blastoceloma y extienden sus extremos apicales para convertirse en células de botella. (B) Estereograma de un embrión de pollo gástrico, que muestra la relación de la línea primitiva, las células migratorias y las dos capas originales del blastodermo. La capa inferior se convierte en un mosaico de células hipoblastos y endodérmicas; Las células de hipoblasto finalmente se separan para formar una capa debajo del endodermo y contribuyen al saco vitelino.
Tan pronto como el tallo primitivo se ha formado, las células epiblastas comienzan a migrar a través de ella y hacia el blastoceloma (Figura 8.4). El tallo primitivo tiene una población celular en continuo cambio. Las células que migran a través del nodo de Hensen pasan al blastoceloma y migran en la parte anterior, formando las vísceras superiores, cabeza del mesodermo y notocorda; Las células que pasan a través de las porciones laterales de la línea primitiva dan lugar a la mayoría de los tejidos endodérmicos y mesodérmicos. A diferencia del mesodermo de Xenopus, que migra como láminas de células al blastoceloma, las células que ingresan en el interior del embrión aviar ingresan como individuos después de sufrir una transformación epitelial a mesenquimatosa. Se cree que la descomposición de la lámina basal y la liberación de estas células en el embrión se llevan a cabo en el nodo de Hensen a lo largo del tallo primitivo por un factor de dispersión, una proteína de 190 kDa segregada por las células cuando entran en la hendidura. El factor de dispersión puede convertir las láminas epiteliales en células mesenquimales de varias maneras, y es probable que esté involucrado tanto en la disminución de la expresión de E-cadherina como en la prevención del funcionamiento de la E-cadherina.
Formación del mesodermo y endodermo
Las primeras células que migran a través del nodo de Hensen son aquellas destinadas a convertirse en el endodermo faríngeo de las vísceras superiores. Una vez dentro del blastoceloma, estas células endodérmicas migran anteriormente y eventualmente desplazan a las células hipoblastas, lo que hace que las células hipoblastas se confinen a una región en la porción anterior del área pelúcida. Esta región, la media luna germinal, no forma ninguna estructura embrionaria, pero contiene los precursores de las células germinales, que luego migran a través de los vasos sanguíneos a las gónadas. Las siguientes células que entran en el blastoceloma a través del nódulo de Hensen también se mueven en sentido anterior, pero no se mueven tan ventralmente como las células endodérmicas presuntivas. Más bien, permanecen entre el endodermo y el epiblasto para formar la cabeza del mesénquima y el mesodermo de la placa precordena. Estas células de ingreso temprano se mueven en sentido anterior, empujando hacia arriba la región de la línea media anterior del epiblasto para formar el proceso de la cabeza (Figura 8.5). Así, la cabeza del embrión aviar se forma anterior (rostral) al nodo de Hensen. Las siguientes células que migran a través del nodo de Hensen se convierten en células de acordesodermo (notocordas). Estas células se extienden hasta el presunto cerebro medio, donde se encuentran con la placa precordal. El cerebro posterior y el tronco se forman a partir del cordamesodermo a nivel del nodo de Hensen y caudal a él.
Mientras tanto, las células continúan migrando hacia adentro a través de la porción lateral del tallo primitivo. Cuando entran en el blastoceloma, estas células se separan en dos capas. La capa profunda se une al hipoblasto a lo largo de su línea media y desplaza las células del hipoblasto a los lados. Estas células de movimiento profundo dan lugar a todos los órganos endodérmicos del embrión, así como a la mayoría de las membranas extraembrionarias (el hipoblasto forma el resto). La segunda capa migratoria se extiende entre este endodermo y el epiblasto, formando una capa suelta de células. Estas células de la capa media generan las porciones mesodérmicas de las membranas embrionarias y extraembrionarias. A las 22 horas de incubación, la mayoría de las células endodérmicas presuntas se encuentran en el interior del embrión, aunque las células mesodérmicas presuntas continúan migrando hacia adentro durante más tiempo.
Regresión del tallo
Ahora comienza una nueva fase de la gastrulación. Mientras continúa la ingresión mesodérmica, el tallo primitivocomienza a retroceder, moviendo el nodo de Hensen desde cerca del centro de la pelícida del área a una posición más posterior (ver Figura 8.5). En su retirada el tallo deja a su paso el eje dorsal del embrión y la notocorda. A medida que el nodo se mueve hacia atrás, la notocorda se establece, comenzando en el nivel del futuro cerebro medio. Mientras que la parte anterior de la notocorda se forma por la ingresión de células a través del nódulo de Hensen, la notocorda posterior (después de la somita 17 en el pollo) se forma a partir de la condensación del tejido mesodérmico que ha ingerido a través de la línea primitiva (es decir, no a través del nodo de Hensen). Esta parte de la notocorda se extiende posteriormente para formar la cola del embrión. Finalmente, el nodo de Hensen regresa a su posición más posterior, formando la región anal. En este momento, todas las células endodérmicas y mesodérmicas presuntas han ingresado en el embrión, y el epiblasto está compuesto completamente de células ectodérmicas presuntas.

Figura 8.5. La gastrulación de los pollitos de aproximadamente 24 a 28 horas. (A) El tallo primitivo en plena extensión (24 horas). El proceso de la cabeza (notocorda anterior) se puede ver extendiéndose desde el nodo de Hensen. (B) Etapa de dos somitas (25 horas). El endodermo faríngeo se ve anteriormente, mientras que la notocorda anterior empuja hacia arriba el proceso de la cabeza por debajo. El tallo primitivo está retrocediendo. (C) Etapa cuatro-somitas (27 horas). (D) A las 28 horas, el tallo primitivo ha regresado a la porción caudal del embrión. (E) Regresión del tallo primitivo, dejando el notocordio a su paso. Varios puntos de la racha (representados por letras) fueron seguidos después de que alcanzó su longitud máxima. El tiempo representa horas después de alcanzar la longitud máxima (la línea de referencia, aproximadamente 18 horas después de la incubación)
Como consecuencia de la secuencia mediante la cual se establecen el mesodermo de la cabeza y la notocorda, los embriones aviares (y mamíferos) exhiben un claro gradiente anterior-posterior de la madurez del desarrollo. Mientras que las células de las porciones posteriores del embrión están experimentando gastrulación, las células en el extremo anterior ya están comenzando a formar órganos (la cabeza se forma primero y la cola después). Durante los próximos días, el extremo anterior del embrión está más avanzado en su desarrollo (después de haber tenido un "inicio" tardío) con respecto al extremo posterior.
Epibolia del ectodermo
Mientras que las células mesodérmicas y endodérmicas presuntas se están moviendo hacia adentro, los precursores ectodérmicos proliferan. Por otra parte, las células ectodérmicas migran para rodear la yema de huevo por epibolia. El encierro de la yema por el ectodermo (una vez más reminiscencia del epibolio del ectodermo de anfibios) es una tarea hercúlea que toma la mayor parte de los 4 días siguientes para completarse.
Implica la producción continua de nuevo material celular y la migración de las presuntas células ectodérmicas a lo largo del lado inferior de la envoltura vitelina. Curiosamente, solo las células del borde exterior del área de opaca se adhieren firmemente a la envoltura vitelina. Estas células son intrínsecamente diferentes de las otras células de blastodermo, ya que pueden extender enormes procesos citoplásmicos (500 μm) a la envoltura vitelina. Se cree que estos filopodios alargados son el aparato locomotor de estas células marginales, mediante el cual tiran de las otras células ectodérmicas alrededor de la yema. Los filopodios parecen unirse a la fibronectina, una proteína laminar que es un componente de la envoltura vitelina del pollo. Si el contacto entre las células marginales y la fibronectina se rompe experimentalmente (al agregar un polipéptido soluble similar a la fibronectina), la retracción de los filopodios y la migración epidérmica cesa.
Así, a medida que la gastrulación aviar se acerca a su fin, el ectodermo ha rodeado la yema, el endodermo ha reemplazado al hipoblasto y el mesodermo se ha posicionado entre estas dos regiones. Hemos identificado muchos de los procesos involucrados en la gastrulación aviar, pero seguimos ignorando los mecanismos por los cuales muchos de estos procesos se llevan a cabo.
Formación del eje anteroposterior
Mientras que la formación de los ejes corporales se logra durante la gastrulación, su especificación ocurre antes, durante la etapa de escisión.
Importancia de la acidez
El eje dorsal-ventral (parte posterior del vientre) es crítico para la formación del hipoblasto y para el desarrollo posterior del embrión. Este eje se establece cuando las células de escisión del blastodermo establecen una barrera entre la albúmina básica (pH 9.5) que se encuentra por encima del blastodisco y el espacio subgerminal ácido (pH 6.5) que se encuentra debajo. El agua y los iones de sodio se transportan desde la albúmina a través de las células hasta el espacio subgerminal, creando una diferencia de potencial de membrana de 25 mV a través de la capa de células epiblastas (positiva en el lado ventral de las células). Este proceso distingue dos lados del epiblasto: un lado que mira a la albúmina negativa y básica, que se convierte en el lado dorsal, y un lado que mira al fluido del espacio subgerminal positivo y ácido, que se convierte en el lado ventral. Este eje se puede invertir experimentalmente invirtiendo el gradiente de pH o invirtiendo la diferencia de potencial a través de la capa celular.
Importancia de la gravedad
La conversión del blastodermo radialmente simétrico en una estructura bilateralmente simétrica está determinada por la gravedad. A medida que el óvulo atraviesa el tracto reproductivo de la gallina, gira alrededor de 20 horas en la glándula de la concha. Esta hilatura, a una velocidad de 10 a 12 revoluciones por hora, desplaza la yema de modo que sus componentes más ligeros se encuentran debajo de un lado del blastodermo. Esto alza ese extremo del blastodermo, y ese extremo se convierte en la parte posterior del embrión, la parte donde comienza la formación del tallo primitivo.

Figura 8.6. Organización anterior-posterior por gravedad del embrión del pollo. La rotación en la glándula de la cáscara (A) da como resultado que los componentes más ligeros de la yema empujen hacia arriba a un lado del blastodermo (B). Esa región más elevada se convierte en la parte posterior del embrión (C).
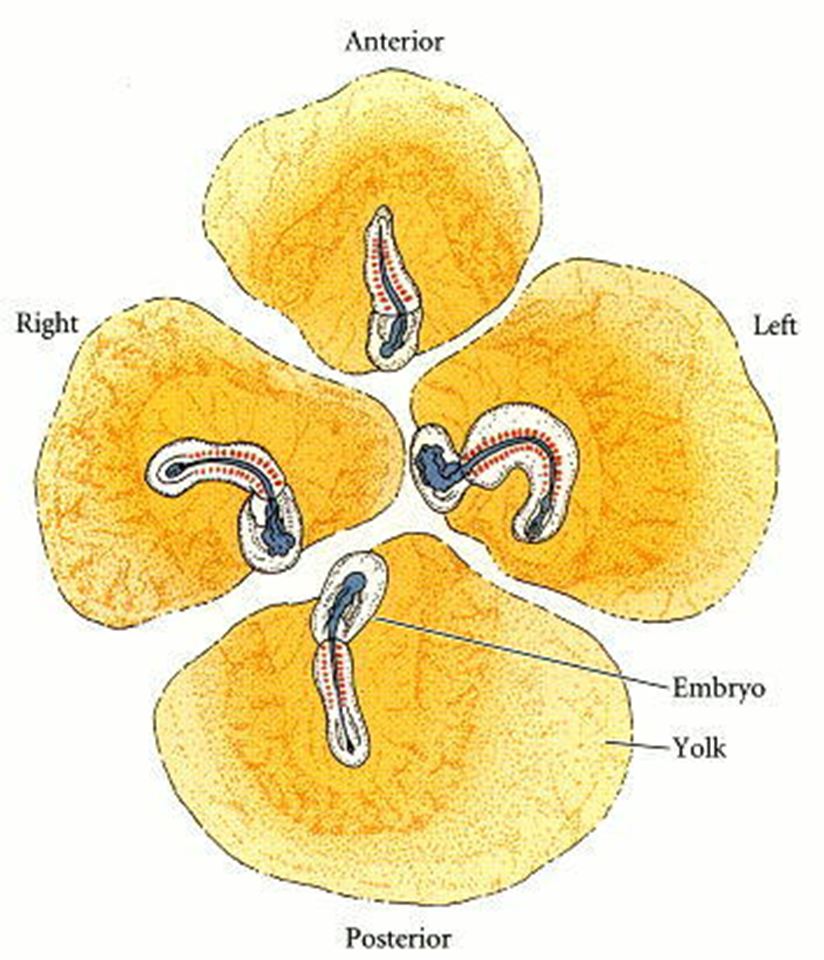
Figura 8.7. Regulación del blastodermo de pollo. Cuando el blastodermo se divide en cuatro partes, cada parte puede iniciar la gastrulación y dar lugar a un embrión (clonación embrionaria). Por lo general, solo las células de la zona marginal posterior pueden formar una raya primitiva e inhiben a otras áreas de la zona marginal para que no lo hagan.
No se sabe qué interacciones hacen que esta porción del blastodermo se convierta en el margen posterior e inicie la gastrulación. La capacidad de iniciar el tallo primitivo se encuentra en toda la zona marginal, y si el blastodermo se separa en partes, cada una con su propia zona marginal, cada parte formará su propio tallo primitivo (Figura 8.7). Sin embargo, una vez que se ha formado una zona marginal posterior (PMZ por sus siglas en inglés), controla las otras regiones del margen. Estas células PMZ no solo inician la gastrulación, sino que también evitan que otras regiones del margen formen sus propios tallos primitivos.
Ahora parece evidente que la PMZ contiene células que actúan como el equivalente del centro Nieuwkoop de anfibios. Cuando se coloca en la región anterior de la zona marginal, un injerto de tejido de la zona marginal posterior (posterior y sin incluir la hoz de Koller) puede inducir una línea primitiva y un nodo de Hensen sin que las células contribuyan a ninguna de las estructuras. Al igual que el centro Nieuwkoop de anfibios, se cree que esta región es el lugar donde la localización de la β-catenina en el núcleo y la señal de la familia TGF-β coinciden. La zona marginal posterior es la única región del Vg1 secretor de embriones.
Las células de epiblasto y capa media en la porción anterior de la hoz de Koller se convierten en el nodo de Hensen. La parte posterior de la hoz de Koller contribuye a la parte posterior de la línea primitiva (Figura 8.8).

Figura 8.8. Formación del nodo de Hensen a partir de la hoz de Koller. (A) Diagrama del extremo posterior de un embrión temprano (antes de la raya), que muestra las células marcadas con tintes fluorescentes en las fotografías. (B) Justo antes de la gastrulación, las células en el extremo anterior de la hoz de Koller (el epiblasto y la capa media) se marcaron con colorante verde. Las células de la porción posterior de la hoz de Koller se marcaron con tinte rojo. A medida que las células migran, las células anteriores formaron el nodo de Hensen y sus derivados de notocordas. Las células posteriores formaron la región posterior de la línea primitiva. El tiempo después de la inyección de tinte está etiquetado en cada fotografía.
Desde hace tiempo se sabe que el nodo de Hensen es el equivalente aviar del labio blastoporo dorsal de anfibios, ya que es (1) el sitio donde comienza la gastrulación, (2) la región cuyas células se convierten en el cordamesodermo, y (3) la región cuyas células pueden organice un segundo eje embrionario cuando se trasplanta en otras ubicaciones de la gástrula. Además, el trasplante de la hoz de Koller también puede causar la formación de nuevos ejes, y las células de la capa media en la hoz de Koller expresan goosecoides, al igual que las células del organizador de Spemann.
Como es el caso en todos los vertebrados, el mesodermo dorsal es capaz de inducir la formación del sistema nervioso central en el ectodermo que lo recubre. Las células del nodo de Hensen y sus derivados actúan como el organizador de los anfibios, y secretan proteínas Cordina, Noggina y Nodal. Los factores de crecimiento de fibroblastos se producen en el nodo de Hensen y en la línea primitiva, y las perlas que contienen ciertos FGF pueden inducir la expresión neuronal del tronco y del cerebro posterior en las células epiblastas.

Figura 8.9. Para la asimetría izquierda-derecha en el embrión de pollo. (A) Lado izquierdo: en el lado izquierdo del nodo de Hensen, se expresan sonic hedgehog (shh) y lefty-1. Lefty-1 bloquea la expresión fgf8, mientras que sonic hedgehog activa la expresión de caronte. Mientras tanto, el ácido retinoico permite la expresión de Lefty-1 a lo largo de la línea media ventral. La expresión de Caronte bloquea las proteínas morfogenéticas óseas (BMP) en el lado izquierdo, que de otro modo bloquearían la expresión de nodal y Lefty-2. En presencia de Nodal y Lefty-2, el gen pitx2 se activa y el gen del caracol (cSnR) se reprime. Pitx2 está activo en los diversos primordios de órganos y especifica el lado que se dejará. En el lado derecho del embrión, la activina se expresa junto con el receptor de activina IIa. Esto activa FGF8, una proteína que bloquea la expresión de caronte. En ausencia de Caronte, BMP reprime la activación de nodal y lefty-2. Esto permite que el gen caracol esté activo mientras que el gen pitx2 permanece reprimido. (B, C) Vistas dorsales y en primer plano de la hibridación in situ del ARNm de sonic hedgehog. (D, E) Vistas dorsales y en primer plano del mensaje del receptor IIa de activina.
Formación del eje lateral
Como hemos visto, el cuerpo de los vertebrados no es simétrico. Más bien, tiene distintos lados derecho e izquierdo. El corazón y el bazo, por ejemplo, están generalmente en el lado izquierdo del cuerpo, mientras que el hígado está generalmente en el lado derecho. La distinción entre los lados derecho e izquierdo de los vertebrados está regulada por dos proteínas principales: el factor paracrino nodal y el factor de transcripción Pitx2. Sin embargo, el mecanismo por el cual la expresión génica nodal se activa en el lado izquierdo del cuerpo difiere entre las clases de vertebrados. La facilidad con que pueden manipularse los embriones de pollo ha permitido a los científicos dilucidar las vías de discriminación de izquierda a derecha en aves más fácilmente que en otros vertebrados.
A medida que la línea primitiva alcanza su longitud máxima, se genera una cascada de genes homeoticos diferencial entre la derecha y la izquierda. Esto permite generar los órganos asimétricos como el hígado, el páncreas, los pulmones y el corazón.
Organogénesis
El endodermo origina el epitelio de revestimiento interno de los sistemas digestivo, inmunológico y respiratorio, y los órganos pueden visualizarse desde el segundo día (48 h) cuando se forma el hígado. La formación del sistema digestivo se completó en el día 12. Los órganos del sistema respiratorio comienzan en el cuarto día como un tejido desorganizado e indiferenciado. Su diferenciación completa se observa a los 10 días de la incubación, sin embargo, hasta los 19 días no se observó la siringe. La formación del sistema inmune al décimo día se observó con la formación del bazo, el timo y la bolsa cloacal. El estudio de la organogénesis del pollo basado en capas germinales es muy complejo y está poco explorado, y el estudio de la embriología del pollo es muy importante debido a la importancia económica y al crecimiento del uso de este modelo animal, como los estudios genéticos (Alcântara et al., 2013).
9. Los huevos
9. Los huevos
Las aves son el único grupo principal de vertebrados sin al menos una especie vivípara: los peces, los anfibios, los reptiles y los mamíferos incluyen, al menos, algunas especies vivíparas.

Figura 9.1. Anatomía del huevo
(a) Cascara: De textura rugosa y granulada, una cáscara de huevo está cubierta con hasta 17,000 poros diminutos. La cáscara del huevo está hecha casi completamente de cristales de carbonato de calcio (CaCO3). Es una membrana semipermeable, lo que significa que el aire y la humedad pueden pasar a través de sus poros. La cáscara también tiene una capa externa más delgada llamada floración o cutícula que ayuda a mantener fuera las bacterias y el polvo.
(b) Membranas interna y externa: Situadas entre la cáscara del huevo y la clara de huevo “albumen”, estas dos membranas de proteínas transparentes proporcionan una defensa eficaz contra la invasión bacteriana. Si le das un tirón a estas capas, encontrarás que son sorprendentemente fuertes. Están hechos en parte de queratina, una proteína que también se encuentra en el cabello humano.
(c) Celda de aire: Un espacio de aire se forma cuando el contenido del huevo se enfría y se contrae después de que se pone el huevo. La celda de aire generalmente descansa entre las membranas externa e interna en el extremo más grande del huevo, y explica el cráter que se ve a menudo al final de un huevo duro. Crece a medida que un huevo envejece. Al comer un huevo tibio rompa por el lado romo.
(d) Albumina: La clara de huevo se conoce como la albúmina, que proviene de albus, la palabra latina para “blanco”. Cuatro capas alternas de albúmina gruesa y delgada contienen aproximadamente 40 proteínas diferentes, los componentes principales de la clara de huevo además del agua.
(e) Chalazas: Cuerdas opacas de clara de huevo, las chalazas sostienen la yema en el centro del huevo. Como pequeñas anclas, sujetan la envoltura de la yema a la membrana que recubre la cáscara del huevo. Cuanto más prominentes son, más fresco es el huevo.
Membrana de la yema: La clara envoltura que encierra la yema.
(f) La yema “yolk”: La yema contiene menos agua y más proteínas que la clara, algo de grasa y la mayoría de las vitaminas y minerales del huevo. Estos incluyen hierro, vitamina A, vitamina D, fósforo, calcio, tiamina y riboflavina. La yema es también una fuente de lecitina, un emulsionante eficaz. El color de la yema varía desde un toque de amarillo hasta un magnífico color naranja intenso, de acuerdo con el alimento y la raza de la gallina.
Exactamente por qué no han evolucionado las aves vivíparas sigue siendo un poco misterioso, pero una idea convincente es que tener crías vivas pesaría a las aves hembras durante la gestación y, por lo tanto, dificultan su capacidad de vuelo o limitan el tamaño de su cría. Sin embargo, el hecho de que las hembras murciélagos tengan éxito en dar a luz crías vivas mientras vuelan compitiendo durante todo su embarazo presenta un argumento en contra de esta hipótesis. La colocación de huevos externos también permite a ambos padres aviares al menos ayudar potencialmente en su cuidado, así como la oportunidad de abandonar los huevos si las condiciones o circunstancias son adversas, lo que brinda una oportunidad para que los padres sobrevivan.
Todos los huevos aviares tienen la función principal de proteger el embrión en el interior a medida que se desarrolla. Sin embargo, los huevos de varias especies de aves difieren ampliamente en tamaño y forma, coloración y estructura de la cáscara, y las proporciones relativas de albúmina y yema. Desde el blanco suave de algunos huevos domésticos de gallina (Gallus gallus) hasta la cápsula granulada como aguacate del Emu (Dromaius novaehollandiae), los huevos de aves presentan una deslumbrante variedad de diversidad. El estudio de la diversidad de huevos es una rama especializada de la ornitología denominada ovología.

Figura 9.2. Gallus gallus, comúnmente conocido como gallo de corral o gallina doméstica, es una especie de ave de la familia Phasianidae. Originario del sureste de Asia, es ampliamente criado en todo el mundo por su carne y huevos. Los gallos son reconocidos por sus coloridos plumajes y prominentes crestas en la cabeza. Las gallinas, por otro lado, tienen un plumaje más discreto y son conocidas por incubar y cuidar a sus crías. Además de su importancia económica y alimentaria, el Gallus gallus ha sido estudiado como un modelo de investigación en genética y comportamiento animal. Su adaptabilidad y relación simbiótica con los humanos lo convierten en uno de los animales más domesticados y familiarizados en la historia de la humanidad.
Anatomía del huevo aviar
Los huevos de peces y la mayoría de los anfibios solo pueden sobrevivir en el agua, atando a estos animales a una existencia que depende de los ambientes acuáticos. Los huevos de aves, por el contrario, pueden retener su contenido acuoso y desarrollarse en tierra firme en una gran variedad de condiciones ambientales. Los archosaurios amniotas ancestrales desarrollaron por primera vez un huevo de cáscara dura con membranas internas. Debido a que sus huevos contenían el medio acuoso requerido por los embriones para desarrollarse, y bolsas especializadas para retener los desechos y proteger al embrión de los movimientos bruscos del ambiente seco, estos animales ya no necesitaban vivir en o cerca del agua; así comenzó la larga evolución de una gran rama de animales terrestres, incluidas las aves.

Figura 9.3. Anatomía del huevo 2. La estructura de un huevo empollado es diferente, las aves son amniotas y por ende presentan 4 sacos embrionarios, el primero lo tienen todos los organismos con huevo y es el vitelio que contiene la yema donde salen los nutrientes, el segundo es el alantoides donde van a parar los desechos. Observe que el alantoides rodea de manera externa al embrión, esto se debe a que los embriones de ave son muy demandantes en metabolismo, por lo que este alantoides no puede con tener todos los desechos, así que los evapora a través de poros en la cáscara. El amnios es la membrana que rodea al embrión para protegerlo, mientras que el corion que no se puesta rodea al alantoides.
Los huevos de aves son grandes. Los únicos mamíferos con huevos similares a las aves son los monotremas, las cuatro especies de equidnas (osos hormigueros espinosos) y el ornitorrinco (Ornithorhynchus anatinus), que ponen huevos suaves y correosos en lugar de dar a luz a crías vivas. Los embriones contenidos dentro de los grandes huevos externos de aves, reptiles y monotremas deben ser totalmente independientes de los recursos durante el desarrollo temprano, a diferencia de todos los otros mamíferos, cuyas madres internamente proporcionan sustento al embrión. Por lo tanto, un huevo de ave debe estar lleno de todas las proteínas, carbohidratos, grasas y agua que el embrión necesitará hasta que eclosione.

Figura 9.4. Diagrama de un huevo de gallina en su 9° día. Membranas: alantoides, corion, amnios, vitelo / yema. Aunque se muestran separadas, es normal que el corion y el alantoides se combinen formando la membrana corioalantoica, como si fueran una sola.
Una forma de entender la estructura de un huevo es seguirlo a través del proceso de desarrollo. Inicialmente, el óvulo es expulsado de las células foliculares que lo rodean en el ovario, y generalmente es fecundado por un espermatozoide cuando comienza su viaje a través del infundíbulo hasta el oviducto. A medida que el huevo se mueve hacia abajo a través del oviducto, pasa a través de una sucesión de células glandulares que segregan capas sucesivas de albúmina y membranas de la cáscara; La cáscara se agrega al final, muy cerca del término del oviducto. Debido a que ninguna de estas capas agregadas implica la adición de nuevas células con sus propios núcleos, básicamente estamos tratando con una célula que se va agrandado capa por capa, así, el huevo de un avestruz (Struthio camelus), con su único núcleo protegido por capas de material no celular, es una de las células vivas más grandes que podemos observar hoy.
Para familiarizarse con las características principales de un huevo, imagine o explore un huevo de gallina desde su cocina (Figura 9.1). A medida que rompe un huevo de gallina, observe cómo una membrana delgada mantiene unida la frágil capa de mineral, adherida a su superficie interna, algo similar a las capas de vidrio de seguridad para automóviles. Esta membrana se adhiere fuertemente a la concha. Otra membrana rodea la albúmina, o clara de huevo; esta capa puede ser frustrante para pelar un huevo duro.
Una vez que haya atravesado estas dos membranas externas, podrá derramar el contenido del huevo en un tazón; Puedes ver la yema rodeada por la albumina o clara, ahora liberada de su membrana. Aunque es difícil de ver, la yema misma está rodeada y sujeta por la membrana vitelina; Esta membrana es lo que rompes si "rompes" una yema. Puedes notar que la yema gira de manera que una mancha blanca y circular en su superficie está hacia arriba en el plato. Este tejido, denominado blastodermo, se desarrollaría en el embrión si el óvulo fuera fertilizado. Si alguna vez ha tratado de separar una yema del blanco de un huevo roto, se ha encontrado con las partes gelatinosas y fibrosas de la albúmina que a menudo son retorcidas y de color blanco lechoso. Difícil de separar de la yema, estos tejidos son las chalazas. Antes de que el huevo se abriera, las chalazas se conectaban a la cara interna de la Cáscara en los extremos distantes del huevo, suspendiendo así la yema en el centro de la albúmina, protegiéndola y manteniéndola en su lugar dentro del huevo. Ahora consideraremos la estructura y función de cada componente con mayor detalle, comenzando con la yema.
La yema
La yema contiene todos los lípidos (grasas) y la mayor parte de la proteína necesaria para nutrir el embrión en desarrollo hasta que eclosione. Los tamaños relativos de la yema y la clara varían entre los grupos de aves: las yemas tienden a constituir una mayor proporción de la masa total de huevos en especies con pollos precoces, que están muy bien desarrollados en la eclosión (como aves acuáticas, aves costeras y faisanes), en comparación con las especies con crías altriciales (como los paseriformes) en las que las yemas son más pequeñas. Esta diferencia se debe al hecho de que los pollitos precoces generalmente se desarrollan en el huevo durante un período de tiempo más largo y, por lo tanto, requieren más sustento y, por ende, una yema más grande.
La yema es la primera parte del huevo producido por la hembra ponedora. Antes de que la yema pase por el oviducto para la adición de albúmina y cáscara, experimenta un rápido crecimiento durante un período de aproximadamente 5 días a 2 semanas. Sin embargo, muchas aves, especialmente las aves cantoras, generalmente ponen un huevo por día, un huevo único por puesta. Debido a que la preparación de la yema requiere más de un día, las hembras a menudo preparan las yemas en “filas” para una nidada completa antes de poner su primer huevo. Durante este período de preparación, que a veces se denomina etapa de "enyemado", la yema se deposita dentro de la membrana vitelina en bandas alternas de yema más oscura y más clara, produciendo una estructura que se asemeja a los anillos de crecimiento en un árbol cuando se observa en condiciones especiales de laboratorio.

Figura 9.5. Composición de la cáscara de huevo. Esta micrografía de una cáscara de huevo, junto con un esquema ampliado, muestra la ubicación de los poros y la membrana interna de la cáscara.
Debido a que estas bandas se alternan en un ciclo diario (las bandas oscuras se depositan durante el día, cuando la hembra ingiere alimentos ricos en pigmentos), la estructura del anillo de la yema revela cuánto tiempo necesita cada hembra para preparar sus yemas para la puesta. La única estructura que interrumpe estos anillos concéntricos es un cilindro de yema de color claro que se extiende desde el núcleo de la yema hasta su superficie, donde se encuentra el punto germinal. El embrión se desarrollará aquí, comenzando como el blastodermo similar a un disco aplanado. Dado que la mancha germinal y su columna son más ligeras que el resto de la yema, el embrión en desarrollo siempre flota hacia la parte superior, sin importar de qué manera se gire el huevo. La yema flotaría contra la cáscara si la chalaza no la suspendiera en el centro del huevo.
La clara
La albúmina, que varía en viscosidad según la proporción de agua y proteína que contiene, consiste en una serie de capas alrededor de la yema. Una capa muy delgada de albúmina viscosa rodea la yema y se extiende hacia la chalaza, seguida por una capa delgada de albúmina acuosa, una capa gruesa de albúmina más viscosa y fibrosa variable (el componente más grande), y finalmente una capa delgada de albúmina acuosa derecha debajo de la cáscara. Además de proporcionar casi toda el agua y gran parte de la proteína para el embrión en desarrollo, estas capas de albúmina sirven admirablemente para proteger al embrión del daño físico, siempre que la cáscara no esté rota.
La concha
La concha es la línea de defensa externa del embrión en desarrollo (Figura 9.5). Las cáscaras de huevos aviares son mucho más gruesas y fuertes que las cáscaras de otros vertebrados que ponen huevos. Sin embargo, este mayor grosor dificulta el intercambio de gas a través de la carcasa. El crecimiento y la respiración de un embrión en desarrollo requieren oxígeno y producen dióxido de carbono.
El huevo no puede contener todo el oxígeno necesario para alimentar el metabolismo del embrión, ni puede contener todo el dióxido de carbón y los desechos solubles residuales producidas por el embrión antes de eclosionar. Por lo tanto, el embrión en desarrollo debe poder "respirar" a través de los poros en su cáscara. Un huevo pierde, en promedio, el 18% de su masa entre la puesta y la eclosión, principalmente por la pérdida de agua a través de la cáscara. La porosidad de la cáscara también puede poner en riesgo al embrión.
Contaminantes como el aceite pueden entrar al huevo y envenenar el embrión o recubrir la superficie del huevo, cortando el intercambio de gases. Más del 90% de los embriones de ánades reales (Anas platyrhynchos) murieron cuando sus cáscaras de huevo se expusieron a solo una décima parte de una gota de petróleo crudo (Hoffman & Moore, 1979).

Figura 9.6. El ánade real (Anas platyrhynchos) es una especie de pato silvestre ampliamente distribuido en Eurasia y América del Norte. Conocido también como pato común o pato azulón, es reconocido por su característico plumaje con cabeza verde iridiscente, pecho blanco y cuerpo marrón. Los machos tienen un llamativo pico amarillo y exhiben un comportamiento de cortejo elaborado durante la temporada de reproducción. Son aves acuáticas versátiles que se adaptan a diversos hábitats, desde lagos y ríos hasta estanques y parques urbanos. El ánade real es apreciado tanto por su belleza como por su importancia ecológica en la dispersión de semillas y control de insectos acuáticos.
El huevo fertilizado
Si tuviera que abrir un huevo de pollo fértil en una etapa avanzada de desarrollo embrionario, toda la superficie interna de la cáscara se cubriría con una membrana llena de vasos sanguíneos que facilitan el intercambio de gases.

Figura 9.7. La anatomía de la membrana cambia a medida que se desarrolla el embrión. Con una infusión de capilares sanguíneos, la membrana corioalantoica está conectada a los órganos internos de este embrión de pollo (Gallus gallus). El espacio de aire en la parte superior del huevo se expande gradualmente a medida que el contenido de agua disminuye a través del consumo del embrión y la evaporación a través de la cáscara permeable.
Esta membrana corioalantoica (Figura 9.7) está formada por la fusión de dos sacos embrionarios, el corion y la alantois, que están conectados a los órganos internos del corazón del embrión en desarrollo, muy similar a la conexión de un embrión humano con la placenta de su madre. a través del cordón umbilical. El corion es la membrana externa que rodea todo el embrión aviar; Es evolutivamente homólogo a la membrana del mismo nombre en los mamíferos, que forma gran parte de la placenta de los mamíferos. La alantois contiene cualquier desecho metabólico que no pueda evaporarse a través de la cáscara, como los cristales de ácido úrico. Como una fusión de estas membranas, la membrana corioalantoica sirve como el equivalente funcional de los pulmones del embrión a lo largo de su desarrollo dentro del huevo (Figura 9.7).
La membrana corioalantoica cubre cada vez más la superficie interna de la concha a medida que se desarrolla el embrión. En un huevo recién puesto, la membrana que encierra la albúmina se adhiere directamente a la membrana justo debajo de la cáscara, excepto en el extremo romo del huevo, donde se produce un pequeño espacio de aire. Este es el mismo espacio de aire que puede encontrar al pelar un huevo duro fresco. A medida que el agua dentro del huevo se evapora a través de la cáscara o es consumida por el embrión en desarrollo, este espacio de aire entre las dos membranas se expande gradualmente (Figura 9.7). La flotabilidad impartida por este espacio aéreo en crecimiento es valiosa para los ornitólogos de campo que necesitan verificar el progreso del desarrollo de un huevo: la medida en que el huevo flota o se hunde cuando se coloca en el agua a menudo indica cuánto más debe desarrollarse el embrión antes de la eclosión.
Colores
Diferentes animales ponen huevos coloreados de distintas maneras. Los pigmentos protoporfirina, biliverdina y quelato de cinc de biliverdina son los responsables de la diversidad de colores en los huevos. Estos pigmentos son segregados por las células de las paredes del oviducto y pueden producir manchas en el cascarón si el color es añadido inmediatamente antes de poner el huevo. Los huevos de los reptiles modernos son todos blancos, y se cree que así era para todos los animales prehistóricos.
Es posible que las manchas de los huevos de muchas aves sirvan de camuflaje. Las aves que anidan en cavidades (tales como los pájaros carpinteros y el martín pescador) no necesitan huevos camuflados. Sus huevos son normalmente de un blanco brillante, de modo que los padres puedan localizarlos fácilmente. En las especies que anidan en grandes grupos, como el arao común (Uria aalge), los huevos de cada hembra presentan marcas muy diferentes entre sí, facilitando que cada hembra reconozca su propio huevo.

Figura 9.8. El arao común (Uria aalge) es una especie de ave marina perteneciente a la familia Alcidae. También conocido como arao aliblanco, se encuentra en las regiones costeras del Atlántico Norte y el Ártico. Estos pájaros tienen un plumaje principalmente negro con un vientre blanco, y se distinguen por su pico largo y afilado. Son excelentes nadadores y buceadores, alimentándose principalmente de peces y calamares. Los araos comunes forman colonias en acantilados y acuden a tierra solo para reproducirse. Son conocidos por sus llamativas exhibiciones de cortejo y su habilidad para volar a gran velocidad. Estas aves desempeñan un papel importante en los ecosistemas marinos y son un símbolo icónico de las regiones costeras del norte.
Textura del cascarón
Normalmente los huevos son lisos, pero hay excepciones. El huevo de los cormoranes, por ejemplo, es muy áspero y parecido a la tiza. En contraste, los huevos de los tinamúes son muy brillantes, y los de los patos son grasos e impermeables, y de color amarillo. Otra variación es el huevo muy picado de los casuarios. El cascarón posee poros minúsculos para permitir al pichón respirar. El huevo de la gallina doméstica tiene unos 7500 poros.
Forma
La mayoría de huevos de aves tienen una forma oval característica, con un extremo redondeado y el otro más aguzado. Esta forma ocurre ya que el huevo es forzado a través del oviducto (previamente al paso por el oviducto, y aún con la cáscara blanda la forma suele ser esférica): los músculos contraen el oviducto por detrás del huevo, empujándolo. Como en ese momento la cáscara del huevo todavía es deformable, la parte posterior del huevo adopta su forma puntiaguda.
La forma oval tiene grandes ventajas: ocupa más eficientemente el espacio dentro de un nido (y en un recipiente) que una forma esférica; evita que los vuelcos sean en posiciones muy aleatorias; generalmente el vuelco es lateral y es así suavizada su fuerza por el deslizamiento que posibilita la forma; la misma forma oval también aporta más resistencia de la cáscara a las presiones en los "polos" (por ejemplo, la presión del peso del ave que está empollando).
Depredación
Hay muchos animales que se alimentan de los huevos. Depredadores principales del ostrero negro americano, por ejemplo, incluyen a los mapaches, las mofetas, las nutrias (de río y marinas), varias especies de gaviota, el cuervo y los zorros. El armiño (Mustela erminea) y la comadreja (Mustela frenata) les roban los huevos a los patos. Las serpientes devoradoras de huevos (géneros Dasypeltis y Elachistodon) son especialistas en comer los huevos.
Puesta
Una vez que todas las capas de membrana se han depositado y la cáscara y la pigmentación están completas, el huevo pasa a través de la cloaca. Aunque no se conocen los tiempos de puesta precisos para la mayoría de las especies, un patrón parece mantenerse: cuanto más grande es el huevo, más largo es el proceso de puesta. Los parásitos de la cría han evolucionado para poner sus huevos notablemente rápido, antes de que el huésped vuelva a su nido. Los cucos comunes (Cuculus canorus), por ejemplo, pueden poner un huevo en el nido de otra ave y desaparecer en menos de 10 segundos.

Figura 9.9. El cuco común (Cuculus canorus) es un ave migratoria de la familia Cuculidae. Es conocido por su comportamiento parasitario, ya que pone sus huevos en los nidos de otras especies de aves, dejando que sean estas últimas quienes críen a sus polluelos. El cuco común es de tamaño mediano, con un plumaje gris y blanco, y una llamativa cresta en forma de flecha en la cabeza. Su canto característico, un sonido repetitivo y distintivo de "cu-co", es ampliamente reconocido. Estas aves se encuentran en una amplia variedad de hábitats, desde bosques hasta áreas urbanas. Su estrategia de cría única y su distintivo canto los hacen fascinantes para los observadores de aves.
El tiempo requerido para que el oviducto segregue las capas alrededor del óvulo determina el intervalo entre los huevos puestos. Las aves playeras pequeñas, los pollos domésticos, los pájaros carpinteros, y la mayoría de los paseriformes ponen huevos con aproximadamente 24 horas de diferencia. Por el contrario, las aves como avestruces, rheas, garzas, cigüeñas, grúas, avutardas, gaviotas, palomas, búhos, colibríes, vencejos, martines pescadores, así como algunos halcones y cucos generalmente ponen huevos cada dos días. Las especies más grandes tienden a necesitar más tiempo entre los huevos, tal vez porque los procesos de secreción en el oviducto toman más tiempo para los huevos más grandes. Entre las aves acuáticas, por ejemplo, muchas especies de patos ponen huevos todos los días, mientras que los grandes gansos y cisnes ponen cada dos días.

Figura 9.10. La puesta es un momento vulnerable para las aves, ya que deben depositar sus huevos en nidos seguros para asegurar la supervivencia de sus crías. Durante este proceso, las aves buscan lugares protegidos y bien camuflados para evitar la detección de depredadores. Construyen nidos con materiales disponibles, como ramitas, hojas y barro, y algunos incluso utilizan tejidos de arañas para reforzar la estructura. Una vez que los huevos son depositados, la hembra incuba los huevos mientras el macho se encarga de proveer alimento y protección. Durante esta etapa, las aves son especialmente sensibles a perturbaciones y alteraciones en su entorno, ya que cualquier interrupción puede resultar en el abandono de los huevos. Es importante respetar y preservar los hábitats naturales de las aves durante el período de puesta, evitando la destrucción de nidos y la interferencia humana en sus áreas de reproducción. De esta manera, contribuimos a garantizar la supervivencia de estas maravillosas criaturas y a mantener el equilibrio de los ecosistemas en los que habitan. (YouTube)
Anidación
Solo los insectos superan a las aves en la diversidad y sofisticación de sus nidos. Una gran cantidad de literatura, en su mayoría del siglo XIX, describe el comportamiento de las aves en la construcción de nidos. Sin embargo, en todo el espectro de especies de aves, desde aves de traspatio comunes hasta especies exóticas en tierras lejanas, todavía no se han estudiado los aspectos de la biología de anidación de la mayoría de las especies. En términos de construcción y colocación, cada tipo de nido de ave incurre en sus propios costos (como el esfuerzo de los padres y el riesgo de depredación o sabotaje) y los beneficios (que incluyen una mayor seguridad del nido y la eficiencia de la termorregulación). Para explorar estos diversos costos y beneficios, es necesario apreciar las funciones que sirven los nidos y la diversidad de formas en que se logran estos objetivos. Dada la extensión de este capítulo, nos enfocaremos en las funciones de los nidos.
La mayoría de las aves construyen nidos principalmente para mantener y proteger sus huevos, y para mantenerlos juntos de modo que un ave progenitora pueda incubarlos a la temperatura adecuada para su desarrollo. Un simple rasguño en el suelo puede servir como un nido adecuado para algunas aves. Unos cuantos pájaros que ponen solo un huevo a la vez han abandonado por completo los nidos e incuban su único huevo de otras maneras. Sin embargo, la mayoría de las aves construyen algún tipo de nido. Los nidos a veces cumplen funciones adicionales: algunas aves se posan en sus nidos incluso más allá de la temporada de reproducción, mientras que otros construyen nidos como un tipo de exhibición para atraer parejas. Para algunas especies, la construcción mutua de nidos es un componente común de la formación de pares y la unión de pareja. Sin embargo, casi todos los nidos deben proporcionar principalmente un refugio seguro contra los depredadores y los elementos.
Un nido exitoso generalmente frustra a los depredadores a través de su ocultamiento y / o inaccesibilidad. Los nidos de la mayoría de las especies de aves son, por lo tanto, difíciles de encontrar. Por ejemplo, el riesgo de depredación de nidos es particularmente alto en muchos hábitats de bosques tropicales, y muchos paseriformes tropicales, en consecuencia, construyen nidos especialmente pequeños que no son visibles fácilmente. La mayoría de las aves construyen nidos con materiales comunes, generalmente evitando sustancias de colores brillantes. Para mejorar aún más el camuflaje, algunas especies fijan líquenes o trozos de corteza en el exterior de un nido. Los nidos de algunos paseriformes incluyen un colgante largo y arrastrado, tal vez para romper el perfil visual de la forma del nido.
La mayoría de las especies de aves también son muy adeptas a la construcción de nidos fuera de alcance. Por ejemplo, las aves en grupos tan diversos como los vireos, las espinosas y las garzas a menudo prefieren anidar precariamente en las puntas de las ramas largas y altas, lugares a los que la mayoría de los depredadores que trepan, como roedores o serpientes, no pueden llegar. Otras aves comúnmente construyen nidos en cavidades de árboles, dentro de rincones de acantilados, o en lo alto de las paredes de los edificios.
Entre las ubicaciones de nidos más interesantes se encuentran las construidas cerca de animales que disuaden a los depredadores. Muchas aves tropicales ponen sus huevos dentro o muy cerca de los nidos de avispas agresivas u hormigas, las cuales atacarán a un depredador que se aproxima “comensalismo”. En los neotrópicos, algunas especies de trogones, martines pescadores y loros cavan túneles de nidos dentro de termitas activas o montículos de hormigas. Cómo (o por qué) estas aves que anidan se salvan de los ataques de los insectos sigue siendo un misterio.
Los nidos también proporcionan refugio de los elementos. El microclima alrededor y dentro de muchos nidos es mucho más favorable para los huevos y los pichones que el ambiente circundante. Por ejemplo, los colibríes de anidación templada a menudo eligen los sitios de anidación bajo los árboles colgantes para minimizar la pérdida de calor en la noche. Algunas gaviotas colocan sus nidos en la sombra para reducir el riesgo de que sus polluelos se sobrecalienten o se deshidraten. De manera similar, en el desierto, donde la vegetación es escasa y la radiación solar es intensa, muchas aves construyen nidos cerrados para proporcionar sombra a sus huevos y a ellos mismos. Los nidos también pueden evitar que los huevos y las crías se maltraten. Los orioles y oropéndolas del Nuevo Mundo, los tejedores del Viejo Mundo y las esposas anchas asiáticas construyen nidos profundos y cerrados en las puntas de las ramas largas y delgadas. Esta combinación de estructura de nido y sitio funciona en tándem: la estructura evita que los huevos se caigan cuando el nido se balancea violentamente con el viento, y la ubicación impide el acceso de depredadores como monos.
10. Nidada e incubación
10. Nidada e incubación
La nidada e incubación en aves es un fascinante proceso de reproducción que desempeña un papel crucial en la supervivencia de las especies aviares. Durante este período, las aves construyen nidos cuidadosamente elaborados y protegen sus huevos mientras los incuban hasta que eclosionan. Este artículo explora en detalle los diferentes aspectos de la nidada e incubación en aves, desde la elección del sitio de anidación y la construcción del nido, hasta el comportamiento de incubación y las estrategias de protección empleadas por las aves durante esta etapa crítica. Además, examinaremos cómo los factores ambientales, la interacción entre los padres y la influencia humana pueden afectar el éxito reproductivo de las aves. A través de un conocimiento más profundo de este proceso, podemos apreciar la belleza y complejidad de la crianza aviar, así como la importancia de conservar y proteger los hábitats naturales donde ocurre.
La nidada
Una nidada es el número total de huevos depositados en una serie ininterrumpida, durante un solo período de anidación, por un ave hembra. En los primeros días de la ornitología, los recolectores de huevos de todo el mundo acumularon grandes colecciones de huevos de aves. Por lo tanto, el tamaño de la nidada es a menudo el único rasgo central de la historia de la vida aviar para el que existe información cuantitativa; como resultado, ha habido un interés considerable durante casi un siglo en tratar de comprender los patrones de variación en el tamaño de la nidada. Durante la década de 1940, David Lack en la Universidad de Oxford (Reino Unido) comenzó a estudiar los tamaños de nidada y las historias de vida de varias especies de aves locales. Muchos de estos estudios continúan hasta el presente, aportando contribuciones a la teoría de la historia de la vida aviar.
Las aves estrechamente relacionadas tienden a tener nidadas de tamaño similar. Por ejemplo, todas las aves marinas tubenosadas (como albatros y petreles) ponen un solo huevo, prácticamente todos los colibríes ponen dos huevos, las aves playeras no ponen más de cuatro, y así sucesivamente. La mayoría de los pájaros cantores ponen entre dos y seis huevos, aunque algunos herrerillos comunes tienen nidadas extraordinariamente grandes de hasta 17 huevos.
La disponibilidad de alimentos puede ser el factor más importante que influye en el tamaño de la nidada. La disponibilidad de alimentos para la hembra ponedora es a menudo una limitación importante; ya que poner un huevo es energéticamente costoso para la hembra, incluso en lugares con abundantes recursos, hacerlo sin un suministro adecuado de alimentos amenaza la supervivencia de la hembra o sus esfuerzos reproductivos posteriores. Por lo tanto, aunque las hembras todavía son fisiológicamente capaces de poner más huevos, a menudo limitan su producción de huevos (y por lo tanto el tamaño de la nidada) cuando la comida es escasa.
Incubación
Todas las aves ponen huevos externos que deben mantenerse en el rango de temperatura adecuado a lo largo de su desarrollo; Esta temperatura se mantiene mediante un proceso llamado incubación.

Figura 10.1. Los chorlitos egipcios (Pluvianus aegyptius) son aves del desierto que habitan en el norte de África y el Medio Oriente. Estas aves pequeñas se distinguen por su plumaje blanco y negro, pico y patas rojas. Su adaptación al entorno desértico les permite encontrar alimento, principalmente insectos y pequeños invertebrados, en la arena caliente. Una característica interesante es su comportamiento de "lavar" las plumas con aceite para repeler el calor y el polvo del desierto. Son aves cooperativas en la crianza, formando parejas monógamas que construyen nidos juntos y comparten las responsabilidades de incubar los huevos y cuidar a los polluelos. Aunque no están en peligro de extinción, su hábitat se enfrenta a desafíos debido a la degradación del desierto.
La temperatura precisa necesaria para el desarrollo embrionario varía entre las especies de aves, pero la mayoría de los embriones aviares requieren temperaturas cercanas a los 37 °C o 38 °C. Debido a que pocos ambientes tienen esta temperatura constante, casi todas las especies de aves deben proporcionar una fuente de calor para sus huevos. La mayoría de las especies usan su propio calor corporal al sentarse sobre los huevos, pero dos grupos emplean otros métodos: los parásitos de cría dejan la incubación de sus huevos a sus huéspedes, y los megapodos proporcionan fuentes alternativas de calor a través de una amplia variedad de métodos, como hacer una pila de material en descomposición, la composta genera calor que incuba a los huevos.

Figura 10.2. El frailecillo de Wilson (Charadrius wilsonia) es un encantador ave playera que habita en la costa oeste de América del Norte. Con su plumaje gris plateado, cabeza negra y pico anaranjado, es fácil de reconocer. Estas aves migratorias se alimentan de pequeños invertebrados marinos, que capturan con destreza utilizando su pico puntiagudo. Durante la temporada de reproducción, forman colonias en playas y dunas costeras, construyendo nidos poco profundos en la arena. Aunque no están en peligro de extinción en la actualidad, enfrentan desafíos como la pérdida de hábitat y la contaminación de los océanos. La protección de sus áreas de reproducción y la concienciación sobre su importancia son fundamentales para preservar a estas hermosas aves a largo plazo.
En casos raros, las aves deben enfriar sus huevos para mantenerlos en el rango de temperatura apropiado. Debido a sus hábitats de reproducción extremadamente calientes a lo largo del Golfo de México, los frailecillos de Wilson (Charadrius wilsonia) sumergen las plumas de sus pechos en agua y corren para enfriar sus huevos con la humedad.

Figura 10.3. El zanco de cuello negro (Himantopus mexicanus) es una elegante ave zancuda que se encuentra en diversas regiones de América, desde Estados Unidos hasta América del Sur. Su característica distintiva es su largo y delgado cuello negro, que contrasta con su plumaje blanco. Estas aves son expertas en vadear aguas poco profundas en busca de alimento, principalmente insectos acuáticos y pequeños crustáceos. Su pico recto y puntiagudo les permite capturar presas con precisión. Son aves gregarias y suelen formar colonias durante la temporada de reproducción, construyendo nidos en áreas pantanosas o costeras. Aunque no están en peligro de extinción, la pérdida de humedales y la contaminación del agua representan desafíos para su conservación.
A lo largo de los bancos de arena de los ríos africanos, los chorlitos egipcios (Pluvianus aegyptius) van un paso más allá, además de enfriar los huevos con plumas de pechuga mojadas, los adultos también pueden enterrar sus huevos (y luego a veces sus polluelos) en arena para protegerlos del sol. Un par de zancos de cuello negro (Himantopus mexicanus) que anidaban en el cálido y árido sur de California (EE. UU.) empaparon las plumas de su vientre y llevaron la humedad refrescante al nido 953 veces en un día. Las aves marinas con nidos abiertos en islas tropicales o desiertos costeros a menudo pasan más tiempo parados sobre sus huevos que cazando.
Rompimiento
Las aves recién nacidas varían ampliamente en su disposición para la vida fuera del huevo. Los pollitos altriciales se incuban desnudos o con poca pluma, sin capacidad para generar suficiente calor para la termorregulación y totalmente dependientes de sus padres para la alimentación. En contraste, los polluelos precoces ya están bien emplumados cuando eclosionan, con poderes sustanciales de termorregulación y locomoción, y un grado considerable de independencia de sus padres para alimentarse. Estas dos categorías —altriciales versus precoz— son puntos finales en un continuo de tipos de desarrollo, y muchas especies tienen pollitos que son mezclas intermedias de estos modos de desarrollo.
Rompimiento (YouTube).
Reconocimiento y cuidado parental
Incluso antes de que eclosionen las aves jóvenes, a menudo tienen contacto social con los padres (o solo uno de los parentales) que asisten al nido. Los jóvenes de algunas especies se asoman desde el interior de su huevo, lo suficientemente alto como para ser audibles desde el exterior. Aparentemente, pueden escuchar a sus padres a cambio, ya que muchos dejarán de mirar furtivamente después de escuchar una respuesta vocal del adulto. Algunas especies, como la codorniz norteña (Colinus virginianus), incluso aprenden a reconocer la llamada de su propia madre cuando aún están en el huevo.
Reconocimiento del padre a la cría
No obstante, solo algunos tipos de aves progenitoras son capaces de reconocer a sus crías. Los investigadores pueden intercambiar a los jóvenes de la mayoría de las especies de paseriformes sin que los padres se den cuenta de que ha ocurrido algún cambio: los adultos generalmente parecen estar dispuestos a alimentar a cualquier polluelo que está mendigando en el nido.

Figura 10.4. La codorniz norteña (Colinus virginianus) es una pequeña ave nativa de América del Norte, conocida por su llamativo plumaje y su distintivo canto. Estas aves terrestres habitan una variedad de hábitats, desde praderas hasta matorrales y pastizales. Su cuerpo compacto y alas cortas les permiten correr rápidamente y evadir a los depredadores. Son aves sociales que forman grupos durante la temporada de reproducción y crían en nidos en el suelo. La codorniz norteña se alimenta principalmente de semillas y vegetación, pero también consume insectos en menor medida. Aunque su población es estable, la pérdida de hábitat y la caza excesiva son amenazas potenciales para su conservación.
Los adultos de muchas especies no paseriformes, especialmente las aves que anidan en colonias, sin embargo, no se engañan tan fácilmente. Por ejemplo, durante 2 o 3 días después de su eclosión joven, la mayoría de las gaviotas aceptarán a pequeñas crías de otros nidos como propias, ya sea que las crías paseen naturalmente o lleguen como parte del experimento de un biólogo. Después de este breve período durante el cual los adultos se acostumbran a los polluelos en su propio territorio, rechazan a cualquier otro; de hecho, los adultos perseguirán y picotearán a los jóvenes callejeros hasta que abandonen el territorio o mueran. Este cambio de desarrollo en el reconocimiento probablemente refleja cambios en la probabilidad de que los hijos se confundan: los polluelos de gaviota muy jóvenes no son tan propensos a vagar en territorios extranjeros como los polluelos de mayor edad, y los pollos jóvenes son, por lo tanto, mucho más propensos a ser la descendencia legítima del adulto.
Reconocimiento de la cría a los padres
La mayoría de las aves jóvenes no reconocen a sus padres por algún tiempo después de la eclosión. Los polluelos altriciales piden a casi cualquier ave que se acerque al nido, incluso a un depredador como un halcón, a menos que estén asustados por el tamaño o el comportamiento del recién llegado o que el padre haga una llamada de advertencia. Sin embargo, a medida que crecen, los polluelos aprenden a reconocer a sus padres y ahorran su energía de mendicidad para los momentos en que es más probable que conduzcan a la entrega de alimentos. En contraste, los jóvenes de muchas aves que anidan en colonias reconocen las llamadas de sus propios padres a una edad temprana.
Este reconocimiento les permite prepararse rápidamente para recibir alimentos cuando llega el padre y, en algunas especies, les impide mendigar a los adultos equivocados, que pueden reaccionar agresivamente. Los pollos reconocen las llamadas de sus padres a los 6 días de edad en gaviotas risueñas (Leucophaeus atricilla) de la costa atlántica de América del Norte, a los 3 días en murres de pico grueso (Uria lomvia) que se crían en acantilados sobre el Océano Ártico, y por al menos 14 días (7 días antes de la fuga) en las urraca piñonera (Gymnorhinus cyanocephalus) del suroeste de Estados Unidos.

Figura 10.5. Los murres de pico grueso (Uria lomvia) son aves marinas que se encuentran en las regiones árticas y subárticas del hemisferio norte. Estas aves son reconocidas por su apariencia distintiva, con un plumaje negro en la parte superior y blanco en la parte inferior, y un pico largo y robusto. Son expertas buceadoras, pudiendo sumergirse a grandes profundidades en busca de peces y otros organismos marinos. Durante la temporada de reproducción, forman grandes colonias en acantilados y acuden en masa para anidar. Aunque su población global es estable, el cambio climático y la contaminación marina representan amenazas potenciales para su hábitat y alimentación. La conservación de los ecosistemas marinos es esencial para garantizar la supervivencia de estas fascinantes aves.
En prácticamente todas las aves, las vocalizaciones de mendicidad se vuelven más intensas cuando los polluelos tienen más hambre, pero en algunos paseriformes que anidan en colonias, como los tejedores espalda dorada (Ploceus jacksoni) de África, las vocalizaciones de los descendientes también contienen información sobre la identidad de los polluelos. Para la mayoría de las especies cuyos jóvenes se juntan en comunidades, los jóvenes reconocen las llamadas de sus padres cuando se forman estos grupos; Esto es cierto para las cacatúas de Galah australianas (Eolophus roseicapilla), los pingüinos de Adelie (Pygoscelis adeliae) y los pelícanos blancos americanos (Pelecanus erythrorhynchos).

Figura 10.6. El pelícano blanco americano (Pelecanus erythrorhynchos) es una majestuosa ave acuática que se encuentra en América del Norte. Su característica más destacada es su enorme pico, que puede alcanzar más de 40 centímetros de longitud. Estos pelícanos tienen un plumaje blanco, excepto por sus alas negras durante la época de reproducción. Son excelentes pescadores, utilizando su bolsa de piel para atrapar peces y otros animales acuáticos. Forman colonias en islas y costas donde construyen nidos con ramas y vegetación. Aunque su población es estable, la contaminación del agua, la pérdida de hábitat y la pesca comercial representan desafíos para su conservación. Proteger sus áreas de anidación y promover prácticas pesqueras sostenibles es crucial para preservar a este magnífico ave.
Todos los polluelos altriciales y la mayoría de los polluelos precoces requieren algún tipo de cuidado parental. Desde el momento en que nacen hasta que alcanzan la independencia, los jóvenes altriciales dependen en gran medida de sus padres. Los padres de jóvenes altriciales encuentran y alimentan a sus polluelos grandes cantidades de comida, a menudo mantienen el nido limpio al desechar sus heces y las defienden contra los depredadores. Incluso la mayoría de los polluelos precoces, que normalmente se alimentan a sí mismos, dependen de sus padres para vigilar a los depredadores y protegerse de los elementos.
R. Referencias bibliográficas
R. Referencias bibliográficas
Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molecular biology of the cell (5th ed.). Garland Science.
Alcântara, D., Rodrigues, M. N., Franciolli, A. L. R., Da Fonseca, E. T., Silva, F. M. O., Carvalho, R. C., … Miglino, M. A. (2013). Embryonic development of endoderm in chicken (Gallus gallus domesticus). Microscopy Research and Technique, 76(8), 803.
Belk, C., & Maier, V. B. (2013). Biology Science for Life with physiology. (Pearson, Ed.) (4th ed.).
Berta, A., Sumich, J. L., & Kovacs, K. M. (2015). Marine mammals: evolutionary biology. Academic Press.
Brusca, R., Brusca, G. J., & Haver, N. J. (2003). Invertebrates (2nd ed.). Sinauer Associates.
Campbell, M. K., & Farrell, S. O. (2009). Biochemistry (6th ed.). USA: Thomsom Brooks/Cole.
Cox, M. M., Doudna, J. A., & O’Donnell, M. (2012). Molecular biology (1st ed.). Freeman.
Deviche, P., Hurley, L. L., & Fokidis, H. B. (2011). Avian testicular structure, function, and regulation. In Hormones and reproduction of vertebrates (pp. 27–70). Elsevier.
Gilbert, S. F. (2000). Early Development in Birds. In Developmental Biology (6th ed.). Sunderland (MA): Sinauer Associates.
Goodenough, J., & McGuire, B. (2012). Biology of Humans, Concepts, Applications and Issues (4th ed.). San Francisco: Pearson, Benjamin Cummings.
Hoefnagels, M. (2015). Biology: concepts and investigations (3rd ed.). McGraw-Hill New York.
Hoffman, D. J., & Moore, J. M. (1979). Teratogenic effects of external egg applications of methyl mercury in the mallard, Anas platyrhynchos. Teratology, 20(3), 453–461.
Huxley, T. H. (1867). On the classification of birds: and on the taxonomic value of the modifications of certain of the cranial bones observed in that class. The Society.
Kardong, K. V. (2011). Vertebrates, comparative anatomy, function, evolution (6th ed.). McGraw-Hill New York.
Kardong, K. V. (2014). Vertebrates: Comparative Anatomy, Function, Evolution (7th ed.). McGraw-Hill Education.
Karp, G. C. (2013). Cell and Molecular Biology, Concepts and Experiments (7th ed.). USA: Wiley Online Library.
Lovette, I. J., & Fitzpatrick, J. W. (2016). Handbook of bird biology. John Wiley & Sons.
Mackean, D. G., & Hayward, D. (2014). Biology (3rd ed.). IGCSE Cambridge.
Mader, S. S. (2010). Biology (10th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2015). Biology (12th ed.). McGraw-Hill Education.
Mader, S. S., & Windelspecht, M. (2018). Essentials of biology (5th ed.). McGraw-Hill Education.
Marieb, E. N., Wilhelm, P. B., & Mallatt, J. (2017). Human Anatomy (8th ed.). Pearson.
Mason, K. A., Losos, J. B., Singer, S. R., & Raven, P. H. (2014). Biology (7th ed.). McGraw-Hill New York.
Murray, R. K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V., & Weil, A. (2012). Harpers Illustrated Biochemistry (29th ed.). McGraw-Hill Medical.
Otter, K., & Ratcliffe, L. (1996). Female initiated divorce in a monogamous songbird: abandoning mates for males of higher quality. Proceedings of the Royal Society of London. Series B: Biological Sciences, 263(1368), 351–355.
Pollard, T. D., Earnshaw, W. C., Lippincott-Schwartz, J., & Johnson, G. T. (2017). Cell Biology (3rd ed.). Elsevier.
Pomiankowski, A., & Iwasa, Y. (1998). Runaway ornament diversity caused by Fisherian sexual selection. Proceedings of the National Academy of Sciences, 95(9), 5106–5111.
Reece, J. B., Urry, L. A., Wasserman, S. A., Cain, M. L., Minorsky, P. V, & Jackson, R. B. (2014). Campbell Biology (10th ed.). Pearson Higher Ed.
Ritchison, G. (2015). Bird biogeography. RITCHISON, G. Avian Biology: Everything You Wanted to Know about Birds… and More. Eastern Kentucky University,[Sd]. Disponível Em:< Http://People. Eku. Edu/Ritchisong/Birdbiogeography1. Htm>. Acesso Em, 9.
Sadava, D., Berenbaum, M., & Hillis, D. (2014). Life the Science of Biology (10th ed.). Sinauer & MacMillian.
Sadler, T. W. (2012). Medical Embryology. (C. Taylor, Ed.) (12th ed.). Baltimore: Lippincott Williams & Wilkins.
Shier, D., Butler, J., & Lewis, R. (2015). Hole’s Human Anatomy & Physiology (14th ed.). McGraw-Hill Education.
Simon, E. J., Dickey, J. L., & Reece, J. B. (2013). Essential biology (Pearson).
Solomon, E., Martin, C., Martin, D. W., & Berg, L. R. (2014). Biology (10th ed.). Cengage Learning.
Starr, C., Evers, C., & Starr, L. (2013). Biology: Today and Tomorrow With Physiology (4th ed.). Brooks/Cole.
Taylor, M. R., Simon, E. J., Dickey, J. L., Hogan, K., & Reece, J. B. (2018). Biology, Concepts and conections (9th ed.). Boston: Pearson education.
Vitt, L. J., & Caldwell, J. P. (2014). Herpetology An Introductory Biology of Amphibians and Reptiles (4th ed.). Oklahoma: Elsevier Inc.
Weaver, R. F. (2012). Molecular biology (5th ed.). McGraw-Hill Education.
Woolley, S. C., & Doupe, A. J. (2008). Social context–induced song variation affects female behavior and gene expression. PLoS Biology, 6(3), e62.
No hay comentarios:
Publicar un comentario